
Description of Four Novel Species in Pleosporales Associated with Coffee in Yunnan, China

 ,
,  ,
,  ,
,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection, Morphological Study, and Isolation
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Sequence Alignment and Phylogenetic Analyses
3. Results
3.1. Phylogenetic Analyses
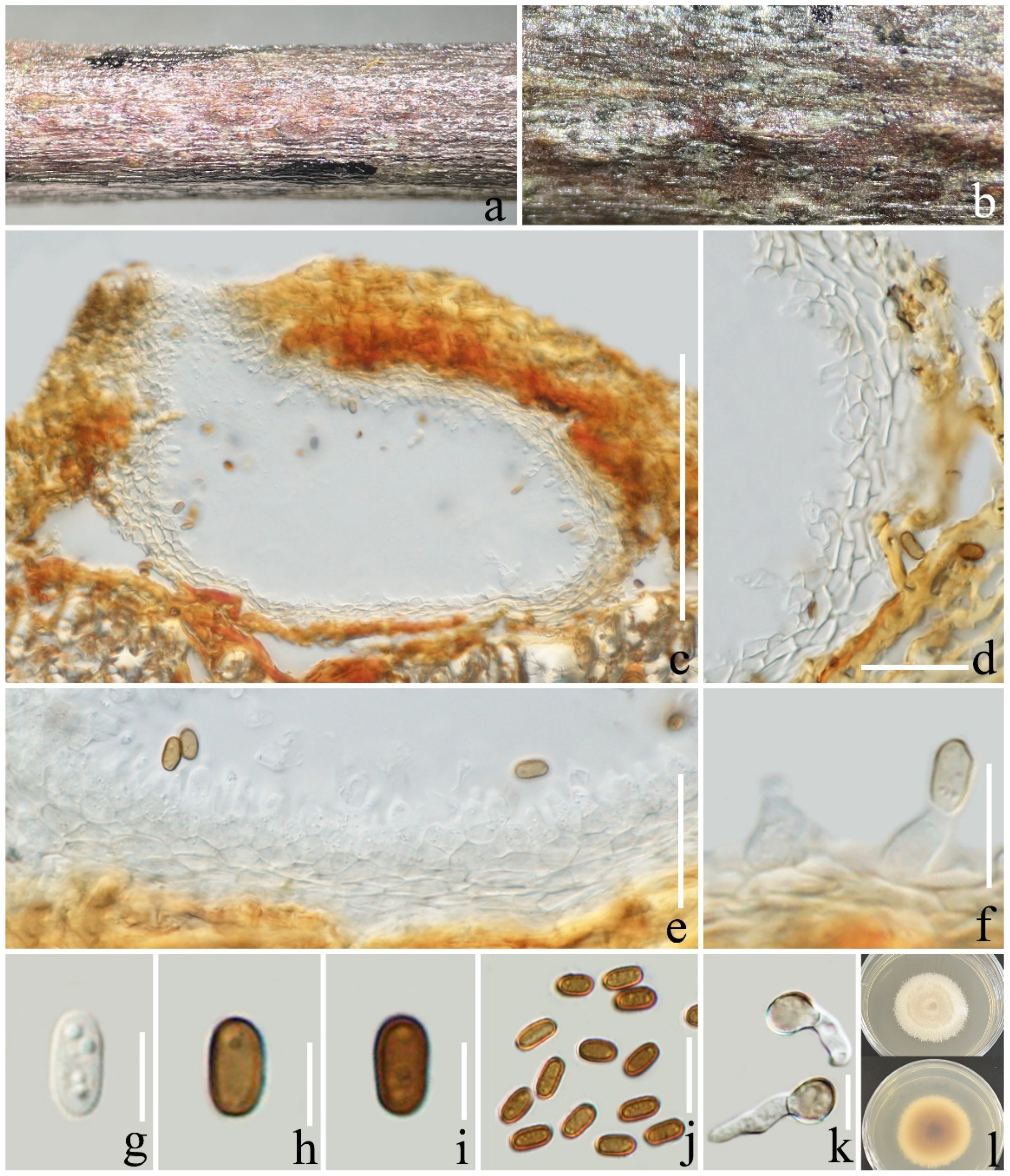
3.2. Taxonomy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coffea L. Available online: https://www.gbif.org/species/2895315 (accessed on 2 March 2022).
