
Trichoderma asperelloides PSU-P1 Induced Expression of Pathogenesis-Related Protein Genes against Gummy Stem Blight of Muskmelon (Cucumis melo) in Field Evaluation




Abstract
:1. Introduction
2. Materials and Methods
2.1. Sources of Trichoderma and Pathogens
2.2. Plant Inoculation
2.3. RNA Extraction and Quantitative RT-PCR
2.4. Protein Extraction and Enzyme Assays
2.5. Scanning Electron Microscopy
2.6. Effect of T. asperelloides PSU-P1 on Plant Growth and Postharvest Quality
2.7. Plant Inoculation and Disease Assessment
2.8. Statistical Analysis
3. Results
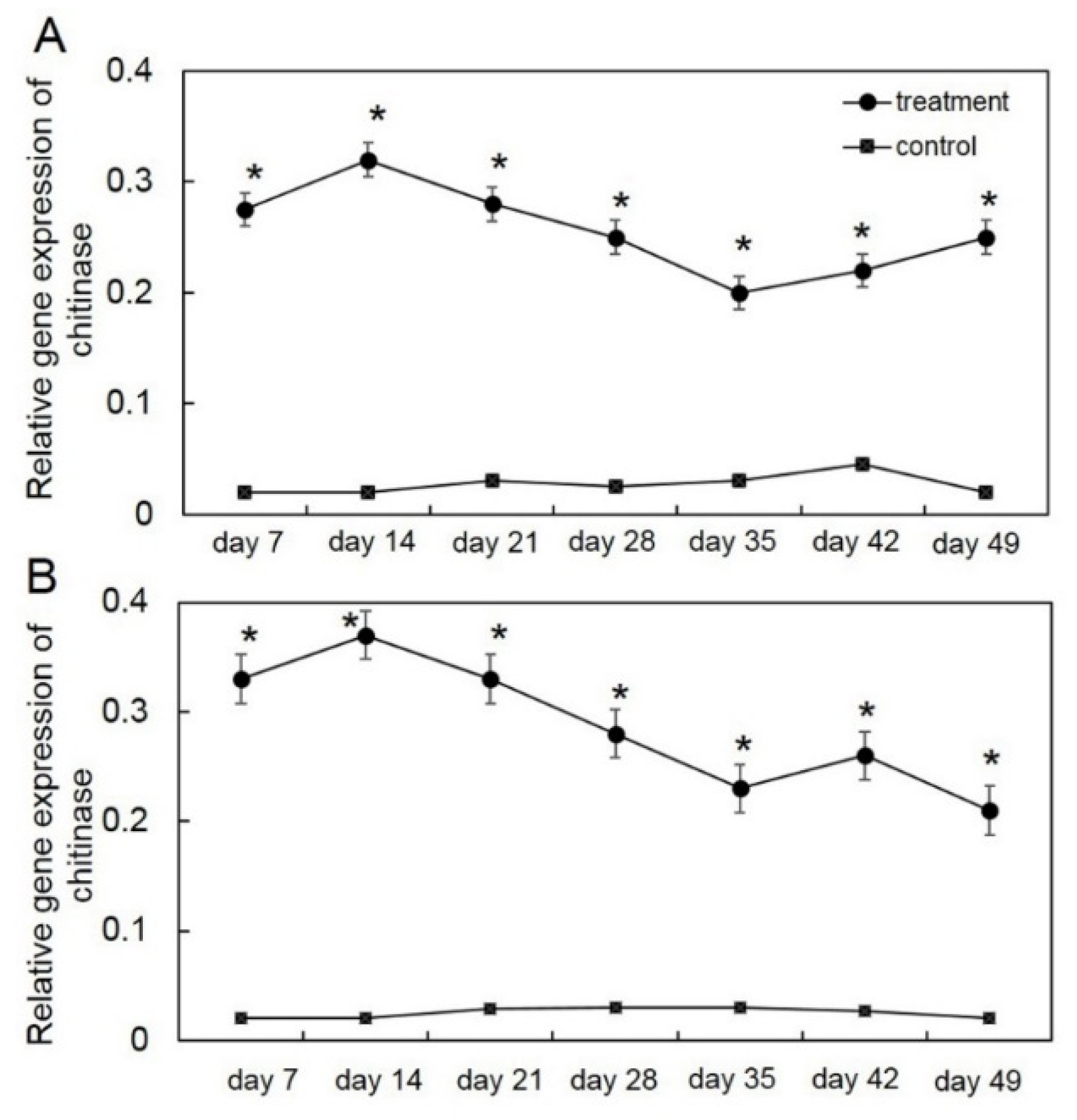
3.1. Effect of Biotic Stress by T. asperelloides PSU-P1 on Gene Expression
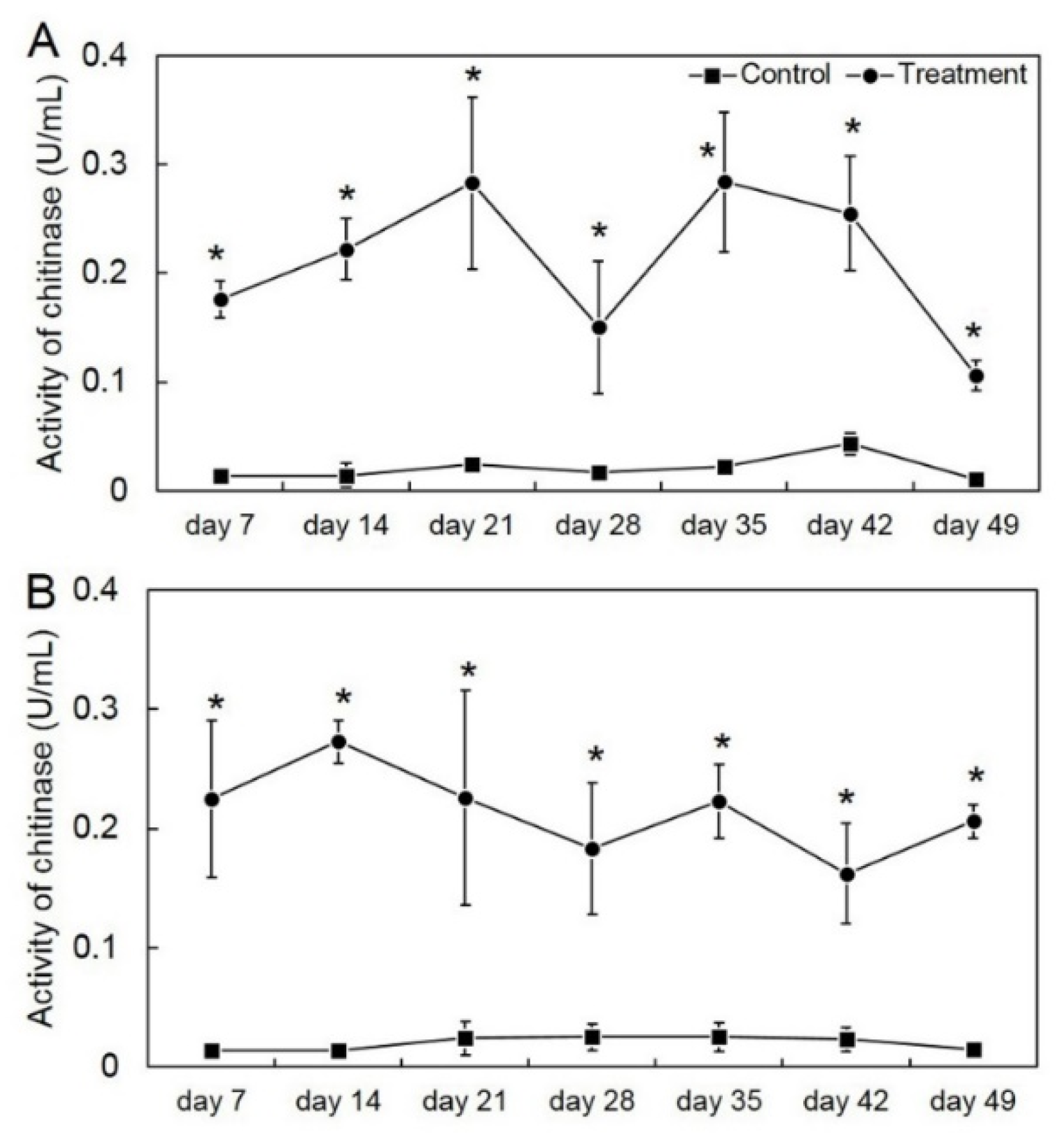
3.2. Application of T. asperelloides PSU-P1 Elevated Enzyme Activity in Muskmelon
3.3. Effect of Crude Metabolites on Fungal Morphology
3.4. Effect of T. asperelloides PSU-P1 on Plant Growth and Postharvest Quality of Muskmelon Fruit
3.5. Trichoderma asperelloides PSU-P1 Reduces Disease Severity in Field Crop
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aveskamp, M.M.; de Gruyter, J.; Woudenberg, J.H.C.; Verkleyand, G.J.M.; Crous, P.W. Highlights of the Didymellaceae: A polyphasic approach to characterize Phoma and related pleosporalean genera. Stud. Mycol. 2010, 65, 281. [Google Scholar] [CrossRef] [PubMed]
- Nuangmek, W.; Aiduang, W.; Suwannarach, N.; Kumla, J.; Lumyong, S. First report of gummy stem blight caused by Stagonosporopsis cucurbitacearum on cantaloupe in Thailand. Can. J. Plant Pathol. 2018, 40, 306–311. [Google Scholar] [CrossRef]
- Nishad, R.; Ahmed, T.; Rahman, V.J.; Kareem, A. Modulation of plant defense system in response to microbial interactions. Front. Microbiol. 2020, 11, e1298. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Liu, N.; Li, C.; Wang, X.; Xu, X.; Chen, W.; Xing, G.; Zheng, W. The early response during the interaction of fungal phytopathogen and host plant. Open Biol. 2017, 7, e170057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baiyee, B.; Ito, S.; Sunapapo, A. Trichoderma asperellum T1 mediated antifungal activity and induced defense response against leaf spot fungi in lettuce (Lactuca sativa L.). Physiol. Mol. Plant Pathol. 2019, 106, 96–101. [Google Scholar] [CrossRef]
