
Isolation and Characterization of Ruminal Yeast Strain with Probiotic Potential and Its Effects on Growth Performance, Nutrients Digestibility, Rumen Fermentation and Microbiota of Hu Sheep


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolating, Identifying and Characterizing Yeast Isolates
2.2. Animal Trial
2.3. Sample Collection
2.4. Chemical Analysis
2.5. Microbial DNA Extraction, Sequencing and Data Analysis
2.6. Statistical Analysis
3. Results
3.1. Growth Curve of Yeast Isolates
3.2. Animal Performance and Nutrient Digestibility
3.3. Rumen Fermentation Parameters
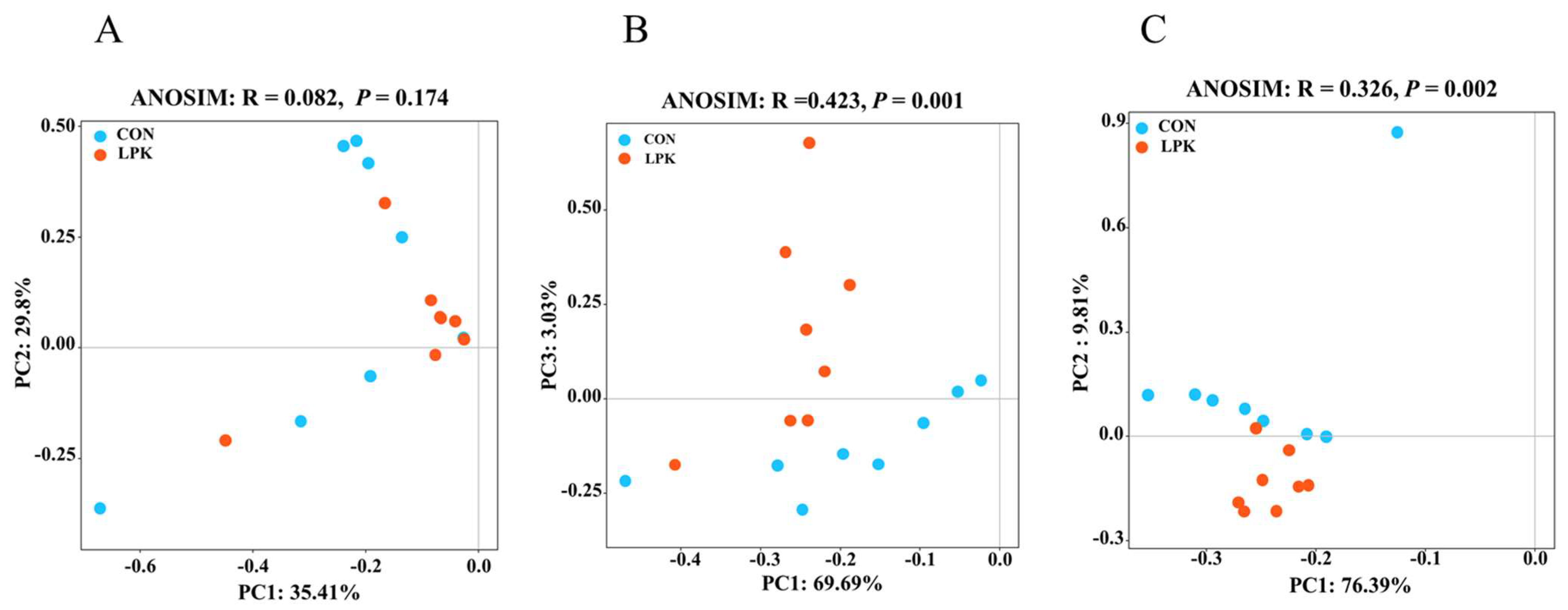
3.4. Rumen Microbial Community Diversity
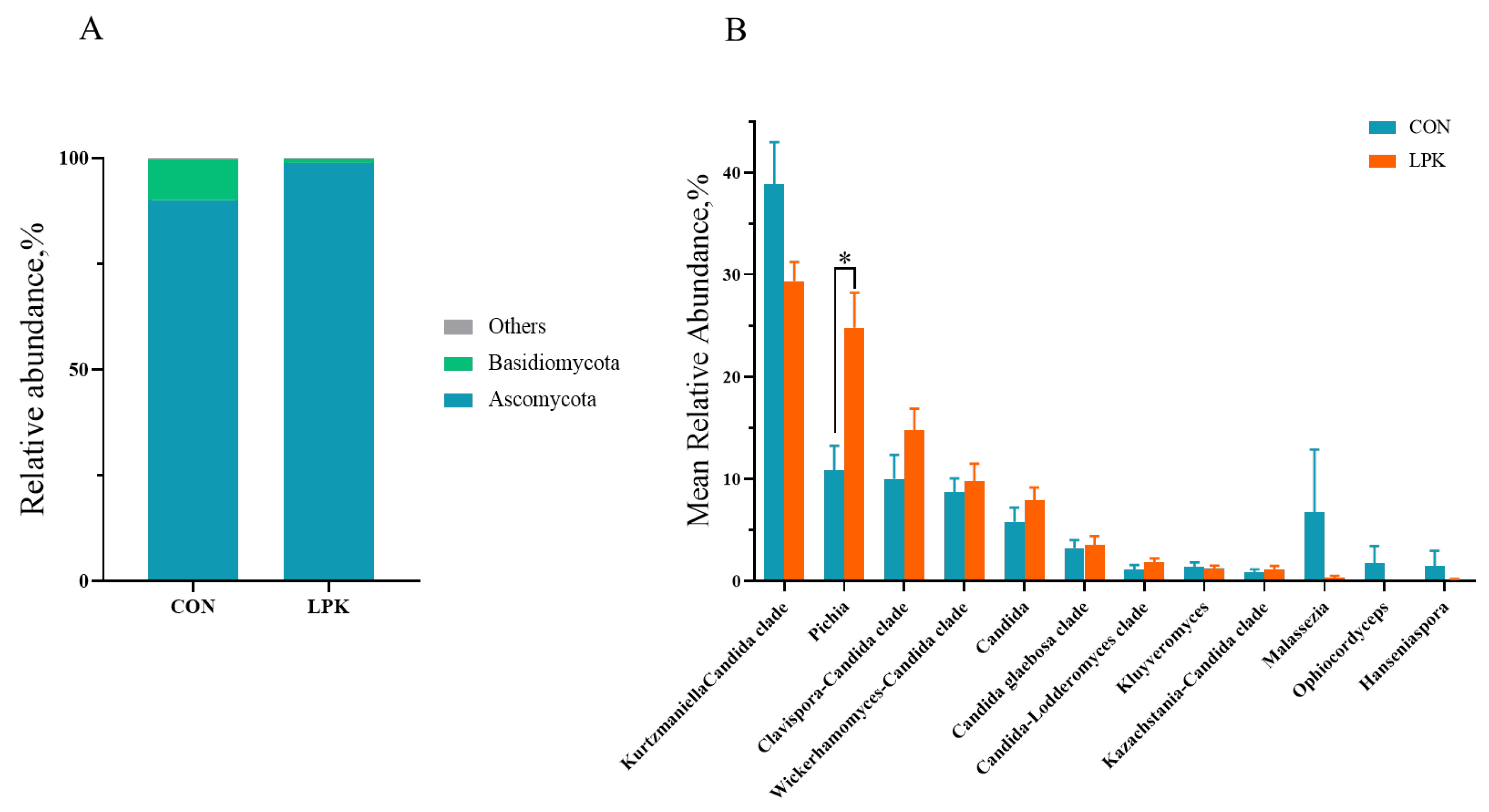
3.5. Rumen Microbiota Composition
3.6. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weimer, P.J. Redundancy, Resilience, and Host Specificity of the Ruminal Microbiota: Implications for Engineering Improved Ruminal Fermentations. Front. Microbiol. 2015, 6, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-L.; Wang, W.-K.; Wu, Q.-C.; Yang, H.-J. The Release and Catabolism of Ferulic Acid in Plant Cell Wall by Rumen Microbes: A Review. Anim. Nutr. 2022, 9, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Fonty, G.; Chaucheyras-Durand, F. Effects and Modes of Action of Live Yeasts in the Rumen. Biologia 2006, 61, 741–750. [Google Scholar] [CrossRef]
- Chaucheyras-Durand, F.; Walker, N.D.; Bach, A. Effects of Active Dry Yeasts on the Rumen Microbial Ecosystem: Past, Present and Future. Anim. Feed Sci. Technol. 2008, 145, 5–26. [Google Scholar] [CrossRef]
- Moya, D.; Ferret, A.; Blanch, M.; Fuentes, M.C.; Fandiño, J.I.; Calsamiglia, S. Effects of Live Yeast (Saccharomyces cerevisiae) and Type of Cereal on Rumen Microbial Fermentation in a Dual Flow Continuous Culture Fermentation System. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1488–1496. [Google Scholar] [CrossRef] [PubMed]
- Pancini, S.; Cooke, R.F.; BrandÃO, A.P.; Dias, N.W.; Timlin, C.L.; Fontes, P.L.P.; Sales, A.F.F.; Wicks, J.C.; Murray, A.; Marques, R.S.; et al. Supplementing a Yeast-Derived Product to Feedlot Cattle Consuming Monensin: Impacts on Performance, Physiological Responses, and Carcass Characteristics. Livest. Sci. 2020, 232, 103907. [Google Scholar] [CrossRef]
- Halfen, J.; Carpinelli, N.; Pino, F.A.B.D.; Chapman, J.D.; Sharman, E.D.; Anderson, J.L.; Osorio, J.S. Effects of Yeast Culture Supplementation on Lactation Performance and Rumen Fermentation Profile and Microbial Abundance in Mid-Lactation Holstein Dairy Cows. J. Dairy Sci. 2021, 104, 11580–11592. [Google Scholar] [CrossRef]