- Herrera, J.C.; Lambot, C. The coffee tree—Genetic diversity and origin. In The Craft and Science of Coffee; Elsevier: Tours, France, 2017; pp. 1–16. [Google Scholar]
- Lu, L.; Tibpromma, S.; Karunarathna, S.; Thiyagaraja, V.; Xu, J.; Jayawardena, R.S.; Lumyong, S.; Hyde, K.D. Taxonomic and phylogenic appraisal of a novel species and a new record of Stictidaceae from coffee in Yunnan Province, China. Phytotaxa 2021, 528, 111–124. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, X.; Li, R.; Wang, X.; Cheng, J.; Yang, Q.; Kong, H. AHP-GIS and MaxEnt for delineation of potential distribution of Arabica coffee plantation under future climate in Yunnan, China. Ecol. Indic. 2021, 132, 108339. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, X.; Wang, X.; Gao, Y.; Yang, Q. Evaluation of coffee ecological adaptability using Fuzzy, AHP, and GIS in Yunnan Province, China. Arab. J. Geosci. 2021, 14, 1366. [Google Scholar] [CrossRef]
- Liu, Y.Z.; Jiao, D. Influence of free trade zone construction on coffee industry in Yunnan, China. World Agric. 2017, 3, 181–184. [Google Scholar]
- Coffee in China. Available online: https://oec.world/en/profile/bilateral-product/coffee/reporter/chn (accessed on 2 March 2022).
- Lu, L.; Karunarathna, S.C.; Hyde, K.D.; Bhat, D.J.; Dai, D.-Q.; Jayawardena, R.S.; Tibpromma, S. Crassiparies yunnanensis sp. nov. (Neohendersoniaceae, Pleosporales) from dead twigs of Coffea arabica in China. Phytotaxa 2022, 543, 244–254. [Google Scholar] [CrossRef]
- Waller, J. Control of coffee diseases. In Coffee; Springer: Boston, MA, USA, 1985; pp. 219–229. [Google Scholar]
- Lu, L.; Tibpromma, S.; Karunarathna, S.C.; Jayawardena, R.S.; Lumyong, S.; Xu, J.; Hyde, K.D. Comprehensive Review of Fungi on Coffee. Pathogens 2022, 11, 411. [Google Scholar] [CrossRef]
- Arias, R.M.; Abarca, G.H. Fungal diversity in coffee plantation systems and in a tropical montane cloud forest in Veracruz, Mexico. Agrofor. Syst. 2014, 88, 921–933. [Google Scholar] [CrossRef]
- Pimenta, C.J.; Angélico, C.L.; Chalfoun, S.M. Challengs in coffee quality: Cultural, chemical and microbiological aspects. Ciênc. Agrotecnol. 2018, 42, 337–349. [Google Scholar] [CrossRef]
- Dos Santos, D.G.; Coelho, C.C.d.S.; Ferreira, A.B.R.; Freitas-Silva, O. Brazilian coffee production and the future microbiome and mycotoxin profile considering the climate change Scenario. Microorganisms 2021, 9, 858. [Google Scholar] [CrossRef]
- Botrel, D.A.; Laborde, M.C.F.; Medeiros, F.H.V.d.; Resende, M.L.V.d.; Ribeiro Júnio, P.M.; Pascholati, S.F.; Gusmão, L.F.P. Saprobic fungi as biocontrol agents of halo blight (Pseudomonas syringae pv. garcae) in coffee clones. Coffee Sci. 2018, 13, 283–291. [Google Scholar] [CrossRef]
- Laborde, M.C.F.; Botelho, D.M.d.S.; Rodríguez, G.A.A.; Resende, M.L.V.d.; Queiroz, M.V.d.; Batista, A.D.; Cardoso, P.G.; Pascholati, S.F.; Gusmão, L.F.P.; Martins, S.J. Phialomyces macrosporus reduces Cercospora coffeicola survival on symptomatic coffee leaves. Coffee Sci. 2019, 14, 1–11. [Google Scholar] [CrossRef]
