
Synopsis of Leptosphaeriaceae and Introduction of Three New Taxa and One New Record from China

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation
2.2. Morphological Observation
2.3. DNA Extraction, PCR Amplification and Sequencing
2.4. Phylogenetic Analysis
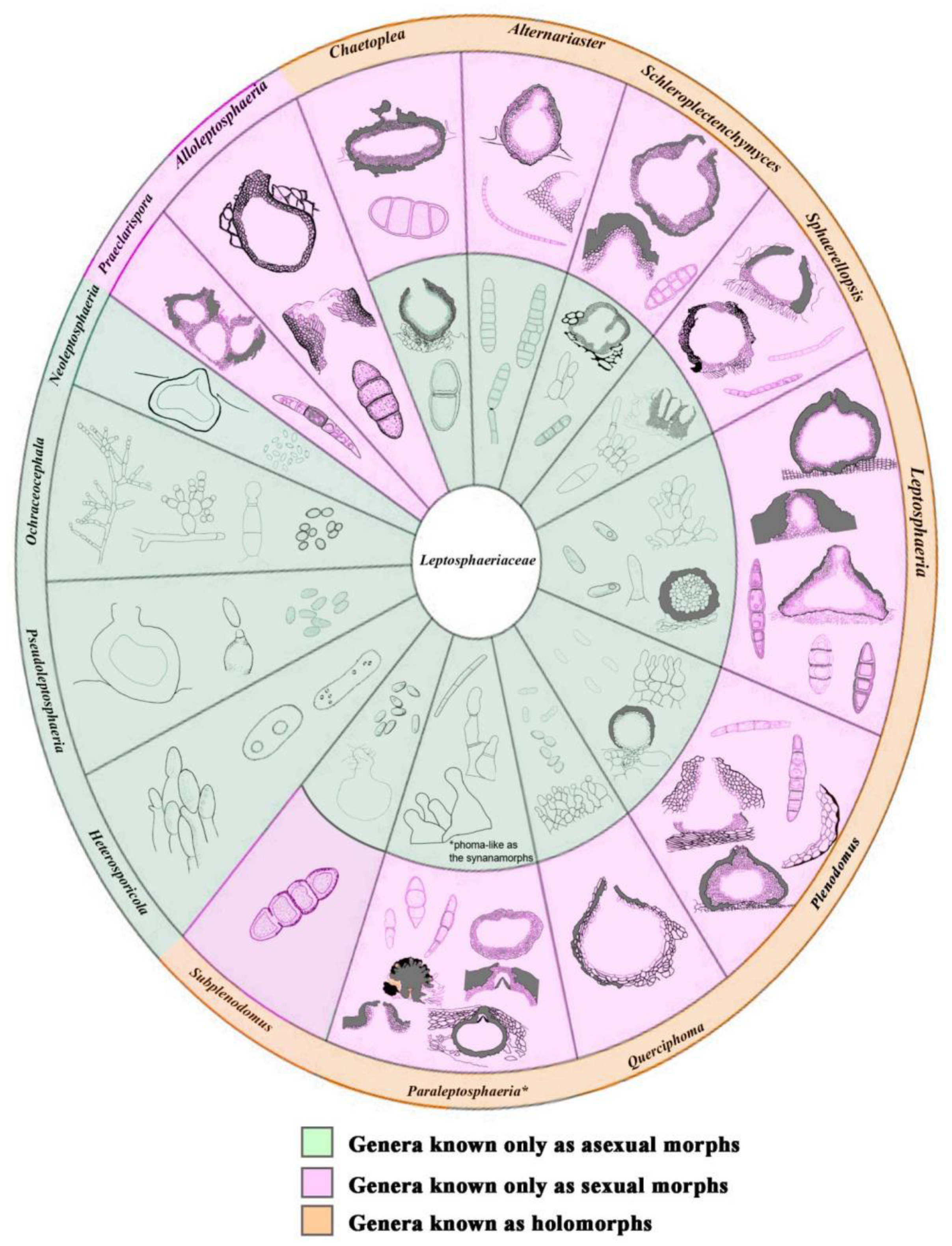
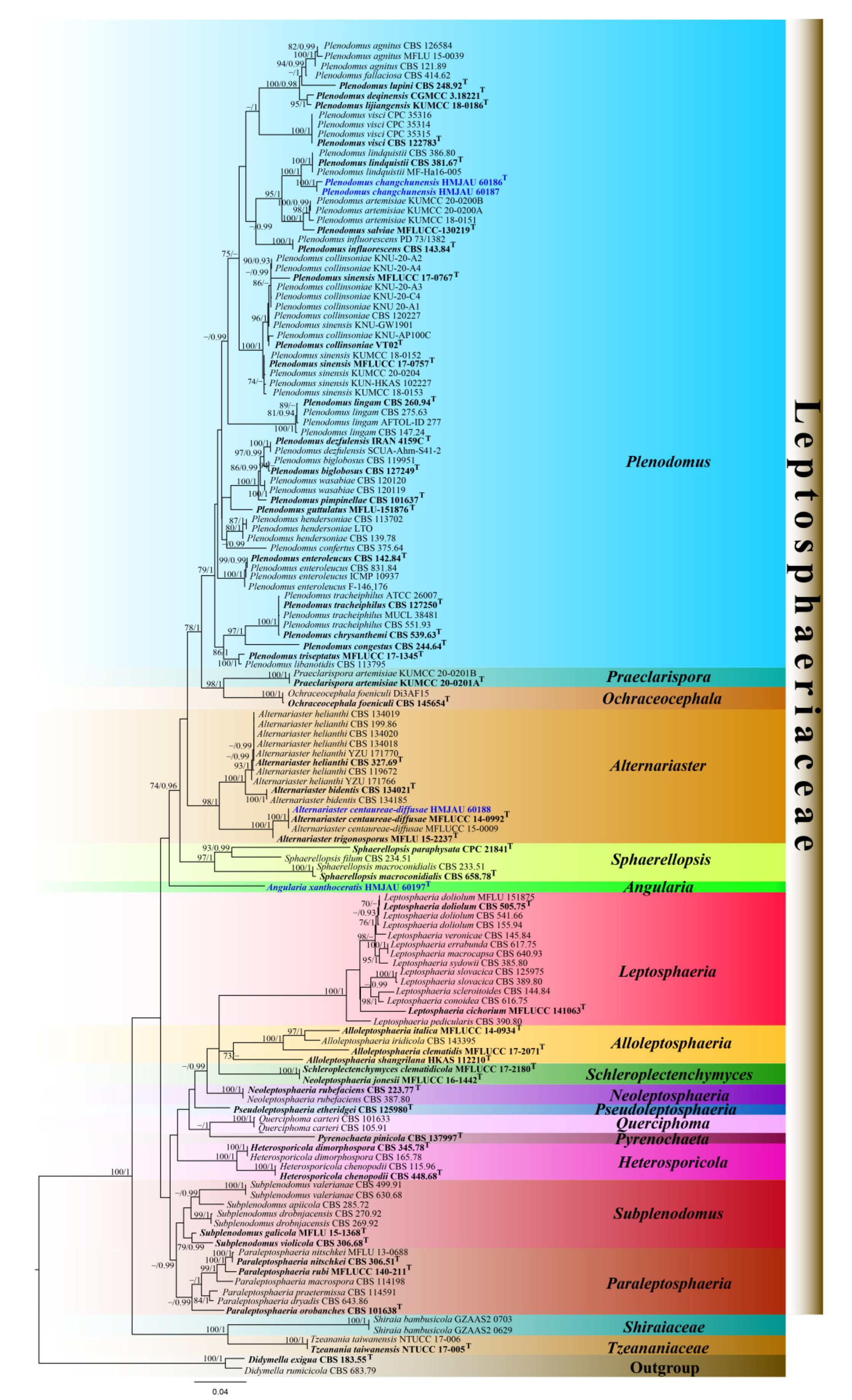
3. Results
3.1. Phylogenetic Analyses
3.2. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barr, M.E. New taxa and combinations in the loculoascomycetes. Mycotaxon 1987, 29, 501–505. [Google Scholar]
- Hyde, K.D.; Jones, E.B.G.; Liu, J.K.; Ariyawansa, H.A.; Boehm, E.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.Q.; et al. Families of Dothideomycetes. Fungal Divers. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Phukhamsakda, C.; Thambugala, K.M.; Bulgakov, T.S.; Wanasinghe, D.N.; Perera, R.H.; Mapook, A.; Camporesi, E.; Kang, J.C.; Jones, E.B.G.; et al. Revision and phylogeny of Leptosphaeriaceae. Fungal Divers. 2015, 74, 19–51. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Crous, P.W.; Kirk, P.M.; Hawksworth, D.L.; Boonmee, S.; Braun, U.; Chomnunti, P.; Dai, D.Q.; D’souza, M.J.; Diederich, P.; et al. Naming and outline of Dothideomycetes–2014 including proposals for the protection or suppression of generic names. Fungal Divers. 2014, 69, 1–55. [Google Scholar] [CrossRef]
- Zhang, Y.; Crous, P.W.; Schoch, C.L.; Hyde, K.D. Pleosporales. Fungal Divers. 2012, 52, 1–225. [Google Scholar] [CrossRef] [Green Version]
- Barr, M.E. Some dictyosporous genera and species of Pleosporales in North America. Mem. New York Bot. Gard. 1990, 62, 1–92. [Google Scholar]
- Crous, P.W.; Groenewald, J.Z. The genera of Fungi–G 4: Camarosporium and Dothiora. IMA Fungus 2017, 8, 131–152. [Google Scholar] [CrossRef] [Green Version]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Phookamsak, R.; Liu, J.K.; McKenzie, E.H.C.; Manamgoda, D.S.; Ariyawansa, H.; Thambugala, K.M.; Dai, D.Q.; Camporesi, E.; Chukeatirote, E.; Wijayawardene, N.N.; et al. Revision of Phaeosphaeriaceae. Fungal Divers. 2014, 68, 159–238. [Google Scholar] [CrossRef]
- Piątek, M.; Rodriguez-Flakus, P.; Domic, A.; Palabral-Aguilera, A.N.; Gómez, M.I.; Flakus, A. Phylogenetic placement of Leptosphaeria polylepidis, a pathogen of Andean endemic Polylepis tarapacana, and its newly dis-covered mycoparasite Sajamaea mycophila gen. et sp. nov. Mycol. Prog. 2020, 19, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Aiello, D.; Vitale, A.; Polizzi, G.; Voglmayr, H. Ochraceocephala foeniculi gen. et sp. nov., a new pathogen causing crown rot of fennel in Italy. MycoKeys 2020, 66, 1–22. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Jones, E.B.G.; Jayarama Bhat, D.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Doilom, M.; Hyde, K.D.; Dong, W.; Liao, C.F.; Suwannarach, N.; Lumyong, S. The plant family Asteraceae is a cache for novel fungal diversity: Novel species and genera with remarkable ascospores in Leptosphaeriaceae. Front. Microbiol. 2021, 12, 660261. [Google Scholar] [CrossRef]
