
A Kinesin Vdkin2 Required for Vacuole Formation, Mycelium Growth, and Penetration Structure Formation of Verticillium dahliae

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Cultivation
2.2. Vector Construction and Fungal Transformation
2.3. Pathogenicity Analysis
2.4. Root Colonization Analysis
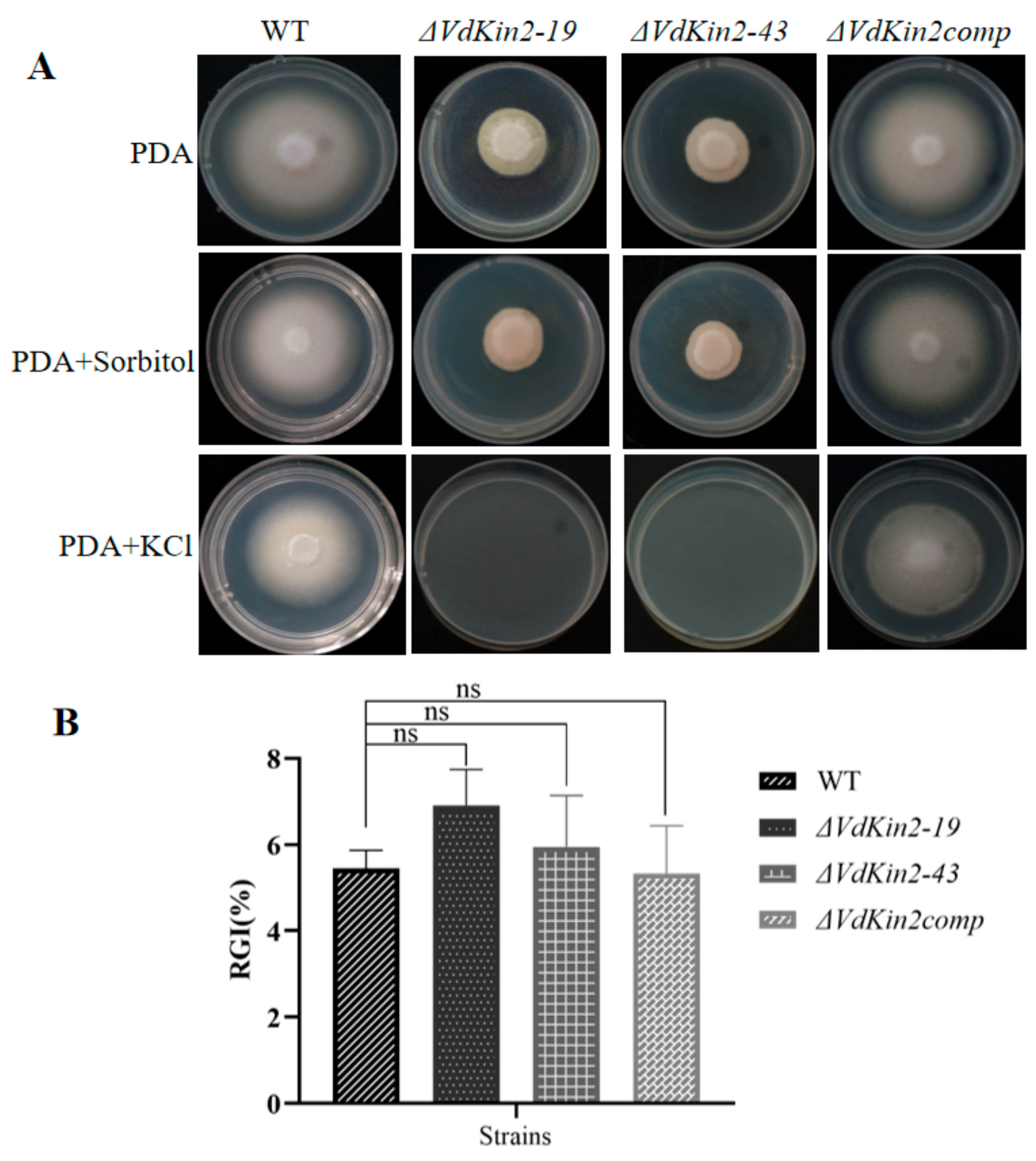
2.5. Salt-Stress Response
2.6. Microsclerotial Formation Assays
2.7. Penetration Assays
2.8. Hyphal Septa, Vacuoles, and Lipid Droplet Staining
2.9. RNA-Seq
2.10. RT-PCR Assay
3. Results
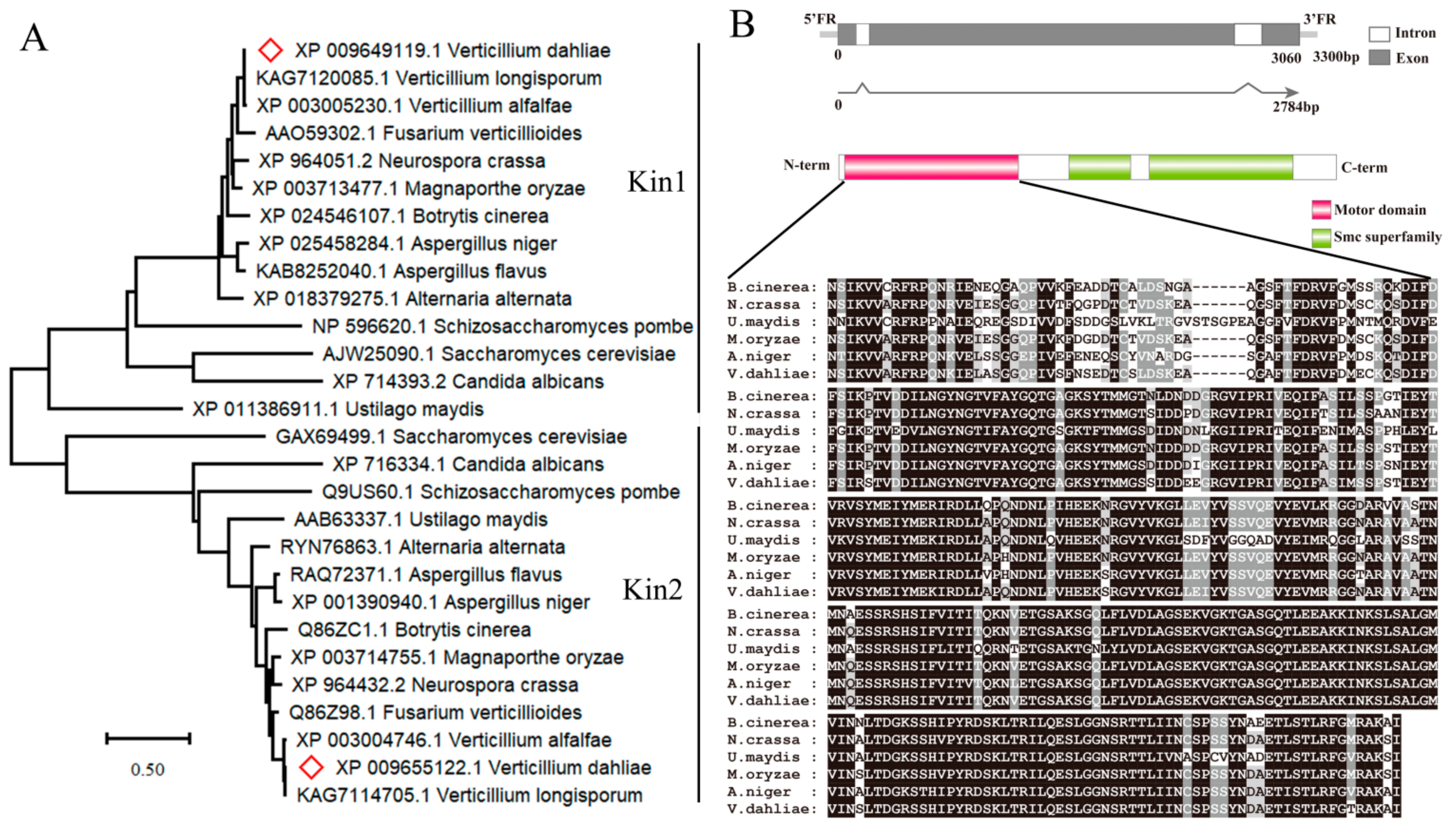
3.1. Identification of Vdkin2
3.2. Vdkin2 Is Required for Virulence in Cotton
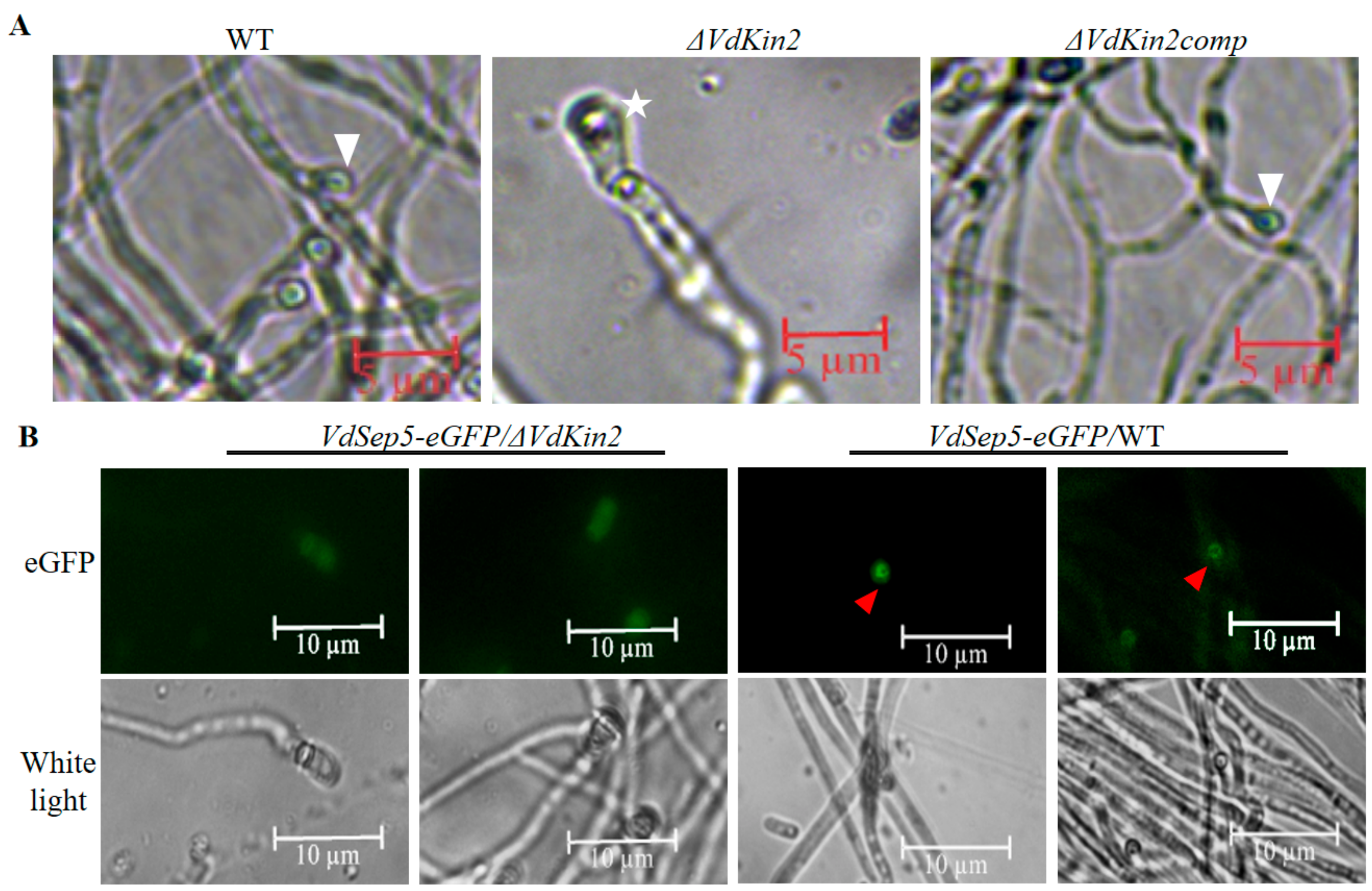
3.3. Deletion of Vdkin2 Impairs Penetration of V. dahliae
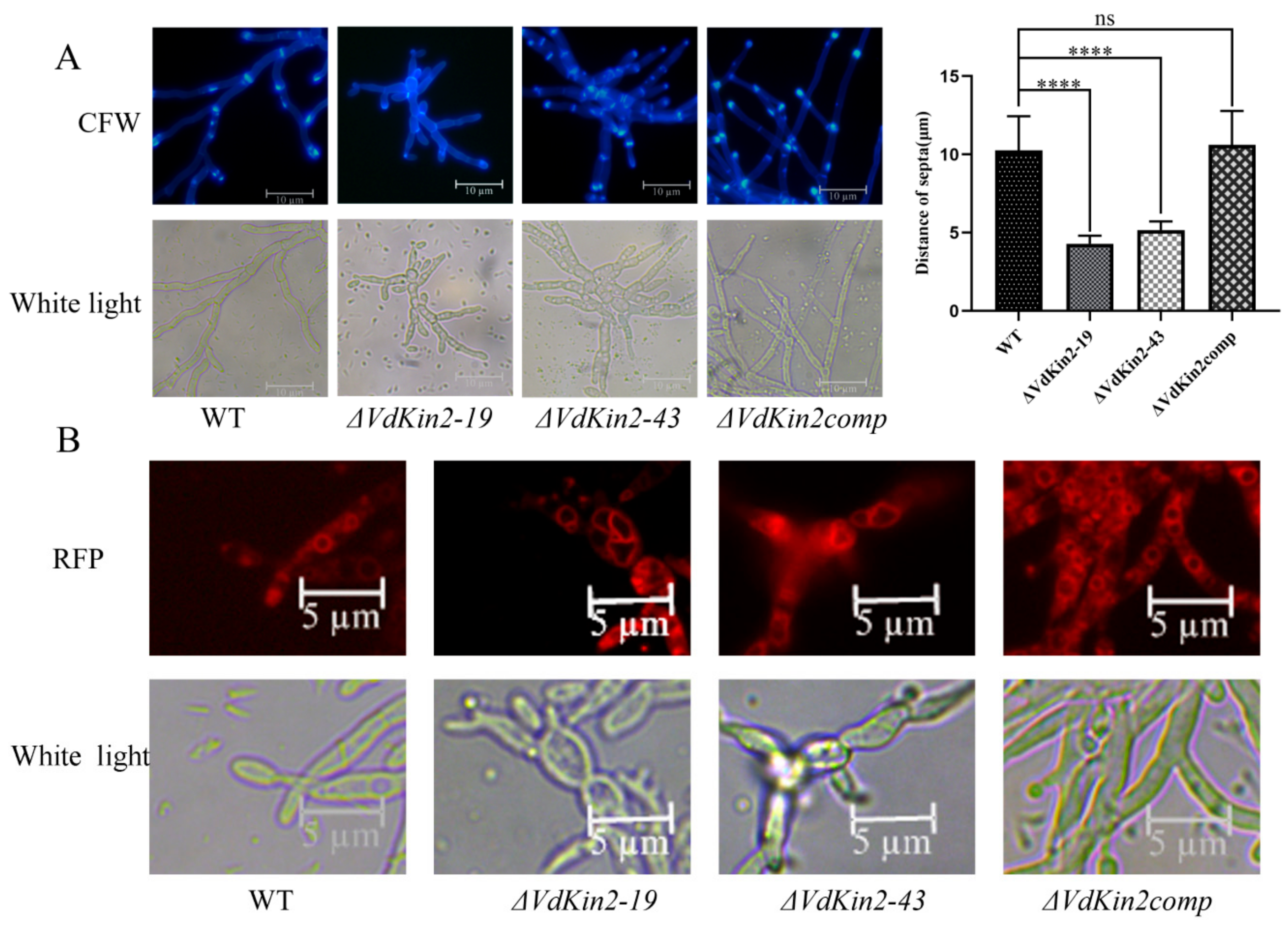
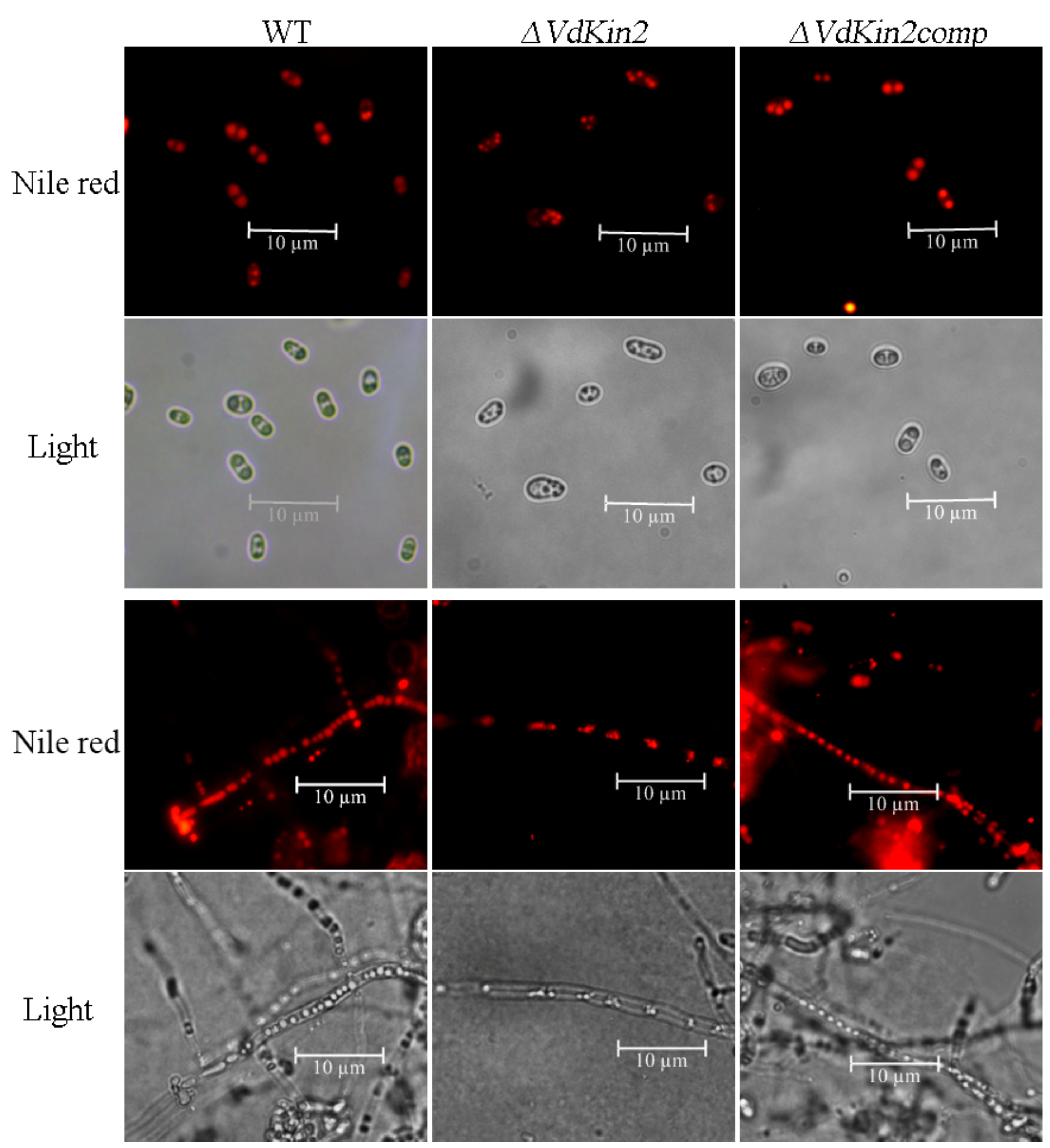
3.4. VdKin2 Deletion Leads to Aberrant Hyphal Growth, Reduced Conidiation, and Abnormal Vacuole Morphology
3.5. VdKin2 Deletion Affects Lipid Storage in the Vacuoles
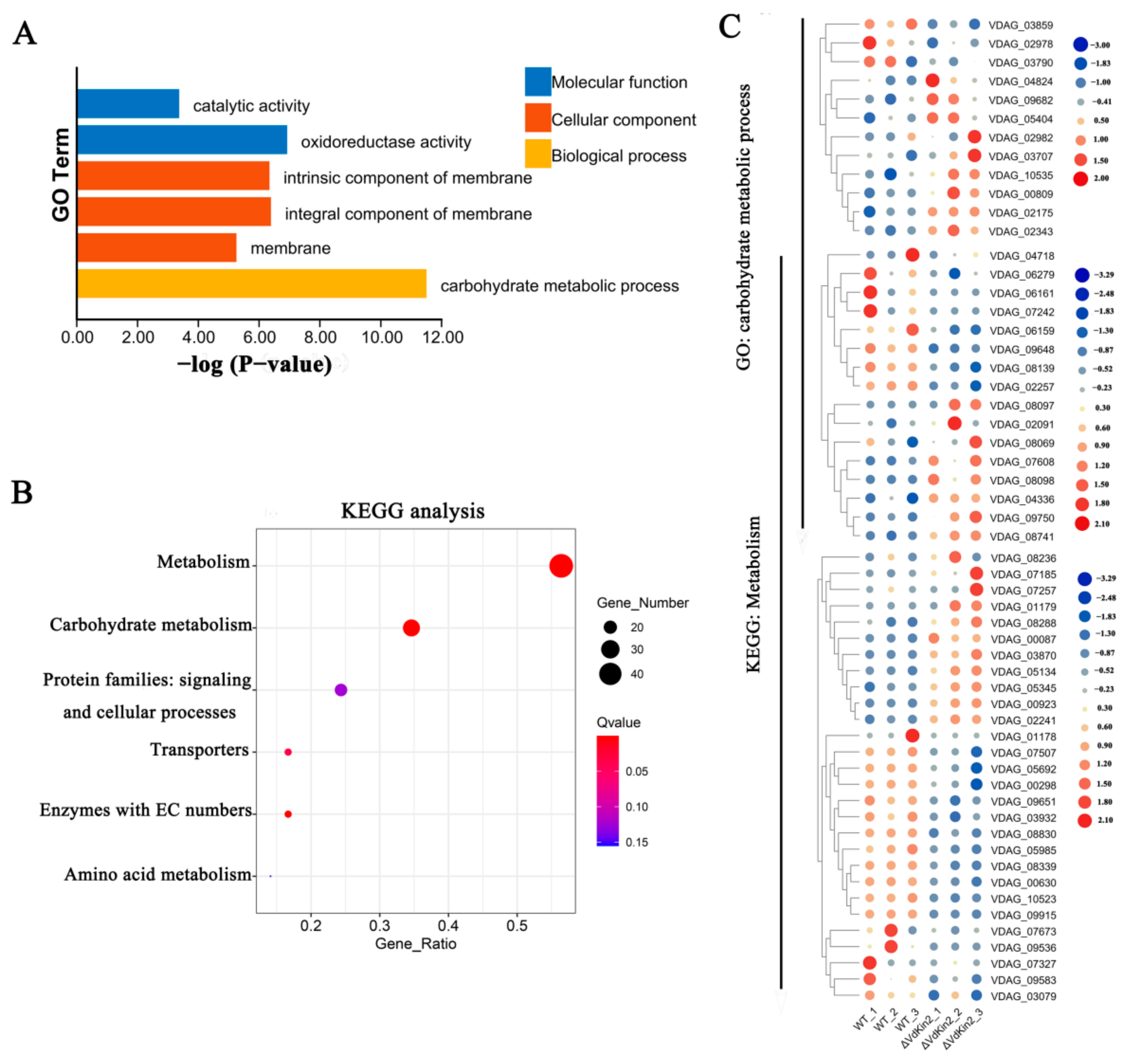