- Niderman, T.; Genetet, I.; Bruyère, T.; Gees, R.; Stintzi, A.; Legrand, M.; Fritig, B.; Mösinger, E. Pathogenesis-related PR-1 proteins are antifungal. Isolation and characterization of three 14-kilodalton proteins of tomato and of a basic PR-1 of tobacco with inhibitory activity against Phytophthora infestans. Plant Physiol. 1995, 108, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Souza, T.P.; Dias, R.O.; Silva-Filho, M.C. Defense-related proteins involved in sugarcane responses to biotic stress. Genet. Mol. Biol. 2017, 40 (Suppl. 1), 360–372. [Google Scholar] [CrossRef] [Green Version]
- Chatterton, S.; Punja, Z.K. Chitinase and beta-1,3-glucanase enzyme production by the mycoparasite Clonostachys rosea f. catenulata against fungal plant pathogens. Can. J. Microbiol. 2009, 55, 356–567. [Google Scholar]
- Gupta, P.; Ravi, I.; Sharma, V. Induction of β-1,3-glucanase and chitinase activity in the defense response of Eruca sativa plants against the fungal pathogen Alternaria brassicicola. J. Plant Interact. 2013, 8, 155–161. [Google Scholar] [CrossRef]
- Vázquez-Garcidueñas, S.; Leal-Morales, C.A.; Herrera-Estrella, A. Analysis of the beta-1,3-glucanolytic system of the biocontrol agent Trichoderma harzianum. Appl. Environ. Microbiol. 1998, 64, 1442–1446. [Google Scholar] [CrossRef] [Green Version]
- Goughenour, K.D.; Whalin, J.; Slot, J.C.; Rappleye, C.A. Diversification of fungal chitinases and their functional differentiation in Histoplasma capsulatum. Mol. Biol. Evol. 2021, 38, 1339–1355. [Google Scholar] [CrossRef]
- Anguelova-Merhar, V.S.; VanDer Westhuizen, A.J.; Pretorius, Z.A. β-1,3-Glucanase and chitinase activities and the resistance response of wheat to leaf rust. J. Phytopath. 2008, 149, 381–384. [Google Scholar] [CrossRef]
- Phoka, N.; Suwannarach, N.; Lumyong, S.; Ito, S.-I.; Matsui, K.; Arikit, S.; Sunpapao, A. Role of volatiles from the endophytic fungus Trichoderma asperelloides PSU-P1 in biocontrol potential and in promoting the plant growth of Arabidopsis thaliana. J. Fungi 2020, 6, 341. [Google Scholar] [CrossRef]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar]
