
A Panoramic View on Grapevine Trunk Diseases Threats: Case of Eutypa Dieback, Botryosphaeria Dieback, and Esca Disease

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. The Complexity of Grapevine Trunk Diseases
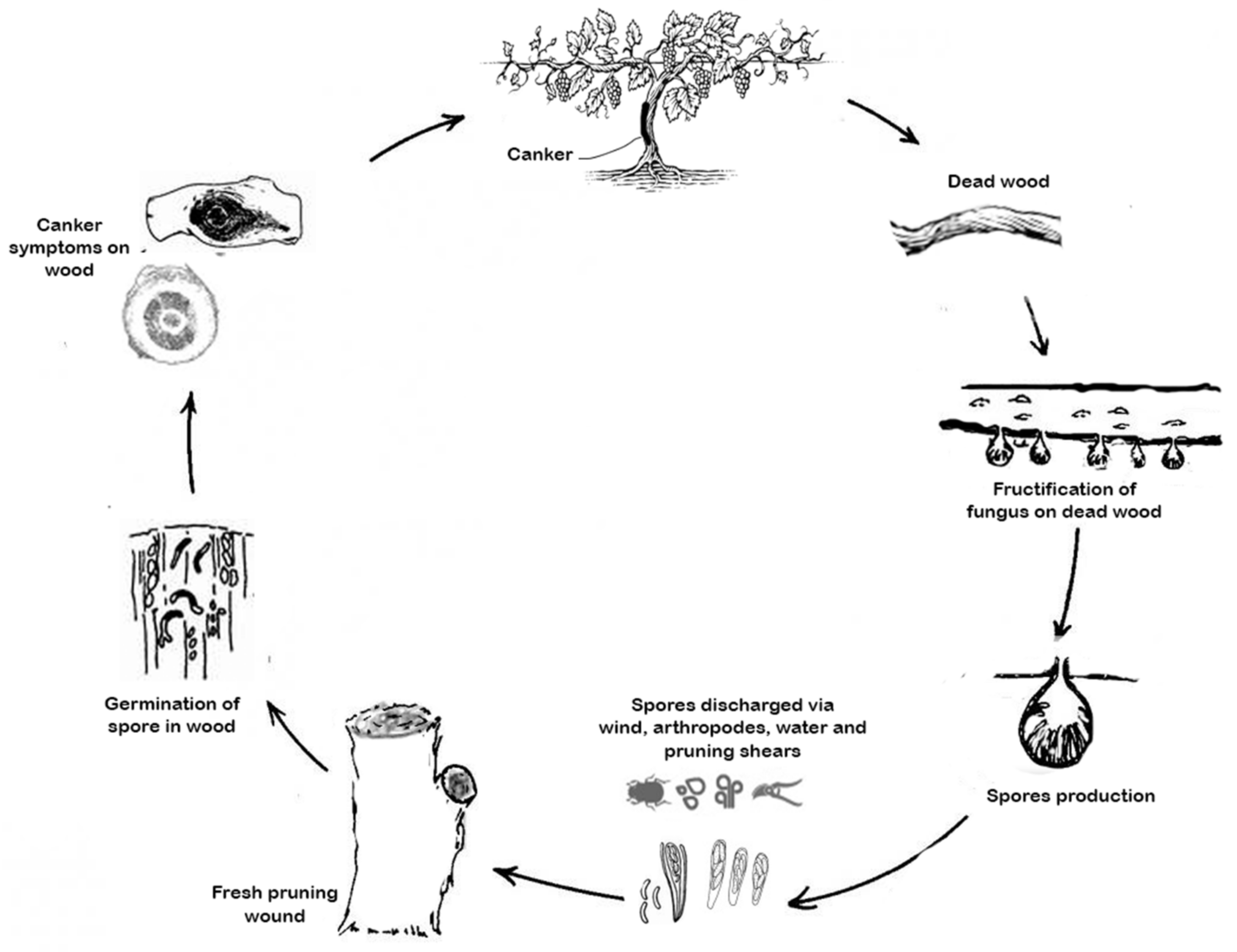
2.1. Eutypa Dieback
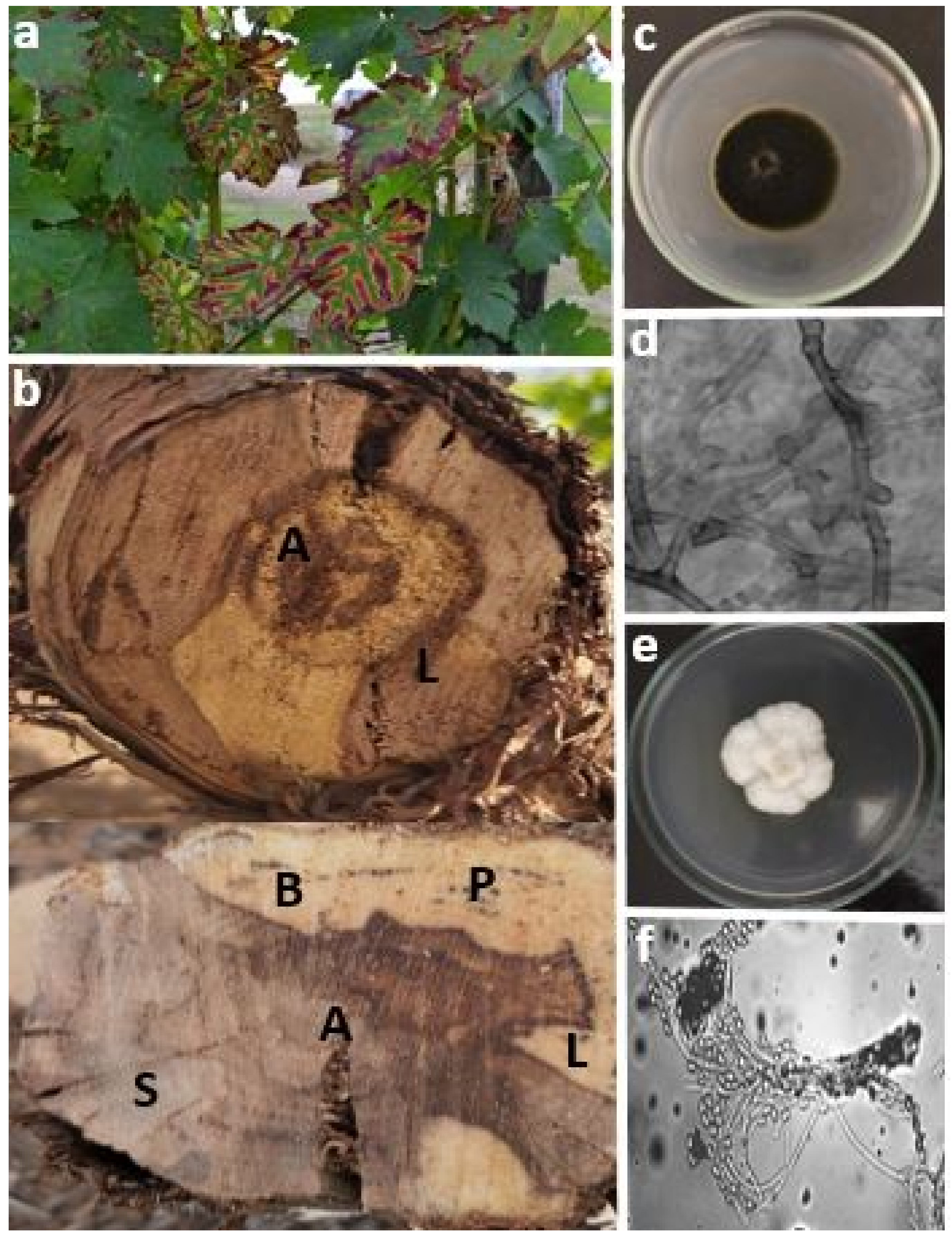
2.2. Esca Disease
2.3. Botryosphaeria Dieback
3. Identification of Pathogens
3.1. Diagnosis of the Disease and Molecular Identification
3.2. Real-Time PCR
4. Grapevine Defense Mechanisms against Fungal Attack
5. Molecular Mechanisms of Fungal Endophytes Colonization
6. Control Methods
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qiu, W.; Feechan, A.; Dry, I. Current understanding of grapevine defense mechanisms against the biotrophic fungus (Erysiphe necator), the causal agent of powdery mildew disease. Hortic. Res. 2015, 2, 15020. [Google Scholar] [CrossRef] [Green Version]
- OIV. State of the World Vitivinicultural Sector in 2020; International Organisation of Vine and Wine: Paris, France, 2021; pp. 1–19. [Google Scholar]
- Gorton, L. Through the Grapevine: Tracing the Origins of Wine; The Ohio State University: Columbus, OH, USA, 2014; ISBN 1687971587. [Google Scholar]
- DRA. La Culture de la Vigne au Maroc. La Direction Régionale de l’Agriculture Méknes. DRA: Perth, Australia, 2019. [Google Scholar]
- García, R.A.A.; Revilla, E. The current status of wild grapevine populations (Vitis vinifera ssp sylvestris) in the Mediterranean basin. In Agricultural and Biological Sciences. The Mediterranean Genetic-Code-Grapevine and Olive; IntechOpen: London, UK, 2013; pp. 51–72. [Google Scholar]
- Pouzoulet, J. Développement D’une Méthodologie PCR en Temps Réel Pour la Détection et la Quantification in Planta des Principaux Champignons Pathogènes Associés aux Maladies du Bois de la Vigne. Ph.D. Thesis. Universite de Toulouse, Toulouse, France, 2012. [Google Scholar]
- Travadon, R.; Lecomte, P.; Diarra, B.; Lawrence, D.P.; Renault, D.; Ojeda, H.; Rey, P.; Baumgartner, K. Grapevine pruning systems and cultivars influence the diversity of wood-colonizing fungi. Fungal Ecol. 2016, 24, 82–93. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, D.P.; Travadon, R.; Nita, M.; Baumgartner, K. TrunkDiseaseID.org: A molecular database for fast and accurate identification of fungi commonly isolated from grapevine wood. Crop Prot. 2017, 102, 110–117. [Google Scholar] [CrossRef] [Green Version]
