

Additions to The Knowledge of Tubakia (Tubakiaceae, Diaporthales) in China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Morphological Characterization
2.2. DNA Extraction, Amplification and Sequencing
2.3. Phylogeny
2.4. Pathogenicity Test
2.5. Statistical Analyses
3. Results
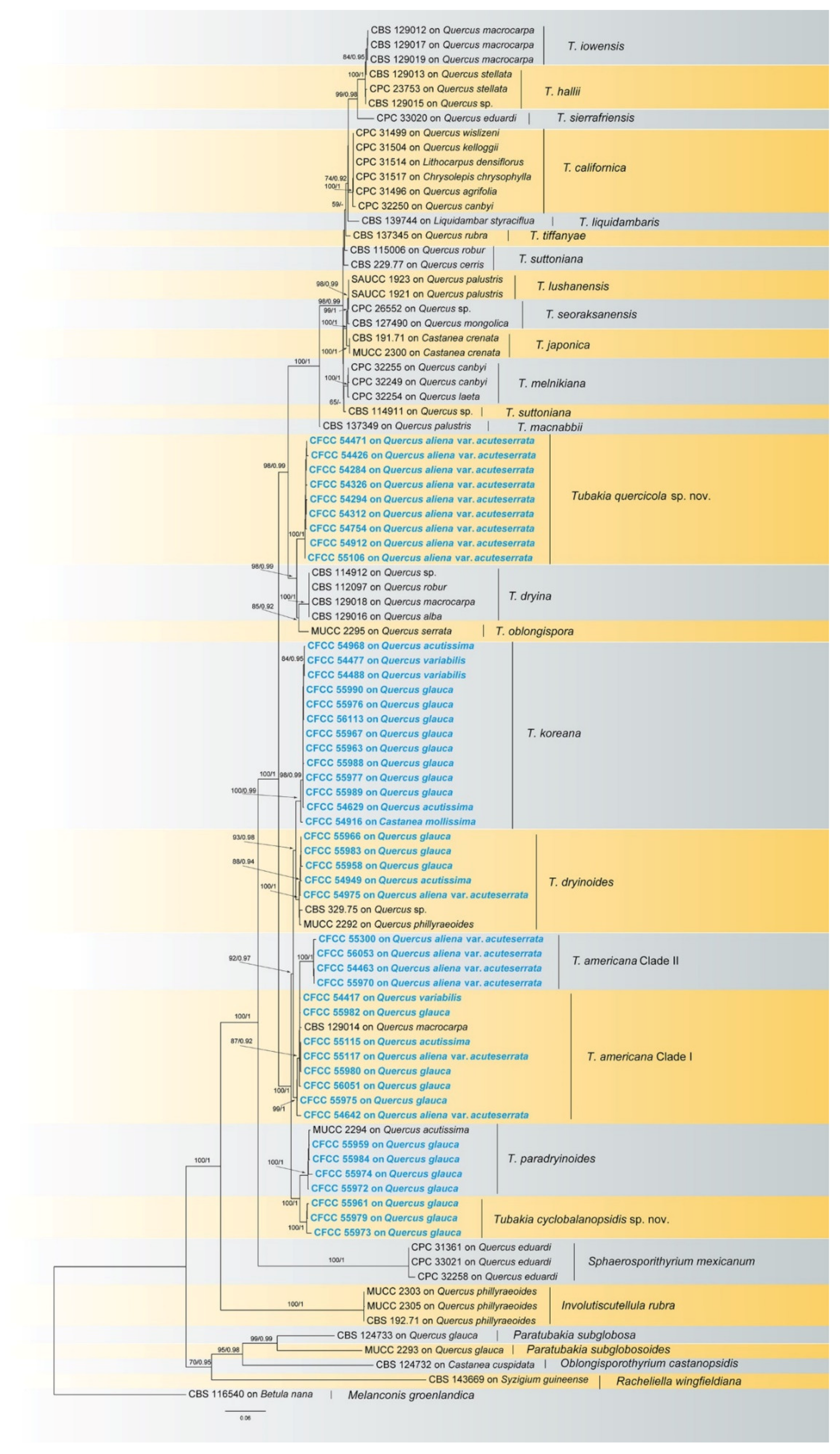
3.1. Phylogenetic Analyses
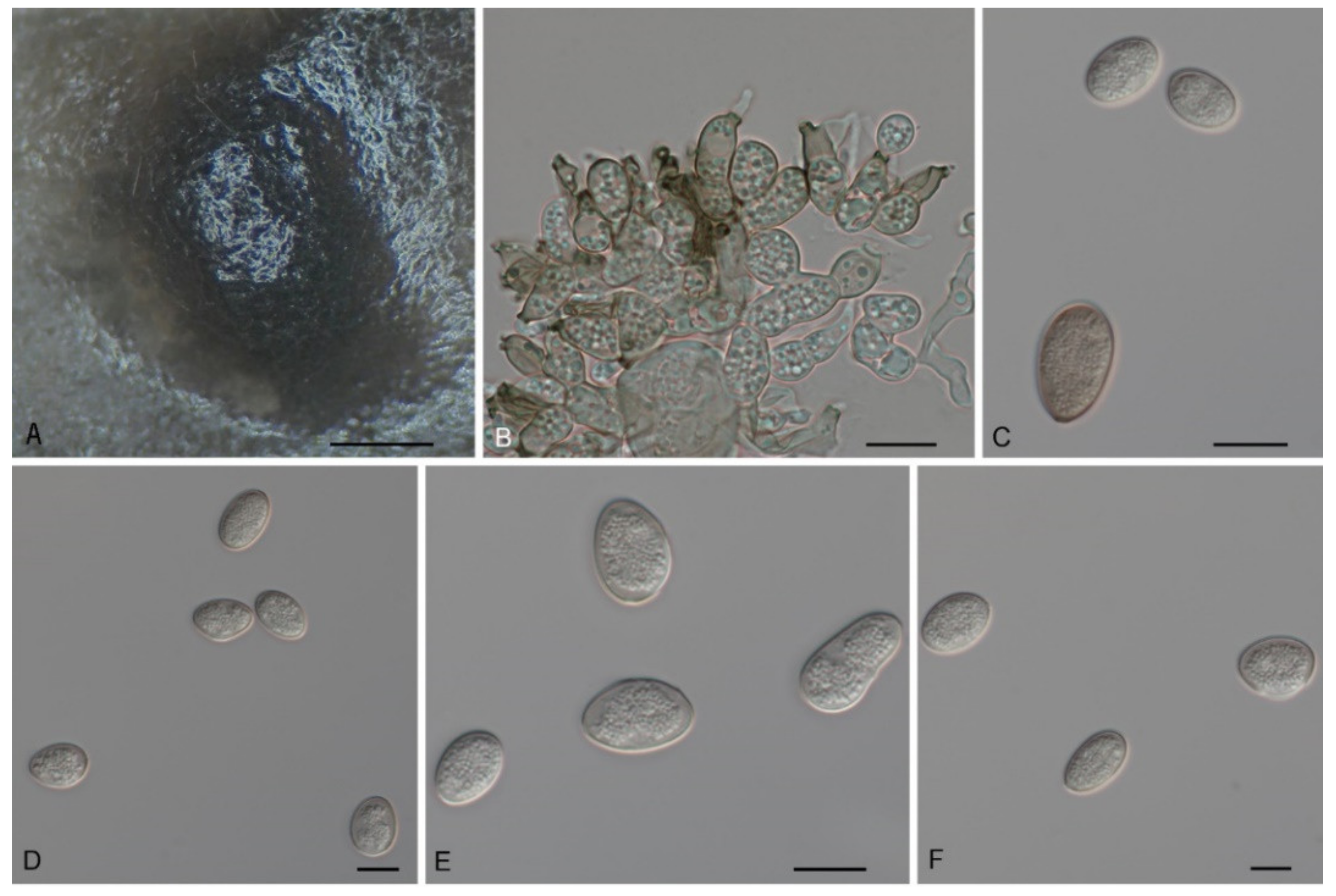
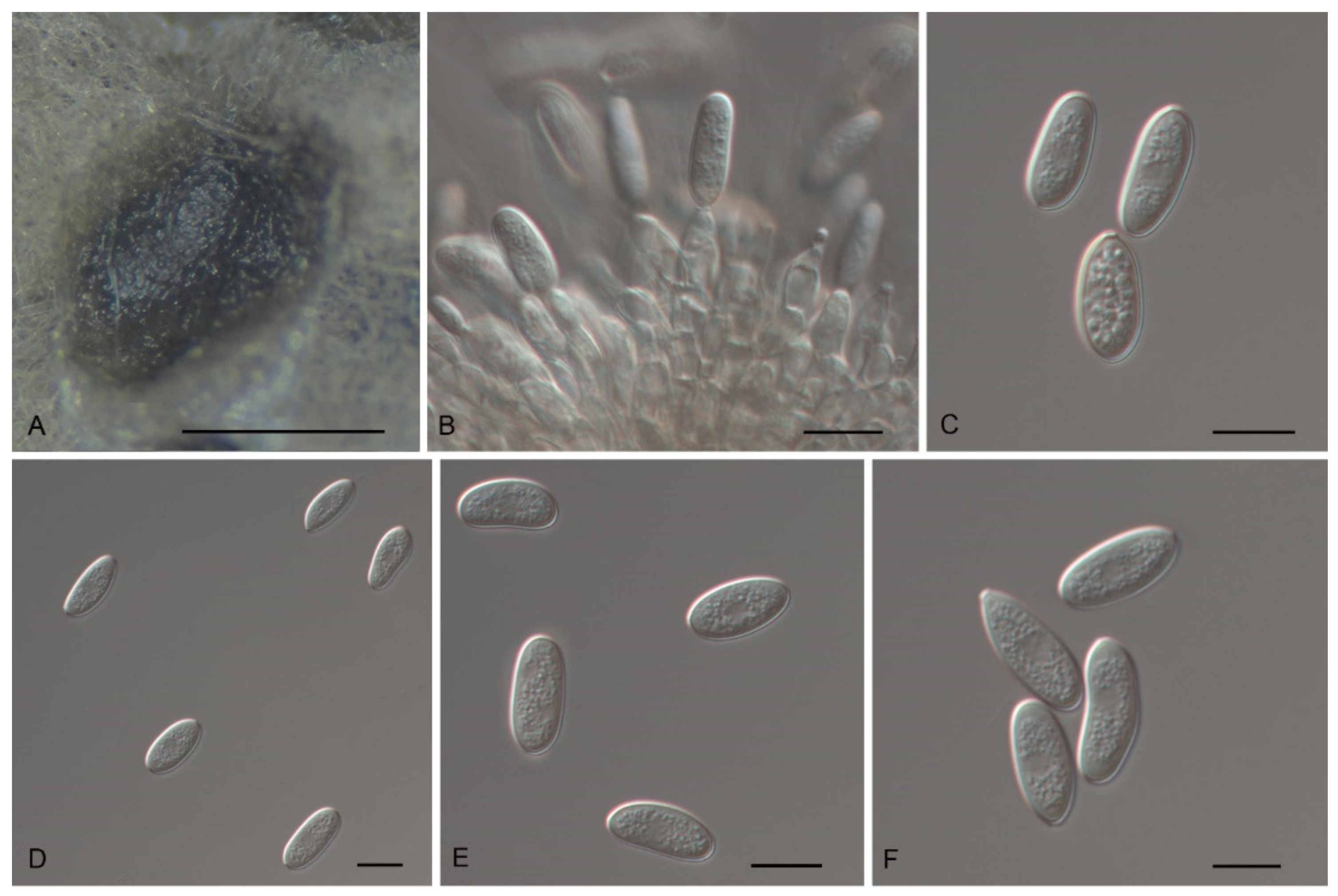
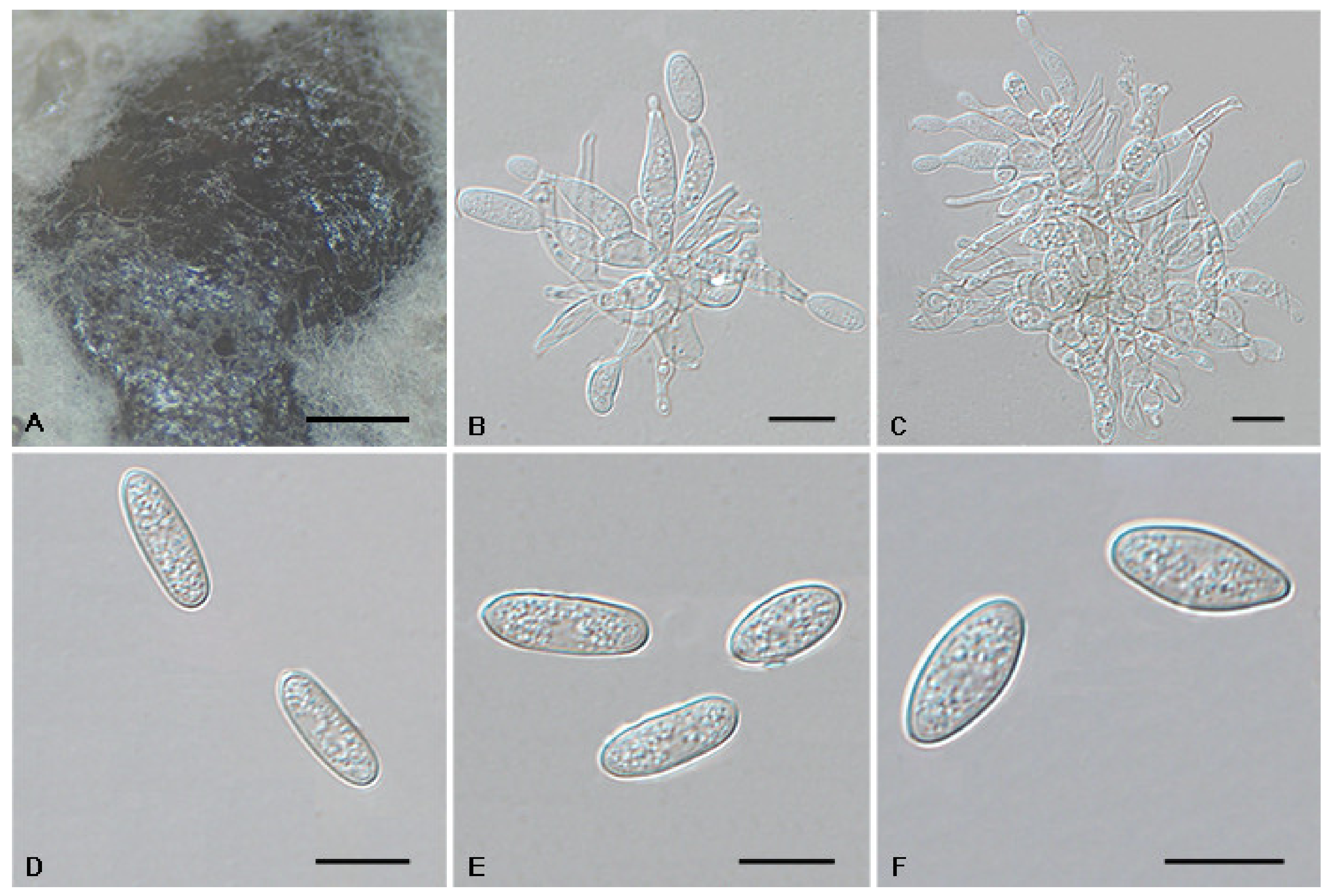
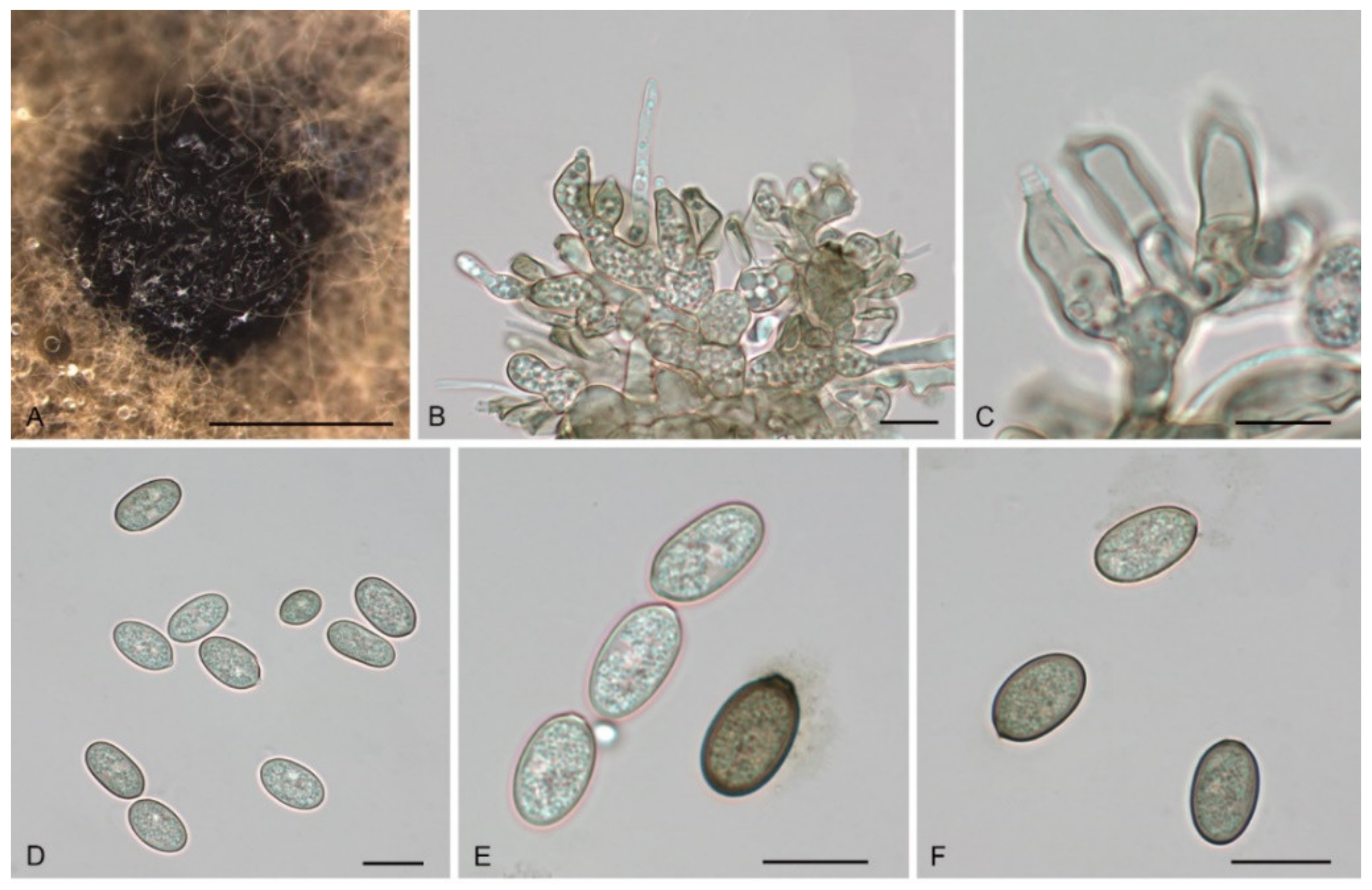
3.2. Morphology
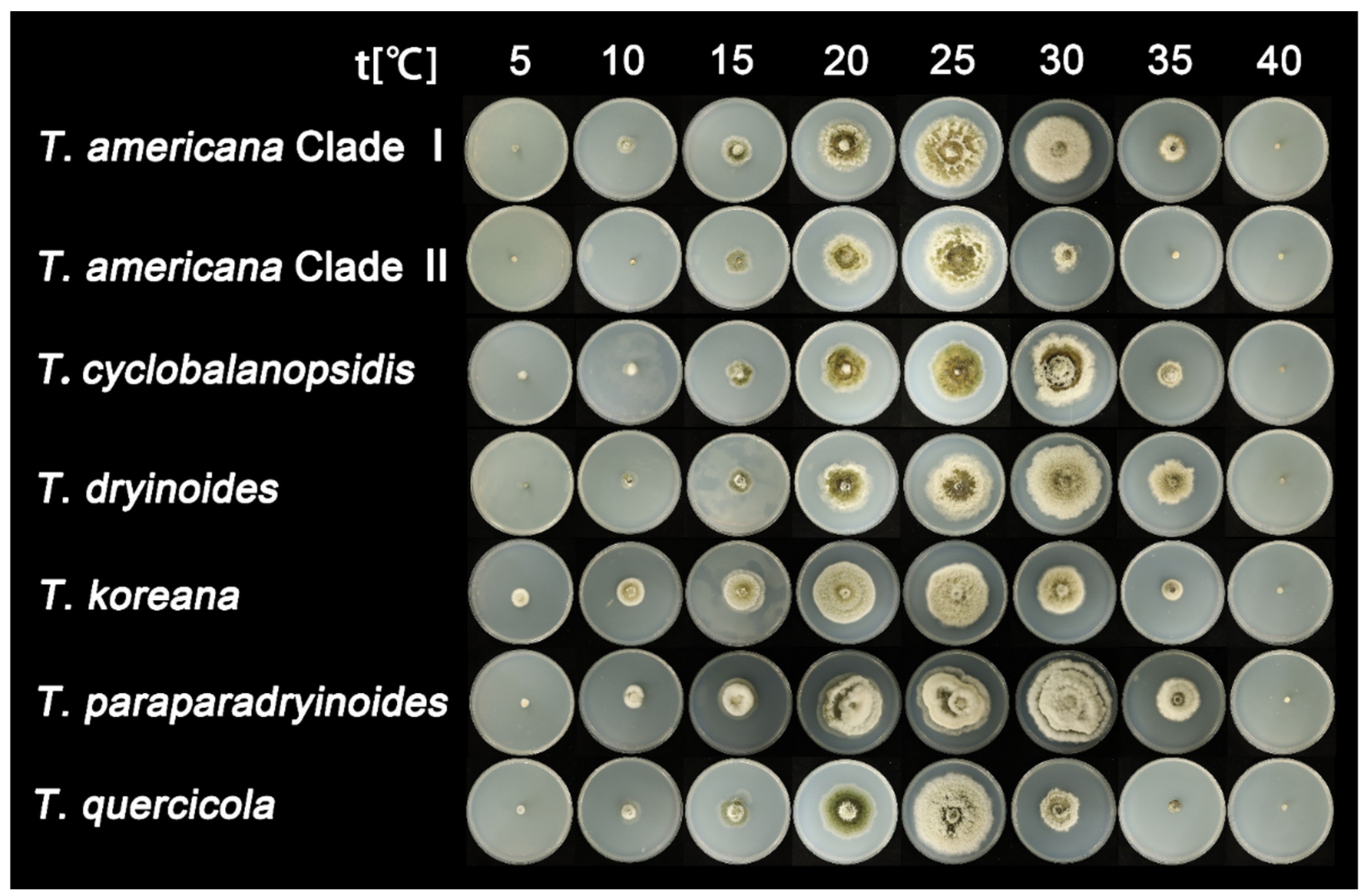
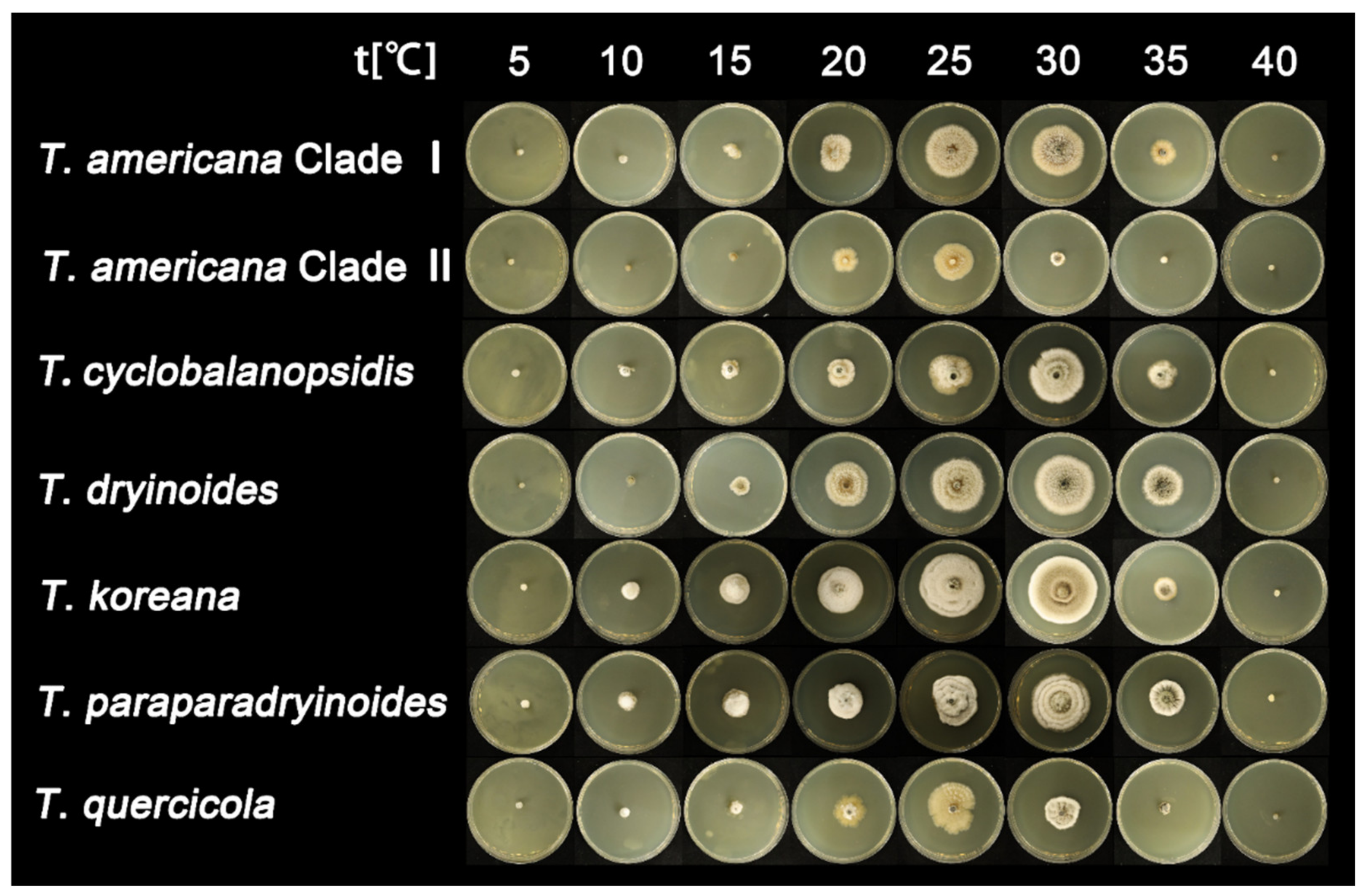
3.3. Statistical Analyses
3.4. Taxonomy
3.5. Pathogenicity Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Strain | Host | GenBank Accession No. | GenBank BLAST Search Results | |||
|---|---|---|---|---|---|---|---|
| Species Identified | Strain | Accession No. | Identities (I), Query Cover (QC) | ||||
| Tubakia americana | CFCC 55115 | Quercus acutissima | OP114595 | T. americana | CBS 129014 | MG591873 | 100.00% (I), 94% (QC) |
| T. americana | CFCC 54642 | Q. aliena var. acuteserrata | OP114596 | T. americana | CBS 129014 | MG591873 | 99.49% (I), 89% (QC) |
| T. americana | CFCC 55117 | Q. aliena var. acuteserrata | OP114597 | T. americana | CBS 129014 | MG591873 | 100.00% (I), 94% (QC) |
| T. americana | CFCC 55980 | Q. glauca | OP114598 | T. americana | CBS 129014 | MG591873 | 100.00% (I), 94% (QC) |
| T. americana | CFCC 55982 | Q. glauca | OP114599 | T. americana | CBS 129014 | MG591873 | 100.00% (I), 94% (QC) |
| T. americana | CFCC 54417 | Q. variabilis | OP114600 | T. americana | CBS 129014 | MG591873 | 100.00% (I), 94% (QC) |
| T. americana | CFCC 55975 | Q. glauca | OP114601 | T. americana | CBS 129014 | MG591873 | 99.66% (I), 95% (QC) |
| T. americana | CFCC 56051 | Q. glauca | OP114602 | T. americana | CBS 129014 | MG591873 | 99.66% (I), 95% (QC) |
| T. americana | CFCC 55970 | Q. aliena var. acuteserrata | OP114603 | T. americana | CBS 129014 | MG591873 | 100.00% (I), 94% (QC) |
| T. americana | CFCC 54463 | Q. aliena var. acuteserrata | OP114604 | T. americana | CBS 129014 | MG591873 | 100.00% (I), 95% (QC) |
| T. americana | CFCC 55300 | Q. aliena var. acuteserrata | OP114605 | T. americana | CBS 129014 | MG591873 | 100.00% (I), 94% (QC) |
| T. americana | CFCC 56053 | Q. aliena var. acuteserrata | OP114606 | T. americana | CBS 129014 | MG591873 | 100.00% (I), 95% (QC) |
| T. cyclobalanopsidis | CFCC 55961 | Q. glauca | OP114638 | T. americana | CBS 129014 | MG591873 | 99.15% (I), 94% (QC) |
| T. cyclobalanopsidis | CFCC 55979 | Q. glauca | OP114639 | T. americana | CBS 129014 | MG591873 | 99.31% (I), 98% (QC) |
| T. cyclobalanopsidis | CFCC 55973 | Q. glauca | OP114640 | T. americana | CBS 129014 | MG591873 | 99.15% (I), 94% (QC) |
| T. dryinoides | CFCC 55958 | Q. glauca | OP114607 | T. dryina | Fs_NI_N09 | KR362909 | 100.00% (I), 98% (QC) |
| T. dryinoides | CFCC 55983 | Q. glauca | OP114608 | T. dryinoides | CBS 329.75 | MG591874 | 100.00% (I), 93% (QC) |
| T. dryinoides | CFCC 55966 | Q. glauca | OP114609 | T. dryina | Fs_NI_N09 | KR362909 | 100.00% (I), 99% (QC) |
