
Molecular Mechanisms Underlying Fungicide Resistance in Citrus Postharvest Green Mold


Abstract
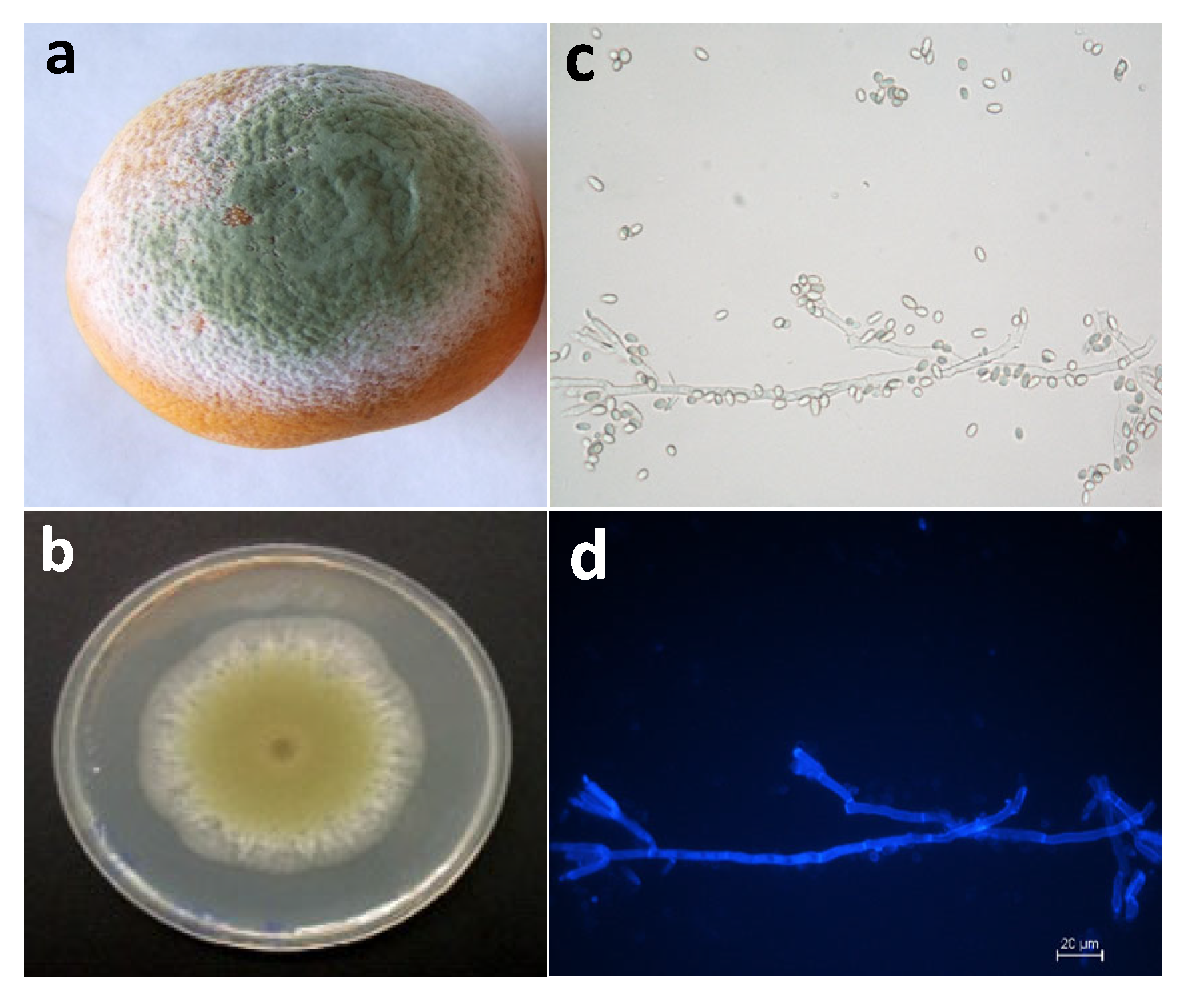
:1. Introduction
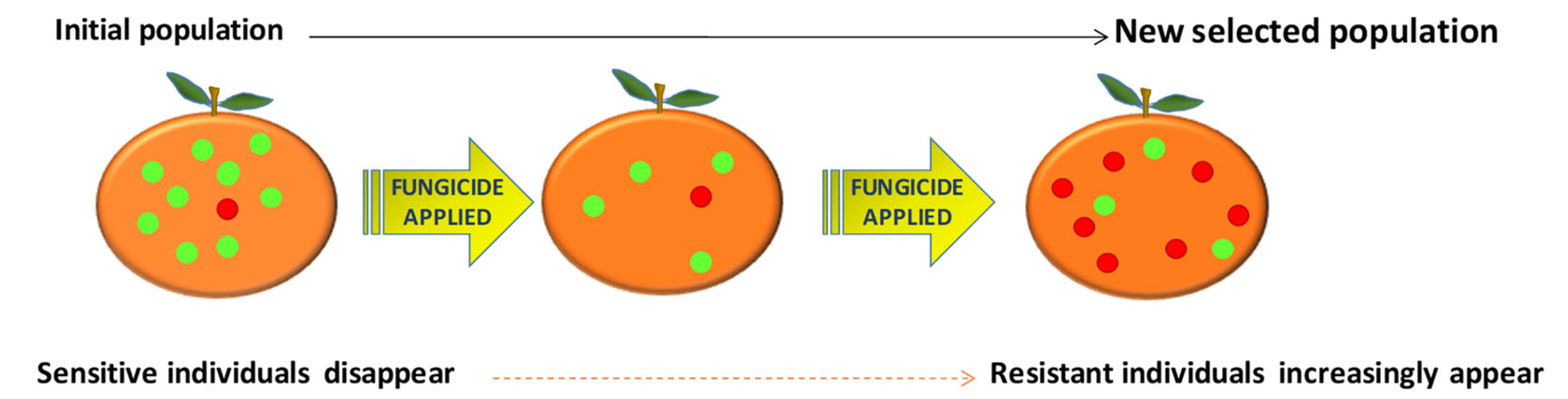
2. Fungicide Resistance Has Become a Major Problem
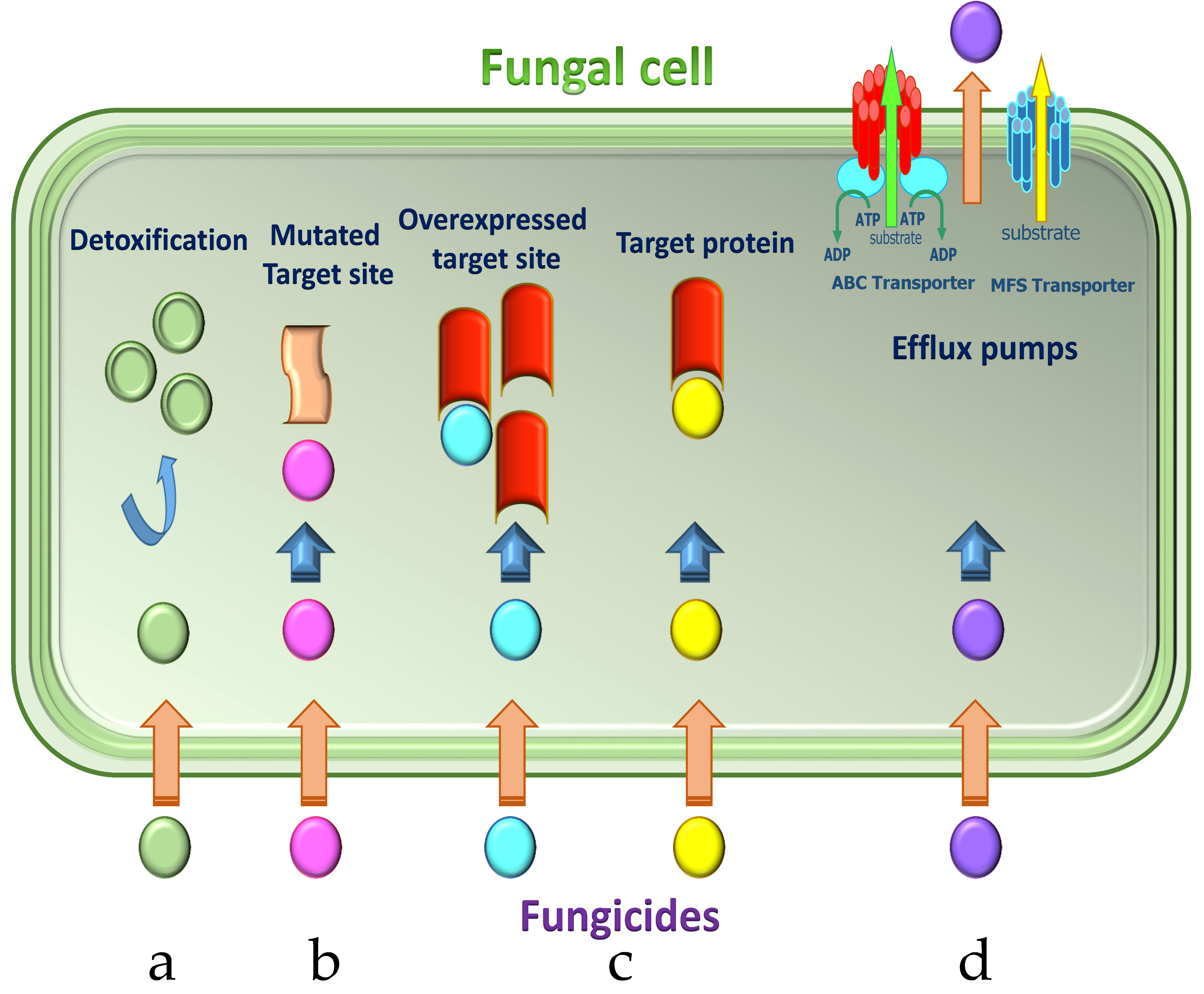
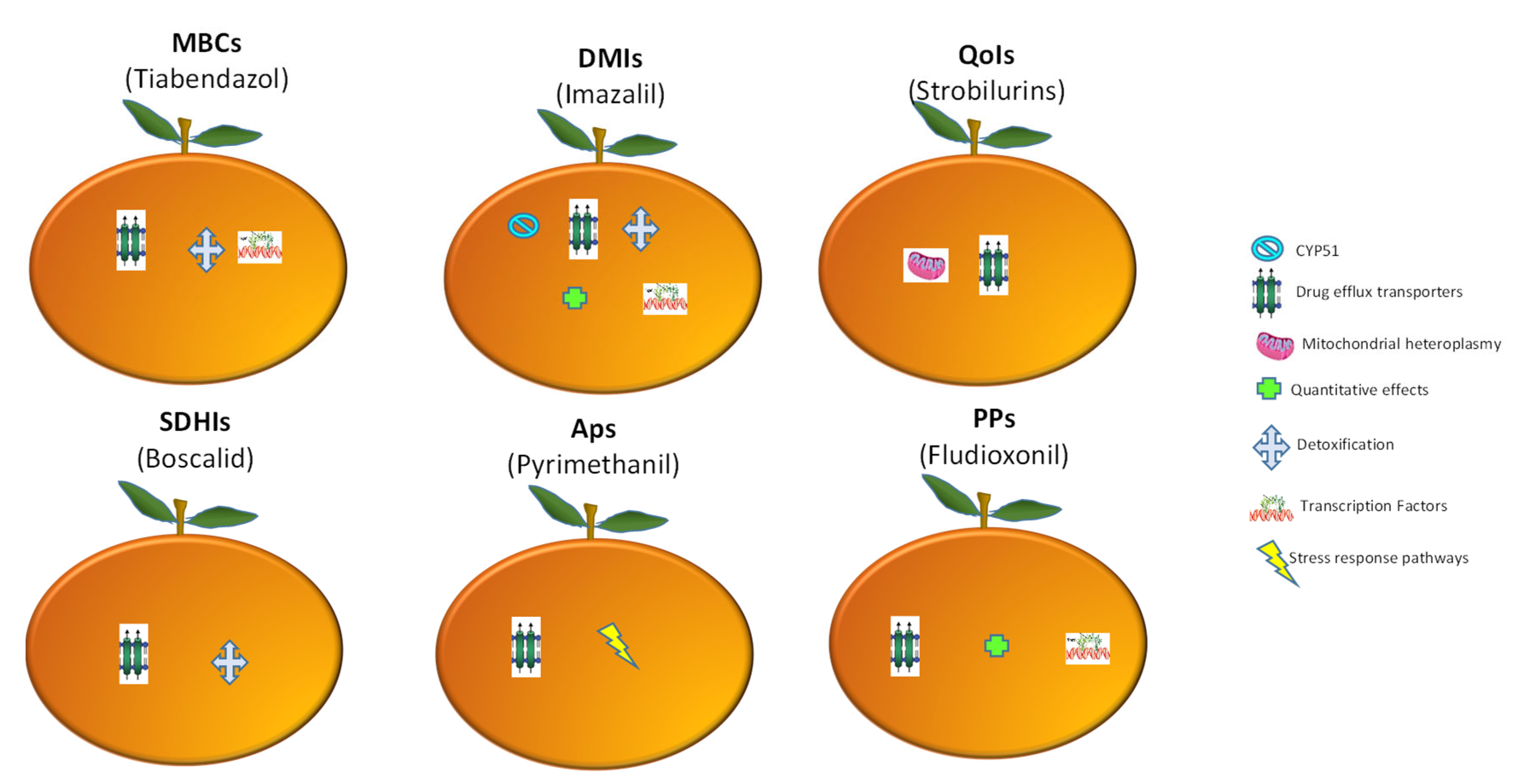
3. Molecular Mechanisms of Fungicide Resistance
3.1. Methyl Benzimidazoles (MBCs)
3.2. Demethylation Inhbithors (DMIs)
3.3. Quinone Outside Inhibitors (QoI)
3.4. Succinate Dehydrogenase Inhibitors (SDHIs)
3.5. Phenylpyrroles (PPs) and Anilinopyrimidines (Aps)
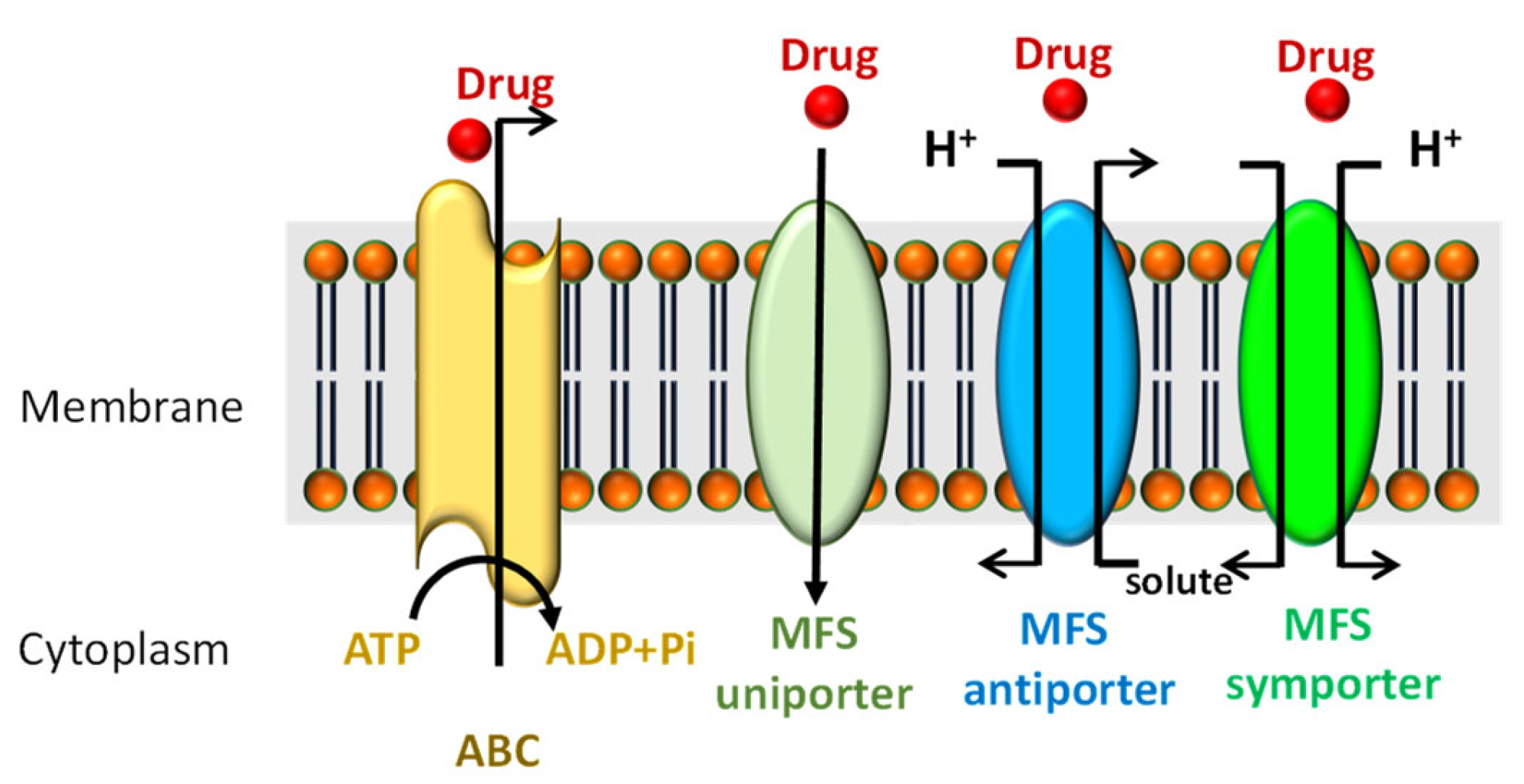
4. Resistance-Mediated Drug Efflux Transporters
4.1. ATP-Binding Cassette Transporters (ABC)
4.2. Major Facilitator Superfamily Transporters (MFS)
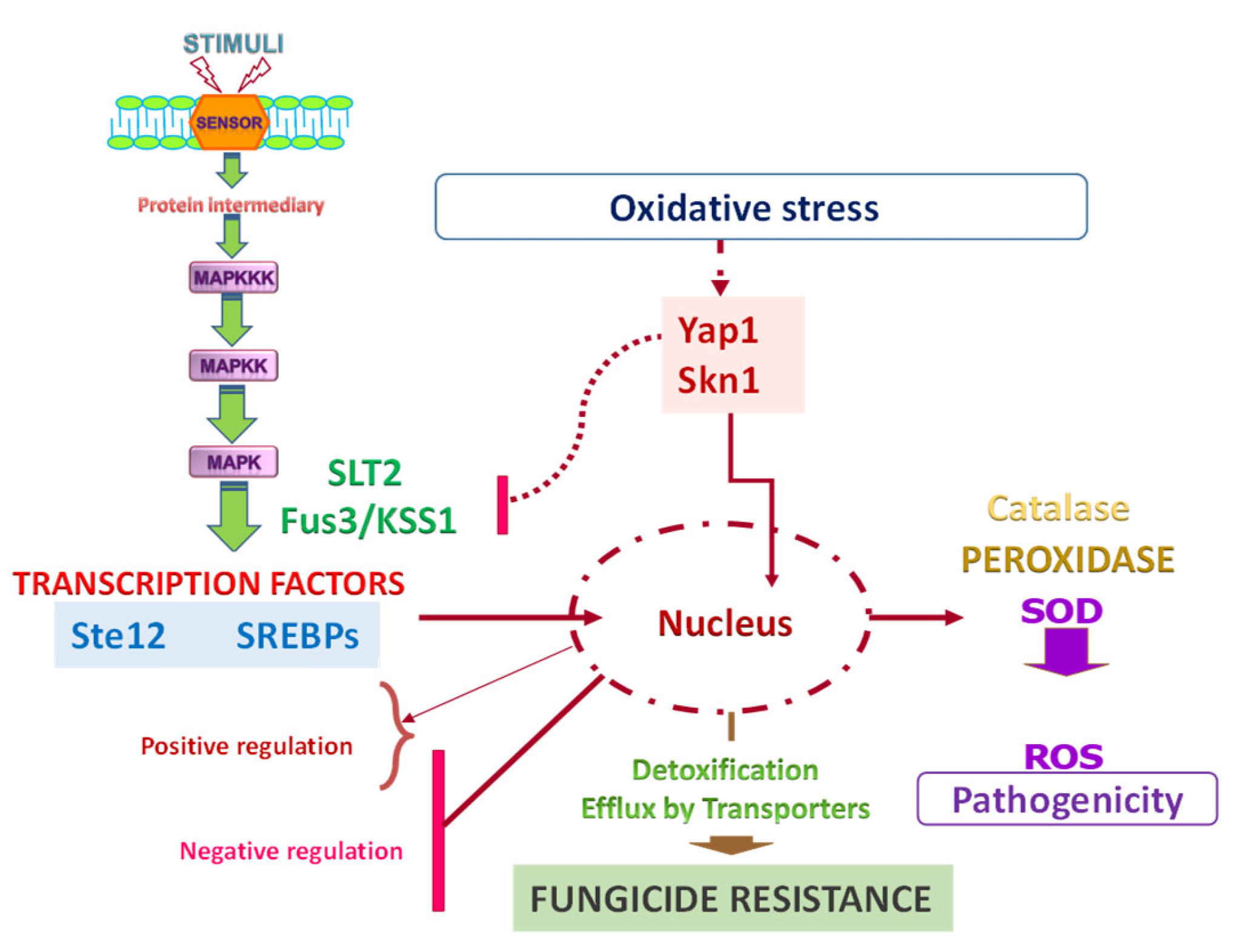
5. Regulation of Fungicide Resistance
5.1. Transcription Factors in Pd Fungicide Resistance
5.2. Protein Kinases in Pd Fungicide Resistance
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Kanetis, L.; Forster, H.; Adaskaveg, J.E. Baseline sensitivities for the new postharvest fungicides against Penicillium species on citrus and multiple resistance evaluations in P. digitatum. Plant Dis. 2008, 92, 261–269. [Google Scholar] [CrossRef]
- Palou, L. Penicillium digitatum and Pencillium italicum (Green Mold, Blue Mold). In Postharvest Decay: Control Strategies; Bautista-Baños, S., Ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 45–102. [Google Scholar]
- Tian, S.; Torres, R.; Ballester, A.R.; Li, B.; Vilanova, L.; Gonzalez-Candelas, L. Molecular aspects in pathogen-fruit interactions: Virulence and resistance. Postharvest Biol. Technol. 2016, 122, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Macarisin, D.; Cohen, L.; Eick, A.; Rafael, G.; Belausov, E.; Wisniewski, M.; Droby, S. Penicillium digitatum suppresses production of hydrogen peroxide in host tissue during infection of citrus fruit. Phytopathology 2007, 97, 1491–1500. [Google Scholar] [CrossRef] [Green Version]
- Marcet-Houben, M.; Ballester, A.R.; de la Fuente, B.; Harries, E.; Marcos, J.F.; González-Candelas, L.; Gabaldón, T. Genome sequence of the necrotrophic fungus Penicillium digitatum, the main postharvest pathogen of citrus. BMC Genom. 2012, 13, 646. [Google Scholar] [CrossRef] [Green Version]
- Perez, M.F.; Ibarreche, J.P.; Isas, A.S.; Sepulveda, M.; Ramallo, J.; Dib, J.R. Antagonistic yeasts for the biological control of Penicillium digitatum on lemons stored under export conditions. Biol. Control 2017, 115, 135–140. [Google Scholar] [CrossRef]
- Ghooshkhaneh, N.G.; Golzarian, M.R.; Mamarabadi, M. Detection and classification of citrus green mold caused by Penicillium digitatum using multispectral imaging. J. Sci. Food Agric. 2018, 98, 3542–3550. [Google Scholar] [CrossRef]
- Vu, T.X.; Ngo, T.T.; Mai, L.T.D.; Bui, T.T.; Le, D.H.; Bui, H.T.V.; Nguyen, H.Q.; Ngo, B.X.; Tran, V.T. A highly efficient Agrobacterium tumefaciens-mediated transformation system for the postharvest pathogen Penicillium digitatum using DsRed and GFP to visualize citrus host colonization. J. Microbiol. Meth. 2018, 144, 134–144. [Google Scholar] [CrossRef]
- Ismail, M.A.; Zhang, J. Post-harvest citrus diseases and their control. Outlooks Pest Manag. 2004, 15, 29–35. [Google Scholar] [CrossRef]
- De Ramón-Carbonell, M.; Sánchez-Torres, P. The transcription factor PdSte12 contributes to Penicillium digitatum virulence during citrus fruit infection. Postharvest Biol. Technol. 2017, 125, 129–139. [Google Scholar] [CrossRef]
- Zhang, T.; Qian, X.; Sun, X.; Li, H. The calcineurin-responsive transcription factor Crz1 is required for conidation, full virulence and DMI resistance in Penicillium digitatum. Microbiol. Res. 2013, 168, 211–222. [Google Scholar] [CrossRef]
- Smilanick, J.L.; Mansour, M.; Gabler, F.M.; Sorenson, D. Control of citrus postharvest green mold and sour rot by potassium sorbate combined with heat and fungicides. Postharvest Biol. Technol. 2008, 47, 226–238. [Google Scholar] [CrossRef]