- Câmara, M.P.S.; Palm, M.E.; van Berkum, P.; O’Neill, N.R. Molecular phylogeny of Leptosphaeria and Phaeosphaeria. Mycologia 2002, 94, 630–640. [Google Scholar] [CrossRef]
- Alves, J.L.; Woudenberg, J.H.C.; Duarte, L.L.; Crous, P.W.; Barreto, R.W. Reappraisal of the genus Alternariaster (Dothideomycetes). Persoonia 2013, 31, 77. [Google Scholar] [CrossRef] [Green Version]
- de Gruyter, J.; Woudenberg, J.H.C.; Aveskamp, M.M.; Verkley, G.J.M.; Groenewald, J.Z.; Crous, P.W. Redisposition of phoma-like anamorphs in Pleosporales. Stud. Mycol. 2013, 75, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Dayarathne, M.C.; Phookamsak, R.; Ariyawansa, H.A.; Jones, E.B.G.; Camporesi, E.; Hyde, K.D. Phylogenetic and morphological appraisal of Leptosphaeria italica sp. nov. (Leptosphaeriaceae, Pleosporales) from Italy. Mycosphere 2015, 6, 634–642. [Google Scholar] [CrossRef]
- Tennakoon, D.S.; Phookamsak, R.; Wanasinghe, D.N.; Yang, J.B.; Lumyong, S.; Hyde, K.D. Morphological and phylogenetic insights resolve Plenodomus sinensis (Leptosphaeriaceae) as a new species. Phytotaxa 2017, 324, 73–82. [Google Scholar] [CrossRef]
- Hyde, K.D.; de Silva, N.; Jeewon, R.; Bhat, D.J.; Phookamsak, R.; Doilom, M.; Boonmee, S.; Jayawardena, R.S.; Maharachchikumbura, S.S.N.; Senanayake, I.C.; et al. AJOM new records and collections of fungi: 1–100. Asian J. Mycol. 2020, 3, 22–294. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Suetrong, S.; Sakayaroj, J.; Bahkali, A.H.; Abdel-Wahab, M.A.; Boekhout, T.; Pang, K.L. Classification of marine Ascomycota, Basidiomycota, Blastocladiomycota and Chytridiomycota. Fungal Divers. 2015, 73, 1–72. [Google Scholar] [CrossRef]
- Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, J.D.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal Diversity Notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Simmons, E.G. Alternaria: An identification manual. In Biodiversity No 6; CBS Fungal Diversity Centre: Utrecht, The Netherlands, 2007. [Google Scholar]
- Trakunyingcharoen, T.; Lombard, L.; Groenewald, J.Z.; Cheewangkoon, R.; To-anun, C.; Alfenas, A.C.; Crous, P.W. Mycoparasitic species of Sphaerellopsis, and allied lichenicolous and other genera. IMA Fungus 2014, 5, 391–414. [Google Scholar] [CrossRef]
- Preuss, C.G.T. Ubersicht untersuchter pilze, besonders aus der umgegend Hoyerswerda. In Linnaea 1851, 24, 99–153. [Google Scholar]
- Torres, M.S.; Bergen, M.; Singh, S.; Bischoff, J.; Sullivan, R.F.; White, J.F. Plenodomus morganjonesii sp. nov. and a discussion of the genus Plenodomus. Mycotaxon 2005, 93, 333–344. [Google Scholar]
- Boerema, G.H.; de Gruyter, J.; Noordeloos, M.E. Contributions towards a monograph of Phoma (Coelomycetes)—IV. Section: Taxa with large sized conidial dimorphs, in vivo sometimes as Stagonosporopsis synanamorphs. Persoonia 1997, 16, 335–371. [Google Scholar]
- Boerema, G.H.; de Gruyter, J.; Noordeloos, M.E.; Hamers, M.E.C. Phoma Identification Manual: Differentiation of Specific and Infra-Specific Taxa in Culture; CABI Publishing: Wallingford, UK, 2004. [Google Scholar]