- Larignon, P.; Fontaine, F.; Farine, S.; Clément, C.; Bertsch, C. Esca et Black Dead Arm: Deux acteurs majeurs des maladies du bois chez la Vigne. Comptes Rendus. Biol. 2009, 332, 765–783. [Google Scholar] [CrossRef] [PubMed]
- Bruez, E.; Larignon, P.; Bertsch, C.; Robert-Siegwald, G.; Lebrun, M.-H.; Rey, P.; Fontaine, F. Impacts of sodium arsenite on wood microbiota of esca-diseased grapevines. J. Fungi 2021, 7, 498. [Google Scholar] [CrossRef] [PubMed]
- Roblin, G.; Luini, E.; Fleurat-Lessard, P.; Larignon, P.; Berjeaud, J.M. Towards a preventive and/or curative treatment of esca in grapevine trunk disease: General basis in the elaboration of treatments to control plant pathogen attacks. Crop Prot. 2019, 116, 156–169. [Google Scholar] [CrossRef]
- Claverie, M.; Notaro, M.; Fontaine, F.; Wéry, J. Current knowledge on Grapevine Trunk Diseases with complex etiology: A systemic approach. Phytopathol. Mediterr. 2020, 59, 29–53. [Google Scholar] [CrossRef]
- Pascoe, I.G.; Edwards, J. Esca or not? Aust. Grapegrow. Winemak. 2002, 14, 439. [Google Scholar]
- Bruez, E.; Lecomte, P.; Grosman, J.; Doublet, B.; Bertsch, C.; Fontaine, F.; Ugaglia, A.; Teissedre, P.L.; Da Costa, J.P.; Guerin-Dubrana, L.; et al. Overview of grapevine trunk diseases in France in the 2000s. Phytopathol. Mediterr. 2013, 52, 262–275. [Google Scholar] [CrossRef]
- Bertsch, C.; Ramírez-Suero, M.; Magnin-Robert, M.; Larignon, P.; Chong, J.; Abou-Mansour, E.; Spagnolo, A.; Clément, C.; Fontaine, F. Grapevine trunk diseases: Complex and still poorly understood. Plant Pathol. 2013, 62, 243–265. [Google Scholar] [CrossRef] [Green Version]
- Murolo, S.; Romanazzi, G. Effects of grapevine cultivar, rootstock and clone on esca disease. Australas. Plant Pathol. 2014, 43, 215–221. [Google Scholar] [CrossRef]
- Sosnowski, M.R.; Luque, J.; Loschiavo, A.P.; Martos, S.; Garcia-Figueres, F.; Wicks, T.J.; Scott, E.S. Studies on the effect of water and temperature stress on grapevines inoculated with Eutypa lata. Phytopathol. Mediterr. 2011, 50, S127–S138. [Google Scholar]