- Hollomon, D.W. Fungicide resistance: Facing the challenge. Plant Protect. Sci. 2015, 51, 170–176. [Google Scholar] [CrossRef]
- Ma, Z.; Michailides, T.J. Advances in understanding molecular mechanisms of fungicide resistance and molecular detection of resistant genotypes in phytopathogenic fungi. Crop Prot. 2005, 24, 853–863. [Google Scholar] [CrossRef]
- Brent, K.J.; Hollomon, D. Fungicide Resistance in Crop Pathogens. How Can It Be Managed, 2nd ed.; FRAC: Brussels, Belgium, 2007. [Google Scholar]
- Hahn, M.; Leroch, M. Multidrug efflux transporters. In Fungicide Resistance in Plant Pathogens; Springer: Berlin/Heidelberg, Germany, 2015; pp. 233–248. [Google Scholar]
- Lucas, J.A.; Hawkins, N.J.; Fraaije, B.A. The evolution of fungicide resistance. Adv. Appl. Microbiol. 2015, 90, 29–92. [Google Scholar]
- Deising, H.B.; Reimann, S.; Pascholati, S.F. Mechanisms and significance of fungicide resistance. Brazilian J. Microbiol. 2008, 39, 286–295. [Google Scholar] [CrossRef] [Green Version]
- Gisi, U.; Chin, K.M.; Knapova, G.; Küng Färber, R.; Mohr, U.; Parisi, S.; Sierotzki, H.; Steinfeld, U. Recent developments in elucidating mode of resistance to phenylamide, DMI, and strobilurin fungicides. Crop Prot. 2000, 19, 863–872. [Google Scholar] [CrossRef]
- Gullino, M.L.; Leroux, P.; Smith, C.M. Uses and challenges of novel compounds for plant disease control. Crop Prot. 2000, 19, 1–11. [Google Scholar] [CrossRef]
- Fluit, A.C.; Visser, M.R.; Schmitz, F.J. Molecular detection of antimicrobial resistance. Clin. Microbiol. Rev. 2001, 14, 836–871. [Google Scholar] [CrossRef] [Green Version]
- Capote, N.; Pastrana, A.M.; Aguado, A.; Sánchez-Torres, P. Molecular Tools for Detection of Plant Pathogenic Fungi and Fungicide Resistance. In Plant Pathology; Cumagun, C.J.R., Ed.; InTech: East Providence, RI, USA, 2012; pp. 151–202. [Google Scholar]
- Langner, T.; Kamoun, S.; Belhaj, K. CRISPR crops: Plant genome editing toward disease resistance. Annu. Rev. Phytopathol. 2018, 56, 479–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burón-Moles, G.; López-Pérez, M.; González-Candelas, L.; Vinas, I.; Teixido, N.; Usall, J.; Torres, R. Use of GFP-tagged strains of Penicillium digitatum and Penicillium expansum to study host-pathogen interactions in oranges and apples. Int. J. Food Microbiol. 2012, 160, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.H.; Bazioli, J.M.; de Moraes Pontes, J.G.; Fill, T.P. Penicillium digitatum infection mechanisms in citrus: What do we know so far? Fungal Biol. 2019, 123, 584–593. [Google Scholar] [CrossRef]
- Holloman, D.W.; Butters, J.A.; Barker, H.; Hall, L. Fungal β-tubulin, expressed as a fusion protein, binds benzimidazole and phenylcarbamate fungicides. Antimicrob. Agents Chemother. 1998, 42, 2171–2173. [Google Scholar] [CrossRef] [Green Version]
- Baraldi, E.; Mari, M.; Chierici, E.; Pondrelli, M.; Bertollini, P.; Pratella, G.C. Studies of Thiabendazole resistance of Penicillium expansum of pears: Pathogenic fitness ang genetic characterization. Plant Pathol. 2003, 52, 362–370. [Google Scholar] [CrossRef]
- Leroux, P.; Fritz, R.; Debieu, D.; Albertini, C.; Lanen, C.; Bach, J.; Gredt, M.; Chapeland, F. Mechanisms of resistance to fungicides in field strains of Botrytis cinerea. Pest Manag. Sci. 2002, 58, 876–888. [Google Scholar] [CrossRef]
- Malandrakis, A.; Markoglou, A.; Ziogas, B. Molecular characterization of benzimidazole-resistant B. cinerea field isolates with reduced or enhanced sensitivity to zoxamide and diethofencarb. Pestic. Biochem. Physiol. 2011, 99, 118–124. [Google Scholar] [CrossRef]
- Fan, J.; Luo, Y.; Michailides, T.J.; Guo, L. Simultaneous quantification of alleles E198A and H6Y in the β-tubulin gene conferring benzimidazole resistance in Monilinia fructicola using a duplex real-time (TaqMan) PCR. Pest Manag. Sci. 2014, 70, 245–251. [Google Scholar] [CrossRef]