- de Gruyter, J.; Aveskamp, M.M.; Woudenberg, J.H.C.; Verkley, G.J.M.; Groenewald, J.Z.; Crous, P.W. Molecular phylogeny of Phoma and allied anamorph genera: Towards a reclassification of the Phoma complex. Mycol. Res. 2009, 113, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Marin-Felix, Y.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Marincowitz, S.; Barnes, I.; Bensch, K.; Braun, U.; Camporesi, E.; Damm, U.; et al. Genera of phytopathogenic fungi: GOPHY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Crous, P.W.; Hou, L.W.; Duan, W.J.; Cai, L.; Ma, Z.Y.; Liu, F. Fungi of quarantine concern for China I: Dothideomycetes. Persoonia 2021, 47, 45–105. [Google Scholar] [CrossRef]
- Alcorn, J.L.; Pont, W. Alternaria helianthi on sunflower. Australas. Plant Path. 1972, 1, 30. [Google Scholar] [CrossRef]
- Ribeiro, I.J.O.; Filho, O.M.; Soave, J.; Corvellini, G.D.S. Ocorrência de Alternaria Helianthi (Hansf.) Tubaki et Nishihara sobre girassol (Heljanthus annuus L.). Bragantia 1974, 33, 81–85. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guid. Methods Appl. 1990, 18, 315–322. [Google Scholar]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Hall, W.J. Unbiased estimation following a group sequential test. Biometrika 1999, 86, 71–78. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thiyagaraja, V.; Wanasinghe, D.N.; Karunarathna, S.C.; Tennakoon, D.S.; Hyde, K.D.; To-anun, C.; Cheewangkoon, R. Alloleptosphaeria shangrilana sp. nov. and first report of the genus (Leptosphaeriaceae, Dothideomycetes) from China. Phytotaxa 2021, 491, 12–22. [Google Scholar] [CrossRef]
- Safi, A.; Mehrabi-Koushki, M.; Farokhinejad, R. Plenodomus dezfulensis sp. nov. causing leaf spot of Rapeseed in Iran. Phytotaxa 2021, 523, 141–154. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest 2.0; Program Distributed by the Author; Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A. FigTree: Tree Figure Drawing Tool; Version 1.2.2; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2008. [Google Scholar]
- Duan, W.-J.; Li, L.; Mo, S.-M.; Duan, L.-J.; Cai, L. Identification of the quarantine fungus Plenodomus lindquistii from the sunflower seeds imported from Kazakhstan. J. Plant Protect. 2015, 42, 795–800. [Google Scholar]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; Mckenzie, E.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Pem, D.; Bhat, D.J.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Phukhamsakda, C.; Jeewon, R.; Promputtha, I.; Hyde, K.D. Integrating different lines of evidence to establish a novel Ascomycete genus and family (Anastomitrabeculia, Anastomitrabeculiaceae) in Pleosporales. J. Fungi 2021, 7, 94. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Staplers, J.A. Dictionary of the Fungi, 10th ed.; CABI Bioscience: Wallingford, UK, 2008. [Google Scholar]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. 2021. Available online: https://nt.ars-grin.gov/fungaldatabases (accessed on 15 January 2022).
