
Photodynamic Inactivation Effectively Eradicates Candida auris Biofilm despite Its Interference with the Upregulation of CDR1 and MDR1 Efflux Genes




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization of C. auris Isolates and Formation of 24 h Biofilm
2.2. MB Efficiency and PDI Assay
2.3. CLSM
2.4. Isolation of RNA and Reverse Transcription to cDNA
2.5. Relative Change in Gene Expression Using RT-qPCR
2.6. Statistical Analysis
3. Results
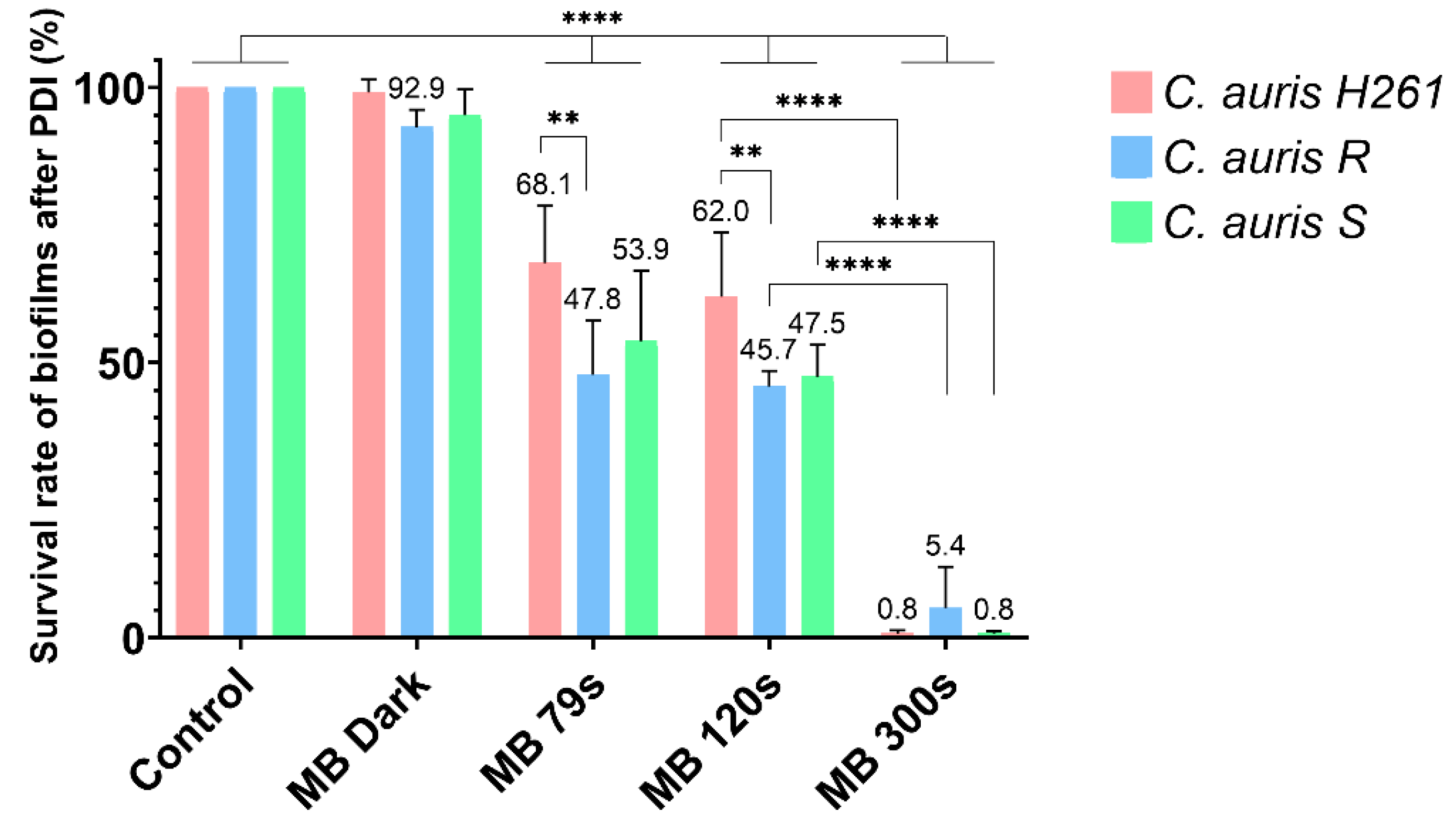
3.1. Efficiency of MB and PDI Assay
3.2. CLSM Confirmation of PDI Effect
3.3. Relative Change in Gene Expression of Efflux Transporters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spivak, E.S.; Hanson, K.E. Candida Auris: An Emerging Fungal Pathogen. J. Clin. Microbiol. 2018, 56, e01588-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, K.R.; Camara, K.; Daniel-Ivad, M.; Trilles, R.; Pimentel-Elardo, S.M.; Fossen, J.L.; Marchillo, K.; Liu, Z.; Singh, S.; Muñoz, J.F.; et al. An Oxindole Efflux Inhibitor Potentiates Azoles and Impairs Virulence in the Fungal Pathogen Candida Auris. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Prakash, A.; Sharma, C.; Kordalewska, M.; Kumar, A.; Sarma, S.; Tarai, B.; Singh, A.; Upadhyaya, G.; Upadhyay, S.; et al. A Multicentre Study of Antifungal Susceptibility Patterns among 350 Candida Auris Isolates (2009–17) in India: Role of the ERG11 and FKS1 Genes in Azole and Echinocandin Resistance. J. Antimicrob. Chemother. 2018, 73, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.C.; Prakash, A.; Meletiadis, J.; Sharma, C.; Chowdhary, A. Comparison of EUCAST and CLSI Reference Microdilution Mics of Eight Antifungal Compounds for Candida Auris and Associated Tentative Epidemiological Cutoff Values. Antimicrob. Agents Chemother. 2017, 61, e00485-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirabet, V.; Salvador, C.; Valentín, A.; Escobedo-Lucea, C.; Navarro, L.; Gimeno, C.; Pemán, J. Risk Assessment of Arterial Allograft Contamination from Tissue Donors Colonized by Candida Auris. J. Hosp. Infect. 2021, 112, 49–53. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Candida Auris: A Rapidly Emerging Cause of Hospital-Acquired Multidrug-Resistant Fungal Infections Globally. PLoS Pathog. 2017, 13, e1006290. [Google Scholar] [CrossRef]
- Cannon, R.D.; Lamping, E.; Holmes, A.R.; Niimi, K.; Baret, P.V.; Keniya, M.V.; Tanabe, K.; Niimi, M.; Goffeau, A.; Monk, B.C. Efflux-Mediated Antifungal Drug Resistance. Clin. Microbiol. Rev. 2009, 22, 291–321. [Google Scholar] [CrossRef] [Green Version]
- Prasad, R.; Rawal, M.K.; Shah, A.H. Candida Efflux ATPases and Antiporters in Clinical Drug Resistance. Adv. Exp. Med. Biol. 2016, 892, 351–376. [Google Scholar] [CrossRef]
- Rao, H.; Choo, S.; Mahalingam, S.R.R.; Adisuri, D.S.; Madhavan, P.; Akim, A.M.; Chong, P.P. Approaches for Mitigating Microbial Biofilm-Related Drug Resistance: A Focus on Micro- and Nanotechnologies. Molecules 2021, 26, 1870. [Google Scholar] [CrossRef]