- Albertini, C.; Gredt, M.; Leroux, P. Mutations of the β-tubulin gene associated with different phenotypes of benzimidazole resistance in the cereal eyespot fungi Tapesia yallundae and Tapesia acuformis. Pestic. Biochem. Physiol. 1999, 64, 17–31. [Google Scholar] [CrossRef]
- Yarden, O.; Katan, T. Mutations leading to substitutions at amino acids 198 and 200 of beta-tubulin that correlates with benomyl-resistance phenotypes of field strains of Botrytis cinerea. Phytopathology 1993, 83, 1478–1483. [Google Scholar] [CrossRef]
- Koenraadt, H.; Somerville, S.C.; Jones, A.L. Characterization of mutations in the beta-tubulin gene of benomyl-resistant field strains of Venturia inaequalis and other plant pathogenic fungi. Phytopathology 1992, 82, 1348–1354. [Google Scholar] [CrossRef]
- Schmidt, L.S.; Ghosoph, J.M.; Margosan, D.A.; Smilanick, J.L. Mutation at β-tubulin codon 200 indicated Thiabendazole resistance in Penicillium digitatum collected from California citrus packinghouses. Plant Dis. 2006, 90, 765–770. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Torres, P.; Tuset, J.J. Molecular insights into fungicide resistance in sensitive and resistant Penicillium digitatum strains infecting citrus. Postharvest Biol. Technol. 2011, 59, 159–165. [Google Scholar] [CrossRef]
- Lee, M.-H.; Pan, S.-M.; Ng, T.-W.; Chen, P.-S.; Wang, L.-Y.; Chung, K.-R. Mutations of β-tubulin codon 198 or 200 indicate thiabendazole resistance among isolates of Penicillium digitatum collected from citrus in Taiwan. Int. J. Food Microbiol. 2011, 150, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Vela-Corcía, D.; Romero, D.; de Vicente, A.; Pérez-García, A. Analysis of β-tubulin-carbendazim interaction reveals that binding site for MBC fungicides does not include residues involved in fungicide resistance. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Brent, K.J.; Hollomon, D.W. Fungicide Resistance: The Assessment of Risk, 2nd ed.; Global Crop Protection Federation: Brussels, Belgium, 1998; pp. 1–48. [Google Scholar]
- Aoyama, Y.; Yoshida, Y. Different substrate specificities of lanosterol 14a-demethylase (P-45014DM) of Saccharomyces cerevisiae and rat liver for 24-methylene-24,25-dihydrolanosterol and 24,25-dihydrolanosterol. Biochem. Biophys. Res. Commun. 1991, 178, 1064–1071. [Google Scholar] [CrossRef]
- Hamamoto, H.; Hasegawa, K.; Nakaune, R.; Lee, Y.J.; Makizumi, Y.; Akutsu, K.; Hibi, T. Tandem repeat of a transcription enhancer upstream of the sterol 14_demethylase gene (CYP51) Penicillium digitatum. Appl. Environ. Microbiol. 2000, 66, 3421–3426. [Google Scholar] [CrossRef] [Green Version]
- Nakaune, R.; Hamamoto, H.; Imada, J.; Akutsu, K.; Hibi, T. A novel ABC transporter gene, PMR5, is involved in multidrug resistance in the phytopathogenic fungus Penicillium digitatum. Mol. Genet. Genom. 2002, 267, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Holmes, G.J.; Eckert, J.W. Relative fitness of Imazalil-resistant and -sensitive biotypes of Penicillium digitatum. Plant Dis. 1995, 79, 1068–1073. [Google Scholar] [CrossRef]
- Parks, L.W.; Casey, W.M. Physiological implications of sterol biosynthesis in yeast. Annu. Rev. Microbiol. 1995, 49, 95–116. [Google Scholar] [CrossRef]
- Ali, E.M.; Amiri, A. Selection pressure pathways and mechanisms of resistance to the demethylation inhibitor difenoconazole in Penicillium expansum. Front. Microbiol. 2018, 9, 2472. [Google Scholar] [CrossRef]
- Snelders, E.; Camps, S.M.T.; Karawajczyk, A.; Rijs, A.J.M.M.; Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Genotype–phenotype complexity of the TR46/Y121F/T289A cyp51A azole resistance mechanism in Aspergillus fumigatus. Fungal Genet. Biol. 2015, 82, 129–135. [Google Scholar] [CrossRef]