- Zeng, M.W. Study on Identification of Coffee Root Exudates and Allelopathic Effects of Coffee Root Exudates on Coffee Seedlings. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2015. [Google Scholar]
- Wang, K.; Chen, S.L.; Dai, Y.C.; Jia, Z.F.; Li, T.H.; Liu, T.Z.; Phurbu, D.; Mamut, R.; Sun, G.Y.; Bau, T. Overview of China’s nomenclature novelties of fungi in the new century (2000–2020). Mycosystema 2021, 40, 822–833. [Google Scholar]
- Luttrell, E.S. The ascostromatic ascomycetes. Mycologia 1955, 47, 511–532. [Google Scholar] [CrossRef]
- Barr, M. Prodromus to Class Loculoascomycetes; University of Massachusetts: Amherst, MA, USA, 1987; 168p. [Google Scholar]
- Pem, D.; Jeewon, R.; Chethana, K.W.T.; Hongsanan, S.; Doilom, M.; Suwannarach, N.; Hyde, K.D. Species concepts of Dothideomycetes: Classification, phylogenetic inconsistencies and taxonomic standardization. Fungal Divers. 2021, 109, 283–319. [Google Scholar] [CrossRef]
- Yang, E.-F.; Tibpromma, S.; Karunarathna, S.C.; Phookamsak, R.; Xu, J.-C.; Zhao, Z.-X.; Karunanayake, C.; Promputtha, I. Taxonomy and phylogeny of novel and extant taxa in Pleosporales associated with Mangifera indica from Yunnan, China (Series I). J. Fungi 2022, 8, 152. [Google Scholar] [CrossRef]
- Wijayawardene, N.; Hyde, K.; Dai, D.; Sánchez-García, M.; Goto, B.; Saxena, R.; Erdogdu, M.; Selçuk, F.; Rajeshkumar, K.; Aptroot, A. Outline of Fungi and fungus-like taxa-2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Kruys, Å.; Eriksson, O.E.; Wedin, M. Phylogenetic relationships of coprophilous Pleosporales (Dothideomycetes, Ascomycota), and the classification of some bitunicate taxa of unknown position. Mycol. Res. 2006, 110, 527–536. [Google Scholar] [CrossRef]
- Zhang, Y.; Crous, P.W.; Schoch, C.L.; Hyde, K.D. Pleosporales. Fungal Divers. 2012, 53, 1–221. [Google Scholar] [CrossRef] [Green Version]
- Tennakoon, D.S.; Kuo, C.-H.; Maharachchikumbura, S.S.; Thambugala, K.M.; Gentekaki, E.; Phillips, A.J.; Bhat, D.J.; Wanasinghe, D.N.; de Silva, N.I.; Promputtha, I. Taxonomic and phylogenetic contributions to Celtis formosana, Ficus ampelas, F. septica, Macaranga tanarius and Morus australis leaf litter inhabiting microfungi. Fungal Divers. 2021, 108, 1–215. [Google Scholar] [CrossRef]
- Boonmee, S.; D’souza, M.J.; Luo, Z.; Pinruan, U.; Tanaka, K.; Su, H.; Bhat, D.J.; McKenzie, E.H.; Jones, E.; Taylor, J.E. Dictyosporiaceae fam. nov. Fungal Divers. 2016, 80, 457–482. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Mortimer, P.E.; Bezerra, J.D.P. Fungal Systematics and Biogeography. Front. Microbiol. 2022, 12, 827725. [Google Scholar] [CrossRef]
- Munk, A. The system of the Pyrenomycetes. Dansk Bot. Ark. 1953, 15, 1–163. [Google Scholar]
- Ariyawansa, H.A.; Tanaka, K.; Thambugala, K.M.; Phookamsak, R.; Tian, Q.; Camporesi, E.; Hongsanan, S.; Monkai, J.; Wanasinghe, D.N.; Mapook, A. A molecular phylogenetic reappraisal of the Didymosphaeriaceae (=Montagnulaceae). Fungal Divers. 2014, 68, 69–104. [Google Scholar] [CrossRef]
- Zhang, M.D.; Wang, R.; Li, Y.; Hu, X.Q.; Li, M.; Zhang, M.; Duan, C.C. Ecological suitability zoning of Coffea arabica L. in Yunnan Province. Chin. J. Eco-Agric. 2020, 28, 168–178. [Google Scholar]
