
Metabolomic and Physiological Changes in Fagus sylvatica Seedlings Infected with Phytophthora plurivora and the A1 and A2 Mating Types of P. ×cambivora

 , , , ,
, , , ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Inoculation
2.3. Sampling for Microscopic Studies
2.4. Re-Isolation of Phytophthora spp.
2.5. Physiological Measurements
2.6. Metabolomic Analyses
2.7. Data Analysis
2.7.1. Mortality
2.7.2. Physiology
2.7.3. Metabolomic Profiling
2.7.4. Visualization
3. Results
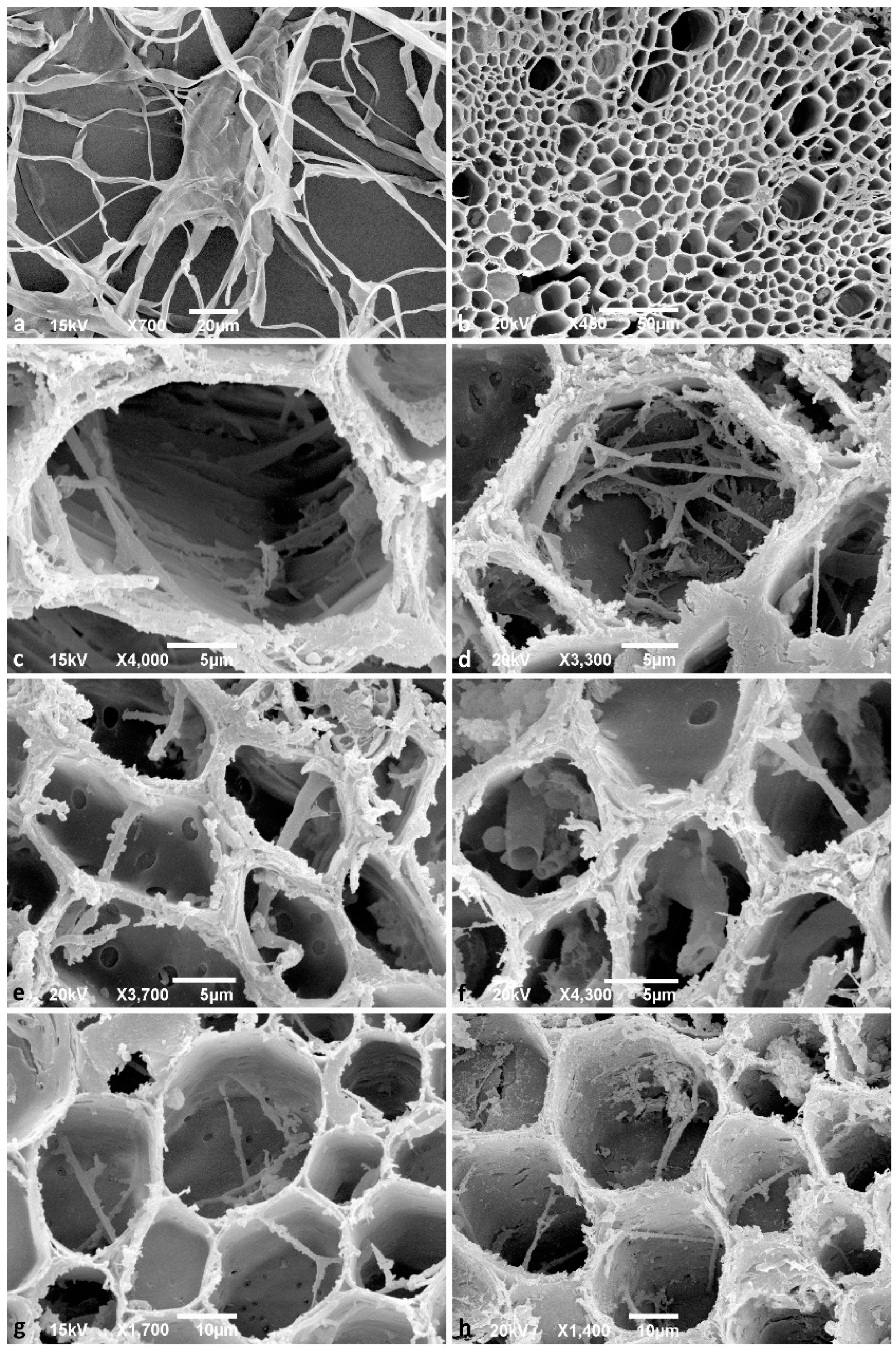
3.1. Viability of Isolates and Light and Scanning Electron Microscopy Observations
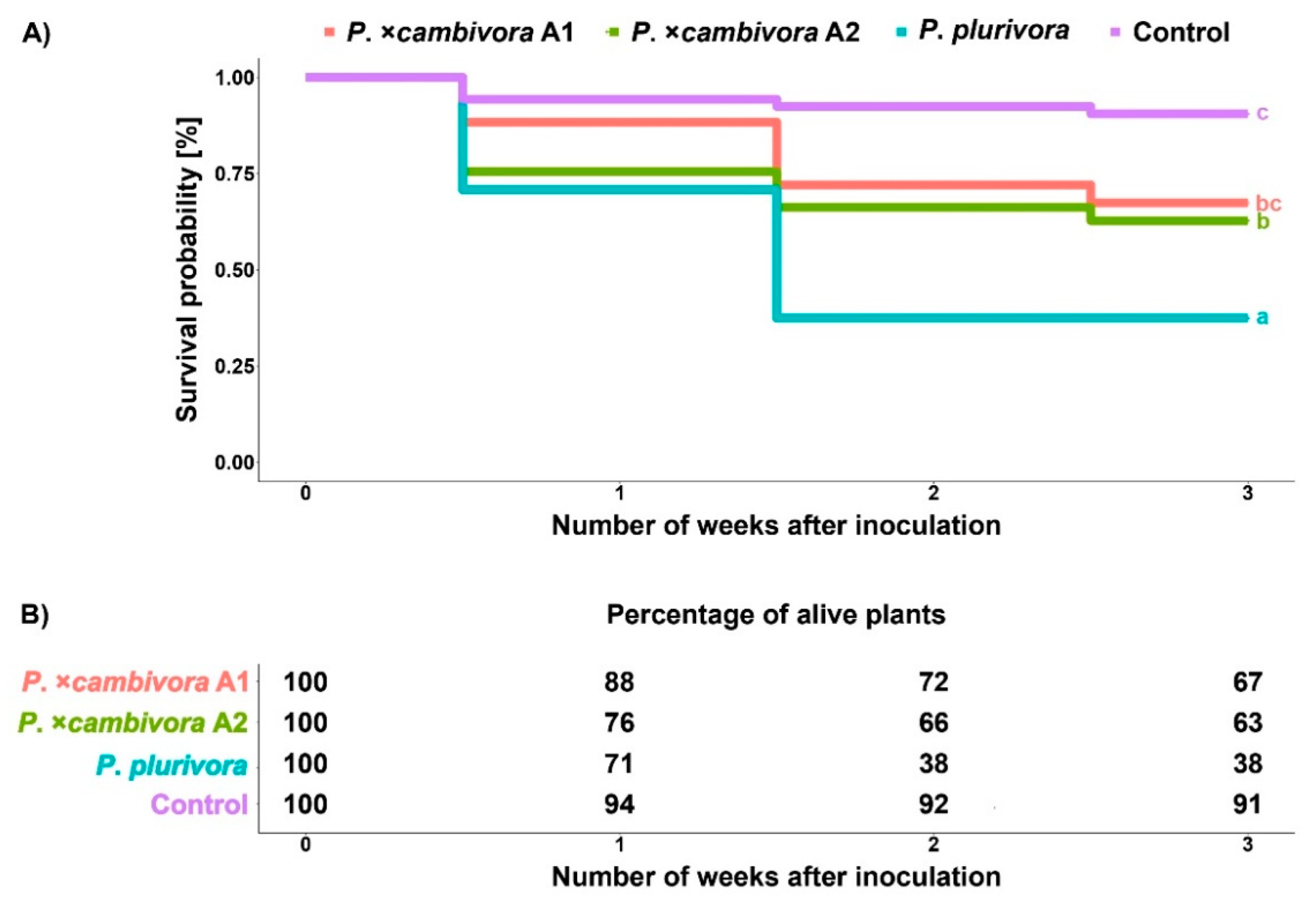
3.2. Survival Analysis
3.3. Physiological Measurements
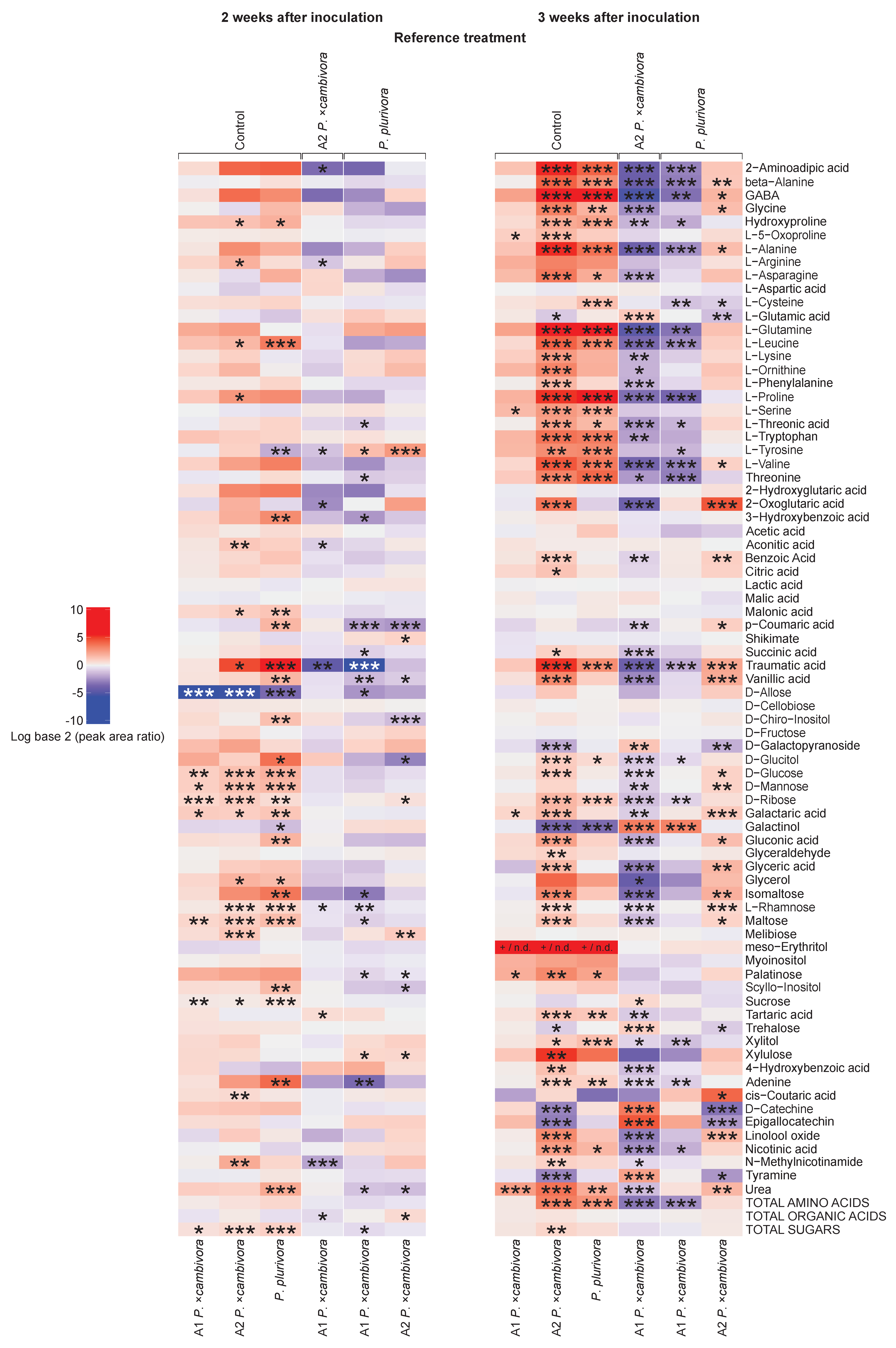
3.4. Root and Leaf Metabolomic Profiling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jung, T.; Pérez-Sierra, A.; Durán, A.; Jung, M.H.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Persoonia 2018, 40, 182–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, T.; Blaschke, H.; Neumann, P. Isolation, identification and pathogenicity of Phytophthora species from declining oak stands. Eur. J. For. Pathol. 1996, 26, 253–272. [Google Scholar] [CrossRef]
- Jung, T. Phytophthora quercina sp. nov., causing root rot of European oaks. Mycol. Res. 1999, 103, 785–798. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, H.; Oßwald, W. Involvement of soilborne Phytophthora Species in Central European oak decline and the effect of site factors on the disease. Plant Pathol. 2000, 49, 706–718. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Barzanti, G.P.; Bianco, M.C.; Ragazzi, A.; Capretti, P.; Paoletti, E.; Luisi, N.; Anselmi, N.; Vannini, A. Occurrence of Phytophthora species in oak stands in Italy and their association with declining oak trees. For. Pathol. 2002, 32, 19–28. [Google Scholar] [CrossRef]
- Jung, T. Beech decline in Central Europe driven by the interaction between Phytophthora infections and climatic extremes. For. Pathol. 2009, 39, 73–94. [Google Scholar] [CrossRef]
- Weiland, J.E.; Nelson, A.H.; Hudler, G.W. Aggressiveness of Phytophthora cactorum, P. citricola I, and P. plurivora from European beech. Plant Dis. 2010, 94, 1009–1014. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Vettraino, A.M.; Cech, T.L.; Vannini, A. The Impact of invasive Phytophthora species on European forests. In Phytophthora: A Global Perspective; Lamour, K., Ed.; CABI: Wallingford, UK, 2013; pp. 146–158. [Google Scholar]
- Corcobado, T.; Cech, T.L.; Brandstetter, M.; Daxer, A.; Hüttler, C.; Kudláček, T.; Jung, M.H.; Jung, T. Decline of European Beech in Austria: Involvement of Phytophthora spp. and contributing biotic and abiotic factors. Forests 2020, 11, 895. [Google Scholar] [CrossRef]
- Gibbs, J.; Lipscombe, M.; Peace, A. The impact of Phytophthora disease on riparian populations of Common alder (Alnus glutinosa) in Southern Britain. Eur. J. For. Pathol. 1999, 29, 39–50. [Google Scholar] [CrossRef]