- Wang, J.; Yu, J.; Liu, J.; Yuan, Y.; Li, N.; He, M.; Qi, T.; Hui, G.; Xiong, L.; Liu, D. Novel mutations in CYP51B from Penicillium digitatum involved in prochloraz resistance. J. Microbiol. 2014, 52, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Urban, M.; Parker, J.E.; Brewer, H.C.; Kelly, S.L.; Hammond-Kosack, K.E.; Fraaije, B.A.; Liu, X.; Cools, H.J. Characterization of the sterol 14α-demethylases of Fusarium graminearum identifies a novel genus-specific cyp51 function. New Phytol. 2013, 198, 821–835. [Google Scholar] [CrossRef] [Green Version]
- Hamamoto, H.; Nawata, O.; Hasegawa, K.; Nakaune, R.; Lee, Y.J.; Makizumi, Y.; Akutsu, K.; Hibi, T. The role of the ABC transporter gene PMR1 in demethylation inhibitor resistance in Penicillium digitatum. Pestic. Biochem. Phys. 2001, 70, 19–26. [Google Scholar] [CrossRef]
- Sun, X.; Wang, J.; Feng, D.; Ma, Z.; Li, H. PdCYP51B, a new putative sterol 14α-demethylase gene of Penicillium digitatum involved in resistance to imazalil and other fungicides inhibiting ergosterol synthesis. Appl. Microbiol. Biot. 2011, 91, 1107–1119. [Google Scholar] [CrossRef]
- Délye, C.; Laigret, F.; Corio-Costet, M.F. A mutation in the 14α-Demethylase gene of Uncinula necator that correlatos with resistance to a sterol biosynthesis inhibitor. Appl. Environ. Microb. 1997, 63, 2966–2970. [Google Scholar] [CrossRef] [Green Version]
- Délye, C.; Bousset, L.; Corio-Costet, M.F. PCR cloning and detection of point mutations in the eburicol 14alpha-demethylase (CYP51) gene from Erysiphe graminis f. sp. hordei, a “recalcitrant” fungus. Curr. Genet. 1998, 34, 399–403. [Google Scholar]
- Rallos, L.E.E.; Baudoin, A.B. Co-occurrence of two allelic variants of CYP51 in Erysiphe necator and their correlation with over-expression for DMI resistance. PLoS ONE 2016, 11, e0148025. [Google Scholar] [CrossRef] [PubMed]
- Wyand, R.A.; Brown, J.K.M. Sequence variation in the CYP51 gene of Blumeria graminis associated with resistance to sterol demethylase inhibiting fungicides. Fungal Genet. Biol. 2005, 42, 726–765. [Google Scholar] [CrossRef]
- Albertini, C.; Gredt, M.; Leroux, P. Polymorphism of the 14α demethylase gene (CYP51) in the cereal eyespot fungi Tapesia acuformis and Tapesia yallundae. Eur. J. Plant Pathol. 2003, 109, 117–128. [Google Scholar] [CrossRef]
- Joseph-Horne, T.; Hollomon, D.W. Molecular mechanisms of azole resistance. FEMS Microbiol. Lett. 1997, 149, 141–149. [Google Scholar] [CrossRef]
- Butters, J.A.; Zhou, M.; Hollomon, D.W. The mechanism of resistance to sterol 14α-demethylation inhibitors in a mutant (Erg40) of Ustilago maydis. Pest Manag. Sci. 2000, 56, 257–263. [Google Scholar] [CrossRef]
- Ma, Z.; Proffer, T.J.; Jacobs, J.L.; Sundin, G.W. Overexpression of the 14α-Demethylase Target Gene (CYP51) Mediates Fungicide Resistance in Blumeriella jaapii. Appl. Environ. Microbiol. 2006, 72, 2581–2585. [Google Scholar] [CrossRef] [Green Version]
- Leroux, P.; Walker, A.S. Activity of fungicides and modulators of membrane drug transporters in field strains of Botrytis cinerea displaying multidrug resistance. Eur. J. Plant Pathol. 2013, 135, 683–693. [Google Scholar] [CrossRef]
- De Ramón-Carbonell, M.; Sánchez-Torres, P. Significance of 195 bp-enhancer of PdCYP51B in the acquisition of Penicillium digitatum DMI resistance and increase of fungal virulence. Pestic. Biochem. Physiol. 2020, 165, 104522. [Google Scholar] [CrossRef]
- Cools, H.J.; Bayon, C.; Atkins, S.; Lucas, J.A.; Fraaije, B.A. Overexpression of the sterol 14 alpha-demethylase gene (MgCYP51) in Mycosphaerella graminicola isolates confers a novel azole fungicide sensitivity phenotype. Pest Manag. Sci. 2012, 68, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Schnabel, G.; Jones, A.L. The 14α-demethylasse (CYP51A1) gene is overexpressed in Venturia Inaequalis strains resistant to myclobutanil. Phytopathology 2001, 91, 102–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, C.X.; Schnabel, G. The cytochrome P450 lanosterol 14α-demethylase gene is a demethylation inhibitor fungicide resistance determinant in Monilinia fructicola field isolates from Georgia. Appl. Environ. Microbiol. 2008, 74, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Ghosoph, J.M.; Schmidt, L.S.; Margosan, D.A.; Smilanick, J.L. Imazalil resistance linked to a unique insertion sequence in the PdCYP51 promoter region of Penicillium digitatum. Postharvest Biol. Technol. 2007, 44, 9–18. [Google Scholar] [CrossRef]