| T. dryinoides | CFCC 54949 | Q. acutissima | OP114610 | T. dryina | Fs_NI_N09 | KR362909 | 100.00% (I), 99% (QC) |
| T. dryinoides | CFCC 54975 | Q. aliena var. acuteserrata | OP114611 | T. dryina | Fs_NI_N09 | KR362909 | 99.84% (I), 99% (QC) |
| T. koreana | CFCC 55990 | Q. glauca | OP114616 | T. dryinoides | CBS 329.75 | MG591874 | 100.00% (I), 94% (QC) |
| T. koreana | CFCC 55976 | Q. glauca | OP114617 | T. dryinoides | CBS 329.75 | MG591874 | 100.00% (I), 92% (QC) |
| T. koreana | CFCC 56113 | Q. glauca | OP114618 | T. dryinoides | CBS 329.75 | MG591874 | 100.00% (I), 95% (QC) |
| T. koreana | CFCC 54629 | Q. acutissima | OP114619 | T. dryina | Fs_NI_N09 | KR362909 | 99.84% (I), 99% (QC) |
| T. koreana | CFCC 55967 | Q. glauca | OP114620 | T. dryina | Fs_NI_N09 | KR362909 | 100.00% (I), 98% (QC) |
| T. koreana | CFCC 55977 | Q. glauca | OP114621 | T. dryina | Fs_NI_N09 | KR362909 | 100.00% (I), 98% (QC) |
| T. koreana | CFCC 55963 | Q. glauca | OP114622 | T. dryina | Fs_NI_N09 | KR362909 | 100.00% (I), 98% (QC) |
| T. koreana | CFCC 54968 | Q. acutissima | OP114623 | T. dryinoides | CBS 329.75 | MG591874 | 99.83% (I), 98% (QC) |
| T. koreana | CFCC 54488 | Q. variabilis | OP114624 | T. dryinoides | CBS 329.75 | MG591874 | 99.83% (I), 91% (QC) |
| T. koreana | CFCC 54477 | Q. variabilis | OP114625 | T. dryina | Fs_NI_N09 | KR362909 | 99.83% (I), 99% (QC) |
| T. koreana | CFCC 55989 | Q. glauca | OP114626 | T. dryinoides | CBS 329.75 | MG591874 | 100.00% (I), 95% (QC) |
| T. koreana | CFCC 55988 | Q. glauca | OP114627 | T. dryina | Fs_NI_N09 | KR362909 | 100.00% (I), 99% (QC) |
| T. koreana | CFCC 54916 | C. mollissima | OP114628 | T. dryina | Fs_NI_N09 | KR362909 | 100.00% (I), 98% (QC) |
| T. paradryinoides | CFCC 55984 | Q. glauca | OP114612 | T. dryinoides | CBS 329.75 | MG591874 | 100.00% (I), 95% (QC) |
| T. paradryinoides | CFCC 55959 | Q. glauca | OP114613 | T. dryinoides | CBS 329.75 | MG591874 | 100.00% (I), 95% (QC) |
| T. paradryinoides | CFCC 55974 | Q. glauca | OP114614 | T. dryina | Fs_NI_N09 | KR362909 | 100.00% (I), 98% (QC) |
| T. paradryinoides | CFCC 55972 | Q. glauca | OP114615 | T. dryina | Fs_NI_N09 | KR362909 | 100.00% (I), 98% (QC) |
| T. quercicola | CFCC 54426 | Q. aliena var. acuteserrata | OP114629 | T. dryina | CBS 129018 | MG591871 | 98.54% (I), 99% (QC) |
| T. quercicola | CFCC 54471 | Q. aliena var. acuteserrata | OP114630 | T. dryina | CBS 129018 | MG591871 | 98.90% (I), 99% (QC) |
| T. quercicola | CFCC 54284 | Q. aliena var. acuteserrata | OP114631 | T. dryina | 41G | MZ078778 | 99.06% (I), 100% (QC) |
| T. quercicola | CFCC 54326 | Q. aliena var. acuteserrata | OP114632 | T. dryina | CBS 114919 | MG591855 | 98.85% (I), 94% (QC) |
| T. quercicola | CFCC 54312 | Q. aliena var. acuteserrata | OP114633 | T. dryina | CBS 129018 | MG591871 | 98.89% (I), 100% (QC) |
| T. quercicola | CFCC 54754 | Q. aliena var. acuteserrata | OP114634 | T. dryina | CBS 129018 | MG591871 | 98.94% (I), 99% (QC) |
| T. quercicola | CFCC 55106 | Q. aliena var. acuteserrata | OP114635 | T. dryina | 41G | MZ078778 | 99.22% (I), 99% (QC) |
| T. quercicola | CFCC 54912 | Q. aliena var. acuteserrata | OP114636 | T. dryina | CBS 129018 | MG591871 | 99.21 % (I), 100% (QC) |