3.6. RNA-Sequencing Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Tang, C.; Li, T.Y.; Klosterman, S.J.; Tian, C.M.; Wang, Y.L. The bZIP transcription factor VdAtf1 regulates virulence by mediating nitrogen metabolism in Verticillium dahliae. New Phytol. 2020, 226, 1461–1479. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Jin, X.; Klosterman, S.J.; Wang, Y. Convergent and distinctive functions of transcription factors VdYap1, VdAtf1, and VdSkn7 in the regulation of nitrosative stress resistance, microsclerotia formation, and virulence in Verticillium dahliae. Mol. Plant Pathol. 2020, 21, 1451–1466. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-L.; Zhou, T.-T.; Guo, H.-S. Hyphopodium-Specific VdNoxB/VdPls1-Dependent ROS-Ca2+ Signaling Is Required for Plant Infection by Verticillium dahliae. PLoS Pathog. 2016, 12, e1005793. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.-T.; Zhao, Y.-L.; Guo, H.-S. Secretory proteins are delivered to the septin-organized penetration interface during root infection by Verticillium dahliae. PLoS Pathog. 2017, 13, e1006275. [Google Scholar] [CrossRef]
- Qin, J.; Wang, K.; Sun, L.; Xing, H.; Wang, S.; Li, L.; Chen, S.; Guo, H.-S.; Zhang, J. The plant-specific transcription factors CBP60g and SARD1 are targeted by a Verticillium secretory protein VdSCP41 to modulate immunity. eLife 2018, 7, e34902. [Google Scholar] [CrossRef]
- Jenkinson, C.B.; Jones, K.; Zhu, J.; Dorhmi, S.; Khang, C.H. The appressorium of the rice blast fungus Magnaporthe oryzae remains mitotically active during post-penetration hyphal growth. Fungal Genet. Biol. 2017, 98, 35–38. [Google Scholar] [CrossRef]
- Wang, X.L.; Lu, D.X.; Tian, C.M. CgEnd3 Regulates Endocytosis, Appressorium Formation, and Virulence in the Poplar Anthracnose Fungus Colletotrichum gloeosporioides. Int. J. Mol. Sci. 2021, 22, 4029. [Google Scholar] [CrossRef]
- Richards, A.; Veses, V.; Gow, N.A.R. Vacuole dynamics in fungi. Fungal Biol. Rev. 2010, 24, 93–105. [Google Scholar] [CrossRef]
- Weber, R.W.S. Vacuoles and the fungal lifestyle. Mycologist 2002, 16, 10–20. [Google Scholar] [CrossRef]
- Veses, V.; Richards, A.; Gow, N.A.R. Vacuoles and fungal biology. Curr. Opin. Microbiol. 2008, 11, 503–510. [Google Scholar] [CrossRef]