- Thrune, M.; Bach, A.; Ruiz-Moreno, M.; Stern, M.D.; Linn, J.G. Effects of Saccharomyces Cerevisiae on Ruminal PH and Microbial Fermentation in Dairy Cows. Livest. Sci. 2009, 124, 261–265. [Google Scholar] [CrossRef]
- Ishaq, S.L.; AlZahal, O.; Walker, N.; McBride, B. An Investigation into Rumen Fungal and Protozoal Diversity in Three Rumen Fractions, during High-Fiber or Grain-Induced Sub-Acute Ruminal Acidosis Conditions, with or without Active Dry Yeast Supplementation. Front. Microbiol. 2017, 8, 1943. [Google Scholar] [CrossRef] [Green Version]
- Zoumpopoulou, G.; Kazou, M.; Alexandraki, V.; Angelopoulou, A.; Papadimitriou, K.; Pot, B.; Tsakalidou, E. Probiotics and Prebiotics in Animal Health and Food Safety; Gioia, D.D., Biavati, B., Eds.; Springer International Publishing: Berlin, Germany, 2018; pp. 1–34. [Google Scholar]
- Lund, A. Yeasts and Moulds in the Bovine Rumen. Microbiology 2000, 81, 453–462. [Google Scholar] [CrossRef]
- Almeida, P.N.M.D.; Duarte, E.R.; AbrÃO, F.O.; Freitas, C.E.S.; Geraseev, L.C.; Rosa, C.A. Aerobic Fungi in the Rumen Fluid from Dairy Cattle Fed Different Sources of Forage. Rev. Bras. Zootec. 2012, 41, 2336–2342. [Google Scholar] [CrossRef] [Green Version]
- Abrão, F.O.; Freitas, C.E.S.; Duarte, E.R.; Geraseev, L.C.; Barreto, S.M.P.; Medeiros, A.O.; Rosa, C.A. Leveduras No Rúmen De Caprinos E Bovinos De Corte Criados Em Pastagem Tropicais. Arq. Bras. Med. Veterinária E Zootec. 2011, 63, 526–529. [Google Scholar] [CrossRef]
- Fernandes, T.; Carvalho, B.F.; Mantovani, H.C.; Schwan, R.F.; Ávila, C.L.S. Identification and Characterization of Yeasts from Bovine Rumen for Potential Use as Probiotics. J. Appl. Microbiol. 2019, 127, 845–855. [Google Scholar] [CrossRef]
- Suntara, C.; Cherdthong, A.; Wanapat, M.; Uriyapongson, S.; Leelavatcharamas, V.; Sawaengkaew, J.; Chanjula, P.; Foiklang, S. Isolation and Characterization of Yeasts from Rumen Fluids for Potential Use as Additives in Ruminant Feeding. Vet. Sci. 2021, 8, 52. [Google Scholar] [CrossRef]
- Ji, Y.; Dong, X.; Liu, Z.; Wang, W.; Yan, H.; Liu, X. Effects of Bovine Pichia kudriavzevii T7, Candida glabrata B14, and Lactobacillus plantarum Y9 on Milk Production, Quality and Digestive Tract Microbiome in Dairy Cows. Microorganisms 2022, 10, 842. [Google Scholar] [CrossRef]