- Kean, R.; Delaney, C.; Sherry, L.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R.; Williams, C.; Ramage, G. Transcriptome Assembly and Profiling of Candida Auris Reveals Novel Insights into Biofilm-Mediated Resistance. Msphere 2018, 3, e00334-18. [Google Scholar] [CrossRef]
- Benov, L. Photodynamic Therapy: Current Status and Future Directions. Med. Princ. Pract. 2015, 24, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Maliszewska, I.; Wanarska, E.; Thompson, A.C.; Samuel, I.D.W.; Matczyszyn, K. Biogenic Gold Nanoparticles Decrease Methylene Blue Photobleaching and Enhance Antimicrobial Photodynamic Therapy. Molecules 2021, 26, 623. [Google Scholar] [CrossRef]
- Chu, S.; Stochaj, U. Exploring Near-Infrared Absorbing Nanocarriers to Overcome Cancer Drug Resistance. Cancer Drug Resist. 2020, 3, 302–333. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, M.R. Antimicrobial Photodynamic Inactivation: A Bright New Technique to Kill Resistant Microbes. Curr. Opin. Microbiol. 2016, 33, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tardivo, J.P.; del Giglio, A.; de Oliveira, C.S.; Gabrielli, D.S.; Junqueira, H.C.; Tada, D.B.; Severino, D.; de Fátima Turchiello, R.; Baptista, M.S. Methylene Blue in Photodynamic Therapy: From Basic Mechanisms to Clinical Applications. Photodiagnosis Photodyn. Ther. 2005, 2, 175–191. [Google Scholar] [CrossRef]
- del Grande, M.P.; Miyake, A.M.; Nagamine, M.K.; Leite, J.V.P.; da Fonseca, I.I.M.; Massoco, C.d.O.; Dagli, M.L.Z. Methylene Blue and Photodynamic Therapy for Melanomas: Inducing Different Rates of Cell Death (Necrosis and Apoptosis) in B16-F10 Melanoma Cells According to Methylene Blue Concentration and Energy Dose. Photodiagnosis Photodyn. Ther. 2022, 37, 102635. [Google Scholar] [CrossRef]
- Prates, R.A.; Kato, I.T.; Ribeiro, M.S.; Tegos, G.P.; Hamblin, M.R. Influence of Multidrug Efflux Systems on Methylene Blue-Mediated Photodynamic Inactivation of Candida Albicans. J. Antimicrob. Chemother. 2011, 66, 1525. [Google Scholar] [CrossRef]
- Dekkerová, J.; Černáková, L.; Kendra, S.; Borghi, E.; Ottaviano, E.; Willinger, B.; Bujdáková, H. Farnesol Boosts the Antifungal Effect of Fluconazole and Modulates Resistance in Candida Auris through Regulation of the CDR1 and ERG11 Genes. J. Fungi 2022, 8, 783. [Google Scholar] [CrossRef]
- Černáková, L.; Dižová, S.; Bujdáková, H. Employment of Methylene Blue Irradiated with Laser Light Source in Photodynamic Inactivation of Biofilm Formed by Candida Albicans Strain Resistant to Fluconazole. Med. Mycol. 2017, 55, 748–753. [Google Scholar] [CrossRef] [Green Version]
- Rybak, J.M.; Doorley, L.A.; Nishimoto, A.T.; Barker, K.S.; Palmer, G.E.; Rogers, P.D. Abrogation of Triazole Resistance upon Deletion of CDR1 in a Clinical Isolate of Candida Auris. Antimicrob. Agents Chemother. 2019, 63, e00057-19. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Legéňová, K.; Kovalčíková, M.; Černáková, L.; Bujdáková, H. The Contribution of Photodynamic Inactivation vs. Corsodyl Mouthwash to the Control of Streptococcus Mutans Biofilms. Curr. Microbiol. 2020, 77, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.C.; Shen, M.; Huang, Y.J.; Lin, H.C.; Chen, C.T. Photodynamic Inactivation Potentiates the Susceptibility of Antifungal Agents against the Planktonic and Biofilm Cells of Candida Albicans. Int. J. Mol. Sci. 2018, 19, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiench, R.; Skaba, D.; Matys, J.; Grzech-Leśniak, K. Efficacy of Toluidine Blue—Mediated Antimicrobial Photodynamic Therapy on Candida Spp. A Systematic Review. Antibiotics 2021, 10, 349. [Google Scholar] [CrossRef] [PubMed]