- Veneault-Fourrey, C.; Barooah, M.; Egan, M.; Wakley, G.; Talbot, N.J. Autophagic Fungal Cell Death Is Necessary for Infection by the Rice Blast Fungus. Science 2006, 312, 580–583. [Google Scholar] [CrossRef]
- Luna-Tapia, A.; DeJarnette, C.; Sansevere, E.; Reitler, P.; Butts, A.; Hevener, K.E.; Palmer, G.E. The Vacuolar Ca2+ ATPase Pump Pmc1p Is Required for Candida albicans Pathogenesis. mSphere 2019, 4, e00715-18. [Google Scholar] [CrossRef]
- Yu, Q.; Wang, F.; Zhao, Q.; Chen, J.; Zhang, B.; Ding, X.; Wang, H.; Yang, B.; Lu, G.; Zhang, B.; et al. A novel role of the vacuolar calcium channel Yvc1 in stress response, morphogenesis and pathogenicity of Candida albicans. Int. J. Med Microbiol. 2013, 304, 339–350. [Google Scholar] [CrossRef]
- Takano, Y.; Kikuchi, T.; Kubo, Y.; Hamer, J.E.; Mise, K.; Furusawa, I. The Colletotrichum lagenarium MAP Kinase Gene CMK1 Regulates Diverse Aspects of Fungal Pathogenesis. Mol. Plant-Microbe Interact. 2000, 13, 374–383. [Google Scholar] [CrossRef]