| T. quercicola | CFCC 54294 | Q. aliena var. acuteserrata | OP114637 | T. dryina | CBS 129018 | MG591871 | 98.81 % (I), 100% (QC) |
References
- Braun, U.; Nakashima, C.; Crous, P.W.; Groenewald, J.Z.; Moreno-Rico, O.; Rooney-Latham, S.; Blomquist, C.L.; Haas, J.; Marmolejo, J. Phylogeny and taxonomy of the genus Tubakia s. lat. Fungal Syst. Evol. 2018, 1, 41–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saccardo, P.A. Notae mycologicae. Series XVI. Ann. Mycol. 1913, 11, 493–511. [Google Scholar]
- Sutton, B.C. Tubakia nom. nov. Trans. Brit. Mycol. Soc. 1973, 60, 164–165. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Crous, P.W.; Groenewald, J.Z.; Maharachchikumbura, S.S.N.; Jeewon, R.; Phillips, A.J.L.; Bhat, J.D.; Perera, R.H.; Li, Q.R.; Li, W.J.; et al. Families of Diaporthales based on morphological and phylogenetic evidence. Stud. Mycol. 2017, 86, 217–296. [Google Scholar] [CrossRef] [PubMed]
- Harrington, T.C.; McNew, D.L. A re-evaluation of Tubakia, including three new species on Quercus and six new combinations. Anton. Leeuw. Int. J. G. 2018, 111, 1003–1022. [Google Scholar] [CrossRef]
- Von Höhnel, F. Neue Fungi imperfecti. 5. Mitteilung. In Mitteilungen des Botanischen Instituts der Technischen Hochschule Wien; Botanischen Instituts der Technischen Hochschule Wien: Vienna, Austria, 1925; Volume 2, pp. 65–73. [Google Scholar]
- Yokoyama, T.; Tubaki, K. Cultural and taxonomical studies on the genus Actinopelte. Res. Commun. Inst. Ferment Osaka. 1971, 5, 43–77. [Google Scholar]
- Yun, H.Y.; Kim, Y.H. Tubakia koreana sp. nov. causing Quercus leaf blight. Mycotaxon 2020, 135, 223–229. [Google Scholar] [CrossRef]
- Zhang, Z.; Mu, T.; Liu, S.; Liu, R.; Zhang, X.; Xia, J. Morphological and phylogenetic analyses reveal a new genus and two new species of Tubakiaceae from China. MycoKeys 2021, 84, 185. [Google Scholar] [CrossRef]
- Boroń, P.; Grad, B. The occurrence of Tubakia dryina in Poland—New hosts and ITS variation. For. Pathol. 2017, 47, e12294. [Google Scholar] [CrossRef]
- Harrington, T.C.; McNew, D.; Yun, H.Y. Bur oak blight, a new disease on Quercus macrocarpa caused by Tubakia iowensis sp. nov. Mycologia 2012, 104, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Harrington, T.C.; McNew, D.L. Distribution and intensification of bur oak blight in Iowa and the Midwest. In Forest Health Monitoring: National Status, Trends, and Analysis 2015; Porter, K.M., Conkling, B.L., Eds.; General Technical Report; Forest Service & Research Station, Southern Research Station: Asheville, NC, USA, 2016; Volume 213, pp. 105–110. [Google Scholar]
- Liu, X.F.; Liu, X.W. Tubakia on the leaves of Quercus, the newly recorded genus in China. J. Fungal Res. 2010, 8, 23–25. [Google Scholar]
- Proffer, T.J. Tubakia leaf spot. Florida Department of Agriculture and Consumer Services, Division of Plant Industry, Gainesville. Plant Pathol. Circular 1990, 337, 2. [Google Scholar]
- Braun, U.; Bien, S.; Hantsch, L.; Heuchert, B. Tubakia chinensis sp. nov. and a key to the species of the genus Tubakia. Schlechtendalia 2014, 28, 23–28. [Google Scholar]