- Gramaje, D.; Di Marco, S. Identifying practices likely to have impacts on grapevine trunk disease infections: A European nursery survey. Phytopathol. Mediterr. 2015, 53, 313–324. [Google Scholar]
- Mostert, L.; Groenewald, J.Z.; Summerbell, R.C.; Gams, W.; Crous, P.W. Taxonomy and pathology of Togninia (Diaporthales) and its Phaeoacremonium anamorphs. Stud. Mycol. 2006, 54, 1–113. [Google Scholar] [CrossRef] [Green Version]
- Serra, S.; Mannoni, M.A.; Ligios, V. Studies on the susceptibility of pruning wounds to infection by fungi involved in grapevine wood diseases in Italy. Phytopathol. Mediterr. 2008, 47, 234–246. [Google Scholar]
- Guan, X.; Essakhi, S.; Laloue, H.; Nick, P.; Bertsch, C.; Chong, J. Mining new resources for grape resistance against Botryosphaeriaceae: A focus on Vitis vinifera subsp. sylvestris. Plant Pathol. 2016, 65, 273–284. [Google Scholar] [CrossRef]
- Hrycan, J.; Hart, M.; Bowen, P.; Forge, T.; Úrbez-Torres, J.R. Grapevine trunk disease fungi: Their roles as latent pathogens and stress factors that favour disease development and symptom expression. Phytopathol. Mediterr. 2020, 59, 395–424. [Google Scholar] [CrossRef]
- Waraich Research Collection. Brisk Binary Robust Invariant Scalable Keypoints. In Proceedings of the 2011 International Conference on Computer Vision, Barcelona, Spain, 6–13 November 2011; pp. 12–19. [Google Scholar]
- Larignon, P. Réflexions Sur L’esca: Ce Que L’on Sait Déjà Montre Qu’il en Reste Beaucoup à Apprendre: Vigne; Firenze University Press: Florence, Italy, 2004. [Google Scholar]
- Peros, J.-P.; Berger, G. A rapid method to assess the aggressiveness of Eutypa lata isolates and the susceptibility of grapevine cultivars to Eutypa dieback. Agronomie 1994, 14, 515–523. [Google Scholar] [CrossRef] [Green Version]
- Moller, W.J.; Kasimatis, A.N. Dieback of grapevines caused by Eutypa armeniacae. Plant Dis. Report. 1978, 62, 254–258. [Google Scholar]
- Rappaz, F. Les espèces sanctionnées du genre Eutypa (Diatrypaceae, ascomycetes): Étude taxonomique et nomenclaturale. Mycotaxon 1984, 20, 567–586. [Google Scholar]
- Bolay, A.; Carter, M.V. Newly recorded hosts of Eutypa lata (=E. armeniacae) in Australia. Plant Prot. Q. 1985, 1, 11. [Google Scholar]
