
DNA Metabarcoding and Isolation by Baiting Complement Each Other in Revealing Phytophthora Diversity in Anthropized and Natural Ecosystems

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
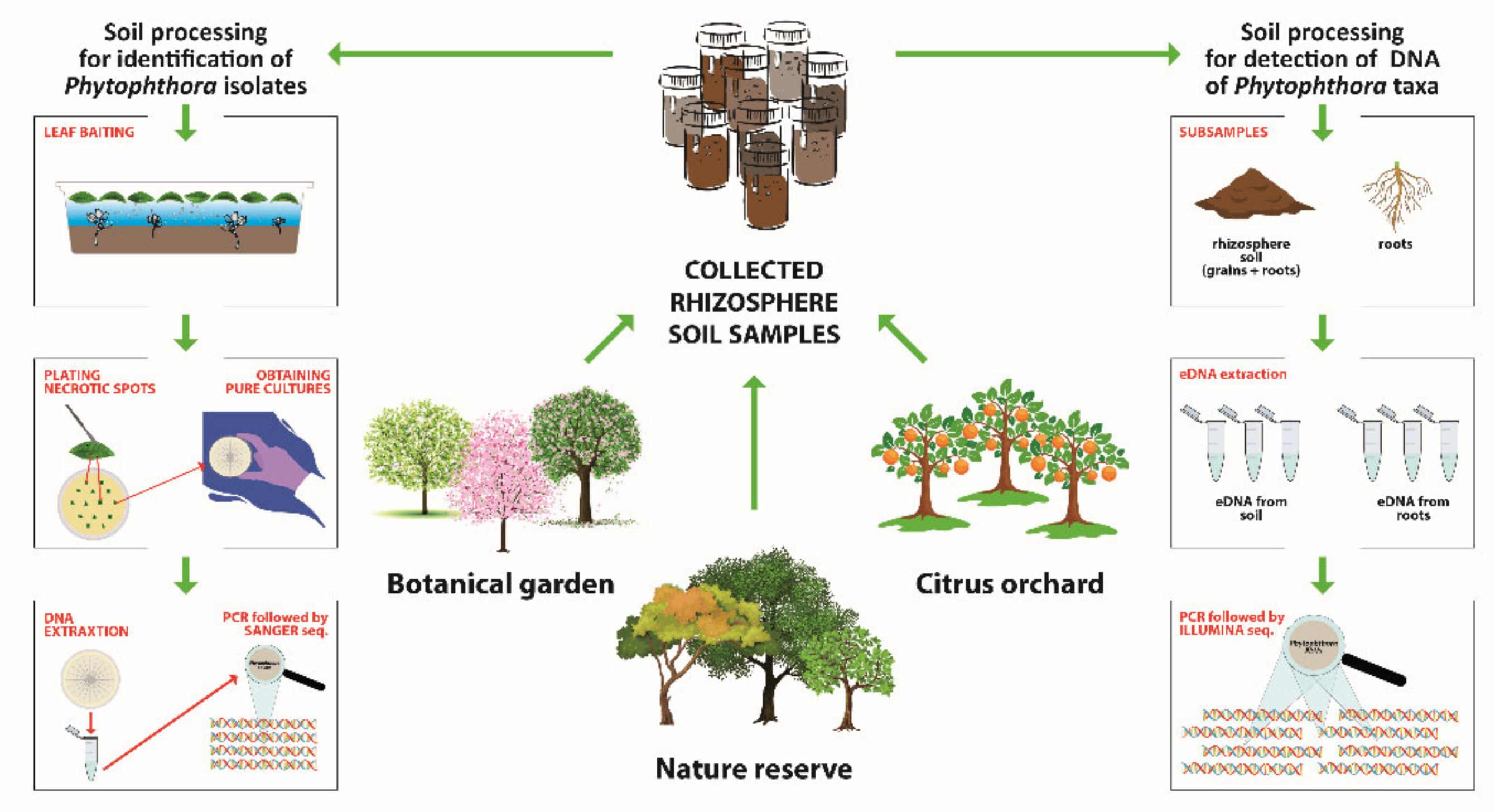
2.1. Sampling Areas and Collection of Rhizosphere Soil Samples
2.2. DNA Extraction from Rhizosphere Soil Samples for Metabarcoding Analyses
2.3. PCR Reactions
2.4. Illumina Sequencing Library Preparation and Sequencing
2.5. Analyses of Illumina Data
2.6. Isolation by Baiting and Identification of Phytophthora Isolates
3. Results
3.1. Phytophthora Species Detected by Illumina Sequencing from Environmental DNA
3.1.1. Nature Reserve
3.1.2. Botanical Garden
3.1.3. Citrus Orchard
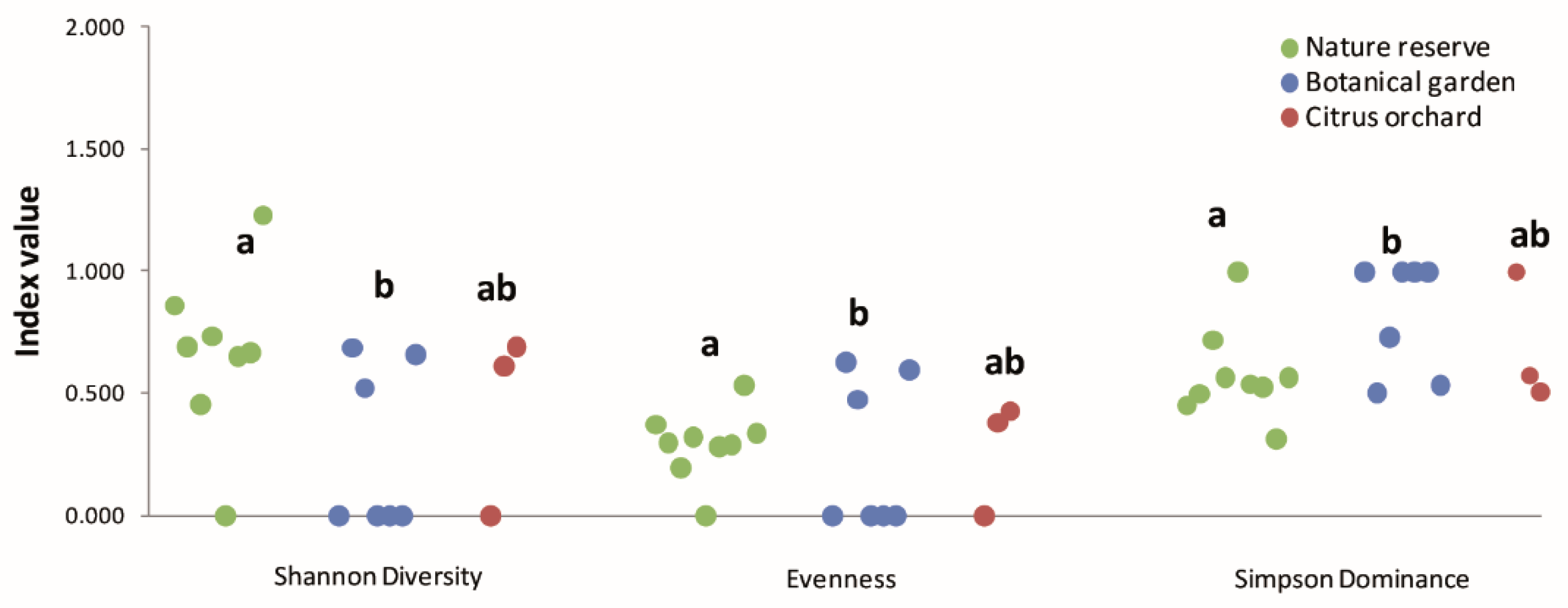
3.1.4. Analyses of Biodiversity Resulting from DNA Metabarcoding
3.2. Phytophthora Species Isolated by Baiting
3.2.1. Nature Reserve
3.2.2. Botanical Garden
3.2.3. Citrus Orchard
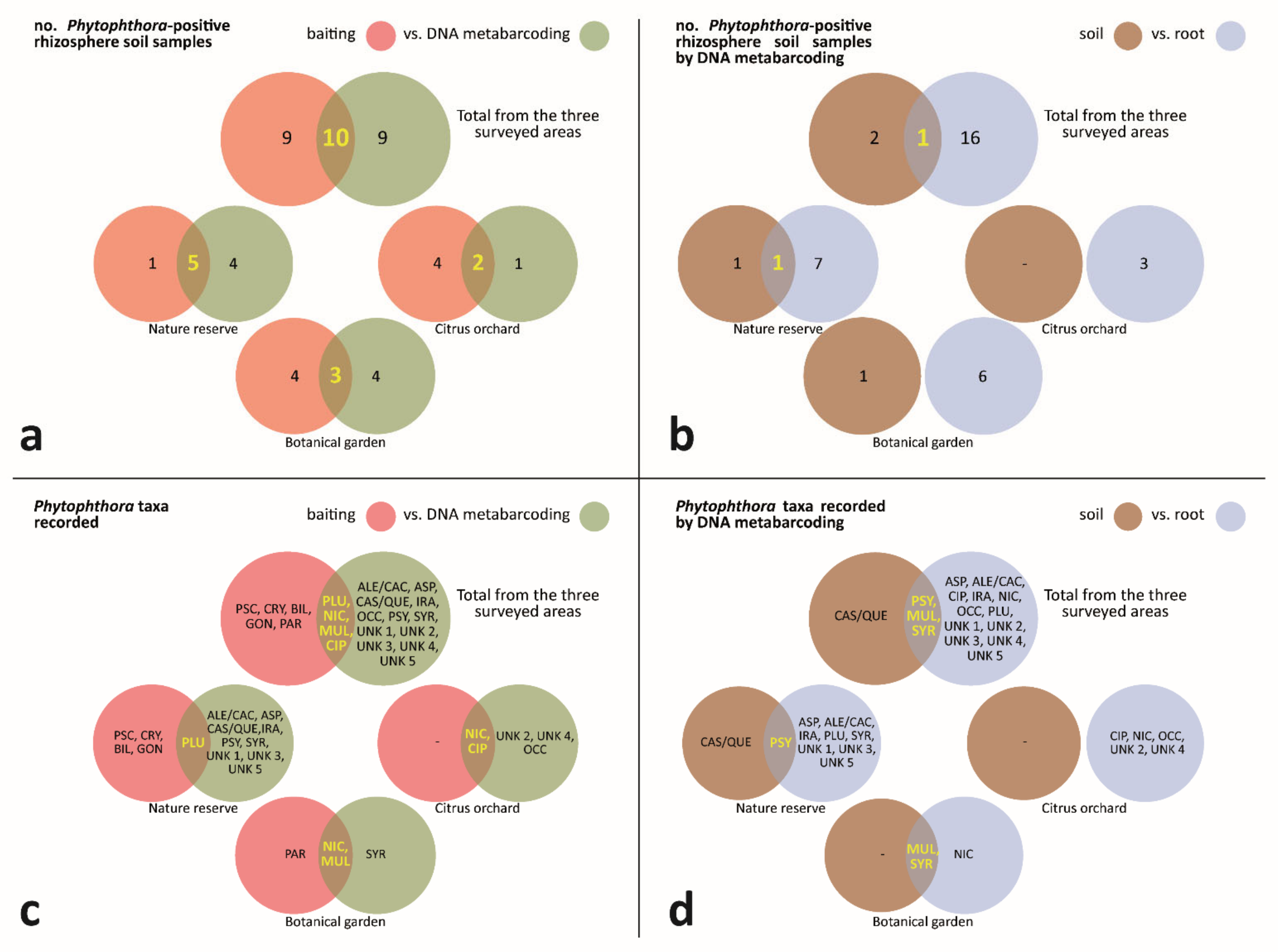
3.3. Phytophthora-Positive Rhizosphere Soil Samples—Comparison of Outcomes from Baiting and DNA Metabarcoding
3.3.1. Phytophthora-Positive Rhizosphere Soil Samples—Comparison of DNA Metabarcoding Outcomes from Processed Samples of Soil and Root
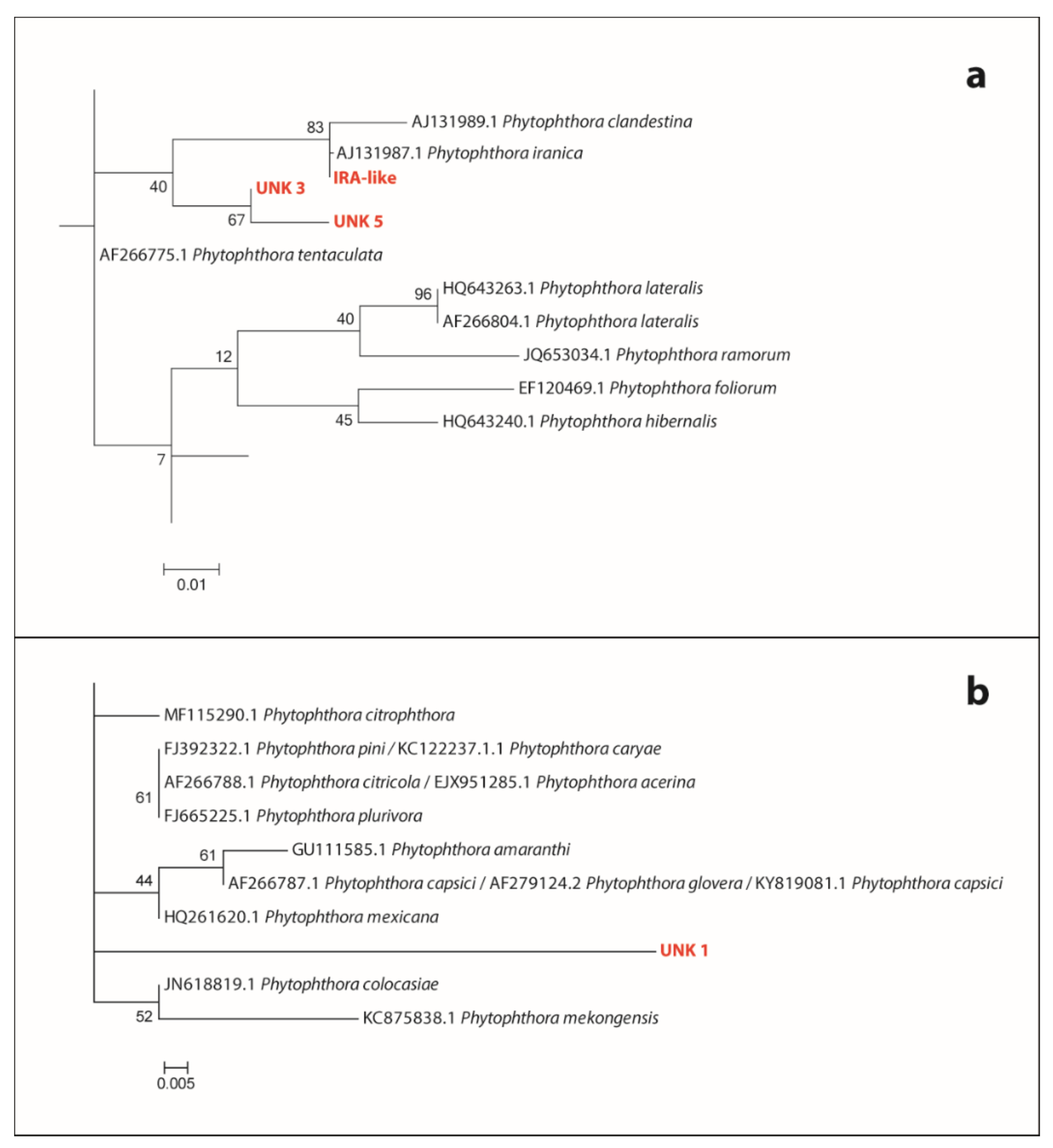
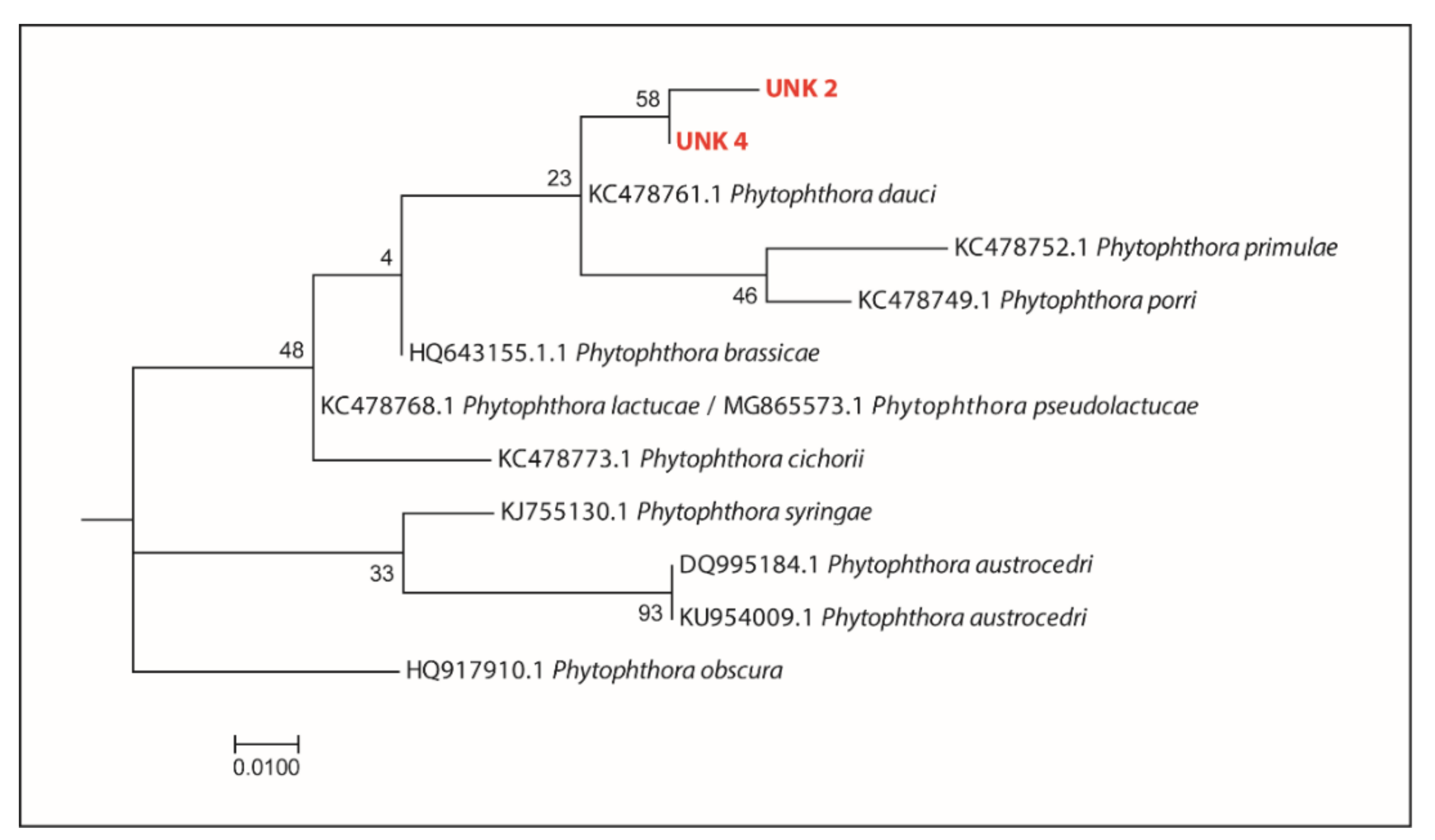
3.3.2. Phytophthora Taxa Recorded—Comparison of Outcomes from Baiting and DNA Metabarcoding
3.3.3. Phytophthora Taxa Recorded—Comparison of DNA Metabarcoding Outcomes from Processed Samples of Soil and Root
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riddell, C.E.; Frederickson-Matika, D.; Armstrong, A.C.; Elliot, M.; Forster, J.; Hedley, P.E.; Morris, J.; Thorpe, P.; Cooke, D.E.L.; Pritchard, L.; et al. Metabarcoding reveals a high diversity of woody host-associated Phytophthora spp. in soils at public gardens and amenity woodlands in Britain. PeerJ 2019, 7, e6931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scanu, B.; Jung, T.; Masigol, H.; Linaldeddu, B.T.; Jung, M.H.; Brandano, A.; Mostowfizadeh-Ghalamfarsa, R.; Janoušek, J.; Riolo, M.; Cacciola, S.O. Phytophthora heterospora sp. nov., a New Pseudoconidia-Producing Sister Species of P. palmivora. J. Fungi 2021, 7, 870. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C.M. Phytophthora cinnamomi and oak decline in southern Europe. Environmental constraints including climate change. Ann. Des. Sci. For. 1996, 53, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Cooke, D.E.L.; Schena, L.; Cacciola, S.O. Tools to detect, identify and monitor Phytophthora species in natural ecosystems. J. Plant Pathol. 2007, 89, 13–28. [Google Scholar]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; American Phytopathological Society (APS Press): St. Paul, MN, USA, 1996; ISBN 0890542120. [Google Scholar]
- Jung, T.; Jung, M.H.; Scanu, B.; Seress, D.; Kovács, G.M.; Maia, C.; Pérez-Sierra, A.; Chang, T.T.; Chandelier, A.; Heungens, K.; et al. Six new Phytophthora species from ITS Clade 7a including two sexually functional heterothallic hybrid species detected in natural ecosystems in Taiwan. Persoonia-Mol. Phylogeny Evol. Fungi 2017, 38, 100–135. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Jung, M.H.; Cacciola, S.O.; Cech, T.; Bakonyi, J.; Seress, D.; Mosca, S.; Schena, L.; Seddaiu, S.; Pane, A.; et al. Multiple new cryptic pathogenic Phytophthora species from Fagaceae forests in Austria, Italy and Portugal. IMA Fungus 2017, 8, 219–244. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; La Spada, F.; Pane, A.; Aloi, F.; Evoli, M.; Jung, M.H.; Scanu, B.; Faedda, R.; Rizza, C.; Puglisi, I.; et al. Diversity and Distribution of Phytophthora Species in Protected Natural Areas in Sicily. Forests 2019, 10, 259. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, S.; Furzer, O.; Jones, J.D.G.; Judelson, H.S.; Ali, G.S.; Dalio, R.J.D.; Roy, S.G.; Schena, L.; Zambounis, A.; Panabières, F.; et al. The Top 10 oomycete pathogens in molecular plant pathology. Mol. Plant Pathol. 2015, 16, 413–434. [Google Scholar] [CrossRef]