- Senanayake, I.; Rathnayaka, A.; Marasinghe, D.; Calabon, M.; Gentekaki, E.; Lee, H.; Hurdeal, V.; Pem, D.; Dissanayake, L.; Wijesinghe, S. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.-C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: http://www.indexfungorum.org/Names/IndexFungorumPartnership.htm (accessed on 2 March 2022).
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press, Inc.: Cambridge, MA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Geneious. Available online: https://www.geneious.com/ (accessed on 2 March 2022).
- GenBank. Available online: http://www.ncbi.nlm.nih.gov (accessed on 2 March 2022).
- Mafft. Available online: https://mafft.cbrc.jp/alignment/server/ (accessed on 2 March 2022).
- trimAl. Available online: http://trimal.cgenomics.org (accessed on 2 March 2022).
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dissanayake, A.; Bhunjun, C.; Maharachchikumbura, S.; Liu, J. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2652–2676. [Google Scholar] [CrossRef]
- Miller, M.; Pfeiffer, W. Schwartz Terri. In Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees; Gateway Computing Environments Workshop (GCE): New Orleans, LA, USA, 2010; pp. 1–8. [Google Scholar]
- Nylander, J. MrModeltest v25; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree 1.4.2 Software; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2014. [Google Scholar]
- Figshare. Available online: https://figshare.com/account/home#/data (accessed on 2 September 2022).
- Jiang, H.-B.; Jeewon, R.; Karunarathna, S.C.; Phukhamsakda, C.; Doilom, M.; Kakumyan, P.; Suwannarach, N.; Phookamsak, R.; Lumyong, S. Reappraisal of Immotthia in Dictyosporiaceae, Pleosporales: Introducing Immotthia bambusae sp. nov. and Pseudocyclothyriella clematidis comb. et gen. nov. based on morphology and phylogeny. Front. Microbiol. 2021, 12, 656235. [Google Scholar] [CrossRef]
- Verkley, G.; Dukik, K.; Renfurm, R.; Göker, M.; Stielow, J. Novel genera and species of coniothyrium-like fungi in Montagnulaceae (Ascomycota). Pers.-Mol. Phylogeny Evol. Fungi 2014, 32, 25–51. [Google Scholar] [CrossRef] [Green Version]
- Ariyawansa, H.A.; Maharachchikumbura, S.S.; Karunarathne, S.C.; Chukeatirote, E.; Bahkali, A.H.; Kang, J.; Bhat, D.; Hyde, K.D. Deniquelata barringtoniae gen. et sp. nov., associated with leaf spots of Barringtonia asiatica. Phytotaxa 2013, 105, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; Hyde, K.D.; Mapook, A.; Mortimer, P.E.; Xu, J.; Karunarathna, S.C.; Tibpromma, S. Morphology and phylogenetic analyses reveal Montagnula puerensis sp. nov.(Didymosphaeriaceae, Pleosporales) from southwest China. Phytotaxa 2021, 514, 1–25. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Tsai, I.; Thambugala, K.M.; Chuang, W.-Y.; Lin, S.-R.; Hozzein, W.N.; Cheewangkoon, R. Species diversity of Pleosporalean taxa associated with Camellia sinensis (L.) Kuntze in Taiwan. Sci. Rep. 2020, 10, 12762. [Google Scholar] [CrossRef]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Kudo, H.