- Ruangwong, O.-U.; Pornsuriya, C.; Pitija, K.; Sunpapao, A. Biocontrol mechanisms of Trichoderma koningiopsis PSU3-2 against postharvest anthracnose of chili pepper. J. Fungi 2021, 7, 276. [Google Scholar] [CrossRef]
- Intana, W.; Kheawleng, S.; Sunpapao, A. Trichoderma asperellum T76-14 released volatile organic compounds against postharvest fruit rot in muskmelons (Cucumis melo) caused by Fusarium incarnatum. J. Fungi 2021, 7, 46. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Beltrán-Peña, E.; Herrera-Estrella, A.; López-Bucio, J. Trichoderma-induced plant immunity likely involves both hormonal- and camalexin-dependent mechanisms in Arabidopsis thaliana and confers resistance against necrotrophic fungi Botrytis cinerea. Plant Signal. Behav. 2011, 6, 1554–1563. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Cortés-Penagos, C.; López-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [Green Version]
- Wonglom, P.; Daengsuwan, W.; Ito, S.; Sunpapao, A. Biological control of Sclerotium fruit rot of snake fruit and stem rot of lettuce by Trichoderma sp. T76-12/2 and the mechanism involved. Physiol. Mol. Plant Pathol. 2019, 107, 1–7. [Google Scholar] [CrossRef]
- Ruangwong, O.-U.; Wonglom, P.; Phoka, N.; Suwannarach, N.; Lumyong, S.; Ito, S.-I.; Sunpapao, A. Biological control activity of Trichoderma asperelloides PSU-P1 against gummy stem blight in muskmelon (Cucumis melo). Physiol. Mol. Plant Pathol. 2021, 115, 101663. [Google Scholar] [CrossRef]
- Dumhai, R.; Wanchana, S.; Saensuk, C.; Choowongkomon, K.; Mahatheeranont, S.; Kraithong, T.; Toojinda, T.; Vanavichit, A.; Arikit, S. Discovery of a novel CnAMADH2 allele associated with higher levels of 2-acetyl-1-pyroline (2AP) in yellow dwarf coconut (Cocos nucifera L.). Sci. Hort. 2019, 243, 490–497. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Annl. Biochem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Sunpapao, A.; Chairin, T.; Ito, S. The biocontrol by Streptomyces and Trichoderma of leaf spot disease caused by Curvularia oryzae in oil palm seedlings. Biol. Control 2018, 123, 36–42. [Google Scholar] [CrossRef]