- Kouyeas, H. Eutypa armeniacae on lemon in Greece. J. Phytopathol. 1978, 91, 235–237. [Google Scholar] [CrossRef]
- Glawe, D.A.; Dilley, M.A.; Moller, W.J. Isolation and identification of Eutypa armeniacae from Malus domestica in Washington State. Mycotaxon 1983, 18, 315–318. [Google Scholar]
- Carter, M.V. Additional hosts of Eutypa armeniacae in Australia. Australas. Plant Pathol. 1982, 11, 46–48. [Google Scholar] [CrossRef]
- Rumbos, I.C. Isolation and identification of Eutypa lata from Pistacia vera in Greece. J. Phytopathol. 1986, 116, 352–357. [Google Scholar] [CrossRef]
- Halleen, F.; Volkmann, A.S.; Fourie, P.H. Incidence of Eutypa-like symptoms in Cabernet Sauvignon vineyards in the greater Stellenbosch area. Wynboer 2001, 143, 12–14. [Google Scholar]
- Berraf, A.; Peros, J.-P. Importance de l’eutypiose et de l’esca en Algérie et structure de la communauté fongique associée. J. Int. Des Sci. La Vigne Du Vin 2005, 39, 121–128. [Google Scholar]
- Chliyeh, M.; Rhimini, Y.; Selmaoui, K.; Touhami, A.O.; Filali-Maltouf, A.; El Modafar, C.; Moukhli, A.; Oukabli, A.; Benkirane, R.; Douira, A. Survey of the fungal species associated to olive-tree (Olea europaea L.) in Morocco. Int. J. Recent Biotechnol. 2014, 2, 15–32. [Google Scholar]
- Teliz, D. Colapso, a new disease of grapevines in northern Mexico associated with a species of Ganoderma. Plant Dis. Report. 1979, 63, 705–707. [Google Scholar]
- Ramirez, E.; Pineda, J. Distribución de Eutypa lata y Lasiodiplodia theobromae en las zonas vitícolas de Venezuela. Rev. La Fac. Agron. 2003, 20, 43–52. [Google Scholar]
- Paradela Filho, O.; Ribeiro, I.J.A.; Braga, F.G.; Nogueira, E.M.C.; Pesce, G.; Kuniyuki, H.; Fossa, E.; Rodrigues, N.J.; Feichtenberger, E. Eutypa lata, the agent of grape decline in Sao Paolo state. Summa Phytopathol. 1993, 19, 86–89. [Google Scholar]
- O’Gorman, D.T.; Haag, P.; Sholberg, P.L. First report of Eutypa dieback and other emerging grapevine diseases in the Okanagan Valley. Can. Plant Dis. Surv. 2010, 90, 158–160. [Google Scholar]
- Pitt, W.M.; Huang, R.; Trouillas, F.P.; Steel, C.C.; Savocchia, S. Evidence that Eutypa lata and other Diatrypaceous species occur in New South Wales vineyards. Australas. Plant Pathol. 2010, 39, 97–106. [Google Scholar] [CrossRef]