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.; Hongsanan, S.; Jeewon, R.; Bhat, D.; McKenzie, E.; Jones, E.; Phookamsa, R.; Ariyawansa, H.; Boonmee, S.; Zhao, Q. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Tennakoon, D.; Bhat, D.; Kuo, C.; Hyde, K. Leaf litter saprobic Dictyosporiaceae (Pleosporales, Dothideomycetes): Pseudocoleophoma zingiberacearum sp. nov. from Hedychium coronarium. Kavaka 2019, 53, 1–7. [Google Scholar] [CrossRef]
- Jayasiri, S.; Hyde, K.; Jones, E.; McKenzie, E.; Jeewon, R.; Phillips, A.; Bhat, D.; Wanasinghe, D.; Liu, J.; Lu, Y. Diversity, morphology and molecular phylogeny of Dothideomycetes on decaying wild seed pods and fruits. Mycosphere 2019, 10, 1–186. [Google Scholar] [CrossRef]
- Li, W.-J.; McKenzie, E.H.; Liu, J.-K.J.; Bhat, D.J.; Dai, D.-Q.; Camporesi, E.; Tian, Q.; Maharachchikumbura, S.S.; Luo, Z.-L.; Shang, Q.-J. Taxonomy and phylogeny of hyaline-spored coelomycetes. Fungal Divers. 2020, 100, 279–801. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; McKenzie, E.H.; Phillips, A.J.; Gareth Jones, E.; Jayarama Bhat, D.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Morin, L.; Shivas, R.; Piper, M.; Tan, Y. Austropleospora osteospermi gen. et sp. nov. and its host specificity and distribution on Chrysanthemoides monilifera ssp. rotundata in Australia. Fungal Divers. 2010, 40, 65–74. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.; Dai, D.Q.; Dai, Y.C.; Daranagama, D.A.; Jayawardena, R.S.; Lücking, R. Fungal diversity notes 111–252—Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Dissanayake, L.S.; Wijayawardene, N.N.; Samarakoon, M.C.; Hyde, K.D.; Kang, J.-C. The taxonomy and phylogeny of Austropleospora ochracea sp. nov.(Didymosphaeriaceae) from Guizhou, China. Phytotaxa 2021, 491, 217–229. [Google Scholar] [CrossRef]
- Alidadi, A.; Kowsari, M.; Javan-Nikkhah, M.; Karami, S.; Ariyawansa, H.A.; Jouzani, G.S. Deniquelata quercina sp. nov., a new endophyte species from Persian oak in Iran. Phytotaxa 2019, 405, 187–194. [Google Scholar] [CrossRef]
- Crous, P.; Cowan, D.; Maggs-Kölling, G.; Yilmaz, N.; Thangavel, R.; Wingfield, M.; Noordeloos, M.; Dima, B.; Brandrud, T.; Jansen, G. Fungal planet description sheets: 1182–1283. Pers. Mol. Phylogeny Evol. Fungi 2021, 46, 313–528. [Google Scholar] [CrossRef]
- Devadatha, B.; Sarma, V.V.; Ariyawansa, H.; Jones, E.G. Deniquelata vittalii sp. nov., a novel Indian saprobic marine fungus on Suaeda monoica and two new records of marine fungi from Muthupet mangroves, East coast of India. Mycosphere 2018, 9, 565–582. [Google Scholar] [CrossRef]
- Berlese, A.N. Icones Fungorum, Pyrenomycetes; Kessinger Publishing, LLC: Whitefish, MT, USA, 1896; Volume 2, pp. 1–216. [Google Scholar]
- Tennakoon, D.; Hyde, K.; Wanasinghe, D.; Bahkali, A.; Camporesi, E.; Khan, S.; Phookamsak, R. Taxonomy and phylogenetic appraisal of Montagnula jonesii sp. nov.(Didymosphaeriaceae, Pleosporales). Mycosphere 2016, 7, 1346–1356. [Google Scholar] [CrossRef]
- Barr, M.E. Some Dictyosporous Genera and Species of Pleosporales in North America; The New York Botanical Garden: New York, NY, USA, 1990; Volume 62, pp. 1–92. [Google Scholar]