- Streito, J.C.; Legrand, P.; Tabary, F.; Jarnouen De Villartay, G. Phytophthora disease of alder (Alnus glutinosa) in France: Investigations between 1995 and 1999. For. Pathol. 2002, 32, 179–191. [Google Scholar] [CrossRef]
- Brasier, C.M.; Kirk, S.A.; Delcan, J.; Cooke, D.E.L.; Jung, T.; Man In’t Veld, W.A. Phytophthora alni sp. nov. and its variants: Designation of emerging heteroploid hybrid pathogens spreading on Alnus trees. Mycol. Res. 2004, 108, 1172–1184. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, M. Phytophthora root and collar rot of alders in Bavaria: Distribution, modes of spread and possible management strategies. Plant Pathol. 2004, 53, 197–208. [Google Scholar] [CrossRef]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Aguín Casal, O.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Pathol. 2016, 46, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Nechwatal, J.; Cooke, D.E.L.; Hartmann, G.; Blaschke, M.; Oßwald, W.F.; Duncan, J.M.; Delatour, C. Phytophthora pseudosyringae sp. nov., a new species causing root and collar rot of deciduous tree species in Europe. Mycol. Res. 2003, 107, 772–789. [Google Scholar] [CrossRef]
- Jung, T.; Hudler, G.W.; Jensen-Tracy, S.L.; Griffiths, H.M.; Fleischmann, F.; Osswald, W. Involvement of Phytophthora species in the decline of European beech in Europe and the USA. Mycologist 2005, 19, 159–166. [Google Scholar] [CrossRef]
- Jung, T.; Jung, M.H.; Scanu, B.; Seress, D.; Kovács, G.M.; Maia, C.; Pérez-Sierra, A.; Chang, T.T.; Chandelier, A.; Heungens, K.; et al. Six new Phytophthora species from ITS Clade 7a including two sexually functional heterothallic hybrid species detected in natural ecosystems in Taiwan. Persoonia 2017, 38, 100–135. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; La Spada, F.; Pane, A.; Aloi, F.; Evoli, M.; Jung, M.H.; Scanu, B.; Faedda, R.; Rizza, C.; Puglisi, I.; et al. Diversity and distribution of Phytophthora Species in protected natural areas in Sicily. Forests 2019, 10, 259. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, S.; Zini, J.; Chandelier, A. Involvement of Phytophthora species in the decline of beech (Fagus sylvatica) in the southern part of Belgium. In Phytophthoras in Forests and Natural Ecosystems: Fourth Meeting of the International Union of Forest Research Organizations (IUFRO) Working Party S07.02.09; Goheen, E., Frankel, S., Eds.; USDA Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2009; p. 320. [Google Scholar]
- Nechwatal, J.; Hahn, J.; Schönborn, A.; Schmitz, G. A twig blight of understorey European beech (Fagus sylvatica) caused by soilborne Phytophthora spp. For. Pathol. 2011, 41, 493–500. [Google Scholar] [CrossRef]