- Valldecabres, A.; Gilmore, S.P.; Embree, J.J.; Zhelev, I.Z.; Gaffney, J.R.; Marotz, C.A.; Yang, F.; Izzo, A.S.; Embree, M.M.; Lago, A. Effects of Rumen-Native Microbial Feed Supplementation on Milk Yield, Composition, and Feed Efficiency in Lactating Dairy Cows. J. Anim. Sci. 2022, 100, skac275. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Zhang, S.; Liu, X.Z.; Wen, H.A.; Wang, M. A Simple Method of Genomic Dna Extraction Suitable for Analysis of Bulk Fungal Strains. Lett. Appl. Microbiol. 2010, 51, 114–118. [Google Scholar] [CrossRef]
- Yang, H. Feeding Standard of Meat-Producing Sheep and Goats. Postdoctoral Thesis, Chinese Academy of Agricultural Sciences (CAAS), Beijing, China, 2003. [Google Scholar]
- Shen, J.S.; Chai, Z.; Song, L.J.; Liu, J.X.; Wu, Y.M. Insertion Depth of Oral Stomach Tubes May Affect the Fermentation Parameters of Ruminal Fluid Collected in Dairy Cows. J. Dairy Sci. 2012, 95, 5978–5984. [Google Scholar] [CrossRef] [Green Version]
- AOAC International. Official Methods of Analysis of AOAC International, 22nd ed.; Jr, G.W.L., Ed.; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- International Organization for Standardization. Animal Feeding Stuffs: Determination of Ash Insoluble in Hydrochloric Acid. ISO 5985:2002/Cor 1:2005. Available online: https://www.iso.org/standard/42078.html (accessed on 1 July 2022).
- Qin, W.L. Determination of Rumen Volatile Fatty Acids by Means of Gas Chromatography. J. Nanjing Agric. Coll. 1982, 4, 110–116. [Google Scholar]
- Baker, S.B.; Summerson, W.H. The Colorimetric Determination of Lactic Acid in Biological Material. J. Biol. Chem. 1941, 138, 535–554. [Google Scholar] [CrossRef]
- Chaney, A.L.; Marbach, E.P. Modified Reagents for Determination of Urea and Ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.S.; Sharma, O.P.; Dawra, R.K.; Negi, S.S. Simple Determination of Microbial Protein in Rumen Liquor. J. Dairy Sci. 1982, 65, 2170–2173. [Google Scholar] [CrossRef]
- Zakrzewski, M.; Goesmann, A.; Jaenicke, S.; Jünemann, S.; Eikmeyer, F.; Szczepanowski, R.; Al-Soud, W.A.; Sørensen, S.; Pühler, A.; Schlüter, A. Profiling of the Metabolically Active Community from a Production-Scale Biogas Plant by Means of High-Throughput Metatranscriptome Sequencing. J. Biotechnol. 2012, 158, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Logares, R.; Audic, S.; Santini, S.; Pernice, M.C.; de Vargas, C.; Massana, R. Diversity Patterns and Activity of Uncultured Marine Heterotrophic Flagellates Unveiled with Pyrosequencing. ISME J. 2012, 6, 1823–1833. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537. [Google Scholar] [CrossRef] [Green Version]