- Yang, Q.; Jiang, N.; Liang, L.Y.; Yang, Y.C.; Tian, C.M. First report of Tubakia americana causing oak seed rot on Quercus variabilis in China. Plant Dis. 2020, 104, 2724. [Google Scholar] [CrossRef]
- Zhang, J.Q.; Zhang, Y.; Zhou, Y.P.; Dou, Z.P.; He, W.; Crous, P.W. First Report of Tubakia seoraksanensis Parasitizing Quercus mongolica in Lesser Khingan Mountains, China. Plant Dis. 2015, 99, 891. [Google Scholar] [CrossRef]
- Chen, M.M. Forest Fungi Phytogeography: Forest Fungi Phytogeography of China, North America, and Siberia and International Quarantine of Tree Pathogens; Pacific Mushroom Research and Education Center: Sacramento, CA, USA, 2003. [Google Scholar]
- Kaneko, R.; Kaneko, S. The effect of bagging branches on levels of endophytic fungal infection in Japanese beech leaves. For. Pathol. 2004, 34, 65–78. [Google Scholar] [CrossRef]
- Matsumura, E.; Morinaga, K.; Fukuda, K. Host Specificity and Seasonal Variation in the Colonization of Tubakia sensu lato Associated with Evergreen Oak Species in Eastern Japan. Microb. Ecol. 2022. [Google Scholar] [CrossRef]
- Yun, H.Y.; Rossman, A.Y. Tubakia seoraksanensis, a new species from Korea. Mycotaxon 2011, 115, 369–373. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microb. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Katoh, K.; Toh, H. Parallelization of the MAFFT multiple sequence alignment program. Bioinformatics 2010, 26, 1899–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees; Institute of Electrical and Electronics Engineers: New Orleans, LA, USA, 2010. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- OriginLab. OriginPro; Version 2018; OriginLab Corporation: Northampton, MA, USA, 2018. [Google Scholar]
- Gonthier, P.; Gennaro, M.; Nicolotti, G. Effects of water stress on the endophytic mycota of Quercus robur. Fungal Divers. 2006, 21, 69–80. [Google Scholar]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fungal Biol. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Park, H.; Eom, A.H. Three unreported endophytic fungi isolated from conifer leaves of Pinus densiflora in Korea. Korean J. Mycol. 2019, 47, 35–42. [Google Scholar]
- Morales-Rodríguez, C.; Bastianelli, G.; Aleandri, M.; Doğmuş-Lehtijärvi, H.T.; Oskay, F.; Vannini, A. Revealing novel interactions between oak and Tubakia species: Evidence of the efficacy of the sentinel arboreta strategy. Biol. Invasions 2021, 23, 3749–3765. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Bian, D.R.; Piao, C.G.; Wang, S.K.; Li, Y. Morphology and phylogeny of Gnomoniopsis (Gnomoniaceae, Diaporthales) from fagaceae leaves in China. J. Fungi 2021, 7, 792. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Voglmayr, H.; Piao, C.G.; Li, Y. Two new species of Diaporthe (Diaporthaceae, Diaporthales) associated with tree cankers in the Netherlands. MycoKeys 2021, 85, 31–56. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Voglmayr, H.; Ma, C.Y.; Xue, H.; Piao, C.G.; Li, Y. A new Arthrinium-like genus of Amphisphaeriales in China. MycoKeys 2022, 92, 27–43. [Google Scholar] [CrossRef]












| Loci | Primers | PCR: Thermal Cycles: (Annealing Temp. in Bold) | Reference |