- Telfer, K.H.; Brurberg, M.B.; Herrero, M.L.; Stensvand, A.; Talgø, V. Phytophthora cambivora found on beech in Norway. For. Pathol. 2015, 45, 415–425. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Morel, O.; Perlerou, C.; Robin, C.; Diamandis, S.; Vannini, A. Occurrence and distribution of Phytophthora Species in European chestnut stands, and their association with Ink disease and crown decline. Eur. J. Plant Pathol. 2005, 111, 169–180. [Google Scholar] [CrossRef]
- Saavedra, A.; Hansen, E.M.; Goheen, D.J. Phytophthora cambivora in Oregon and its pathogenicity to Chrysolepis Chrysophylla. For. Pathol. 2007, 37, 409–419. [Google Scholar] [CrossRef]
- Reeser, P.; Sutton, W.; Hansen, E. Phytophthora species associated with stem cankers on tanoak in Southwestern Oregon. In Proceedings of the Sudden Oak Death Third Science Symposium; Frankel, S.J., Kliejunas, J.T., Palmieri, K.M., Eds.; Gen. Tech. Rep. PSW-GTR-214; US Department of Agriculture, Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2008; pp. 227–229. [Google Scholar]
- Greslebin, A.G.; Hansen, E.M.; Winton, L.M.; Rajchenberg, M. Phytophthora species from declining Austrocedrus chilensis forests in Patagonia, Argentina. Mycologia 2005, 97, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.; Durán, A.; Sanfuentes von Stowasser, E.; Schena, L.; Mosca, S.; Fajardo, S.; González, M.; Navarro Ortega, A.D.; Bakonyi, J.; Seress, D.; et al. Diversity of Phytophthora species in Valdivian rainforests and association with severe dieback symptoms. For. Pathol. 2018, 48, e12443. [Google Scholar] [CrossRef]
- Suzui, T.; Hoshino, Y. Collar rot of apple caused by Phytophthora cambivora (Petri) Buism. Jpn. J. Phytopathol. 1979, 45, 344–352. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Jung, M.H.; Cacciola, S.O.; Cech, T.; Bakonyi, J.; Seress, D.; Mosca, S.; Schena, L.; Seddaiu, S.; Pane, A.; et al. Multiple new cryptic pathogenic Phytophthora species from Fagaceae forests in Austria, Italy and Portugal. IMA Fungus 2017, 8, 219–244. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Burgess, T.I. Re-evaluation of Phytophthora citricola isolates from multiple woody hosts in Europe and North America reveals a new species, Phytophthora plurivora sp. nov. Persoonia 2009, 22, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Fleischmann, F.; Schneider, D.; Matyssek, R.; Oßwald, W.F. Investigations on net CO2 assimilation, transpiration and root growth of Fagus sylvatica infested with four different Phytophthora species. Plant Biol. 2002, 4, 144–152. [Google Scholar] [CrossRef]
- Fleischmann, F.; Göttlein, A.; Rodenkirchen, H.; Lütz, C.; Oßwald, W. Biomass, nutrient and pigment content of beech (Fagus sylvatica) saplings infected with Phytophthora citricola, P. cambivora, P. pseudosyringae and P. undulata. For. Pathol. 2004, 34, 79–92. [Google Scholar] [CrossRef]