- Chaiwan, N.; Gomdola, D.; Wang, S.; Monkai, J.; Tibpromma, S.; Doilom, M.; Wanasinghe, D.N.; Mortimer, P.E.; Lumyong, S.; Hyde, K.D. https://gmsmicrofungi.org: An online database providing updated information of microfungi in the Greater Mekong Subregion. Mycosphere 2021, 12, 1513–1526. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Niskanen, T.; Suwannarach, N.; Wannathes, N.; Chen, Y.J.; McKenzie, E.H.C.; Maharachchikumbura, S.S.N.; Buyck, B.; Zhao, C.L.; Fan, Y.G.; et al. The numbers of fungi: Are the most speciose genera truly diverse? Fungal Divers. 2022. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; Nilsson, R.H.; Bhunjun, C.S.; Gomes de Farias, A.R.; Sun, Y.R.; Wijesinghe, S.N.; Raza, M.; Bao, D.F.; Lu, L.; Tibpromma, S.; et al. The numbers of fungi; contributions from traditional taxonomic studies and challenges of metabarcoding. Fungal Divers. 2022; in press. [Google Scholar]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Phukhamsakda, C.; Jayawardena, R.S.; Jeewon, R.; Promputtha, I.; Hyde, K.D. Investigating species boundaries in Colletotrichum. Fungal Divers. 2021, 107, 107–127. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Dong, Y.; Jayawardena, R.S.; Jeewon, R.; Phukhamsakda, C.; Bundhun, D.; Hyde, K.D.; Sheng, J. A polyphasic approach to delineate species in Bipolaris. Fungal Divers. 2020, 102, 225–256. [Google Scholar] [CrossRef]
- Fitt, B.D.L.; Brun, H.; Barbetti, M.J.; Rimmer, S.R. World-wide importance of Phoma stem canker (Leptosphaeria maculans and L. biglobosa) on oilseed rape (Brassica napus). Eur. J. Plant Pathol. 2006, 114, 3–15. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; Jeewon, R.; Ghobad-Nejhad, M.; Wanasinghe, D.N.; Liu, N.G.; Phillips, A.J.L.; Oliveira-Filho, J.R.C.; da Silva, G.A.; Gibertoni, T.B.; et al. One stop shop II: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 26–50 (2019). Fungal Divers. 2019, 94, 41–129. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Perera, R.H.; de Silva, N.I.; Maharachchikumbura, S.S.N.; Samarakoon, M.C.; Ekanayake, A.H.; et al. One stop shop III: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 51–75 (2019). Fungal Divers. 2019, 98, 77–160. [Google Scholar] [CrossRef]
- Zhang, Y.; Jeewon, R.; Fournier, J.; Hyde, K.D. Multi-gene phylogeny and morphotaxonomy of Amniculicola lignicola: Novel freshwater fungus from France and its relationships to the Pleosporales. Mycol. Res. 2008, 112, 1186–1194. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Host | Strain/Isolate | GenBank Accession Numbers | |||
|---|---|---|---|---|---|---|
| ITS | LSU | SSU | tub2 | |||
| Alloleptosphaeria clematidis | Clematis subumbellata | MFLUCC 17-2071 | MT310604 | MT214557 | MT226674 | _ |
| All. iridicola | Iris sp. | CBS 143395 | MH107919 | MH107965 | _ | _ |
| All. italica | _ | MFLUCC 14-0934 | KT454722 | KT454714 | _ | _ |
| All. shangrilana | _ | HKAS: 112210 | MW431059 | MW431315 | MW431058 | _ |
| Alternariaster bidentis | Bidens sulphurea | CBS 134021 | KC609333 | KC609341 | _ | _ |
| Alt. bidentis | Bidens sulphurea | CBS 134185 | KC609334 | KC609342 | _ | _ |
| Alt. centaureae-diffusae | Centaurea diffusa Lam. | MFLUCC 14-0992 | KT454723 | KT454715 | KT454730 | _ |