- Park, G.; Bruno, K.S.; Staiger, C.J.; Talbot, N.J.; Xu, J.-R. Independent genetic mechanisms mediate turgor generation and penetration peg formation during plant infection in the rice blast fungus. Mol. Microbiol. 2004, 53, 1695–1707. [Google Scholar] [CrossRef]
- Park, G.; Xue, C.; Zheng, L.; Lam, S.; Xu, J.-R. MST12 Regulates Infectious Growth but Not Appressorium Formation in the Rice Blast Fungus Magnaporthe grisea. Mol. Plant-Microbe Interact. 2002, 15, 183–192. [Google Scholar] [CrossRef]
- Kallipolitou, A.; DeLuca, D.; Majdic, U.; Lakämper, S.; Cross, R.; Meyhofer, E.; Moroder, L.; Schliwa, M.; Woehlke, G. Unusual properties of the fungal conventional kinesin neck domain from Neurospora crassa. EMBO J. 2001, 20, 6226–6235. [Google Scholar] [CrossRef]
- Steinberg, G.; Schliwa, M.; Lehmler, C.; Bolker, M.; Kahmann, R.; McIntosh, J.R. Kinesin from the plant pathogenic fungus Ustilago maydis is involved in vacuole formation and cytoplasmic migration. J. Cell Sci. 1998, 111, 2235–2246. [Google Scholar] [CrossRef]
- Lehmler, C.; Steinberg, G.; Snetselaar, K.M.; Schliwa, M.; Kahmann, R.; Bölker, M. Identification of a motor protein required for filamentous growth in Ustilago maydis. EMBO J. 1997, 16, 3464–3473. [Google Scholar] [CrossRef]