- Singh, S.P.; Keswani, C.; Singh, S.P.; Sansinenea, E.; Hoat, T.X. Trichoderma spp. mediated induction of systemic defense response in brinjal against Sclerotinia sclerotiorum. Curr. Res. Microb. Sci. 2021, 2, 100051. [Google Scholar]
- Hafez, E.E.; Hashem, M.; Balbaa, M.M.; El-Saadani, M.A.; Ahmed, S.A. Induction of new defensin genes in tomato plants via pathogens biocontrol agent interaction. J. Plant Pathol. Microb. 2013, 4, 167. [Google Scholar]
- Porteous-Álvarez, A.J.; Mayo-Prieto, S.; Álvarez-García, S.; Reinoso, B.; Casquero, P.A. Genetic Response of Common Bean to the Inoculation with Indigenous Fusarium Isolates. J. Fungi 2020, 6, 228. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, M.; Radhika, K.; Mathiyazhagan, S.; Bhaskaran, R.; Samiyappan, R.; Velazhahan, R. Induction of phenolics and defense-related enzymes in coconut (Cocos nucifera L.) roots treated with biocontrol agents. Braz. J. Plant Physiol. 2006, 18, 367–377. [Google Scholar] [CrossRef]
- Patel, S.; Saraf, M. Biocontrol efficacy of Trichoderma asperellum MSST against tomato wilting by Fusarium oxysporum f. sp. lycopersici. Arch. Phytopathol. Plant Prot. 2017, 50, 228–238. [Google Scholar] [CrossRef]
- Wonglom, P.; Ito, S.; Sunpapao, A. Volatile organic compounds emitted from endophytic fungus Trichoderma asperellum T1 mediate antifungal activity, defense response and plant growth promoting ability in lettuce (Lactuca sativa L.). Fungal Ecol. 2020, 43, 100867. [Google Scholar] [CrossRef]
- Kumar, M.; Ashraf, S. Role of Trichoderma spp. as a Biocontrol Agent of Fungal Plant Pathogens. In Probiotics and Plant Health; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017. [Google Scholar] [CrossRef]
- Asad, S.A.; Ali, N.; Hameed, A.; Khan, S.A.; Ahmad, R.; Bilal, M.; Shahzad, M.; Tabassum, A. Biocontrol efficacy of different isolates of Trichoderma against soil borne pathogen Rhizoctonia solani. Pol. J. Microbiol. 2014, 63, 95–103. [Google Scholar] [CrossRef]