| Alt. centaureae-diffusae | Centaurea diffusa | MFLUCC 150009 | KT454724 | KT454716 | KT454731 | _ |
| Alt. centaureae-diffusae | Clematis spp. | HMJAU 60188 | OL996125 | OL897175 | OL891810 | OL898721 |
| Alt. helianthi | _ | YZU 171766 | MZ702726 | _ | _ | _ |
| Alt. helianthi | _ | YZU 171770 | MZ702727 | _ | _ | _ |
| Alt. helianthi | Helianthus annuus | CBS 327.69 | KC609335 | KC584369 | KC584627 | _ |
| Alt. helianthi | Helianthus annuus | CBS 199.86 | KC609336 | KC609343 | _ | _ |
| Alt. helianthi | Helianthus sp. | CBS 119672 | KC609337 | KC584368 | KC584626 | _ |
| Alt. helianthi | Helianthus annuus | CBS 134018 | KC609338 | KC609344 | _ | _ |
| Alt. helianthi | Helianthus annuus | CBS 134019 | KC609339 | KC609345 | _ | _ |
| Alt. helianthi | Helianthus annuus | CBS 134020 | KC609340 | KC609346 | _ | _ |
| Alt. trigonosporus | Cirsium sp. | MFLU 15-2237 | KY674857 | KY674858 | _ | _ |
| Angularia xanthoceratis | Xanthoceras sorbifolium | HMJAU 60197 | OM295683 | OM295682 | OM295681 | OM304358 |
| Didymella exigua | Rumex arifolius | CBS 183.55 | GU237794 | EU754155 | EU754056 | GU237525 |
| D. rumicicola | Rumex obtusifolius | CBS 683.79 | KT389503 | KT389721 | _ | KT389800 |
| Heterosporicola chenopodii | Chenopodium album | CBS 448.68 | FJ427023 | EU754187 | EU754088 | _ |
| H. chenopodii | Chenopodium album | CBS 115.96 | JF740227 | EU754188 | EU754089 | _ |
| H. dimorphospora | Chenopodium quinoa | CBS 165.78 | JF740204 | JF740281 | JF740098 | _ |
| H. dimorphospora | Chenopodium quinoa | CBS 345.78 | JF740203 | GU238069 | GU238213 | _ |
| Leptosphaeria cichorium | Cichorium intybus | MFLUCC 14-1063 | KT454720 | KT454712 | KT454728 | _ |
| L. conoidea | Lunaria annua | CBS 616.75 | JF740201 | JF740279 | _ | KT389804 |
| L. doliolum | Phlox paniculata | CBS 155.94 | JF740207 | JF740282 | _ | JF740146 |
| L. doliolum | _ | MFLU: 151875 | KT454727 | KT454719 | KT454734 | _ |
| L. doliolum | Rudbeckia sp. | CBS 541.66 | JF740206 | JF740284 | _ | JF740145 |
| L. doliolum | Urtica dioica | CBS 505.75 | JF740205 | GQ387576 | GQ387515 | JF740144 |
| L. errabunda | Solidago sp. | CBS 617.75 | JF740216 | JF740289 | _ | JF740150 |
| L. macrocapsa | Mercurialis perennis | CBS 640.93 | JF740237 | JF740304 | _ | JF740156 |
| L. pedicularis | Pedicularis sp. | CBS 390.80 | JF740224 | JF740294 | _ | JF740155 |
| L. scleroitoides | Medicago sativa | CBS 144.84 | JF740192 | JF740269 | _ | _ |
| L. slovacica | Ballota nigra | CBS 125975 | JF740248 | JF740316 | _ | _ |
| L. slovacica | Balota nigra | CBS 389.80 | JF740247 | JF740315 | JF740101 | _ |
| L. sydowii | Senecio jacobaea | CBS 385.80 | JF740244 | JF740313 | _ | JF740157 |
| L. veronicae | Veronica chamaedrys subsp. chamaedryoides | CBS 145.84 | JF740254 | JF740320 | _ | JF740160 |
| Neoleptosphaeria jonesii | Clematis vitalba | MFLUCC 16-1442 | KY211869 | KY211870 | KY211871 | _ |
| N. rubefaciens | Quercus | CBS 223.77 | JF740243 | JF740312 | _ | _ |
| N. rubefaciens | Tilia sp. | CBS 387.80 | JF740242 | JF740311 | _ | _ |
| Ochraceocephala foeniculi | Foeniculum vulgare | Di3AF1 = CBS 145654 | MN516753 | MN516774 | MN516743 | MN520147 |