- Ristaino, J.B.; Gumpertz, M.L. New Frontiers in the Study of Dispersal and Spatial Analysis of Epidemics Caused by Species in the Genus Phytophthora. Annu. Rev. Phytopathol. 2000, 38, 541–576. [Google Scholar] [CrossRef] [Green Version]
- Burgess, T.I.; Scott, J.K.; Mcdougall, K.L.; Stukely, M.J.C.; Crane, C.; Dunstan, W.A.; Brigg, F.; Andjic, V.; White, D.; Rudman, T.; et al. Current and projected global distribution of Phytophthora cinnamomi, one of the world’s worst plant pathogens. Glob. Chang. Biol. 2017, 23, 1661–1674. [Google Scholar] [CrossRef] [Green Version]
- Frisullo, S.; Lima, G.; di San Lio, G.M.; Camele, I.; Melissano, L.; Puglisi, I.; Pane, A.; Agosteo, G.E.; Prudente, L.; Cacciola, S.O. Phytophthora cinnamomi Involved in the Decline of Holm Oak (Quercus ilex) Stands in Southern Italy. For. Sci. 2018, 64, 290–298. [Google Scholar] [CrossRef]
- Hardham, A.R.; Blackman, L.M. Pathogen profile update Phytophthora cinnamomi. Mol. Plant Pathol. 2018, 19, 260–285. [Google Scholar] [CrossRef] [Green Version]
- Shakya, S.K.; Grünwald, N.J.; Fieland, V.J.; Knaus, B.J.; Weiland, J.E.; Maia, C.; Drenth, A.; Guest, D.I.; Liew, E.C.Y.; Crane, C.; et al. Phylogeography of the wide-host range panglobal plant pathogen Phytophthora cinnamomi. Mol. Ecol. 2021, 30, 5164–5178. [Google Scholar] [CrossRef]
- Scott, P.M.; Burgess, T.I.; Barber, P.A.; Shearer, B.L.; Stukely, M.J.C.; Hardy, G.E.S.J.; Jung, T. Phytophthora multivora sp. nov., a new species recovered from declining Eucalyptus, Banksia, Agonis and other plant species in Western Australia. Persoonia 2009, 22, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Migliorini, D.; Khdiar, M.Y.; Padrón, C.R.; Vivas, M.; Barber, P.A.; Hardy, G.E.S.J.; Burgess, T.I. Extending the host range of Phytophthora multivora, a pathogen of woody plants in horticulture, nurseries, urban environments and natural ecosystems. Urban For. Urban Green. 2019, 46, 126460. [Google Scholar] [CrossRef]
- Biasi, A.; Martin, F.N.; Cacciola, S.O.; Magnano di San Lio, G.; Grünwald, N.J.; Schena, L. Genetic Analysis of Phytophthora nicotianae Populations from Different Hosts Using Microsatellite Markers. Phytopathology 2016, 106, 1006–1014. [Google Scholar] [CrossRef] [Green Version]
- Mammella, M.A.; Cacciola, S.O.; Martin, F.; Schena, L. Genetic characterization of Phytophthora nicotianae by the analysis of polymorphic regions of the mitochondrial DNA. Fungal Biol. 2011, 115, 432–442. [Google Scholar] [CrossRef]
- Mammella, M.A.; Martin, F.N.; Cacciola, S.O.; Coffey, M.D.; Faedda, R.; Schena, L. Analyses of the Population Structure in a Global Collection of Phytophthora nicotianae Isolates Inferred from Mitochondrial and Nuclear DNA Sequences. Phytopathology 2013, 103, 610–622. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Zhang, Q.; Ding, W.; Shan, W. Phytophthora parasitica: A model oomycete plant pathogen. Mycology 2014, 5, 43–51. [Google Scholar] [CrossRef]
- Panabières, F.; Ali, G.S.; Allagui, M.B.; Dalio, R.J.D.; Gudmestad, N.C.; Kuhn, M.L.; Guha Roy, S.; Schena, L.; Zampounis, A. Phytophthora nicotianae diseases worldwide: New knowledge of a long-recognised pathogen. Phytopathol. Mediterr. 2016, 55, 20–40. [Google Scholar]