- Bastakoti, S.; Belbase, S.; Manandhar, S.; Arjyal, C. Trichoderma species as biocontrol agent against soil borne fungal pathogen. Nepal J. Biotechnol. 2017, 5, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Begum, M.F.; Rahman, M.A.; Alam, M.F. Biological control of Alternaria fruit rot of chili by Trichoderma species under field conditions. Mycobiology 2010, 38, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Arzanlou, M.; Khodaei, S.; Narmani, A.; Babai-Ahari, A.; Azar, A.M. Inhibitory effect of Trichoderma isolates on growth of Alternaria alternata, the causal agent of leaf spot disease on sunflower, under laboratory conditions. Arch. Phytopathol. Plant Prot. 2014, 47, 1592–1599. [Google Scholar] [CrossRef]
- Degani, O.; Dor, S. Trichoderma biological control to protect sensitive maize hybrids against late wilt disease in the field. J. Fungi 2021, 7, 315. [Google Scholar] [CrossRef]
- Nuangmek, W.; Aiduang, W.; Kumla, J.; Lumyong, S.; Suwannarach, N. Evaluation of a newly identified, endophytic fungus, Trichoderma phayaoense for plant growth promotion and biological control of gummy stem blight and wilt of muskmelon. Front. Microbiol. 2021, 12, 634772. [Google Scholar] [CrossRef]
- Bononi, L.; Chiaramonte, J.B.; Pansa, C.C.; Moitinho, M.A.; Melo, I.S. Phosphorus solubilizing Trichoderma spp. from Amazon soils improve soybean plant growth. Sci. Rep. 2020, 10, 2858. [Google Scholar] [CrossRef] [Green Version]
- Syam, N.; Sabahannur, S.; Nurdin, A. Effects of Trichoderma and foliar fertilizer on the vegetative growth of black pepper (Piper nigrum L.) seedlings. Int. J. Agron. 2021, 2021, e9953239. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Del Mar Alguacil, M.; Pascual, J.A.; Van Wees, S.C. Phytohormone profiles induced by trichoderma isolates correspond with their biocontrol and plant growth-promoting activity on melon plants. J. Chem. Ecol. 2014, 40, 804–815. [Google Scholar] [CrossRef]
- Aifaa, Y.N.H.; Suhanna, A. Effect of Trichoderma on postharvest quality of Harumanis man. J. Trop. Agric. Food Sci. 2015, 43, 21–28. [Google Scholar]
- Sangeetha, G.; Usharani, S.; Muthukumar, A. Biocontrol with Trichoderma species for the management of post-harvest crown rot of banana. Phytopathol. Mediterr. 2009, 48, 214–225. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession Number | Primer | Sequences (5′→3′) | Product Size (bp) |
|---|---|---|---|---|
| chi | AF241538 | Chi-F | CGTGGACCAATGCAACTCAA | 242 |
| Chi-R | ATTCCCTGTGCTGTCATCCA | |||