| O. foeniculi | Foeniculum vulgare | Di3AF15 | MN516766 | MN516783 | MN516752 | _ |
| Paraleptosphaeria dryadis | Dryas octopetala | CBS 643.86 | JF740213 | GU301828 | _ | _ |
| Pa. macrospora | Rumex domesticus | CBS 114198 | JF740238 | JF740305 | _ | _ |
| Pa. nitschkei | _ | MFLUCC 13-0688 | KR025860 | KR025864 | _ | _ |
| Pa. nitschkei | Cirsium spinosissimum | CBS 306.51 | JF740239 | JF740308 | _ | KT389833 |
| Pa. orobanches | Epifagus virginiana | CBS 101638 | JF740230 | JF740299 | _ | _ |
| Pa. praetermissa | Rubus idaeus | CBS 114591 | JF740241 | JF740310 | _ | _ |
| Pa. rubi | Rubussp. | MFLUCC 14-0211 | KT454726 | KT454718 | KT454733 | _ |
| Plenodomus agnitus | Eupatorium sp. | CBS 121.89 | JF740194 | JF740271 | _ | KY064053 |
| Pl. agnitus | Eupatorium cannabinum | CBS 126584 | JF740195 | JF740272 | _ | _ |
| Pl. agnitus | _ | MFLU 15-0039 | KP744459 | KP744504 | _ | _ |
| Pl. artemisiae | _ | KUMCC 18-0151 | MK387920 | MK387958 | MK387928 | _ |
| Pl. artemisiae | Artemisia argyi | KUMCC 20-0200A | MT957062 | MT957055 | MT957048 | _ |
| Pl. artemisiae | Artemisia argyi | KUMCC 20-0200B | MT957063 | MT957056 | MT957049 | _ |
| Pl. biglobosus | Brassica rapa | CBS 119951 | JF740198 | JF740274 | JF740102 | KY064054 |
| Pl. biglobosus | Brassica juncea | CBS 127249 | JF740199 | JF740275 | _ | _ |
| Pl. changchunensis | Poaceae | HMJAU 60186 | OL996123 | OL897174 | OL984031 | OM009247 |
| Pl. changchunensis | Poaceae | HMJAU 60187 | OL996124 | OL966928 | OL984032 | OL898716 |
| Pl. chrysanthemi | Chrysanthemum sp. | CBS 539.63 | JF740253 | GU238151 | GU238230 | KY064055 |
| Pl. collinsoniae | Vitis coignetiae | CBS 120227 | JF740200 | JF740276 | _ | KY064056 |
| Pl. collinsoniae | _ | VT02 | MN653010 | MN982862 | MN652269 | _ |
| Pl. collinsoniae | _ | KNU-AP100C | LC550566 | LC550568 | _ | _ |
| Pl. collinsoniae | Malus domestica | KNU-20-A1 | LC591836 | _ | _ | LC591846 |
| Pl. collinsoniae | Malus domestica | KNU-20-A2 | LC591837 | _ | _ | LC591847 |
| Pl. collinsoniae | Malus domestica | KNU-20-A3 | LC591838 | _ | _ | LC591848 |
| Pl. collinsoniae | Malus domestica | KNU-20-A4 | LC591839 | _ | _ | LC591849 |
| Pl. collinsoniae | Malus domestica | KNU-20-C4 | LC591840 | _ | _ | LC591850 |
| Pl. confertus | Anacyclus radiatus | CBS 375.64 | AF439459 | JF740277 | _ | KY064057 |
| Pl. congestus | Erigeron canadensis | CBS 244.64 | AF439460 | JF740278 | _ | KY064058 |
| Pl. deqinensis | _ | CGMCC 3.18221 | KY064027 | KY064031 | _ | KY064052 |
| Pl. dezfulensis | Brassica napus | IRAN 4159C = SCUA-Ahm-S41 | MZ048609 | _ | _ | MZ043102 |
| Pl. dezfulensis | Brassica napus | SCUA-Ahm-S41-2 | MZ048610 | _ | _ | MZ043103 |
| Pl. enteroleucus | Catalpa bignonioides | CBS 142.84 | JF740214 | JF740287 | _ | KT266266 |
| Pl. enteroleucus | Triticum aestivum | CBS 831.84 | JF740215 | JF740288 | _ | KT266270 |
| Pl. enteroleucus | Fraxinus angustifolia | F-146,176 | MN910295 | MN910294 | _ | _ |
| Pl. enteroleucus | Citrus sp. | ICMP:10937 | KT309810 | KT309635 | _ | KT309399 |