- Abad, Z.G.; Abad, J.A.; Cacciola, S.O.; Pane, A.; Faedda, R.; Moralejo, E.; Pérez-Sierra, A.; Abad-Campos, P.; Alvarez-Bernaola, L.A.; Bakonyi, J.; et al. Phytophthora niederhauserii sp. nov., a polyphagous species associated with ornamentals, fruit trees and native plants in 13 countries. Mycologia 2014, 106, 431–447. [Google Scholar] [CrossRef]
- Rizzo, D.M.; Garbelotto, M.; Davidson, J.M.; Slaughter, G.W.; Koike, S.T. Phytophthora ramorum as the Cause of Extensive Mortality of Quercus spp. and Lithocarpus densiflorus in California. Plant Dis. 2002, 86, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, D.M.; Garbelotto, M.; Hansen, E.M. Phytophthora ramorum: Integrative research and management of an emerging pathogen in California and Oregon forests. Annu. Rev. Phytopathol. 2005, 43, 309–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grünwald, N.J.; Goss, E.M.; Press, C.M. Phytophthora ramorum: A pathogen with a remarkably wide host range causing sudden oak death on oaks and ramorum blight on woody ornamentals. Mol. Plant Pathol. 2008, 9, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Werres, S.; Marwitz, R.; In’T Veld, W.A.M.; De Cock, A.W.A.M.; Bonants, P.J.M.; De Weerdt, M.; Themann, K.; Ilieva, E.; Baayen, R.P. Phytophthora ramorum sp. nov., a new pathogen on Rhododendron and Viburnum. Mycol. Res. 2001, 105, 1155–1165. [Google Scholar] [CrossRef]
- Scanu, B.; Linaldeddu, B.T.; Peréz-Sierra, A.; Deidda, A.; Franceschini, A. Phytophthora ilicis as a leaf and stem pathogen of Ilex aquifolium in Mediterranean islands. Phytopathol. Mediterr. 2014, 53, 480–490. [Google Scholar]
- Ruano-Rosa, D.; Schena, L.; Agosteo, G.E.; di San Lio, G.M.; Cacciola, S.O. Phytophthora oleae sp. nov. causing fruit rot of olive in southern Italy. Plant Pathol. 2018, 67, 1362–1373. [Google Scholar] [CrossRef]
- Saville, A.C.; La Spada, F.; Faedda, R.; Migheli, Q.; Scanu, B.; Ermacora, P.; Gilardi, G.; Fedele, G.; Rossi, V.; Lenzi, N.; et al. Population structure of Phytophthora infestans collected on potato and tomato in Italy. Plant Pathol. 2021, 70, 2165–2178. [Google Scholar] [CrossRef]
- Santilli, E.; Riolo, M.; La Spada, F.; Pane, A.; Cacciola, S.O. First Report of Root Rot Caused by Phytophthora bilorbang on Olea europaea in Italy. Plants 2020, 9, 826. [Google Scholar] [CrossRef]
- Puglisi, I.; De Patrizio, A.; Schena, L.; Jung, T.; Evoli, M.; Pane, A.; Van Hoa, N.; Van Tri, M.; Wright, S.; Ramstedt, M.; et al. Two previously unknown Phytophthora species associated with brown rot of Pomelo (Citrus grandis) fruits in Vietnam. PLoS ONE 2017, 12, e0172085. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.S.J.; Barber, P.A.; Alvarado, P.; Barnes, C.W.; Buchanan, P.K.; Heykoop, M.; Moreno, G.; et al. Fungal Planet description sheets. Persoonia-Mol. Phylogeny Evol. Fungi 2017, 38, 558–624. [Google Scholar] [CrossRef]
- Cacciola, S.; La Spada, F.; Hoa, N.; Jung, M.; Scanu, B. Nomenclatural novelties. Index Fungorum 2018, 367, 1. [Google Scholar]
- Huai, W.X.; Tian, G.; Hansen, E.M.; Zhao, W.X.; Goheen, E.M.; Grünwald, N.J.; Cheng, C. Identification of Phytophthora species baited and isolated from forest soil and streams in northwestern Yunnan province, China. For. Pathol. 2013, 43, 87–103. [Google Scholar] [CrossRef]
- Pérez-Sierra, A.; López-García, C.; León, M.; García-Jiménez, J.; Abad-Campos, P.; Jung, T. Previously unrecorded low-temperature Phytophthora species associated with Quercus decline in a Mediterranean forest in eastern Spain. For. Pathol. 2013, 43, 331–339. [Google Scholar] [CrossRef]
- Scibetta, S.; Schena, L.; Chimento, A.; Cacciola, S.O.; Cooke, D.E.L. A molecular method to assess Phytophthora diversity in environmental samples. J. Microbiol. Methods 2012, 88, 356–368. [Google Scholar] [CrossRef]