| glu | AF459794 | Glu-F | TGGAGAAGAATGGTGGAGGA | 188 |
| Glu-R | GTCAGACATGGCGAACACAT | |||
| act | AY859055 | ACT-F | TGGTATGGAAGCTGCAGGAA | 158 |
| ACT-R | GGGCTGTGATTTCCTTGCTC |
| Crop | Treatment | Stem Diam. (cm) a | Shoot Length (cm) | Stem Fresh Weight (g) | Fruit Weight (g) | Fruit Width (cm) |
|---|---|---|---|---|---|---|
| 1st | Control | 0.94 ± 0.19 | 193 ± 26.52 | 519.70 ± 22.63 | 1036.60 ± 22.86 | 38.84 ± 2.76 |
| Treatment | 0.98 ± 0.05 | 205.80 ± 30.71 | 589.44 ± 14.27 * | 1274.40 ± 60.78 * | 43.10 ± 3.84 * | |
| 2nd | Control | 1.15 ± 0.05 | 215 ± 10.62 | 862.87 ± 22.17 | 1450.50 ± 52.73 | 45.11 ± 0.68 |
| Treatment | 1.22 ± 0.12 | 211.50 ± 10.52 | 907.25 ± 29.78 * | 1529.90 ± 21.98 * | 48.35 ± 1.12 * |
| Crop | Treatment | Brix (°Brix) a | Texture (bar) |
|---|---|---|---|
| 1st | Control | 14.18 ± 1.94 | 4.80 ± 0.40 |
| Treatment | 14.60 ± 0.96 | 5.16 ± 0.79 | |
| 2nd | Control | 12.53 ± 0.86 | 5.40 ± 0.50 |
| Treatment | 13.00 ± 0.89 | 5.25 ± 0.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Intana, W.; Wonglom, P.; Suwannarach, N.; Sunpapao, A. Trichoderma asperelloides PSU-P1 Induced Expression of Pathogenesis-Related Protein Genes against Gummy Stem Blight of Muskmelon (Cucumis melo) in Field Evaluation. J. Fungi 2022, 8, 156. https://doi.org/10.3390/jof8020156
Intana W, Wonglom P, Suwannarach N, Sunpapao A. Trichoderma asperelloides PSU-P1 Induced Expression of Pathogenesis-Related Protein Genes against Gummy Stem Blight of Muskmelon (Cucumis melo) in Field Evaluation. Journal of Fungi. 2022; 8(2):156. https://doi.org/10.3390/jof8020156
Chicago/Turabian StyleIntana, Warin, Prisana Wonglom, Nakarin Suwannarach, and Anurag Sunpapao. 2022. "Trichoderma asperelloides PSU-P1 Induced Expression of Pathogenesis-Related Protein Genes against Gummy Stem Blight of Muskmelon (Cucumis melo) in Field Evaluation" Journal of Fungi 8, no. 2: 156. https://doi.org/10.3390/jof8020156
APA StyleIntana, W., Wonglom, P., Suwannarach, N., & Sunpapao, A. (2022). Trichoderma asperelloides PSU-P1 Induced Expression of Pathogenesis-Related Protein Genes against Gummy Stem Blight of Muskmelon (Cucumis melo) in Field Evaluation. Journal of Fungi, 8(2), 156. https://doi.org/10.3390/jof8020156