- Sun, X.; Ruan, R.; Lin, L.; Zhu, C.; Zhang, T.; Wang, M.; Li, H.; Yu, D. Genomewide investigation into DNA elements and ABC transporters involved in imazalil resistance in Penicillium digitatum. FEMS Microbiol. Lett. 2013, 348, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzansk, B. The strobilurin fungicides. Pest Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Uesugi, Y. Fungicide classes: Chemistry, uses and mode of action. In Fungicidal Activity—Chemical and Biological Approaches to Plant Protection; Hutson, D., Miyamoto, J., Eds.; John Wiley & Sons: New York, NY, USA, 1998; pp. 23–56. [Google Scholar]
- Kanetis, L.; Forster, H.; Adaskaveg, J.E. Comparative efficacy of the new postharvest fungicides Azoxystrobin, fludioxonil, and pyrimethanil for managing citrus green mold. Plant Dis. 2007, 91, 1502–1511. [Google Scholar] [CrossRef] [Green Version]
- Smilanick, J.L.; Mansour, M.F.; Sorenson, D. Pre- and postharvest treatments tocontrol green mold of citrus fruit during ethylene degreening. Plant Dis. 2006, 90, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.F.; Zhao, Q.; Liu, K.Y.; Zhang, Z.G.; Wang, Y.C.; Zheng, X.B. MgCRZ1, a transcription factor of Magnaporthe grisea, controls growth, development and is involved in full virulence. FEMS Microbiol. Lett. 2009, 293, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Ishii, H.; Fraaije, B.A.; Sugiyama, T.; Noguchi, K.; Nishimura, K.; Takeda, T.; Amano, T.; Hollomon, D.W. Ocurrence and molecular characterization of strobilurin resistance in cucumber powdery mildew and downy mildew. Phytopathology 2001, 91, 1166–1171. [Google Scholar] [CrossRef] [Green Version]
- Fraaije, B.A.; Butters, J.A.; Coelho, J.M.; Jones, D.R.; Hollomon, D.W. Following the dynamics of strobilurin resistance in Blumeria graminis f Sp. tritici using quantitative allele-specific real-time PCR measurements with the fluorescent dye SYBR Green I. Plant Pathol. 2002, 51, 45–54. [Google Scholar]
- Fraaije, B.A.; Cools, H.J.; Fountaine, J.; Lovell, D.; Motteram, J.J.; West, J.S.; Lucas, J.A. Role of ascospores in further QoI-resistant cytochrome b alleles (G143A) in field populations Mycosphaerella graminicola. Phytopathology 2005, 95, 933–941. [Google Scholar] [CrossRef] [Green Version]
- Baumler, S.; Felsenstein, F.G.; Schwarz, G. CAPS and DHPLC analysis of a single nucleotide polymorphism in the cytochrome b gene conferring resistance to strobilurins in field isolates of Blumeria graminis f.sp. hordei. J. Phytopathol. 2003, 151, 149–152. [Google Scholar] [CrossRef]
- Ma, Z.; Yoshimura, M.A.; Michailides, T.J. Identification and characterization of benzimidazole resistance in Monilinia fructicola from stone fruit orchards in California. Appl. Environ. Biol. 2003, 69, 7145–7152. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Dixon, E.W.; Vincelli, P.; Farman, M.L. Field resistance to strobilurin (QoI) fungicides in Pyricularia grisea caused by mutations in the mitochondrial cytochrome b gene. Phytopathology 2003, 93, 891–900. [Google Scholar] [CrossRef] [Green Version]
- Gisi, U.; Sierotzki, H.; Cook, A.; McCaffery, A. Mechanisms influencing the evolution of resistance to Qo inhibitor fungicides. Pest Manag. Sci. 2002, 58, 859–867. [Google Scholar] [CrossRef]
- Sierotzki, H.; Frey, R.; Wullschleger, J.; Palermo, S.; Karlin, S.; Godwin, J.; Gisi, U. Cytochrome b gene sequence and structure of Pyrenophora teres and P. tritici-repentis and implications for QoI resistance. Pestic. Manag. Sci. 2007, 63, 225–233. [Google Scholar] [CrossRef]