- Jung, T.; Chang, T.T.; Bakonyi, J.; Seress, D.; Pérez-Sierra, A.; Yang, X.; Hong, C.; Scanu, B.; Fu, C.H.; Hsueh, K.L.; et al. Diversity of Phytophthora species in natural ecosystems of Taiwan and association with disease symptoms. Plant Pathol. 2017, 66, 194–211. [Google Scholar] [CrossRef]
- Jung, T.; Scanu, B.; Brasier, C.M.; Webber, J.; Milenković, I.; Corcobado, T.; Tomšovský, M.; Pánek, M.; Bakonyi, J.; Maia, C.; et al. A survey in natural forest ecosystems of Vietnam reveals high diversity of both new and described Phytophthora taxa including P. ramorum. Forests 2020, 11, 93. [Google Scholar] [CrossRef] [Green Version]
- Vélez, M.L.; Silva, P.V.; Troncoso, O.A.; Greslebin, A.G. Alteration of physiological parameters of Austrocedrus chilensis by the pathogen Phytophthora austrocedrae. Plant Pathol. 2012, 61, 877–888. [Google Scholar] [CrossRef]
- Angay, O.; Fleischmann, F.; Recht, S.; Herrmann, S.; Matyssek, R.; Oßwald, W.; Buscot, F.; Grams, T.E.E. Sweets for the foe—effects of nonstructural carbohydrates on the susceptibility of Quercus robur against Phytophthora quercina. New Phytol. 2014, 203, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Reeksting, B.J.; Taylor, N.J.; van den Berg, N. Flooding and Phytophthora cinnamomi: Effects on photosynthesis and chlorophyll fluorescence in shoots of non-grafted Persea americana (Mill.) rootstocks differing in tolerance to Phytophthora root rot. S. Afr. J. Bot. 2014, 95, 40–53. [Google Scholar] [CrossRef] [Green Version]
- Gold, K.M.; Townsend, P.A.; Larson, E.R.; Herrmann, I.; Gevens, A.J. Contact reflectance spectroscopy for rapid, accurate, and nondestructive Phytophthora infestans clonal lineage discrimination. Phytopathology 2020, 110, 851–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newby, Z.; Murphy, R.J.; Guest, D.I.; Ramp, D.; Liew, E.C.Y. Detecting symptoms of Phytophthora cinnamomi infection in Australian native vegetation using reflectance spectrometry: Complex effects of water stress and species susceptibility. Australas. Plant Pathol. 2019, 48, 409–424. [Google Scholar] [CrossRef]
- Manter, D.K.; Kelsey, R.G.; Karchesy, J.J. Photosynthetic declines in Phytophthora ramorum-infected plants develop prior to water stress and in response to exogenous application of elicitins. Phytopathology 2007, 97, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Abu-Nada, Y.; Kushalappa, A.C.; Marshall, W.D.; Al-Mughrabi, K.; Murphy, A. Temporal dynamics of pathogenesis-related metabolites and their plausible pathways of induction in potato leaves following inoculation with Phytophthora infestans. Eur. J. Plant Pathol. 2007, 118, 375–391. [Google Scholar] [CrossRef]
- Zhu, L.; Zhou, Y.; Li, X.; Zhao, J.; Guo, N.; Xing, H. Metabolomics analysis of soybean hypocotyls in response to Phytophthora sojae infection. Front. Plant Sci. 2018, 9, 1530. [Google Scholar] [CrossRef] [Green Version]
- Rouse, J.W.; Hass, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. Third ERTS Symp. 1974, 1, 309–317. [Google Scholar]