|---|---|---|---|
| ITS | ITS/ITS4 | (95 °C:30 s, 48 °C:30 s, 72 °C:1 min) × 35 cycles | [23] |
| tef1 | EF1–728F/EF1–986R | (95 °C:15 s, 54 °C:20 s, 72 °C:1 min) × 35 cycles | [24] |
| tub2 | T1(Bt2a)/Bt2b | (95 °C:30 s, 55 °C:30 s, 72 °C:1 min) × 35 cycles | [25,26] |
| Species | Country | Host | Strain | GenBank Accession Number | ||
|---|---|---|---|---|---|---|
| ITS | tef1 | tub2 | ||||
| Involutiscutellula rubra | Japan | Q. phillyraeoides | CBS 192.71 * | MG591899 | MG592086 | MG592180 |
| I. rubra | Japan | Q. phillyraeoides | MUCC2303 | MG591900 | MG592087 | MG592181 |
| I. rubra | Japan | Q. phillyraeoides | MUCC2305 | MG591902 | MG592089 | MG592182 |
| Melanconis groenlandica | Greenland | Betula nana | CBS 116540 * | KU878552 | KU878554 | KU878555 |
| Oblongisporothyrium castanopsidis | Japan | Castanea cuspidata | CBS 124732 * | MG591849 | MG592037 | MG592131 |
| Paratubakia subglobosa | Japan | Q. glauca | CBS 124733 * | MG591913 | MG592102 | MG592194 |
| P. subglobosoides | Japan | Q. glauca | MUCC2293 * | MG591915 | MG592104 | MG592196 |
| Racheliella wingfieldiana | South Africa | Syzigium guineense | CBS 143669 | MG591911 | MG592100 | MG592192 |
| Sphaerosporithyrium mexicanum | Mexico | Q. eduardi | CPC 31361 | MG591894 | MG592081 | MG592175 |
| S. mexicanum | Mexico | Q. eduardi | CPC 32258 | MG591895 | MG592082 | MG592176 |
| S. mexicanum | Mexico | Q. eduardi | CPC 33021 | MG591896 | MG592083 | MG592177 |
| Tubakia americana | USA | Q.macrocarpa | CBS 129014 | MG591873 | MG592058 | MG592152 |
| T. americana | China | Q. acutissima | CFCC 55115 | OP114595 | OP254203 | OP254249 |
| T. americana | China | Q. aliena var. acuteserrata | CFCC 54642 | OP114596 | OP254204 | OP254250 |
| T. americana | China | Q. aliena var. acuteserrata | CFCC 55117 | OP114597 | OP254205 | OP254251 |
| T. americana | China | Q. glauca | CFCC 55980 | OP114598 | OP254206 | OP254252 |
| T. americana | China | Q. glauca | CFCC 55982 | OP114599 | OP254207 | OP254253 |
| T. americana | China | Q. variabilis | CFCC 54417 | OP114600 | OP254208 | OP254254 |
| T. americana | China | Q. glauca | CFCC 55975 | OP114601 | OP254209 | OP254255 |
| T. americana | China | Q. glauca | CFCC 56051 | OP114602 | OP254210 | OP254256 |
| T. americana | China | Q. aliena var. acuteserrata | CFCC 55970 | OP114603 | OP254211 | OP254257 |
| T. americana | China | Q. aliena var. acuteserrata | CFCC 54463 | OP114604 | OP254212 | OP254258 |
| T. americana | China | Q. aliena var. acuteserrata | CFCC 55300 | OP114605 | OP254213 | OP254259 |
| T. americana | China | Q. aliena var. acuteserrata | CFCC 56053 | OP114606 | OP254214 | OP254260 |
| T. californica | USA | Q. agrifolia | CPC 31496 | MG591829 | MG592017 | MG592111 |
| T. californica | USA | Q. wislizeni | CPC 31499 | MG591832 | MG592020 | MG592114 |
| T. californica | USA | Q. kelloggii | CPC 31504 | MG591834 | MG592022 | MG592116 |
| T. californica | USA | Lithocarpus densiflorus | CPC 31514 | MG591843 | MG592031 | MG592125 |
| T. californica | USA | Chrysolepis chrysophylla | CPC 31517 | MG591846 | MG592034 | MG592128 |
| T. californica | Mexico | Q. canbyi | CPC 32250 | MG591847 | MG592035 | MG592129 |