- Wood, P.M.; Hollomon, D.W. A critical evaluation of the role of alternative oxidase in the performance of strobilurin and related fungicides acting at the Qo site of Complex III. Pest Manag. Sci. 2003, 59, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Ortuño, D.; Torés, J.A.; de Vicente, A.; Pérez-García, A. Mechanisms of resistance to QoI fungicides in phytopathogenic fungi. Int. Microbiol. Off. J. Span. Soc. Microbiol. 2008, 11, 1–9. [Google Scholar]
- Avenot, H.F.; Sellam, A.; Karaoglanidis, G.; Michailides, T.J. Characterization of mutations in the iron sulfur subunit of succinate dehydrogenase correlating with boscalid resistance in Alternaria alternata from California pistachio. Phytopathology 2008, 98, 736–742. [Google Scholar] [CrossRef] [Green Version]
- Avenot, H.F.; Michailides, T.J. Progress in understanding molecular mechanisms and evolution of resistance to succinate dehydrogenase inhibiting (SDHI) fungicides in phytopathogenic fungi. Crop Prot. 2010, 29, 643–651. [Google Scholar] [CrossRef]
- Avenot, H.F.; Morgan, D.P.; Michailides, T.J. Resistance to pyraclostrobin, boscalid and multiple resistance to Pristine® (pyraclostrobin + boscalid) fungicide in Alternaria alternata causing alternaria late blight of pistachios in California. Plant Pathol. 2008, 57, 135–140. [Google Scholar] [CrossRef]
- Fernández-Ortuño, D.; Pérez-García, A.; Chamorro, M.; de la Peña, E.; de Vicente, A.; Torés, J.A. Resistance to the SDHI Fungicides Boscalid, Fluopyram, Fluxapyroxad, and Penthiopyrad in Botrytis cinerea from Commercial Strawberry Fields in Spain. Plant Dis. 2017, 101, 17–0067. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, T.; Ishii, H.; Stammler, G.; Koch, A.; Ogawara, T.; Tomita, Y.; Kobori, S. Distribution and molecular characterization of Corynespora cassiicola isolates resistant to boscalid. Plant Pathol. 2010, 59, 873–881. [Google Scholar] [CrossRef]
- Wang, Y.; Duan, Y.; Wang, J.; Zhou, M.A. New point mutation in the iron–sulfur subunit of succinatedehydrogenase confers resistance to boscalid inSclerotinia sclerotiorum. Mol. Plant Pathol. 2015, 16, 653–661. [Google Scholar] [CrossRef]
- Avenot, H.F.; Michailides, T.J. Resistance to boscalid fungicide in Alternaria alternata isolates from pistachio in California. Plant Dis. 2007, 91, 1345–1350. [Google Scholar] [CrossRef] [Green Version]
- Veloukas, T.; Leroch, M.; Hahn, M.; Karaoglanidis, G.S. Detection and molecular characterization of boscalid-resistant Botrytis cinerea isolates from strawberry. Plant Dis. 2011, 95, 1302–1307. [Google Scholar] [CrossRef] [Green Version]
- Fraaije, B.A.; Bayon, C.; Atkins, S.; Cools, H.J.; Lucas, J.A.; Fraaije, M.W. Risk assessment studies on succinate dehydrogenase inhibitors, the new weapons in the battle to control Septoria leaf blotch in wheat. Mol. Plant Pathol. 2012, 13, 263–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalliet, G.; Bowler, J.; Luksch, T.; Kirchhofer-Allan, L.; Steinhauer, D.; Ward, K.; Niklaus, M.; Verras, A.; Csukai, M.; Daina, A.; et al. Mutagenesis and Functional Studies with Succinate Dehydrogenase Inhibitors in the Wheat Pathogen Mycosphaerella graminicola. PLoS ONE 2012, 7, e35429. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Zhang, K.; Fu, Y.; Ma, H.; Zhu, F. Toxic action and baseline sensitivity of boscalid against Penicillium digitatum. Crop Prot. 2020, 137, 105272. [Google Scholar] [CrossRef]
- Murphy, M.P.; Holmgren, A.; Larsson, N.G.; Halliwell, B.; Chang, C.J.; Kalyanaraman, B.; Rhee, S.G.; Thornalley, P.J.; Partridge, L.; Gems, D.; et al. Unraveling the biological roles of reactive oxygen species. Cell Metab. 2011, 13, 361–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FRAC. Pathogen Risk List. Available online: https://www.frac.info/docs/default-source/publications/pathogen-risk/frac-pathogen-list-2019.pdf?sfvrsn=caf3489a_2 (accessed on 19 May 2020).