- Peñuelas, J.; Filella, I.; Gamon, J.A. Assessment of photosynthetic radiation-use efficiency with spectral reflectance. New Phytol. 1995, 131, 291–296. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Zur, Y.; Chivkunova, O.B.; Merzlyak, M.N. Assessing carotenoid content in plant leaves with reflectance spectroscopy. Photochem. Photobiol. 2002, 75, 272–281. [Google Scholar] [CrossRef]
- Berka, M.; Luklová, M.; Dufková, H.; Berková, V.; Novák, J.; Saiz-Fernández, I.; Rashotte, A.M.; Brzobohatý, B.; Černý, M. Barley root proteome and metabolome in response to cytokinin and abiotic stimuli. Front. Plant Sci. 2020, 11, 1647. [Google Scholar] [CrossRef] [PubMed]
- Therneau, T. A Package for Survival Analysis in S. R Package Version. 2015, Volume 2. Available online: https://sites.google.com/site/statsr4us/docs/SurvivalanalysisinR_package.pdf (accessed on 16 February 2022).
- Kassambara, A.; Kosinski, M. Survminer: Drawing Survival Curves Using ‘Ggplot2’. R Package Version 0.3. 2017. Available online: https://rpkgs.datanovia.com/survminer/ (accessed on 16 February 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeileis, A. Econometric computing with HC and HAC covariance matrix estimators. J. Stat. Softw. 2004, 11, 1–17. [Google Scholar] [CrossRef]
- Zeileis, A. Object-oriented computation of sandwich estimators. J. Stat. Softw. 2006, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Herberich, E.; Sikorski, J.; Hothorn, T. A robust procedure for comparing multiple means under heteroscedasticity in unbalanced designs. PLoS ONE 2010, 5, e9788. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Inkscape Project. Inkscape. 2020. Available online: https://inkscape.org/news/2020/ (accessed on 16 February 2022).
- Zarco-Tejada, P.J.; González-Dugo, M.V.; Fereres, E. Seasonal stability of chlorophyll fluorescence quantified from airborne hyperspectral imagery as an indicator of net photosynthesis in the context of precision agriculture. Remote Sens. Environ. 2016, 179, 89–103. [Google Scholar] [CrossRef]
- Sanogo, S.; El-Sebai, O.I.; Sanderson, R. Severity of Verticillium wilt, plant growth, and spectral reflectance indices of Chile pepper under periodic flooding and no-flooding conditions. HortScience 2008, 43, 414–419. [Google Scholar] [CrossRef] [Green Version]
- Sankaran, S.; Ehsani, R.; Inch, S.A.; Ploetz, R.C. Evaluation of visible-near infrared reflectance spectra of avocado leaves as a non-destructive sensing tool for detection of laurel wilt. Plant Dis. 2012, 96, 1683–1689. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.; Bergsträsser, S.; Schmittgen, S.; Müller-Linow, M.; Rascher, U. Non-invasive spectral phenotyping methods can improve and accelerate Cercospora disease scoring in sugar beet breeding. Agriculture 2014, 4, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Dalio, R.J.D.; Fleischmann, F.; Chambery, A.; Eichmann, R.; Massola, N.S.; Pascholati, S.F.; Osswald, W. Immunodepletion of α-Plurivorin effector leads to loss of virulence of Phytophthora plurivora towards Fagus sylvatica. For. Pathol. 2017, 47, e12362. [Google Scholar] [CrossRef]