- Černáková, L.; Chupáčová, J.; Židlíková, K.; Bujdáková, H. Effectiveness of the Photoactive Dye Methylene Blue versus Caspofungin on the Candida Parapsilosis Biofilm in Vitro and Ex Vivo. Photochem. Photobiol. 2015, 91, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Carmello, J.C.; Dovigo, L.N.; Mima, E.G.; Jorge, J.H.; de Souza Costa, C.A.; Bagnato, V.S.; Pavarina, A.C. In Vivo Evaluation of Photodynamic Inactivation Using Photodithazine® against Candida Albicans. Photochem. Photobiol. Sci. 2015, 14, 1319–1328. [Google Scholar] [CrossRef]
- Bapat, P.S.; Nobile, C.J. Photodynamic Therapy Is Effective Against Candida Auris Biofilms. Front. Cell. Infect. Microbiol. 2021, 11, 819. [Google Scholar] [CrossRef]
- Tan, J.; Liu, Z.; Sun, Y.; Yang, L.; Gao, L. Inhibitory Effects of Photodynamic Inactivation on Planktonic Cells and Biofilms of Candida Auris. Mycopathologia 2019, 184, 525–531. [Google Scholar] [CrossRef]
- Dabrzalska, M.; Janaszewska, A.; Zablocka, M.; Mignani, S.; Majoral, J.P.; Klajnert-Maculewicz, B. Complexing Methylene Blue with Phosphorus Dendrimers to Increase Photodynamic Activity. Molecules 2017, 22, 345. [Google Scholar] [CrossRef] [Green Version]
- Amos-Tautua, B.M.; Songca, S.P.; Oluwafemi, O.S. Application of Porphyrins in Antibacterial Photodynamic Therapy. Molecules 2019, 24, 2456. [Google Scholar] [CrossRef]
- Spring, B.Q.; Rizvi, I.; Xu, N.; Hasan, T. The Role of Photodynamic Therapy in Overcoming Cancer Drug Resistance. Photochem. Photobiol. Sci. 2015, 14, 1476–1491. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štefánek, M.; Černáková, L.; Dekkerová, J.; Bujdáková, H. Photodynamic Inactivation Effectively Eradicates Candida auris Biofilm despite Its Interference with the Upregulation of CDR1 and MDR1 Efflux Genes. J. Fungi 2022, 8, 1137. https://doi.org/10.3390/jof8111137
Štefánek M, Černáková L, Dekkerová J, Bujdáková H. Photodynamic Inactivation Effectively Eradicates Candida auris Biofilm despite Its Interference with the Upregulation of CDR1 and MDR1 Efflux Genes. Journal of Fungi. 2022; 8(11):1137. https://doi.org/10.3390/jof8111137
Chicago/Turabian StyleŠtefánek, Matúš, Lucia Černáková, Jaroslava Dekkerová, and Helena Bujdáková. 2022. "Photodynamic Inactivation Effectively Eradicates Candida auris Biofilm despite Its Interference with the Upregulation of CDR1 and MDR1 Efflux Genes" Journal of Fungi 8, no. 11: 1137. https://doi.org/10.3390/jof8111137
APA StyleŠtefánek, M., Černáková, L., Dekkerová, J., & Bujdáková, H. (2022). Photodynamic Inactivation Effectively Eradicates Candida auris Biofilm despite Its Interference with the Upregulation of CDR1 and MDR1 Efflux Genes. Journal of Fungi, 8(11), 1137. https://doi.org/10.3390/jof8111137