- Schirra, M.; D’aquino, S.; Mulas, M.; Melis, R.A.M.; Giobbe, S.; Migheli, Q.; Garau, A.; Angioni, A.; Cabras, P. Efficacy of Heat Treatments with Water and Fludioxonil for Postharvest Control of Blue and Gray Molds on Inoculated Pears and Fludioxonil Residues in Fruit. J Food Prot. 2008, 71, 967–972. [Google Scholar] [CrossRef]
- Li, H.X.; Xiao, C.L. Characterization of fludioxonil-resistant and pyrimethanil-resistant phenotypes of Penicillium expansum from apple. Phytopathology 2008, 98, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.K.; Saito, S.; Xiao, C.L. Occurrence of Fludioxonil Resistance in Penicillium digitatum from Citrus in California. Plant Dis. 2015, 99, 1447. [Google Scholar] [CrossRef]
- Kinay, P.; Mansour, M.F.; Gabler, F.M.; Margosan, D.A.; Smilanick, J.L. Characterization of fungicide-resistan isolates of Penicillium digitatum collected in California. Crop Prot. 2007, 26, 647–656. [Google Scholar] [CrossRef]
- Fritz, R.; Lanen, C.; Chapeland-Leclerc, F.; Leroux, P. Effect of the anilinopyrimidine fungicide pyrimethanil on the cystathionine β-lyase of Botrytis cinerea. Pestic. Biochem. Physiol. 2003, 77, 54–65. [Google Scholar] [CrossRef]
- Del Sorbo, G.; Schoonbeek, H.-J.; de Waard, M.A. Fungal transporters involved in efflux of natural toxic compounds and fungicides. Fungal Gen. Biol. 2000, 30, 1–15. [Google Scholar] [CrossRef]
- Kretschmer, M.; Leroch, M.; Mosbach, A.; Walker, A.-S.; Fillinger, S.; Mernke, D.; Schoonbeek, H.-J.; Pradier, J.-M.; Leroux, P.; De Waard, M.A.; et al. Fungicide-driven evolution and molecular basis of multidrug resistance in field populations of the grey mould fungus Botrytis cinerea. PLoS Pathog. 2009, 5, e1000696. [Google Scholar] [CrossRef]
- Omrane, S.; Sghyer, H.; Audéon, C.; Lanen, C.; Duplaix, C.; Walker, A.-S.; Fillinger, S. Fungicide efflux and the MgMFS 1 transporter contribute to the multidrug resistance phenotype in Zymoseptoria tritici Field Isolates. Environ. Microbiol. 2015, 17, 2805–2823. [Google Scholar] [CrossRef]
- De Ramón-Carbonell, M.; López-Pérez, M.; González-Candelas, L.; Sánchez-Torres, P. PdMFS1 transporter contributes to Penicilliun digitatum fungicide resistance and fungal virulence during citrus fruit infection. J. Fungi 2019, 5, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ramón-Carbonell, M.; Sánchez-Torres, P. Penicillium digitatum MFS transporters can display different roles during pathogen-fruit interaction. Int. J. Food Microbiol. 2021, 337, 108918. [Google Scholar] [CrossRef] [PubMed]
- Perlin, M.H.; Andrews, J.; San Toh, S. Essential letters in the fungal alphabet: ABC and MFS transporters and their roles in survival and pathogenicity. In Advances in Genetics; Elsevier: Amsterdam, The Netherlands, 2014; Volume 85, pp. 201–253. [Google Scholar]
- Kuroda, T.; Tsuchiya, T. Multidrug efflux transporters in the MATE family. Biochim. Biophys. Acta 2009, 1794, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Chen, S. Non-Target Site Mechanisms of Fungicide Resistance in Crop Pathogens: A Review. Microorganisms 2021, 9, 502. [Google Scholar] [CrossRef]
- Stergiopoulos, I.; van Nistelrooy, J.G.; Kema, G.H.; De Waard, M.A. Multiple mechanisms account for variation in base-line sensitivity to azole fungicides in field isolates of Mycosphaerella graminicola. Pest Manag. Sci. 2003, 59, 1333–1343. [Google Scholar] [CrossRef]
- De Waard, M.A.; Andrade, A.C.; Hayashi, K.; Schoonbeek, H.; Stergiopoulos, I.; Zwiers, L.H. Impact of fungal drug transporters on fungicide sensitivity, multidrug resistance and virulence. Pest Manag. Sci. 2006, 62, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Schoonbeek, H.; Raaijmakers, J.M.; De Waard, M.A. Fungal ABC transporters and microbial interactions in natural environments. Mol. Plant Microbe Int. 2002, 15, 1165–1172. [Google Scholar] [CrossRef] [Green Version]
- Nakaune, R.; Adachi, K.; Nawata, O.; Tomiyama, M.; Akutsu, K.; Hibi, T. A novel ATP-binding cassette transporter involved in multidrug resistance in the phytopathogenic fungus Penicillium digitatum. Appl. Environ. Microbiol. 1998, 64, 3983–3988. [Google Scholar] [CrossRef] [Green Version]
- Nakaune, R. ABC transporters genes involved in multidrug resistance in Penicillium digitatum. J. Gen. Pathol. 2001, 67, 251. [Google Scholar] [CrossRef]
- Liu, J.; Yuan, Y.; Wu, Z.; Li, N.; Chen, Y.; Qin, T.; Geng, H.; Xiong, L.; Liu, D. A novel sterol regulatory element-binding protein gene (sreA) identified in Penicillium digitatum is required for prochloraz resistance, full virulence and erg11 (cyp51) regulation. PLoS ONE 2015, 10, e0117115. [Google Scholar]
- Drew, D.; North, R.A.; Nagarathinam, K.; Tanabe, M. Structures and General Transport Mechanisms by the Major Facilitator Superfamily (MFS). Chem. Rev. 2021, 121, 5289–5335. [Google Scholar] [CrossRef]
- Dos Santos, S.C.; Teixeira, M.C.; Dias, P.J.; Sá-Correia, I. MFS transporter required for multidrug/multixenobiotic (MD/MX) resistance in the model yeast: Understanding their physiological function through postgenomic approaches. Front. Physiol. 2014, 5, 180. [Google Scholar] [CrossRef] [Green Version]
- Choquer, M.; Lee, M.; Bau, H.; Chung, K. Deletion of a MFS transporter like gene in Cercospora nicotianae reduces cercosporin toxin accumulation and fungal virulence. FEMS Lett. 2007, 581, 489–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, K.; Schoonbek, H.J.; de Waard, M.A. BcmfsA novel major facilitator superfamily tranporter from Botrytis cinerea, provides tolerance towards the natural toxic compounds camptothecin and cercosporin and towards fungicides. Appl. Environ. Microbiol. 2002, 68, 4996–5004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roohparvar, R.; De Waard, M.A.; Kema, G.H.; Zwiers, L.H. MgMfs1, a major facilitator superfamily transporter from the fungal wheat pathogen Mycosphaerella graminicola, is a strong protectant against natural toxic compounds and fungicides. Fungal Genet. Biol. 2007, 44, 378–388. [Google Scholar] [CrossRef]
- Roohparvar, R.; Mehrabi, R.; Van Nistelrooy, J.G.M.; Zwiers, L.; De Waard, M.A. The drug transporter MgMfs1 can modulate sensitivity of field strains of the fungal wheat pathogen Mycosphaerella graminicola to the strobilurin fungicide trifloxystrobin. Pest Manag. Sci. 2008, 64, 685–693. [Google Scholar] [CrossRef]
- Chen, L.H.; Tsai, H.C.; Chung, K.R. A Major Facilitator Superfamily Transporter-Mediated Resistance to Oxidative Stress and Fungicides Requires Yap1, Skn7, and MAP Kinases in the Citrus Fungal Pathogen Alternaria alternata. PLoS ONE 2017, 12, e0169103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samaras, A.; Ntasiou, P.; Myresiotis, C.; Karaoglanidis, G. Multidrug resistance of Penicillium expansum to fungicides: Whole transcriptome analysis of MDR strains reveals overexpression of efflux transporter genes. Int. J. Food Microbiol. 2020, 335, 108896. [Google Scholar] [CrossRef]