- Jung, T.; Chang, T.T.; Bakonyi, J.; Seress, D.; Pérez-Sierra, A.; Yang, X.; Hong, C.; Scanu, B.; Fu, C.H.; Hsueh, K.L.; et al. Diversity of Phytophthora species in natural ecosystems of Taiwan and association with disease symptoms. Plant Pathol. 2017, 66, 194–211. [Google Scholar] [CrossRef]
- Riolo, M.; Aloi, F.; La Spada, F.; Sciandrello, S.; Moricca, S.; Santilli, E.; Pane, A.; Cacciola, S.O. Diversity of Phytophthora Communities across Different Types of Mediterranean Vegetation in a Nature Reserve Area. Forests 2020, 11, 853. [Google Scholar] [CrossRef]
- Scanu, B.; Vannini, A.; Franceschini, A.; Vettraino, A.M.; Ginetti, B.; Moricca, S. Phytophthora spp. in Mediterranean forests. In Proceedings of the Second International Congress of Silviculture, Florence, Italy, 26–29 November 2014; Ciancio, O., Ed.; Accademia Italiana di Scienze Forestali: Florence, Italy, 2014; pp. 402–407. [Google Scholar]
- Scanu, B.; Linaldeddu, B.T.; Deidda, A.; Jung, T. Diversity of Phytophthora Species from Declining Mediterranean Maquis Vegetation, including Two New Species, Phytophthora crassamura and P. ornamentata sp. nov. PLoS ONE 2015, 10, e0143234. [Google Scholar] [CrossRef] [Green Version]
- Simamora, A.V.; Paap, T.; Howard, K.; Stukely, M.J.C.; Hardy, G.E.S.J.; Burgess, T.I. Phytophthora Contamination in a Nursery and Its Potential Dispersal into the Natural Environment. Plant Dis. 2018, 102, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.; Gryzenhout, M.; Wingfield, B.D.; Wingfield, M.J.; Burgess, T.I. Surveys of soil and water reveal a goldmine of Phytophthora diversity in South African natural ecosystems. IMA Fungus 2013, 4, 123–131. [Google Scholar] [CrossRef]
- Rodríguez-Padrón, C.; Siverio, F.; Pérez-Sierra, A.; Rodríguez, A. Isolation and pathogenicity of Phytophthora species and Phytopythium vexans recovered from avocado orchards in the Canary Islands, including Phytophthora niederhauserii as a new pathogen of avocado. Phytopathol. Mediterr. 2018, 57, 89–106. [Google Scholar]
- Sarker, S.R.; McComb, J.; Burgess, T.I.; Hardy, G.E.S.J. Timing and abundance of sporangia production and zoospore release influences the recovery of different Phytophthora species by baiting. Fungal Biol. 2021, 125, 477–484. [Google Scholar] [CrossRef]
- Vannini, A.; Bruni, N.; Tomassini, A.; Franceschini, S.; Vettraino, A.M. Pyrosequencing of environmental soil samples reveals biodiversity of the Phytophthora resident community in chestnut forests. FEMS Microbiol. Ecol. 2013, 85, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Bose, T.; Wingfield, M.J.; Roux, J.; Vivas, M.; Burgess, T.I. Community composition and distribution of Phytophthora species across adjacent native and non-native forests of South Africa. Fungal Ecol. 2018, 36, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Cooke, D.E.L.; Williams, N.A.; Williamson, B.; Duncan, J.M. An Its-Based Phylogenetic Analysis of the Relationships between Peronospora and Phytophthora. In Advances in Downy Mildew Research; Springer: Dordrecht, The Netherlands, 2005; pp. 161–165. [Google Scholar]
- Davidson, J.M.; Werres, S.; Garbelotto, M.; Hansen, E.M.; Rizzo, D.M. Sudden Oak Death and Associated Diseases Caused by Phytophthora ramorum. Plant Health Prog. 2003, 4, 12. [Google Scholar] [CrossRef]
- Reeser, P.; Sutton, W.; Hansen, E. Phytophthora species in tanoak trees, canopy-drip, soil, and streams in the sudden oak death epidemic area of south-western Oregon, USA. N. Z. J. For. Sci. 2011, 41S, S65–S73. [Google Scholar]
- Schena, L.; Mosca, S.; Cacciola, S.O.; Faedda, R.; Sanzani, S.M.; Agosteo, G.E.; Sergeeva, V.; di San Lio, G.M. Species of the Colletotrichum gloeosporioides and C. boninense complexes associated with olive anthracnose. Plant Pathol. 2014, 63, 437–446. [Google Scholar] [CrossRef]