- García-Ríos, E.; Alonso-Del-Real, J.; Lip, K.Y.F.; Pinheiro, T.; Teixeira, J.; van Gulik, W.; Domingues, L.; Querol, A.; Guillamón, J.M. Genome-Wide Effect of Non-Optimal Temperatures under Anaerobic Conditions on Gene Expression in Saccharomyces cerevisiae. Genomics 2022, 114, 110386. [Google Scholar] [CrossRef]
- Marden, J.P.; Julien, C.; Monteils, V.; Auclair, E.; Moncoulon, R.; Bayourthe, C. How Does Live Yeast Differ from Sodium Bicarbonate to Stabilize Ruminal Ph in High-Yielding Dairy Cows? J. Dairy Sci. 2008, 91, 3528–3535. [Google Scholar] [CrossRef] [Green Version]
- Desnoyers, M.; Giger-Reverdin, S.; Bertin, G.; Duvaux-Ponter, C.; Sauvant, D. Meta-Analysis of the Influence of Saccharomyces cerevisiae Supplementation on Ruminal Parameters and Milk Production of Ruminants. J. Dairy Sci. 2009, 92, 1620–1632. [Google Scholar] [CrossRef] [Green Version]
- Siciliano-Jones, J.; Murphy, M.R. Production of Volatile Fatty Acids in the Rumen and Cecum-Colon of Steers as Affected by Forage: Concentrate and Forage Physical Form. J. Dairy Sci. 1989, 72, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Opsi, F.; Fortina, R.; Tassone, S.; Bodas, R.; López, S. Effects of Inactivated and Live Cells of Saccharomyces cerevisiae on In Vitro Ruminal Fermentation of Diets with Different Forage: Concentrate Ratio. J. Agric. Sci. 2012, 150, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Perdomo, M.C.; Marsola, R.S.; Favoreto, M.G.; Adesogan, A.; Staples, C.R.; Santos, J.E.P. Effects of feeding live yeast at 2 dosages on performance and feeding behavior of dairy cows under heat stress. J. Dairy Sci. 2019, 103, 325–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J.X.; Alugongo, G.M.; Chung, R.; Dong, S.Z.; Li, S.L.; Yoon, I.; Wu, Z.H.; Cao, Z.J. Effects of Saccharomyces cerevisiae Fermentation Products on Dairy Calves: Ruminal Fermentation, Gastrointestinal Morphology, and Microbial Community. J. Dairy Sci. 2016, 99, 5401–5412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strömpl, C.; Tindall, B.J.; Jarvis, G.N.; Lünsdorf, H.; Moore, E.R.B.; Hippe, H. A Re-Evaluation of the Taxonomy of the Genus Anaerovibrio, with the Reclassification of Anaerovibrio glycerini as Anaerosinus glycerini Gen. Nov., Comb. Nov., and Anaerovibrio burkinabensis as Anaeroarcus burkinensis. Gen. Nov., Comb. Nov. Int. J. Syst. Evol. Microbiol. 1999, 49, 1861–1872. [Google Scholar] [CrossRef] [Green Version]