| T. cyclobalanopsidis | China | Q. glauca | CFCC 55961 | OP114638 | OP254246 | OP329289 |
| T. cyclobalanopsidis | China | Q. glauca | CFCC 55979 * | OP114639 | OP254247 | OP329290 |
| T. cyclobalanopsidis | China | Q. glauca | CFCC 55973 | OP114640 | OP254248 | OP329291 |
| T. dryina | Italy | Q. robur | CBS 112097 * | MG591851 | MG592039 | MG592133 |
| T. dryina | Netherlands | Quercus sp. | CBS 114912 | MG591853 | MG592041 | MG592135 |
| T. dryina | USA | Q. alba | CBS 129016 | MG591870 | MG592056 | MG592150 |
| T. dryina | USA | Q. macrocarpa | CBS 129018 | MG591871 | MG592057 | MG592151 |
| T. dryinoides | France | Quercus sp. | CBS 329.75 | MG591874 | MG592059 | MG592153 |
| T. dryinoides | Japan | Q. phillyraeoides | MUCC2292 * | MG591878 | MG592063 | MG592157 |
| T. dryinoides | China | Q. glauca | CFCC 55958 | OP114607 | OP254215 | OP254261 |
| T. dryinoides | China | Q. glauca | CFCC 55983 | OP114608 | OP254216 | OP254262 |
| T. dryinoides | China | Q. glauca | CFCC 55966 | OP114609 | OP254217 | OP254263 |
| T. dryinoides | China | Q. acutissima | CFCC 54949 | OP114610 | OP254218 | OP254264 |
| T. dryinoides | China | Q. aliena var. acuteserrata | CFCC 54975 | OP114611 | OP254219 | OP254265 |
| T. hallii | USA | Q. stellata | CBS 129013 * | MG591880 | MG592065 | MG592159 |
| T. hallii | USA | Q. stellata | CBS 129015 | MG591881 | MG592066 | MG592160 |
| T. hallii | Iran | Quercus sp. | CPC 23753 | MG591884 | MG592069 | MG592163 |
| T. iowensis | USA | Q. macrocarpa | CBS 129012 * | MG591879 | MG592064 | MG592158 |
| T. iowensis | USA | Q. macrocarpa | CBS 129017 | MG591882 | MG592067 | MG592161 |
| T. iowensis | USA | Q. macrocarpa | CBS 129019 | MG591883 | MG592068 | MG592162 |
| T. japonica | Japan | C. crenata | CBS 191.71 | MG591885 | MG592070 | MG592164 |
| T. japonica | Japan | C. crenata | MUCC2300 | MG591887 | MG592074 | MG592168 |
| T. koreana | China | Q. glauca | CFCC 55990 | OP114616 | OP254224 | OP254270 |
| T. koreana | China | Q. glauca | CFCC 55976 | OP114617 | OP254225 | OP254271 |
| T. koreana | China | Q. glauca | CFCC 56113 | OP114618 | OP254226 | OP254272 |
| T. koreana | China | Q. acutissima | CFCC 54629 | OP114619 | OP254227 | OP254273 |
| T. koreana | China | Q. glauca | CFCC 55967 | OP114620 | OP254228 | OP254274 |
| T. koreana | China | Q. glauca | CFCC 55977 | OP114621 | OP254229 | OP254275 |
| T. koreana | China | Q. glauca | CFCC 55963 | OP114622 | OP254230 | OP254276 |
| T. koreana | China | Q. acutissima | CFCC 54968 | OP114623 | OP254231 | OP254277 |
| T. koreana | China | Q. variabilis | CFCC 54488 | OP114624 | OP254232 | OP254278 |
| T. koreana | China | Q. variabilis | CFCC 54477 | OP114625 | OP254233 | OP254279 |
| T. koreana | China | Q. glauca | CFCC 55989 | OP114626 | OP254234 | OP254280 |
| T. koreana | China | Q. glauca | CFCC 55988 | OP114627 | OP254235 | OP254281 |
| T. koreana | China | C. mollissima | CFCC 54916 | OP114628 | OP254236 | OP254282 |
| T. liquidambaris | USA | Liquidambar styraciflua | CBS 139744 | MG605068 | MG603578 | NA |
| T. lushanensis | China | Q. palustris | SAUCC 1921 | MW784677 | MW842262 | MW842265 |