| Pl. fallaciosus | Satureja montana | CBS 414.62 | JF740222 | JF740292 | _ | _ |
| Pl. guttulatus | _ | MFLU 151876 | KT454721 | KT454713 | KT454729 | _ |
| Pl. hendersoniae | Pyrus malus | CBS 139.78 | JF740226 | JF740296 | _ | _ |
| Pl. hendersoniae | Salix cinerea | CBS 113702 | JF740225 | JF740295 | _ | KT266271 |
| Pl. hendersoniae | Salix appendiculata | LTO | MF795790 | _ | _ | _ |
| Pl. influorescens | Fraxinus excelsior | CBS 143.84 | JF740228 | JF740297 | _ | KT266267 |
| Pl. influorescens | Lilium sp. | PD 73/1382 | JF740229 | JF740298 | _ | KT266273 |
| Pl. libanotidis | Seseli libanotis | CBS 113795 | JF740231 | JF740300 | _ | KY064059 |
| Pl. lijiangensis | _ | KUMCC 18-0186 | MK387921 | MK387959 | MK387929 | _ |
| Pl. lindquistii | Helianthus annuus | CBS 381.67 | JF740233 | JF740302 | _ | _ |
| Pl. lindquistii | Helianthus annuus | CBS 386.80 | JF740232 | JF740301 | _ | _ |
| Pl. lindquistii | Helianthus annuus | MF-Ha16-005 | MK495988 | _ | _ | MK501790 |
| Pl. lingam | _ | AFTOL-ID 277 | KT225526 | DQ470946 | DQ470993 | _ |
| Pl. lingam | Brassica oleracea | CBS 260.94 | JF740235 | JF740307 | _ | MZ073915 |
| Pl. lingam | Brassica sp. | CBS 275.63 | MW810266 | JF740306 | _ | MZ073916 |
| Pl. lingam | _ | CBS 147.24 | MW810259 | JX681097 | _ | MZ073914 |
| Pl. lupini | Lupinus mutabilis | CBS 248.92 | JF740236 | JF740303 | _ | KY064061 |
| Pl. pimpinellae | Pimpenella anisum | CBS 101637 | JF740240 | JF740309 | _ | KY064062 |
| Pl. salviae | Salvia glutinosa | MFLUCC: 13-0219 | KT454725 | KT454717 | KT454732 | _ |
| Pl. sinensis | Plukenetia sp. | MFLUCC 17-0757 | MF072722 | MF072718 | MF072720 | _ |
| Pl. sinensis | Tamarindus sp. | MFLUCC 17-0767 | MF072721 | MF072717 | MF072719 | _ |
| Pl. sinensis | _ | KNU-GW1901 | LC550567 | LC550569 | LC550570 | _ |
| Pl. sinensis | Ageratina adenophora | KUMCC 20-0204 | MT957064 | MT957057 | MT957050 | _ |
| Pl. sinensis | _ | KUMCC 18-0153 | MK387922 | MK387960 | MK387930 | _ |
| Pl. sinensis | _ | KUMCC 18-0152 | MK387923 | MK387961 | MK387931 | _ |
| Pl. sinensis | _ | KUN-HKAS 102227 | MK387924 | MK387962 | MK387932 | _ |
| Pl. tracheiphilus | Citrus limonia | CBS 551.93 | JF740249 | JF740317 | JF740104 | MZ073918 |
| Pl. tracheiphilus | Citrus aurantium | CBS 127250 | JF740250 | JF740318 | _ | MZ073919 |
| Pl. tracheiphilus | Citrus limon | MUCL 38481 | MW810293 | MW715037 | _ | MZ073920 |
| Pl. tracheiphilus | Citrus sp. | ATCC 26007 | MZ049614 | MW959165 | _ | MZ073908 |
| Pl. triseptatus | Daucus carota | MFLUCC 17-1345 | MN648452 | MN648451 | _ | _ |
| Pl. visci | Viscum album | CBS 122783 | JF740256 | EU754195 | EU754096 | KY064063 |
| Pl. visci | Viscum album | CPC:35316 | MT223832 | MT223924 | _ | _ |
| Pl. visci | Viscum album | CPC:35315 | MT223831 | MT223923 | _ | _ |
| Pl. visci | Viscum album | CPC:35314 | MT223830 | MT223922 | _ | _ |
| Pl. wasabiae | Eutrema wasabi | CBS 120119 | JF740257 | JF740323 | _ | KT266272 |
| Pl. wasabiae | Eutrema japonicum | CBS 120120 | JF740258 | JF740324 | _ | _ |
| Praeclarispora artemisiae | Artemisia argyi | KUMCC 20-0201A | MT957060 | MT957053 | MT957046 | |