- Hobson, P.N.; Mann, S.O. The Isolation of Glycerol-Fermenting and Lipolytic Bacteria from the Rumen of the Sheep. J. Gen. Microbiol. 1961, 25, 227–240. [Google Scholar] [CrossRef] [Green Version]
- Schauder, R.; Schink, B. Anaerovibrio Glycerini Sp. Nov., an Anaerobic Bacterium Fermenting Glycerol to Propionate, Cell Matter, and Hydrogen. Arch. Microbiol. 1989, 152, 473–478. [Google Scholar] [CrossRef] [Green Version]
- Jennings, D.H. Polyol Metabolism in Fungi. In Advances in Microbial Physiology; Rose, A.H., Tempest, D.W., Eds.; Academic Press: Cambridge, MA, USA, 1985; Volume 25, pp. 149–193. [Google Scholar] [CrossRef]
- Lalucat, J.; Bennasar, A.; Bosch, R.; García-Valdés, E.; Palleroni, N.J. Biology of Pseudomonas stutzeri. Microbiol. Mol. Biol. Rev. 2006, 70, 510–547. [Google Scholar] [CrossRef] [Green Version]
- Priji, P.; Sajith, S.; Unni, K.N.; Anderson, R.C.; Benjamin, S. Pseudomonas sp. BUP6, a Novel Isolate from Malabari Goat Produces an Efficient Rhamnolipid Type Biosurfactant. J. Basic Microbiol. 2017, 57, 21–33. [Google Scholar] [CrossRef]
- Patel, M.; Patel, H.M.; Vohra, N.; Dave, S. Complete Genome Sequencing and Comparative Genome Characterization of the Lignocellulosic Biomass Degrading Bacterium Pseudomonas stutzeri Mp4687 from Cattle Rumen. Biotechnol. Rep. 2020, 28, e00530. [Google Scholar] [CrossRef]
- Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From Microbiology to Diagnostics and Prognostics. J. ISME 2017, 11, 841–852. [Google Scholar] [CrossRef]
- Leahy, S.C.; Higgins, D.G.; Fitzgerald, G.F.; Sinderen, D. Getting Better with Bifidobacteria. J. Appl. Microbiol. 2005, 98, 1303–1315. [Google Scholar] [CrossRef]
- Kelly, W.J.; Cookson, A.L.; Altermann, E.; Lambie, S.C.; Perry, R.; Teh, K.H.; Otter, D.E.; Shapiro, N.; Woyke, T.; Leahy, S.C. Genomic Analysis of Three Bifidobacterium Species Isolated from the Calf Gastrointestinal Tract. Sci. Rep. 2016, 6, 30768. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, M.K.; Karim, S.A. Effect of Yeast Cultures Supplementation on Live Weight Change, Rumen Fermentation, Ciliate Protozoa Population, Microbial Hydrolytic Enzymes Status and Slaughtering Performance of Growing Lamb. Livest. Sci. 2011, 135, 17–25. [Google Scholar] [CrossRef]
- Majewska, M.P.; Miltko, R.; Bełżecki, G.; Kowalik, B. Population of Protozoa and Carbohydrate-Digesting Enzymes in the Rumen of Sheep Fed a Diet Supplemented with Yeast Saccharomyces cerevisiae. Small Rumin. Res. 2021, 205, 106544. [Google Scholar] [CrossRef]