- Faedda, R.; Granata, G.; Cocuzza, G.E.M.; Lo Giudice, V.; Audoly, G.; Pane, A.; Cacciola, S.O. First report of heart rot of pomegranate (Punica granatum) caused by Alternaria alternata in Italy. Plant Dis. 2015, 99, 1446. [Google Scholar] [CrossRef]
- Luo, Y.; Hou, L.; Förster, H.; Pryor, B.; Adaskaveg, J.E. Identification of Alternaria Species Causing Heart Rot of Pomegranates in California. Plant Dis. 2017, 101, 421–427. [Google Scholar] [CrossRef] [Green Version]
- Talhinhas, P.; Loureiro, A.; Oliveira, H. Olive anthracnose: A yield- and oil quality-degrading disease caused by several species of Colletotrichum that differ in virulence, host preference and geographical distribution. Mol. Plant Pathol. 2018, 19, 1797–1807. [Google Scholar] [CrossRef] [Green Version]
- Cacciola, S.O.; Gilardi, G.; Faedda, R.; Schena, L.; Pane, A.; Garibaldi, A.; Gullino, M.L. Characterization of Colletotrichum ocimi Population Associated with Black Spot of Sweet Basil (Ocimum basilicum) in Northern Italy. Plants 2020, 9, 654. [Google Scholar] [CrossRef]
- Aloi, F.; Riolo, M.; Sanzani, S.M.; Mincuzzi, A.; Ippolito, A.; Siciliano, I.; Pane, A.; Gullino, M.L.; Cacciola, S.O. Characterization of Alternaria Species Associated with Heart Rot of Pomegranate Fruit. J. Fungi 2021, 7, 172. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Li Nicosia, M.G.D.; Cacciola, S.O.; Droby, S.; Schena, L. Metabarcoding Analysis of Fungal Diversity in the Phyllosphere and Carposphere of Olive (Olea europaea). PLoS ONE 2015, 10, e0131069. [Google Scholar] [CrossRef] [Green Version]
- Mendoza, M.L.Z.; Sicheritz-Pontén, T.; Thomas Gilbert, M.P. Environmental genes and genomes: Understanding the differences and challenges in the approaches and software for their analyses. Brief. Bioinform. 2015, 16, 745–758. [Google Scholar] [CrossRef] [Green Version]
- Prigigallo, M.I.; Abdelfattah, A.; Cacciola, S.O.; Faedda, R.; Sanzani, S.M.; Cooke, D.E.L.; Schena, L. Metabarcoding Analysis of Phytophthora Diversity Using Genus-Specific Primers and 454 Pyrosequencing. Phytopathology 2016, 106, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Green, S.; Cooke, D.E.L.; Dunn, M.; Barwell, L.; Purse, B.; Chapman, D.S.; Valatin, G.; Schlenzig, A.; Barbrook, J.; Pettitt, T.; et al. Phyto-threats: Addressing threats to uk forests and woodlands from Phytophthora; identifying risks of spread in trade and methods for mitigation. Forests 2021, 12, 1617. [Google Scholar] [CrossRef]
- Prigigallo, M.I.; Mosca, S.; Cacciola, S.O.; Cooke, D.E.L.; Schena, L. Molecular analysis of Phytophthora diversity in nursery-grown ornamental and fruit plants. Plant Pathol. 2015, 64, 1308–1319. [Google Scholar] [CrossRef] [Green Version]
- Burgess, T.I.; White, D.; McDougall, K.M.; Garnas, J.; Dunstan, W.A.; Català, S.; Carnegie, A.J.; Worboys, S.; Cahill, D.; Vettraino, A.M.; et al. Distribution and diversity of Phytophthora across Australia. Pac. Conserv. Biol. 2017, 23, 150–162. [Google Scholar] [CrossRef] [Green Version]
- Català, S.; Pérez-Sierra, A.; Abad-Campos, P. The Use of Genus-Specific Amplicon Pyrosequencing to Assess Phytophthora Species Diversity Using eDNA from Soil and Water in Northern Spain. PLoS ONE 2015, 10, e0119311. [Google Scholar] [CrossRef] [Green Version]
- Català, S.; Berbegal, M.; Pérez-Sierra, A.; Abad-Campos, P. Metabarcoding and development of new real-time specific assays reveal Phytophthora species diversity in holm oak forests in eastern Spain. Plant Pathol. 2017, 66, 115–123. [Google Scholar] [CrossRef]