- Trouillas, F.P.; Gubler, W.D. Host range, biological variation, and phylogenetic diversity of Eutypa lata in California. Phytopathology 2010, 100, 1048–1056. [Google Scholar] [CrossRef] [Green Version]
- Navarrete, F.; Abreo, E.; Martínez, S.; Bettucci, L.; Lupo, S. Pathogenicity and molecular detection of Uruguayan isolates of Greeneria uvicola and Cadophora luteo-olivacea associated with grapevine trunk diseases. Phytopathol. Mediterr. 2011, 50, 166–175. [Google Scholar] [CrossRef]
- Nieder, G.; Reisenzein, H.; Berger, N. Esca in Austria. Phytopathol. Mediterr. 2000, 39, 26–34. [Google Scholar]
- Cardot, C.; Mappa, G.; La Camera, S.; Gaillard, C.; Vriet, C.; Lecomte, P.; Ferrari, G.; Coutos-Thévenot, P. Comparison of the molecular responses of tolerant, susceptible and highly susceptible grapevine cultivars during interaction with the pathogenic fungus Eutypa lata. Front. Plant Sci. 2019, 10, 991. [Google Scholar] [CrossRef] [Green Version]
- Schwappach, P.; Grimm, M. First detection of Eutypa lata (Pers.: Fr.) Tul. with PCR directly out of grapevine trunks in Germany. IOBC Wprs Bull. 2006, 29, 103. [Google Scholar]
- Kouyeas, H. Eutypa Armeniacae on Apricot and Grapevine in Greece. Phytopathol. Z. 1976. [Google Scholar]
- Cortesi, P.; Milgroom, M.G. Outcrossing and diversity of vegetative compatibility types in populations of Eutypa lata from grapevines. J. Plant Pathol. 2001, 83, 79–86. [Google Scholar]
- Rafaila, G.; Oprea, M. Eutypa disease of vine, a severe disease in Romania. In Analele Institutului de Cercetari Pentru Protectia Plantelor (Romania); FAO: Rome, Italy, 1985. [Google Scholar]
- Bolay, A.; Wj, M. Eutypa armeniacae Hansf & Carter, Agent d’un Grave Deperissement de Vignes en Production. Rev. Suisse Vitic Arboric Hortic 1977, 9, 241–251. [Google Scholar]
- Nakov, B. Eutypiosis (Eutypa armeniacae Hansf & Carter) on Grapes. 1986. Available online: https://scholar.google.com.hk/scholar?hl=zh-TW&as_sdt=0%2C5&scioq=Eutypiosis+%28Eutypa+armeniacae+Hansf.+%26+Carter%29+on+Grapes&q=Eutypiosis+%28Eutypa+armeniacae+Hansf.+%26+Carter%29+on+Grapes&btnG= (accessed on 12 February 2022).