- Akkada, A.; Eadie, J.; Howard, B. The Biochemistry of Rumen Protozoa. 7. the Carbohydrases of Polyplastron multivesiculatum (Dogiel & Fedorowa). Biochem. J. 1963, 89, 268–272. [Google Scholar] [CrossRef]
- Chaucheyras-Durand, F.; Ameilbonne, A.; Bichat, A.; Mosoni, P.; Ossa, F.; Forano, E. Live Yeasts Enhance Fibre Degradation in the Cow Rumen through an Increase in Plant Substrate Colonization by Fibrolytic Bacteria and Fungi. J. Appl. Microbiol. 2016, 120, 560–570. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Content |
|---|---|
| Ingredient (air-dried basis) | |
| Mushroom residue 1 | 10.00 |
| Maize | 40.00 |
| Soybean meal | 10.00 |
| Corn germ meal | 10.00 |
| Barley malt sprouts | 5.50 |
| Barley peel | 10.00 |
| Rice husk | 6.00 |
| Rice bran | 3.50 |
| NaCl | 0.80 |
| Dicalcium phosphate | 0.80 |
| Limestone meal | 1.40 |
| Mineral and vitamin mixture 2 | 2.00 |
| Total | 100.00 |
| Nutrient level (dry matter basis) | - |
| Metabolic energy (ME) 3 (MJ·kg−1) | 11.01 |
| Dry matter (DM), % | 87.42 |
| Crude protein (CP), % | 14.00 |
| Ether extract (EE), % | 3.12 |
| Neutral detergent fiber (NDF), % | 43.16 |
| Acid detergent fiber (ADF), % | 23.55 |
| Crude ash (Ash), % | 9.54 |
| Calcium (Ca), % | 1.30 |
| Phosphorus (P), % | 0.40 |
| Items | CON | LPK | p-Value |
|---|---|---|---|
| Dry matter intake, g/d | 1519.06 ± 21.95 | 1551.13 ± 23.29 | 0.906 |
| Initial body weight, kg | 27.97 ± 0.99 | 28.66 ± 1.08 | 0.292 |
| Final body weight, kg | 34.90 ± 1.01 | 36.23 ± 0.93 | 0.033 |
| Average daily gain, g/d | 241.33 ± 19.31 | 266.83 ± 14.81 | 0.906 |
| Feed efficiency | 6.07 ± 0.45 | 5.81 ± 0.50 | 0.300 |
| Items | CON | LPK | p-Value |
|---|---|---|---|
| Dry matter | 60.62 ± 0.37 | 64.41 ± 1.46 | 0.033 |
| Crude protein | 60.50 ± 1.85 | 64.68 ± 1.16 | 0.121 |
| Ether extract | 58.08 ± 2.65 | 67.56 ± 1.41 | 0.010 |
| Neutral detergent fiber | 44.38 ± 1.26 | 51.44 ± 1.28 | 0.003 |
| Acid detergent fiber | 37.65 ± 1.35 | 45.5 ± 1.63 | 0.005 |
| Items | CON | LPK | p-Value |
|---|---|---|---|
| Rumen pH | 6.30 ± 0.08 | 6.18 ± 0.39 | 0.618 |
| Ammonia, mg/dL | 19.23 ± 1.97 | 21.68 ± 2.27 | 0.466 |
| Microbial crud protein, mg/dL | 43.55 ± 0.04 | 38.69 ± 0.06 | 0.562 |
| Lactate, mmol/L | 0.26 ± 0.02 | 0.26 ± 0.02 | 0.988 |
| Acetate, mmol/L | 40.73 ± 1.67 | 52.08 ± 2.02 | <0.001 |
| Propionate, mmol/L | 24.42 ± 2.15 | 20.94 ± 0.91 | 0.137 |