- Khaliq, I.; St. Hardy, G.E.J.; White, D.; Burgess, T.I. eDNA from roots: A robust tool for determining Phytophthora communities in natural ecosystems. FEMS Microbiol. Ecol. 2018, 94, fiy048. [Google Scholar] [CrossRef] [Green Version]
- Ruiz Gómez, F.J.; Navarro-Cerrillo, R.M.; Pérez-de-Luque, A.; Oβwald, W.; Vannini, A.; Morales-Rodríguez, C. Assessment of functional and structural changes of soil fungal and oomycete communities in holm oak declined dehesas through metabarcoding analysis. Sci. Rep. 2019, 9, 5315. [Google Scholar] [CrossRef] [Green Version]
- Chimento, A.; Cacciola, S.O.; Garbelotto, M. Detection of mRNA by reverse-transcription PCR as an indicator of viability in Phytophthora ramorum. For. Pathol. 2012, 42, 14–21. [Google Scholar] [CrossRef]
- Cullen, D.W.; Lees, A.K.; Toth, I.K.; Duncan, J.M. Conventional PCR and real-time quantitative PCR detection of Helminthosporium solani in soil and on potato tubers. Eur. J. Plant Pathol. 2001, 107, 387–398. [Google Scholar] [CrossRef]
- Cooke, D.E.L.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A Molecular Phylogeny of Phytophthora and Related Oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Illumina Inc. 16S Metagenomic Sequencing Library Preparation—Preparing 16S Ribosomal RNA Gene Amplicons for the Illumina MiSeq System; Illumina Inc.: San Diego, CA, USA, 2013. [Google Scholar]
- peterjc/thapbi-pict: THAPBI PICT v0.6.1|Zenodo. Available online: https://zenodo.org/record/6022960#.YjijiqjSLIW (accessed on 21 March 2022).
- Safaiefarahani, B.; Mostowfizadeh-Ghalamfarsa, R.; Hardy, G.E.S.J.; Burgess, T.I. Re-evaluation of the Phytophthora cryptogea species complex and the description of a new species, Phytophthora pseudocryptogea sp. nov. Mycol. Prog. 2015, 14, 108. [Google Scholar] [CrossRef]
- Scanu, B.; Hunter, G.C.; Linaldeddu, B.T.; Franceschini, A.; Maddau, L.; Jung, T.; Denman, S. A taxonomic re-evaluation reveals that Phytophthora cinnamomi and P. cinnamomi var. parvispora are separate species. For. Pathol. 2014, 44, 1–20. [Google Scholar] [CrossRef]
- Aghighi, S.; Hardy, G.E.S.J.; Scott, J.K.; Burgess, T.I. Phytophthora bilorbang sp. nov., a new species associated with the decline of Rubus anglocandicans (European blackberry) in Western Australia. Eur. J. Plant Pathol. 2012, 133, 841–855. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- MEGA—Molecular Evolutionary Genetics Analysis. Available online: https://www.megasoftware.net/ (accessed on 21 March 2022).
- R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 21 March 2022).
- Aloi, F.; Riolo, M.; La Spada, F.; Bentivenga, G.; Moricca, S.; Santilli, E.; Pane, A.; Faedda, R.; Cacciola, S.O. Phytophthora Root and Collar Rot of Paulownia, a New Disease for Europe. Forests 2021, 12, 1664. [Google Scholar] [CrossRef]
- Jung, T.; Burgess, T.I. Re-evaluation of Phytophthora citricola isolates from multiple woody hosts in Europe and North America reveals a new species, Phytophthora plurivora sp. nov. Persoonia 2009, 22, 95–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasier, C.M.; Cooke, D.E.L.; Duncan, J.M.; Hansen, E.M. Multiple new phenotypic taxa from trees and riparian ecosystems in Phytophthora gonapodyides-P. megasperma ITS Clade 6, which tend to be high-temperature tolerant and either inbreeding or sterile. Mycol. Res. 2003, 107, 277–290. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- FinchTV|Digital World Biology. Available online: https://digitalworldbiology.com/FinchTV (accessed on 21 March 2022).
- BLAST: Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 21 March 2022).
- GenBank. Available online: https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 21 March 2022).