- Ariyawansa, H.A.; Camporesi, E.; Thambugala, K.M.; Mapook, A.; Kang, J.-C.; Alias, S.A.; Chukeatirote, E.; Thines, M.; Mckenzie, E.H.; Hyde, K.D. Confusion surrounding Didymosphaeria—Phylogenetic and morphological evidence suggest Didymosphaeriaceae is not a distinct family. Phytotaxa 2014, 176, 102–119. [Google Scholar] [CrossRef] [Green Version]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.; Bhat, D.J.; Phillips, A.J.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.; Phookamsak, R.; Wanasinghe, D.; McKenzie, E.; Sarma, V.; Boonmee, S.; Lücking, R.; Bhat, D.; Liu, N. Refined families of Dothideomycetes: Orders and families incertae sets in Dothideomycetidae. Mycosphere J. Fungal Biol. 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.; Jones, E.; Bhat, D.J.; Jeewon, R.; Stadler, M.; Samarakoon, M.C.; Malaithong, M.; Tanunchai, B. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 1–175. [Google Scholar] [CrossRef]
- U.S. National Fungus Collections Fungal Database. Available online: https://nt.ars-grin.gov/fungaldatabases (accessed on 2 March 2022).
- Verkley, G.J.; da Silva, M.; Wicklow, D.T.; Crous, P.W. Paraconiothyrium, a new genus to accommodate the mycoparasite Coniothyrium minitans, anamorphs of Paraphaeosphaeria, and four new species. Stud. Mycol. 2004, 50, 323–335. [Google Scholar]
- Wang, J.; Shao, S.; Liu, C.; Song, Z.; Liu, S.; Wu, S. The genus Paraconiothyrium: Species concepts, biological functions, and secondary metabolites. Crit. Rev. Microbiol. 2021, 47, 781–810. [Google Scholar] [CrossRef]
- Gonçalves, M.F.; Esteves, A.C.; Alves, A. Revealing the hidden diversity of marine fungi in Portugal with the description of two novel species, Neoascochyta fuci sp. nov. and Paraconiothyrium salinum sp. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 5337–5354. [Google Scholar] [CrossRef]
- Crous, P.; Wingfield, M.; Guarro, J.; Hernández-Restrepo, M.; Sutton, D.; Acharya, K.; Barber, P.; Boekhout, T.; Dimitrov, R.; Dueñas, M. Fungal planet description sheets: 320–370. Pers.-Mol. Phylogeny Evol. Fungi 2015, 34, 167–266. [Google Scholar] [CrossRef]
- Wijayawardene, N.; Hyde, K.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Becerra, A.G.; Schnittler, M.; Shchepin, O.; Novozhilov, Y.; Silva-Filho, A. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Wijayawardene, N.; Tibpromma, S.; Hyde, K.; An, Y.; Camporesi, E.; Wang, Y. Additions to Pseudocamarosporium; two new species from Italy. Stud. Fungi 2016, 1, 1–10. [Google Scholar] [CrossRef]
- Khodaei, S.; Arzanlou, M.; Babai-Ahari, A.; Rota-Stabelli, O.; Pertot, I. Phylogeny and evolution of Didymosphaeriaceae (Pleosporales): New Iranian samples and hosts, first divergence estimates, and multiple evidences of species mis-identifications. Phytotaxa 2019, 424, 131–146. [Google Scholar] [CrossRef]