| Isobutyrate, mmol/L | 0.17 ± 0.05 | 0.29 ± 0.05 | 0.098 |
| Butyrate, mmol/L | 16.27 ± 1.26 | 17.36 ± 1.03 | 0.444 |
| Isovalerate, mmol/L | 0.47 ± 0.08 | 0.47 ± 0.05 | 0.997 |
| Valerate, mmol/L | 1.77 ± 0.20 | 1.56 ± 0.15 | 0.382 |
| Acetate/propionate | 1.76 ± 0.19 | 2.52 ± 0.14 | 0.011 |
| Total volatile fatty acids, mmol/L | 83.84 ± 2.58 | 92.69 ± 1.72 | 0.005 |
| Item | CON | LPK | p-Value |
|---|---|---|---|
| Rumen bacteria | |||
| Observed ASVs 2 | 167.38 ± 17.58 | 230.75 ± 19.12 | 0.029 |
| Coverage | 1.00 ± 0.0002 | 1.00 ± 0.0002 | 0.255 |
| Chao 1 | 170.38 ± 18.50 | 234.25 ± 18.93 | 0.030 |
| Shannon | 3.24 ± 0.26 | 3.80 ± 0.20 | 0.110 |
| Simpson | 0.15 ± 0.04 | 0.09 ± 0.02 | 0.213 |
| Rumen protozoa | |||
| Observed ASVs | 11.00 ± 2.30 | 10.50 ± 1.09 | 0.847 |
| Coverage | 0.98 ± 0.01 | 0.98 ± 0.003 | 0.618 |
| Chao 1 | 13.13 ± 3.01 | 11.38 ± 1.12 | 0.600 |
| Shannon | 1.41 ± 0.30 | 1.52 ± 0.16 | 0.740 |
| Simpson | 0.43 ± 0.11 | 0.34 ± 0.06 | 0.512 |
| Rumen fungi | |||
| Observed ASVs | 24.38 ± 3.18 | 23.38 ± 1.53 | 0.781 |
| Coverage | 0.98 ± 0.01 | 0.98 ± 0.003 | 0.868 |
| Chao 1 | 28.00 ± 4.53 | 29.00 ± 4.74 | 0.881 |
| Shannon | 2.20 ± 0.13 | 2.28 ± 0.08 | 0.597 |
| Simpson | 0.20 ± 0.03 | 0.16 ± 0.01 | 0.148 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, Z.; Jin, W.; Mao, S. Isolation and Characterization of Ruminal Yeast Strain with Probiotic Potential and Its Effects on Growth Performance, Nutrients Digestibility, Rumen Fermentation and Microbiota of Hu Sheep. J. Fungi 2022, 8, 1260. https://doi.org/10.3390/jof8121260
Wang Y, Li Z, Jin W, Mao S. Isolation and Characterization of Ruminal Yeast Strain with Probiotic Potential and Its Effects on Growth Performance, Nutrients Digestibility, Rumen Fermentation and Microbiota of Hu Sheep. Journal of Fungi. 2022; 8(12):1260. https://doi.org/10.3390/jof8121260
Chicago/Turabian StyleWang, Yao, Zihao Li, Wei Jin, and Shengyong Mao. 2022. "Isolation and Characterization of Ruminal Yeast Strain with Probiotic Potential and Its Effects on Growth Performance, Nutrients Digestibility, Rumen Fermentation and Microbiota of Hu Sheep" Journal of Fungi 8, no. 12: 1260. https://doi.org/10.3390/jof8121260
APA StyleWang, Y., Li, Z., Jin, W., & Mao, S. (2022). Isolation and Characterization of Ruminal Yeast Strain with Probiotic Potential and Its Effects on Growth Performance, Nutrients Digestibility, Rumen Fermentation and Microbiota of Hu Sheep. Journal of Fungi, 8(12), 1260. https://doi.org/10.3390/jof8121260