- In’t Veld, W.A.M.; Rosendahl, K.C.H.M.; Van Rijswick, P.C.J.; Meffert, J.P.; Westenberg, M.; Van De Vossenberg, B.T.L.H.; Denton, G.; Van Kuik, F.A.J. Phytophthora terminalis sp. nov. and Phytophthora occultans sp. nov., two invasive pathogens of ornamental plants in Europe. Mycologia 2015, 107, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Vettraino, A.M.; Roques, A.; Yart, A.; Fan, J.T.; Sun, J.H.; Vannini, A. Sentinel trees as a tool to forecast invasions of alien plant pathogens. PLoS ONE 2015, 10, e0120571. [Google Scholar] [CrossRef] [PubMed]
- Riolo, M.; La Spada, F.; Aloi, F.; del Galdo, G.G.; Santilli, E.; Pane, A.; Cacciola, S.O. Phytophthora Diversity in a Sentinel Arboretum and in a Nature Reserve Area. Biol. Life Sci. Forum 2020, 4, 51. [Google Scholar]
- Riolo, M.; La Spada, F.; Aloi, F.; del Galdo, G.G.; Santilli, E.; Pane, A.; Cacciola, S.O. Phytophthora Diversity in Two Different Types of Plant Conservation Sites. In Proceedings of the IECPS 2020—The 1st International Electronic Conference on Plant Science, Online, 1–15 December 2020; pp. 1–12. [Google Scholar]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Casal, O.A.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Pathol. 2016, 46, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Maseko, B.; Burgess, T.I.; Coutinho, T.A.; Wingfield, M.J. Two new Phytophthora species from South African Eucalyptus plantations. Mycol. Res. 2007, 111, 1321–1338. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Hansen, E.M.; Winton, L.; Oswald, W.; Delatour, C. Three new species of Phytophthora from European oak forests. Mycol. Res. 2002, 106, 397–411. [Google Scholar] [CrossRef]
- Frankel, S.J.; Conforti, C.; Hillman, J.; Ingolia, M.; Shor, A.; Benner, D.; Alexander, J.M.; Bernhardt, E.; Swiecki, T.J. Phytophthora Introductions in Restoration Areas: Responding to Protect California Native Flora from Human-Assisted Pathogen Spread. Forests 2020, 11, 1291. [Google Scholar] [CrossRef]
- Migliorini, D.; Ghelardini, L.; Tondini, E.; Luchi, N.; Santini, A. The potential of symptomless potted plants for carrying invasive soilborne plant pathogens. Divers. Distrib. 2015, 21, 1218–1229. [Google Scholar] [CrossRef]
- Sims, L.; Tjosvold, S.; Chambers, D.; Garbelotto, M. Control of Phytophthora species in plant stock for habitat restoration through best management practices. Plant Pathol. 2019, 68, 196–204. [Google Scholar] [CrossRef] [Green Version]
- Cacciola, S.O.; di San Lio, G.M.; Belisario, A. Phytophthora italica sp. nov. on myrtle. Phytopathol. Mediterr. 1996, 35, 177–190. [Google Scholar]
- Jung, T.; Cooke, D.E.L.; Blaschke, H.; Duncan, J.M.; Oßwald, W. Phytophthora quercina sp. nov., causing root rot of European oaks. Mycol. Res. 1999, 103, 785–798. [Google Scholar] [CrossRef]
- Martin, F.N.; Abad, G.Z.; Balci, Y.; Ivors, K. Identification and Detection of Phytophthora: Reviewing Our Progress, Identifying Our Needs. Plant Dis. 2012, 96, 1080–1103. [Google Scholar] [CrossRef] [Green Version]
- Bertier, L.; Brouwer, H.; de Cock, A.W.A.M.; Cooke, D.E.L.; Olsson, C.H.B.; Höfte, M. The expansion of Phytophthora clade 8b: Three new species associated with winter grown vegetable crops. Persoonia Mol. Phylogeny Evol. Fungi 2013, 31, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Tyler, B.M.; Hong, C. An expanded phylogeny for the genus Phytophthora. IMA Fungus 2017, 8, 355–384. [Google Scholar] [CrossRef] [Green Version]
- Kroon, L.P.N.M.; Brouwer, H.; de Cock, A.W.A.M.; Govers, F. The Genus Phytophthora Anno 2012. Phytopathology 2012, 102, 348–364. [Google Scholar] [CrossRef] [Green Version]
- Bose, T.; Hulbert, J.M.; Burgess, T.I.; Paap, T.; Roets, F.; Wingfield, M.J. Two novel Phytophthora species from the southern tip of Africa. Mycol. Prog. 2021, 20, 755–767. [Google Scholar] [CrossRef]
- Ginetti, B.; Moricca, S.; Squires, J.N.; Cooke, D.E.L.; Ragazzi, A.; Jung, T. Phytophthora acerina sp. nov., a new species causing bleeding cankers and dieback of Acer pseudoplatanus trees in planted forests in northern Italy. Plant Pathol. 2014, 63, 858–876. [Google Scholar] [CrossRef] [Green Version]
- Burgess, T.I.; López-Villamor, A.; Paap, T.; Williams, B.; Belhaj, R.; Crone, M.; Dunstan, W.; Howard, K.; Hardy, G.E.S.J. Towards a best practice methodology for the detection of Phytophthora species in soils. Plant Pathol. 2021, 70, 604–614. [Google Scholar] [CrossRef]
- Linzer, R.E.; Rizzo, D.M.; Cacciola, S.O.; Garbelotto, M. AFLPs detect low genetic diversity for Phytophthora nemorosa and P. pseudosyringae in the US and Europe. Mycol. Res. 2009, 113, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Biasi, A.; Martin, F.; Schena, L. Identification and validation of polymorphic microsatellite loci for the analysis of Phytophthora nicotianae populations. J. Microbiol. Methods 2015, 110, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Schena, L.; Hughes, K.J.D.; Cooke, D.E.L. Detection and quantification of Phytophthora ramorum, P. kernoviae, P. citricola and P. quercina in symptomatic leaves by multiplex real-time PCR. Mol. Plant Pathol. 2006, 7, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Schena, L.; Abdelfattah, A.; Mosca, S.; Li Nicosia, M.G.D.; Agosteo, G.E.; Cacciola, S.O. Quantitative detection of Colletotrichum godetiae and C. acutatum sensu stricto in the phyllosphere and carposphere of olive during four phenological phases. Eur. J. Plant Pathol. 2017, 149, 337–347. [Google Scholar] [CrossRef]
- ITS1 Metabarcoding Revealing Phytophthora Diversity in Anthropized and Natural Ecosystems in Sicily, Italy | Zenodo. Available online: https://zenodo.org/record/6044741#.YjiywKjSLIU (accessed on 21 March 2022).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Area | Rhizosphere Soil Sample ID. | Location-Country and Geographic Coordinates (DATUM WGS84) | Sampled Tree Species (Baiting/DNA Metabarcoding Phytophthora-Positive (+) or Negative (−)) | Baited Phytophthora Taxa 1 | DNA Metabarcoding Detected Phytophthora Taxa 1 | Phytophthora Spp. (Baiting + DNA Metabarcoding) 1 |