| T. lushanensis | China | Q. palustris | SAUCC 1923 * | MW784678 | MW842261 | MW842264 |
| T. macnabbii | USA | Q. palustris | CBS 137349 * | MG605069 | MG603579 | NA |
| T. melnikiana | Mexico | Q. canbyi | CPC 32249 | MG591889 | MG592076 | MG592170 |
| T. melnikiana | Mexico | Q. laeta | CPC 32254 | MG591892 | MG592079 | MG592173 |
| T. melnikiana | Mexico | Q. canbyi | CPC 32255 * | MG591893 | MG592080 | MG592174 |
| T. oblongispora | Japan | Q. serrata | MUCC2295 * | MG591897 | MG592084 | MG592178 |
| T. paradryinoides | Japan | Q. acutissima | MUCC2294 * | MG591898 | MG592085 | MG592179 |
| T. paradryinoides | China | Q. glauca | CFCC 55984 | OP114612 | OP254220 | OP254266 |
| T. paradryinoides | China | Q. glauca | CFCC 55959 | OP114613 | OP254221 | OP254267 |
| T. paradryinoides | China | Q. glauca | CFCC 55974 | OP114614 | OP254222 | OP254268 |
| T. paradryinoides | China | Q. glauca | CFCC 55972 | OP114615 | OP254223 | OP254269 |
| T. quercicola | China | Q. aliena var. acuteserrata | CFCC 54426 | OP114629 | OP254237 | OP254283 |
| T. quercicola | China | Q. aliena var. acuteserrata | CFCC 54471 | OP114630 | OP254238 | OP254284 |
| T. quercicola | China | Q. aliena var. acuteserrata | CFCC 54284 | OP114631 | OP254239 | OP254285 |
| T. quercicola | China | Q. aliena var. acuteserrata | CFCC 54326 | OP114632 | OP254240 | OP254286 |
| T. quercicola | China | Q. aliena var. acuteserrata | CFCC 54312 | OP114633 | OP254241 | OP254287 |
| T. quercicola | China | Q. aliena var. acuteserrata | CFCC 54754 | OP114634 | OP254242 | OP254288 |
| T. quercicola | China | Q. aliena var. acuteserrata | CFCC 55106 * | OP114635 | OP254243 | OP254289 |
| T. quercicola | China | Q. aliena var. acuteserrata | CFCC 54912 | OP114636 | OP254244 | OP254290 |
| T. quercicola | China | Q. aliena var. acuteserrata | CFCC 54294 | OP114637 | OP254245 | OP254291 |
| T. seoraksanensis | South Korea | Q. mongolica | CBS 127490 * | MG591907 | MG592094 | MG592186 |
| T. seoraksanensis | China | Q. mongolica | CPC 26552 | MG591909 | MG592097 | MG592189 |
| T. sierrafriensis | Mexico | Q. eduardi | CPC 33020 * | MG591910 | MG592099 | MG592191 |
| T. suttoniana | Netherlands | Quercus sp. | CBS 114911 | MG591916 | MG592105 | MG592197 |
| T. suttoniana | Netherlands | Q. robur | CBS 115006 | MG591917 | MG592106 | MG592198 |
| T. suttoniana | New Zealand | Q. cerris | CBS 229.77 | MG591919 | MG592108 | MG592200 |
| T. tiffanyae | USA | Q. rubra | CBS 137345 * | MG605081 | MG603581 | NA |
| Species | Media | Lesion Diameter 1 (cm) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 5 °C | 10 °C | 15 °C | 20 °C | 25 °C | 30 °C | 35 °C | 40 °C | ||
| Tubakia americana clade I | PDA | 0 | 1.7 ± 0.1 | 2.7 ± 0.1 | 4.8 ± 0.2 | 6.5 ± 0.2 | 6 ± 0.3 | 2.7 ± 0.1 | 0 |
| MEA | 0 | 0 | 1.4 ± 0.1 | 3 ± 0.1 | 4.7 ± 0.1 | 4.6 ± 0.2 | 2.2 ± 0.1 | 0 | |
| Tubakia americana clade II | PDA | 0 | 0 | 2.4 ± 0.1 | 4.4 ± 0.2 | 6.8 ± 0.1 | 2.8 ± 0.1 | 0 | 0 |
| MEA | 0 | 0 | 0.7 | 2.2 ± 0.1 | 3.2 ± 0.1 | 1.2 ± 0.1 | 0 | 0 | |
| Tubakia cyclobalanopsidis | PDA | 0.7 | 1.4 ± 0.1 | 2.4 ± 0.1 | 4.6 ± 0.1 | 5.4 ± 0.1 | 5.8 ± 0.2 | 2.5 ± 0.1 | 0 |