- Xie, C.J.; Li, Q.L.; Yang, X.Y. Characterization of VdASP F2 Secretory Factor from Verticillium dahliae by a Fast and Easy Gene Knockout System. Mol. Plant-Microbe Interact. 2017, 30, 444–454. [Google Scholar] [CrossRef]
- Santhanam, P.; Thomma, B.P.H.J. Verticillium dahliae Sge1 Differentially Regulates Expression of Candidate Effector Genes. Mol. Plant-Microbe Interact. 2013, 26, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Fradin, E.F.; Zhang, Z.; Ayala, J.C.J.; Castroverde, C.D.M.; Nazar, R.N.; Robb, J.; Liu, C.-M.; Thomma, B.P.H.J. Genetic Dissection of Verticillium Wilt Resistance Mediated by Tomato Ve1. Plant Physiol. 2009, 150, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Z.-K.; Sun, H.-H.; Ying, S.-H.; Feng, M.-G. Characterization of the Hog1 MAPK pathway in the entomopathogenic fungus Beauveria bassiana. Environ. Microbiol. 2017, 19, 1808–1821. [Google Scholar] [CrossRef] [PubMed]
- Browning, H.; Hayles, J.; Mata, J.; Aveline, L.; Nurse, P.; McIntosh, J.R. Tea2p Is a Kinesin-like Protein Required to Generate Polarized Growth in Fission Yeast. J. Cell Biol. 2000, 151, 15–28. [Google Scholar] [CrossRef]
- Konzack, S.; Rischitor, P.E.; Enke, C.; Fischer, R. The Role of the Kinesin Motor KipA in Microtubule Organization and Polarized Growth ofAspergillus nidulans. Mol. Biol. Cell 2005, 16, 497–506. [Google Scholar] [CrossRef]
- Seidel, C.; Moreno-Velásquez, S.D.; Riquelme, M.; Fischer, R. Neurospora crassa NKIN2, a Kinesin-3 Motor, Transports Early Endosomes and Is Required for Polarized Growth. Eukaryot. Cell 2013, 12, 1020–1032. [Google Scholar] [CrossRef]
- Schuchardt, I.; Aßmann, D.; Thines, E.; Schuberth, C.; Steinberg, G. Myosin-V, Kinesin-1, and Kinesin-3 Cooperate in Hyphal Growth of the Fungus Ustilago maydis. Mol. Biol. Cell 2005, 16, 5191–5201. [Google Scholar] [CrossRef]
- Weber, R.W.S.; Wakley, G.E.; Thines, E.; Talbot, N.J. The vacuole as central element of the lytic system and sink for lipid droplets in maturing appressoria of Magnaporthe grisea. Protoplasma 2001, 216, 101–112. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Guo, C.; Chen, C.; Hu, Z.; Zheng, X.; Xu, S.; Yang, X.; Xie, C. A Kinesin Vdkin2 Required for Vacuole Formation, Mycelium Growth, and Penetration Structure Formation of Verticillium dahliae. J. Fungi 2022, 8, 391. https://doi.org/10.3390/jof8040391
Yang X, Guo C, Chen C, Hu Z, Zheng X, Xu S, Yang X, Xie C. A Kinesin Vdkin2 Required for Vacuole Formation, Mycelium Growth, and Penetration Structure Formation of Verticillium dahliae. Journal of Fungi. 2022; 8(4):391. https://doi.org/10.3390/jof8040391
Chicago/Turabian StyleYang, Xing, Cuimei Guo, Chi Chen, Zhijuan Hu, Xinyao Zheng, Shan Xu, Xingyong Yang, and Chengjian Xie. 2022. "A Kinesin Vdkin2 Required for Vacuole Formation, Mycelium Growth, and Penetration Structure Formation of Verticillium dahliae" Journal of Fungi 8, no. 4: 391. https://doi.org/10.3390/jof8040391
APA StyleYang, X., Guo, C., Chen, C., Hu, Z., Zheng, X., Xu, S., Yang, X., & Xie, C. (2022). A Kinesin Vdkin2 Required for Vacuole Formation, Mycelium Growth, and Penetration Structure Formation of Verticillium dahliae. Journal of Fungi, 8(4), 391. https://doi.org/10.3390/jof8040391