- Sghaier-Hammami, B.; Valero-Galvàn, J.; Romero-Rodríguez, M.C.; Navarro-Cerrillo, R.M.; Abdelly, C.; Jorrín-Novo, J. Physiological and proteomics analyses of Holm Oak (Quercus ilex subsp. ballota [Desf.] Samp.) responses to Phytophthora cinnamomi. Plant Physiol. Biochem. 2013, 71, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Saiz-Fernández, I.; Milenković, I.; Berka, M.; Černý, M.; Tomšovský, M.; Brzobohatý, B.; Kerchev, P. Integrated proteomic and metabolomic profiling of Phytophthora cinnamomi attack on Sweet chestnut (Castanea sativa) reveals distinct molecular reprogramming proximal to the infection site and away from it. Int. J. Mol. Sci. 2020, 21, 8525. [Google Scholar] [CrossRef]
- Saiz-Fernández, I.; De Diego, N.; Brzobohatý, B.; Muñoz-Rueda, A.; Lacuesta, M. The imbalance between C and N metabolism during high nitrate supply inhibits photosynthesis and overall growth in maize (Zea mays L.). Plant Physiol. Biochem. 2017, 120, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Clemenz, C.; Fleischmann, F.; Häberle, K.H.; Matyssek, R.; Oßwald, W. Photosynthetic and leaf water potential responses of Alnus glutinosa saplings to stem-base inoculaton with Phytophthora alni subsp. alni. Tree Physiol. 2008, 28, 1703–1711. [Google Scholar] [CrossRef] [Green Version]
- Tauzin, A.S.; Giardina, T. Sucrose and invertases, a part of the plant defense response to the biotic stresses. Front. Plant Sci. 2014, 5, 293. [Google Scholar] [CrossRef]
- Sperdouli, I.; Moustakas, M. Differential response of photosystem II photochemistry in young and mature leaves of Arabidopsis thaliana to the onset of drought stress. Acta Physiol. Plant. 2012, 34, 1267–1276. [Google Scholar] [CrossRef]
- Maurel, M.; Robin, C.; Simonneau, T.; Loustau, D.; Dreyer, E.; Desprez-Loustau, M.L. Stomatal conductance and root-to-shoot signalling in chestnut saplings exposed to Phytophthora cinnamomi or partial soil drying. Funct. Plant Biol. 2004, 31, 41–51. [Google Scholar] [CrossRef]
- Oßwald, W.; Fleischmann, F.; Rigling, D.; Coelho, A.C.; Cravador, A.; Diez, J.; Dalio, R.J.; Horta Jung, M.; Pfanz, H.; Robin, C.; et al. Strategies of attack and defence in woody plant-Phytophthora interactions. For. Pathol. 2014, 44, 169–190. [Google Scholar] [CrossRef]
- Ruiz Gómez, F.J.; Navarro-Cerrillo, R.M.; Sánchez-Cuesta, R.; Pérez-de-Luque, A. Histopathology of infection and colonization of Quercus ilex fine roots by Phytophthora cinnamomi. Plant Pathol. 2015, 64, 605–616. [Google Scholar] [CrossRef]
- Ruiz-Gómez, F.J.; Pérez-de-Luque, A.; Navarro-Cerrillo, R.M. The involvement of Phytophthora root rot and drought stress in holm oak decline: From ecophysiology to microbiome influence. Curr. For. Rep. 2019, 5, 251–266. [Google Scholar] [CrossRef]