| MEA | 0 | 1 | 1.6 ± 0.1 | 2.5 | 3.8 ± 0.1 | 4.9 ± 0.2 | 2.4 | 0 | |
| Tubakia dryinoides | PDA | 0 | 1.6 ± 0.1 | 2.5 ± 0.1 | 4.8 ± 0.1 | 6.1 ± 0.2 | 6.9 ± 0.3 | 4.2 ± 0.1 | 0 |
| MEA | 0 | 0 | 1.6 ± 0.1 | 3.8 ± 0.1 | 5 ± 0.1 | 5.1 ± 0.2 | 3.6 ± 0.1 | 0 | |
| Tubakia koreana | PDA | 1.2 | 2.5 ± 0.1 | 4.1 ± 0.1 | 5.4 ± 0.2 | 5.6 ± 0.1 | 4.5 ± 0.1 | 2 ± 0.1 | 0 |
| MEA | 0 | 1.6 ± 0.1 | 2.6 ± 0.1 | 4.1 ± 0.1 | 5.6 ± 0.2 | 5.8 ± 0.2 | 2.1 | 0 | |
| Tubakia paradryinoides | PDA | 0 | 1.9 ± 0.1 | 3.6 ± 0.1 | 5.8 ± 0.2 | 6.1 ± 0.2 | 7.6 ± 0.2 | 4 ± 0.1 | 0 |
| MEA | 0 | 1.6 ± 0.1 | 2.4 ± 0.1 | 3 ± 0.1 | 4.5 ± 0.1 | 5.1 ± 0.1 | 3.1 ± 0.1 | 0 | |
| Tubakia quercicola | PDA | 0 | 2.2 ± 0.1 | 2.6 ± 0.1 | 5 ± 0.1 | 7.3 ± 0.3 | 3.6 ± 0.1 | 1.1 | 0 |
| MEA | 0 | 0 | 1.4 ± 0.1 | 2.7 ± 0.1 | 4.2 ± 0.1 | 3 ± 0.1 | 1 | 0 | |
| Species | Days | Lesion Diameter (mm) | Incidence Rate (%) | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Castanea mollissima | Quercus acutissima | Q. variabilis | Q. aliena var.acuteserrata | C. mollissima | Q. acutissima | Q. variabilis | Q. aliena var.acuteserrata | |||
| Tubakia americana clade I | 7 | 0–15 | 0–8 | 0–6 | 7–8 | 66.7% | 66.7% | 83.3% | 100% | 79.18% |
| 14 | 0–23 | 0–11 | 5–19 | 2–26 | 66.7% | 66.7% | 83.3% | 100% | 79.18% | |
| Tubakia americana clade II | 7 | 0–5 | 0–2 | 0–2 | 9–11 | 50% | 33.3% | 33.3% | 100% | 54.15% |
| 14 | 0–9 | 0–5 | 0–11 | 22–29 | 83.3% | 83.3% | 83.3% | 100% | 87.48% | |
| Tubakia cyclobalanopsidis | 7 | 3–22 | 0–5 | 0–1 | 6–9 | 100% | 66.7% | 83.3% | 100% | 87.5% |
| 14 | 4–25 | 0–7 | 0–1 | 6–11 | 100% | 66.7% | 83.3% | 100% | 87.5% | |
| Tubakia dryinoides | 7 | 0–11 | 5–8 | 4–12 | 3–6 | 83.3% | 100% | 100% | 100% | 95.83% |
| 14 | 0–23 | 8–15 | 5–2 | 6–16 | 83.3% | 100% | 100% | 100% | 95.83% | |
| Tubakia koreana | 7 | 0.2–15 | 2–18 | 5–10 | 0–10 | 100% | 100% | 100% | 83.3% | 95.83% |
| 14 | 0.5–25 | 5–29 | 1–35 | 8–29 | 100% | 100% | 100% | 100% | 100% | |
| Tubakia paradryinoides | 7 | 0–13 | 6–8 | 12–2 | 0 | 83.3% | 100% | 100% | 0 | 70.83% |
| 14 | 0–27 | 8–23 | 23–49 | 0 | 83.3% | 100% | 100% | 0 | 70.83% | |
| Tubakia quercicola | 7 | 18–50 | 1–9 | 6–2 | 5–16 | 100% | 100% | 100% | 100% | 100% |
| 14 | whole | 5–17 | 6–22 | whole | 100% | 100% | 100% | 100% | 100% | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.-Q.; Jiang, N.; Dou, Z.-P.; Xue, H.; Piao, C.-G.; Li, Y. Additions to The Knowledge of Tubakia (Tubakiaceae, Diaporthales) in China. J. Fungi 2022, 8, 1143. https://doi.org/10.3390/jof8111143
Zhu Y-Q, Jiang N, Dou Z-P, Xue H, Piao C-G, Li Y. Additions to The Knowledge of Tubakia (Tubakiaceae, Diaporthales) in China. Journal of Fungi. 2022; 8(11):1143. https://doi.org/10.3390/jof8111143
Chicago/Turabian StyleZhu, Ya-Quan, Ning Jiang, Zhi-Peng Dou, Han Xue, Chun-Gen Piao, and Yong Li. 2022. "Additions to The Knowledge of Tubakia (Tubakiaceae, Diaporthales) in China" Journal of Fungi 8, no. 11: 1143. https://doi.org/10.3390/jof8111143
APA StyleZhu, Y. -Q., Jiang, N., Dou, Z. -P., Xue, H., Piao, C. -G., & Li, Y. (2022). Additions to The Knowledge of Tubakia (Tubakiaceae, Diaporthales) in China. Journal of Fungi, 8(11), 1143. https://doi.org/10.3390/jof8111143