- Wang, J.Y.; Sun, X.P.; Lin, L.Y.; Zhang, T.Y.; Ma, Z.H.; Li, H.Y. PdMfs1, a major facilitator superfamily transporter from Penicillium digitatum, is partially involved in the imazalil-resistance and pathogenicity. Afr. J. Microbiol. Res. 2012, 6, 95–105. [Google Scholar]
- Wu, Z.; Wang, S.Q.; Yuan, Y.Z.; Zhang, T.F.; Liu, J.; Liu, D.L. A novel major facilitator superfamily transporter in Penicillium digitatum (PdMFS2) is required for prochloraz resistance, conidiation and full virulence. Biotechnol. Lett. 2016, 38, 1349–1357. [Google Scholar] [CrossRef]
- Du, D.; Wang-Kan, X.; Neuberger, A.; van Veen, H.W.; Pos, K.M.; Piddock, L.J.V.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Jin, J.; Kim, H.; Kim, J.C.; Yun, S.H.; Lee, Y.W. GIP2, a putative transcription factor that regulates the aurofusarin biosynthetic gene cluster in Gibberella zeae. Appl. Environ. Microbiol. 2006, 72, 1645–1652. [Google Scholar] [CrossRef] [Green Version]
- Hellauer, K.; Akache, B.; MacPherson, S.; Sirard, E.; Turcotte, B. Zinc cluster protein Rdr1p is a transcriptional repressor of the PDR5 gene encoding a multidrug transporter. J. Biol. Chem. 2002, 277, 17671–17676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bien, C.M.; Espenshade, P.J. Sterol regulatory element binding proteins in fungi: Hypoxic transcription factors linked to pathogenesis. Eukaryot. Cell. 2010, 9, 352–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, R.; Wang, M.; Liu, X.; Sun, X.; Chung, K.R.; Li, H. Functional analysis of two sterol regulatory element binding proteins in Penicillium digitatum. PLoS ONE 2017, 12, e0176485. [Google Scholar]
- Thakur, J.K.; Arthanari, H.; Yang, F.; Pan, S.J.; Fan, X.; Breger, J.; Frueh, D.P.; Gulshan, K.; Li, D.K.; Mylonakis, E.; et al. A nuclear receptor-like pathway regulating multidrug resistance in fungi. Nature 2008, 452, 604–609. [Google Scholar] [CrossRef]
- Sang, H.; Hulvey, J.P.; Green, R.; Xu, H.; Im, J.; Chang, T.; Jung, G. A xenobiotic detoxification pathway through transcriptional regulation in filamentous fungi. mBio 2018, 9, e00457-18. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Moye-Rowley, W.S. Multidrug resistance in fungi: Regulation of transporter-encoding gene expression. Front Physiol. 2014, 5, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rispail, N.; Di Pietro, A. The homeodomain transcription factor Ste12: Connecting fungal MAPK signaling to plant pathogenicity. Commun. Integr. Biol. 2010, 3, 327–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilanova, L.; Teixido, N.; Torres, R.; Usall, J.; Vinas, I.; Sánchez-Torres, P. Relevance of the transcription factor PdSte12 in Penicillium digitatum conidiation and virulence during citrus fruit infection. Int. J. Food Microbiol. 2016, 235, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.C.; Yu, P.L.; Chen, L.H.; Tsai, H.C.; Chung, K.R. A Major Facilitator Superfamily Transporter Regulated by the Stress-Responsive Transcription Factor Yap1 Is Required for Resistance to Fungicides, Xenobiotics, and Oxidants and Full Virulence in Alternaria alternata. Front Microbiol. 2018, 9, 2229. [Google Scholar] [CrossRef]
- Chung, K.R. Reactive oxygen species in the citrus fungal pathogen Alternaria alternata: The roles of NADPH oxidase. Physiol. Mol. Plant Pathol. 2014, 88, 10–17. [Google Scholar] [CrossRef]
- Turra, D.; Segorbe, D.; Di Pietro, A. Protein kinases in plant-pathogenic fungi: Conserved regulators of infection. Annu. Rev. Phytopathol. 2014, 52, 267–288. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.H.; Mehrabi, R.; Xu, J.R. Mitogen-activated protein kinase pathways and fungal pathogenesis. Eukaryot. Cell. 2007, 6, 1701–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.J.; Sun, X.P.; Wang, M.S.; Gai, Y.P.; Chung, K.R.; Li, H.Y. The citrus postharvest pathogen Penicillium digitatum depends on the PdMpkB kinase for developmental and virulence functions. Int. J. Food Microbiol. 2016, 236, 167–176. [Google Scholar] [CrossRef] [PubMed]
- De Ramón-Carbonell, M.; Sánchez-Torres, P. PdSlt2 Penicillium digitatum mitogen-activated-protein kinase controls sporulation and virulence during citrus fruit infection. Fungal Biol. 2017, 121, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, C.; Zhu, C.; Sun, X.; Ruan, R.; Li, H. Os2 MAP kinase-mediated osmostress tolerance in Penicillium digitatum is associated with its positive regulation on glycerol synthesis and negative regulation on ergosterol synthesis. Microbiol. Res. 2014, 169, 511–521. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Z.K.; Sun, H.H.; Ying, S.H.; Feng, M.G. Characterization of the Hog1 MAPK pathway in the entomopathogenic fungus Beauveria bassiana. Environ. Microbiol. 2017, 19, 1808–1821. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FRAC Code | Fungicide Class | Celular Function Affected | Target Protein | Risk Resistance Development |
|---|---|---|---|---|
| 1 | Methyl benzimidazoles (MBCs) | Cytoskeleton | β-tubulin | High |
| 3 | Demethylation inhibitors (DMIs) | Membrane biosynthesis | Sterol 14α-demethylase (CYP51) | Medium |
| 11 | Quinone outside inhibitors (QoIs) | Respiration | Mitochondrial cytochrome b | High |
| 7 | Succinate dehydrogenase inhibitors (SDHIs) | Respiration | Succinate dehydrogenase | Medium to High |
| 12 | Phenylpyrroles (PPs) | Altered target site (protein kinase involved in osmoregulation) | Protein kinase | Low to medium |
| 9 | Anilino-pyrimidines (APs) | Altered target site (protein kinase involved in osmoregulation) | Protein kinase | Medium |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Torres, P. Molecular Mechanisms Underlying Fungicide Resistance in Citrus Postharvest Green Mold. J. Fungi 2021, 7, 783. https://doi.org/10.3390/jof7090783
Sánchez-Torres P. Molecular Mechanisms Underlying Fungicide Resistance in Citrus Postharvest Green Mold. Journal of Fungi. 2021; 7(9):783. https://doi.org/10.3390/jof7090783
Chicago/Turabian StyleSánchez-Torres, Paloma. 2021. "Molecular Mechanisms Underlying Fungicide Resistance in Citrus Postharvest Green Mold" Journal of Fungi 7, no. 9: 783. https://doi.org/10.3390/jof7090783
APA StyleSánchez-Torres, P. (2021). Molecular Mechanisms Underlying Fungicide Resistance in Citrus Postharvest Green Mold. Journal of Fungi, 7(9), 783. https://doi.org/10.3390/jof7090783