- Hyde, K.D.; Abd-Elsalam, K.; Cai, L. Morphology: Still essential in a molecular world. Mycotaxon 2010, 114, 439–451. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; Nilsson, R.H.; Bhunjun, C.S.; de Farias, A.R.G.; Sun, Y.-R.; Wijesinghe, S.N.; Raza, M.; Bao, D.-F.; Lu, L.; Tibpromma, S. The numbers of fungi: Contributions from traditional taxonomic studies and challenges of metabarcoding. Fungal Divers. 2022, 114, 327–386. [Google Scholar] [CrossRef]
- Liu, J.K.; Hyde, K.D.; Jones, E.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.; McKenzie, E.H.; Phookamsak, R.; Phukhamsakda, C. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Boerema, G.; Loerakker, W. Contributions towards a monograph of Phoma (Coelomycetes)—III. 2. Misapplications of the type species name and the generic synonyms of section Plenodomus (Excluded species). Pers. Mol. Phylogeny Evol. Fungi 1996, 16, 141–189. [Google Scholar]
- Chaiwan, N.; Gomdola, D.; Wang, S.; Monkai, J.; Tibpromma, S.; Doilom, M.; Wanasinghe, D.; Mortimer, P.; Lumyong, S.; Hyde, K. https://gmsmicrofungi.org: An online database providing updated information of microfungi in the Greater Mekong Subregion. Mycosphere 2021, 12, 1409–1422. [Google Scholar] [CrossRef]
- Wang, K.; Cai, L.; Yao, Y. Overview of nomenclature novelties of fungi in the world and China (2020). Biodivers. Sci. 2021, 29, 1064. [Google Scholar] [CrossRef]
- Reverchon, F.; María del Ortega-Larrocea, P.; Pérez-Moreno, J. Saprophytic fungal communities change in diversity and species composition across a volcanic soil chronosequence at Sierra del Chichinautzin, Mexico. Ann. Microbiol. 2010, 60, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Purwanto, B.; Sumadi, S.; Nuraini, A.; Setiawati, M.R. Effect of water content on conidia of Trichoderma spp., indole acetic acid content, electrical conductivity, and pH. Biodivers. J. Biol. Divers. 2022, 23, 2553–2560. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, L.; Karunarathna, S.C.; Dai, D.-Q.; Xiong, Y.-R.; Suwannarach, N.; Stephenson, S.L.; Elgorban, A.M.; Al-Rejaie, S.; Jayawardena, R.S.; Tibpromma, S. Description of Four Novel Species in Pleosporales Associated with Coffee in Yunnan, China. J. Fungi 2022, 8, 1113. https://doi.org/10.3390/jof8101113
Lu L, Karunarathna SC, Dai D-Q, Xiong Y-R, Suwannarach N, Stephenson SL, Elgorban AM, Al-Rejaie S, Jayawardena RS, Tibpromma S. Description of Four Novel Species in Pleosporales Associated with Coffee in Yunnan, China. Journal of Fungi. 2022; 8(10):1113. https://doi.org/10.3390/jof8101113
Chicago/Turabian StyleLu, Li, Samantha C. Karunarathna, Dong-Qin Dai, Yin-Ru Xiong, Nakarin Suwannarach, Steven L. Stephenson, Abdallah M. Elgorban, Salim Al-Rejaie, Ruvishika S. Jayawardena, and Saowaluck Tibpromma. 2022. "Description of Four Novel Species in Pleosporales Associated with Coffee in Yunnan, China" Journal of Fungi 8, no. 10: 1113. https://doi.org/10.3390/jof8101113
APA StyleLu, L., Karunarathna, S. C., Dai, D. -Q., Xiong, Y. -R., Suwannarach, N., Stephenson, S. L., Elgorban, A. M., Al-Rejaie, S., Jayawardena, R. S., & Tibpromma, S. (2022). Description of Four Novel Species in Pleosporales Associated with Coffee in Yunnan, China. Journal of Fungi, 8(10), 1113. https://doi.org/10.3390/jof8101113