|---|---|---|---|---|---|---|
| Complesso Speleologico Villasmundo S. Alfio Regional Nature Reserve | NR_1903_S1 | Melilli-37°13′17.54″ N; 15°6′19.52″ E | Salix pedicellata (+/−) | PSC | - | PSC |
| NR_1903_S2 | Melilli-37°13′17.66″ N; 15°6′19.28″ E | S. pedicellata (+/+) | CRY | PSY (r) 2, UNK 3 (r), UNK 5 (r) | CRY, PSY (r), UNK 3 (r), UNK 5 (r) | |
| NR_1903_S3 | Melilli-37°13′17.753″ N; 15°6′18.93″ E | Platanus orientalis (−/−) | - | - | - | |
| NR_1903_S4 | Melilli-37°13′17.86″ N; 15°6′18.81″ E | P. orientalis (+/+) | BIL | PSY (r), UNK 1 (r) | BIL, PSY (r), UNK 1 (r) | |
| NR_1903_S5 | Melilli-37°13′17.25″ N; 15°6′15.30″ E | Euphorbia dendroides (−/−) | - | - | - | |
| NR_1903_S6 | Melilli-37°13′17.48″ N; 15°6′15.31″ E | Cynara cardunculus (−/+) | - | IRA-like (r), PSY (r) | IRA-like (r), PSY (r) | |
| NR_1903_S7 | Melilli-37°13′17.60″ N; 15°6′15.30″ E | Asphodelus sp. (−/−) | - | - | - | |
| NR_1903_S8 | Melilli-37°13′11.75″ N; 15°6′1.20″ E | Quercus ilex (+/+) | GON | CAS/QUE (s) 2, PSY (s), SYR (r) | GON, CAS/QUE (s), PSY (s), SYR (r) | |
| NR_1903_S9 | Melilli-37°13′11.00″ N; 15°5′59.69″ E | Q. ilex (+/+) | PLU | PSY (s) | PLU, PSY (s) | |
| NR_1903_S10 | Melilli-37°13′10.93″ N; 15°5′59.95″ E | Q. ilex (−/−) | - | - | - | |
| NR_1903_S11 | Melilli-37°13′11.788″ N; 15°6′0.547″ E | Q. pubescens sensu latu (+/) | GON | PLU (r), PSY (r) | GON, PLU (r), PSY (r) | |
| NR_1903_S12 | Melilli-37°13′17.52″ N; 15°6′7.94″ E | Sarcopoterium spinosum (−/−) | - | - | - | |
| NR_1903_S13 | Melilli-37°13′17.50″ N; 15°6′8.57″ E | S. spinosum (−/−) | - | - | - | |
| NR_1903_S14 | Melilli-37°13′17.28″ N; 15°6′4.77″ E | Pistacia lentiscus (−/+) | - | SYR (r), UNK 1 (r) | SYR (r), UNK 1 (r) | |
| NR_1903_S15 | Melilli-37°13′17.50″ N; 15°6′5.13″ E | P. lentiscus and Pyrus sp., mixed sample (−/−) | - | - | - | |
| NR_1903_S16 | Melilli-37°13′16.94″ N; 15°6′7.66″ E | P. lentiscus (−/+) | - | ALE/CAC (r), ASP (r), SYR (r), UNK 1 (r) | ALE/CAC (r), ASP (r), SYR (r), UNK 1 (r) | |
| NR_1903_S17 | Melilli-37°13′16.93″ N; 15°6′6.24″ E | P. lentiscus (−/+) | - | ALE/CAC (r), SYR (r), UNK 1 (r) | ALE/CAC (r), SYR (r), UNK 1 (r) | |
| Botanical garden of Catania | BG_1903_S1 | Catania-37°30′57.29″ N; 15°5′2.27″ E | Araucaria cookii (+/−) | NIC, MUL | - | NIC, MUL |
| BG_1903_S2 | Catania-37°30′55.92″ N; 15°5′1.95″ E | Phytolacca dioica (+/−) | NIC, MUL | - | NIC, MUL | |
| BG_1903_S3 | Catania-37°30′55.08″ N; 15°4′59.75″ E | Grevillea robusta (−/+) | - | MUL (r) | MUL (r) | |
| BG_1903_S4 | Catania-37°30′57.56″ N; 15°5′1.47″ E | Pistacia atlantica (−/+) | - | MUL (s), SYR (s) | MUL (s), SYR (s) | |
| BG_1903_S5 | Catania-37°30′57.47″ N; 15°5′0.81″ E | Sterculia diversifolia (+/−) | MUL | - | MUL | |
| BG_1903_S6 | Catania-37°30′57.69″ N; 15°5′1.80″ E | Eucalyptus citridora (−/+) | - | NIC (r), MUL (r), SYR (r) | NIC (r), MUL (r), SYR (r) | |
| BG_1903_S7 | Catania-37°30′53.46″ N; 15°5′2.38″ E | Zelkowa sicula (+/+) | MUL | NIC (r) | MUL, NIC (r) | |
| BG_1903_S8 | Catania-37°30′53.35″ N; 15°5′1.89″ E | Q. suber (+/+) | NIC, MUL | SYR (r) | NIC, MUL, SYR (r) | |
| BG_1903_S9 | Catania-37°30′53.19″ N; 15°5′2.42″ E | Olea europea (+/+) | MUL, NIC | MUL (r) | NIC, MUL, MUL (r) | |
| BG_1903_S10 | Catania-37°30′53.34″ N; 15°5′2.40″ E | Pistacia lentiscus (+/) | - | MUL (r), SYR (r) | MUL (r), SYR (r) | |
| BG_1903_S11 | Catania-37°30′57.92″ N; 15°5′0.74″ E | Coffea arabica (+/−) | PAR | - | PAR | |
| BG_1903_S12 | Catania-37°30′57.95″ N; 15°5′0.86″ E | Mangifera indica (−/−) | - | - | - | |
| Citrus orchard-Tenuta Serravalle | CO_1905_S1 | Mineo-37°19′39.38″ N; 14°41′10.65″ E | Citrus × sinensis ′Tarocco′ nested on C. × aurantium (−/+) | - | - | - |
| CO_1905_S2 | Mineo-37°19′39.38″ N; 14°41′10.65″ E | //(−/+) | - | NIC (r) | NIC (r) | |
| CO_1905_S3 | Mineo-37°19′40.29″ N; 14°41′11.05″ E | //(+/−) | NIC, CIP | - | NIC, CIP | |
| CO_1905_S4 | Mineo-37°19′40.54″ N; 14°41′12.78″ E | //(+/+) | NIC | UNK 2 (r), UNK 4 (r) | NIC, UNK 2 (r), UNK 4 (r) | |
| CO_1905_S5 | Mineo-37°19′41.35″ N; 14°41′12.90″ E | //(+/+) | NIC | CIP (r), OCC (r) | NIC, CIP (r), OCC (r) | |
| Citrus orchard-Tenuta Serravalle | CO_1905_S6 | Mineo-37°19′39.36″ N; 14°41′7.75″ E | //(+/−) | NIC, CIP | - | NIC, CIP |
| CO_1905_S7 | Mineo-37°19′41.05″ N; 14°41′6.45″ E | //(+/−) | NIC | - | NIC | |
| CO_1905_S8 | Mineo-37°19′41.69″ N; 14°41′7.73″ E | //(+/−) | NIC | - | NIC | |
| CO_1905_S9 | Mineo-37°19′41.97″ N; 14°41′9.30″ E | //(−/−) | - | - | - | |
| CO_1905_S10 | Mineo-7°19′43.16″ N; 14°41′7.12″ E | //(−/−) | - | - | - |
| Sampling Area | |||
|---|---|---|---|
| Diversity Index | Nature Reserve | Botanical Garden | Citrus Orchard |
| Shannon diversity | 0.674 a | 0.267 b | 0.436 ab |
| Pielou’s evenness | 0.29 a | 0.24 b | 0.27 ab |
| Simpson dominance | 0.57 a | 0.82 b | 0.69 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
La Spada, F.; Cock, P.J.A.; Randall, E.; Pane, A.; Cooke, D.E.L.; Cacciola, S.O. DNA Metabarcoding and Isolation by Baiting Complement Each Other in Revealing Phytophthora Diversity in Anthropized and Natural Ecosystems. J. Fungi 2022, 8, 330. https://doi.org/10.3390/jof8040330
La Spada F, Cock PJA, Randall E, Pane A, Cooke DEL, Cacciola SO. DNA Metabarcoding and Isolation by Baiting Complement Each Other in Revealing Phytophthora Diversity in Anthropized and Natural Ecosystems. Journal of Fungi. 2022; 8(4):330. https://doi.org/10.3390/jof8040330
Chicago/Turabian StyleLa Spada, Federico, Peter J. A. Cock, Eva Randall, Antonella Pane, David E. L. Cooke, and Santa Olga Cacciola. 2022. "DNA Metabarcoding and Isolation by Baiting Complement Each Other in Revealing Phytophthora Diversity in Anthropized and Natural Ecosystems" Journal of Fungi 8, no. 4: 330. https://doi.org/10.3390/jof8040330
APA StyleLa Spada, F., Cock, P. J. A., Randall, E., Pane, A., Cooke, D. E. L., & Cacciola, S. O. (2022). DNA Metabarcoding and Isolation by Baiting Complement Each Other in Revealing Phytophthora Diversity in Anthropized and Natural Ecosystems. Journal of Fungi, 8(4), 330. https://doi.org/10.3390/jof8040330