- Vandana, V.V.; Suseela Bhai, R.; Ramakrishnan Nair, R.; Azeez, S. Role of cell wall and cell membrane integrity in imparting defense response against Phytophthora capsici in black pepper (Piper nigrum L.). Eur. J. Plant Pathol. 2019, 154, 359–375. [Google Scholar] [CrossRef]
- Parker, D.; Beckmann, M.; Zubair, H.; Enot, D.P.; Caracuel-Rios, Z.; Overy, D.P.; Snowdon, S.; Talbot, N.J.; Draper, J. Metabolomic analysis reveals a common pattern of metabolic re-programming during invasion of three host plant species by Magnaporthe grisea. Plant J. 2009, 59, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Draper, J.; Rasmussen, S.; Zubair, H. Metabolite analysis and metabolomics in the study of biotrophic interactions between plants and microbes. Annu. Plant Rev. 2018, 43, 25–59. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Bhat, J.A.; Siddiqui, M.H.; Rinklebe, J.; Ahmad, P. Integration of silicon and secondary metabolites in plants: A significant association in stress tolerance. J. Exp. Bot. 2020, 71, 6758–6774. [Google Scholar] [CrossRef]
- Tenenboim, H.; Brotman, Y. Omic relief for the biotically stressed: Metabolomics of plant biotic interactions. Trends Plant Sci. 2016, 21, 781–791. [Google Scholar] [CrossRef]
- Judelson, H.S.; Tani, S.; Narayan, R.D. Metabolic adaptation of Phytophthora infestans during growth on leaves, tubers and artificial media. Mol. Plant Pathol. 2009, 10, 843–855. [Google Scholar] [CrossRef]
- Rodenburg, S.Y.; Seidl, M.F.; Judelson, H.S.; Vu, A.L.; Govers, F.; de Ridder, D. Metabolic model of the Phytophthora infestans-tomato interaction reveals metabolic switches during host colonization. MBio 2019, 10, e00454-19. [Google Scholar] [CrossRef] [Green Version]
- Dumschott, K.; Richter, A.; Loescher, W.; Merchant, A. Post photosynthetic carbon partitioning to sugar alcohols and consequences for plant growth. Phytochemistry 2017, 144, 243–252. [Google Scholar] [CrossRef]
- Lee, D.K.; Ahn, S.; Cho, H.Y.; Yun, H.Y.; Park, J.H.; Lim, J.; Lee, J.; Kwon, S.W. Metabolic response induced by parasitic plant-fungus interactions hinder amino sugar and nucleotide sugar metabolism in the host. Sci. Rep. 2016, 6, 37434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, J.D.; Jennings, D.B.; Guo, W.W.; Pharr, D.M.; Ehrenshaft, M. Sugar alcohols, salt stress, and fungal resistance: Polyols—multifunctional plant protection? J. Am. Soc. Hortic. Sci. 2002, 127, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Calmes, B.; Guillemette, T.; Teyssier, L.; Siegler, B.; Pigné, S.; Landreau, A.; Iacomi, B.; Lemoine, R.; Richomme, P.; Simoneau, P. role of mannitol metabolism in the pathogenicity of the necrotrophic fungus Alternaria brassicicola. Front. Plant Sci. 2013, 4, 131. [Google Scholar] [CrossRef] [Green Version]