| Pr. artemisiae | Artemisia argyi | KUMCC 20-0201B | MT957061 | MT957054 | MT957047 | |
| Pseudoleptosphaeria etheridgei | Populus tremuloides | CBS 125980 | JF740221 | JF740291 | _ | _ |
| pyrenochaeta pinicola | Pinus sp. | CBS 137997 | KJ869152 | KJ869209 | _ | KJ869249 |
| Querciphoma carteri | Quercus robur | CBS 105.91 | KF251209 | GQ387594 | GQ387533 | KF252700 |
| Q. carteri | Quercus sp. | CBS 101633 | KF251210 | GQ387593 | GQ387532 | KF252701 |
| Schleroplectenchymyces clematidis | Clematis vitalba | MFLUCC 17-2180 | MT310605 | MT214558 | MT226675 | _ |
| Shiraia bambusicola | Phyllostachys sp. | GZAAS2 0703 | GQ845412 | KC460981 | _ | _ |
| Sh. bambusicola | Pleioblastus sp. | GZAAS2 0629 | GQ845415 | KC460980 | _ | _ |
| Sphaerellopsis filum | _ | CBS 234.51 | KP170655 | KP170723 | _ | KP170704 |
| Sp. macroconidialis | Dianthus caryophyllus | CBS 233.51 | KP170658 | KP170726 | _ | KP170707 |
| Sp. macroconidialis | Allium schoenoprasum | CBS 658.78 | KP170659 | KP170727 | _ | KP170708 |
| Sp. paraphysata | Cenchrus sp. | CPC 21841 | KP170662 | KP170729 | _ | KP170710 |
| Subplenodomus apiicola | Apium graveolens var. rapaceum | CBS 285.72 | JF740196 | GU238040 | GU238211 | _ |
| Su. drobnjacensis | Eustoma exaltatum | CBS 269.92 | JF740211 | JF740285 | JF740100 | _ |
| Su. drobnjacensis | Gentiana sp. | CBS 270.92 | JF740212 | JF740286 | _ | _ |
| Su. galicola | Galium sp. | MFLU 15-1368 | KY554204 | KY554199 | _ | _ |
| Su. valerianae | Valeriana officinalis | CBS 499.91 | JF740252 | JF740319 | _ | _ |
| Su. valerianae | Valeriana phu | CBS 630.68 | JF740251 | GU238150 | GU238229 | _ |
| Su. violicola | Viola tricolor | CBS 306.68 | FJ427083 | GU238156 | GU238231 | KT389849 |
| Tzeanania taiwanensis | Ophiocordyceps macroacicularis | NTUCC 17-005 | MH461123 | MH461120 | MH461126 | MH461132 |
| T. taiwanensis | Ophiocordyceps macroacicularis | NTUCC 17-006 | MH461124 | MH461121 | MH461127 | MH461133 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, R.; Su, W.; Tian, S.; Bhunjun, C.S.; Tibpromma, S.; Hyde, K.D.; Li, Y.; Phukhamsakda, C. Synopsis of Leptosphaeriaceae and Introduction of Three New Taxa and One New Record from China. J. Fungi 2022, 8, 416. https://doi.org/10.3390/jof8050416
Xu R, Su W, Tian S, Bhunjun CS, Tibpromma S, Hyde KD, Li Y, Phukhamsakda C. Synopsis of Leptosphaeriaceae and Introduction of Three New Taxa and One New Record from China. Journal of Fungi. 2022; 8(5):416. https://doi.org/10.3390/jof8050416
Chicago/Turabian StyleXu, Rong, Wenxin Su, Shangqing Tian, Chitrabhanu S. Bhunjun, Saowaluck Tibpromma, Kevin D. Hyde, Yu Li, and Chayanard Phukhamsakda. 2022. "Synopsis of Leptosphaeriaceae and Introduction of Three New Taxa and One New Record from China" Journal of Fungi 8, no. 5: 416. https://doi.org/10.3390/jof8050416
APA StyleXu, R., Su, W., Tian, S., Bhunjun, C. S., Tibpromma, S., Hyde, K. D., Li, Y., & Phukhamsakda, C. (2022). Synopsis of Leptosphaeriaceae and Introduction of Three New Taxa and One New Record from China. Journal of Fungi, 8(5), 416. https://doi.org/10.3390/jof8050416