- Eshel, G.; Shaked, R.; Kazachkova, Y.; Khan, A.; Eppel, A.; Cisneros, A.; Acuna, T.; Gutterman, Y.; Tel-Zur, N.; Rachmilevitch, S.; et al. Anastatica hierochuntica, an Arabidopsis desert relative, is tolerant to multiple abiotic stresses and exhibits species-specific and common stress tolerance strategies with its halophytic relative, Eutrema (Thellungiella) salsugineum. Front. Plant Sci. 2017, 7, 1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkama-Ohtsu, N.; Oikawa, A.; Zhao, P.; Xiang, C.; Saito, K.; Oliver, D.J. A γ-glutamyl transpeptidase-independent pathway of glutathione catabolism to glutamate via 5-oxoproline in Arabidopsis. Plant Physiol. 2008, 148, 1603–1613. [Google Scholar] [CrossRef] [Green Version]
- Gullner, G.; Komives, T.; Király, L.; Schröder, P. Glutathione S-transferase enzymes in plant-pathogen interactions. Front. Plant Sci. 2018, 9, 1836. [Google Scholar] [CrossRef] [Green Version]
- Vasyukova, N.I.; Chalenko, G.I.; Gerasimova, N.G.; Ozeretskovskaya, O.L. Wound repair in plant tissues (review). Appl. Biochem. Microbiol. 2011, 47, 229–233. [Google Scholar] [CrossRef]
- Pietryczuk, A.; Czerpak, R. Effect of traumatic acid on antioxidant activity in Chlorella vulgaris (Chlorophyceae). Plant Growth Regul. 2011, 65, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corcobado, T.; Milenković, I.; Saiz-Fernández, I.; Kudláček, T.; Plichta, R.; Májek, T.; Bačová, A.; Ďatková, H.; Dálya, L.B.; Trifković, M.; et al. Metabolomic and Physiological Changes in Fagus sylvatica Seedlings Infected with Phytophthora plurivora and the A1 and A2 Mating Types of P. ×cambivora. J. Fungi 2022, 8, 298. https://doi.org/10.3390/jof8030298
Corcobado T, Milenković I, Saiz-Fernández I, Kudláček T, Plichta R, Májek T, Bačová A, Ďatková H, Dálya LB, Trifković M, et al. Metabolomic and Physiological Changes in Fagus sylvatica Seedlings Infected with Phytophthora plurivora and the A1 and A2 Mating Types of P. ×cambivora. Journal of Fungi. 2022; 8(3):298. https://doi.org/10.3390/jof8030298
Chicago/Turabian StyleCorcobado, Tamara, Ivan Milenković, Iñigo Saiz-Fernández, Tomáš Kudláček, Roman Plichta, Tomáš Májek, Aneta Bačová, Henrieta Ďatková, László Benedek Dálya, Miloš Trifković, and et al. 2022. "Metabolomic and Physiological Changes in Fagus sylvatica Seedlings Infected with Phytophthora plurivora and the A1 and A2 Mating Types of P. ×cambivora" Journal of Fungi 8, no. 3: 298. https://doi.org/10.3390/jof8030298
APA StyleCorcobado, T., Milenković, I., Saiz-Fernández, I., Kudláček, T., Plichta, R., Májek, T., Bačová, A., Ďatková, H., Dálya, L. B., Trifković, M., Mureddu, D., Račko, V., Kardošová, M., Ďurkovič, J., Rattunde, R., & Jung, T. (2022). Metabolomic and Physiological Changes in Fagus sylvatica Seedlings Infected with Phytophthora plurivora and the A1 and A2 Mating Types of P. ×cambivora. Journal of Fungi, 8(3), 298. https://doi.org/10.3390/jof8030298