- Ivić, D.; Cvjetković, B. Eutypa lata u kontekstu odumiranja marelice. Glas. Biljn. Zaštite 2005, 1, 321–324. [Google Scholar]
- Luque, J.; Garcia-Figueres, F.; Legorburu, F.J.; Muruamendiaraz, A.; Armengol, J.; Trouillas, F.P. Species of Diatrypaceae associated with grapevine trunk diseases in Eastern Spain. Phytopathol. Mediterr. 2012, 51, 528–540. [Google Scholar] [CrossRef]
- Moller, W.J.; Kasimatis, A.N.; Ramos, D.E.; English, W.H.; Bowers, K.W.; Kissler, J.J.; Rough, D.; Sanborn, R.R. Eutypa Dieback of Apricot and Grape in California; Leaflet-University of California: Berkeley, CA, USA, 1980. [Google Scholar]
- Lucas Tomaz, I.; Pereira Da Costa, M.E.A. Eutipiose e outras doenças do lenho da videira. An. Do Inst. Super. De Agron. 1983, 233, 1984. [Google Scholar]
- Delibašić, G.; Aćimović, S.; Gajić, S. Identifikacija Eutypa lata, parazita vinove loze. Pestic. I Fitomedicina Beogr. 2006, 21, 193–203. [Google Scholar]
- Šafránková, I. Poruchy, Poškození a Choroby Révy Vinné; Mendelova Zemědělská a Lesnická Univerzita: Brno, Czech Republic, 2007; ISBN 8073751003. [Google Scholar]
- Sosnowski, M.R.; Mundy, D.C. Pruning wound protection strategies for simultaneous control of eutypa and botryosphaeria dieback in New Zealand. Plant Dis. 2019, 103, 519–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, L.; Ruyi, L.; Hua, W.; Yuwen, C. Discovery and identify Eutypa dieback of grapevine in China. In Proceedings of the 5th International Symposium on Vine and Wine 2; Shaanxi People Publishing House: Xi’an, China, 2007. [Google Scholar]
- Mohammadi, H.; Banihashemi, Z.; Gramaje, D.; Armengol, J. Fungal pathogens associated with grapevine trunk diseases in Iran. JAST 2013, 15, 137–150. [Google Scholar]
- Ammad, F.; Benchabane, M.; Toumi, M. Diversity of fungal trunk pathogens associated with grapevine dieback of grapevine in Algeria. Jordan J. Biol. Sci. 2014, 7, 35–39. [Google Scholar] [CrossRef] [Green Version]
- Choueiri, E.; Jreijiri, F.; Chlela, P.; Mayet, V.; Comont, G.; Liminana, J.M.; Mostert, L.; Fischer, M.; Lecomte, P. Fungal community associated with grapevine wood lesions in Lebanon. J. Int. Des Sci. La Vigne Du Vin 2014, 48, 293–302. [Google Scholar] [CrossRef]
- Narmani, A.; Arzanlou, M. Quambalaria cyanescens, a new fungal trunk pathogen associated with grapevine decline in Iran. Crop Prot. 2019, 124, 104875. [Google Scholar] [CrossRef]
- Ari, M.E. A General Approach for Esca Disease in the Vineyards of Turkey. Phytopathol. Mediterr. 2000, 39, 35–37. [Google Scholar] [CrossRef]
- Larignon, P. Maladies Cryptogamiques du Bois de la Vigne: Symptomatologie et Agents Pathogen; Institut Français de la Vigne et du Vin, Grau du Roi dans le Gard: Le Grau-du-Roi, France, 2016; p. 165. [Google Scholar]
- Rolshausen, P.E.; Baumgartner, K.; Travadon, R.; Fujiyoshi, P.; Pouzoulet, J.; Wilcox, W.F. Identification of Eutypa spp. Causing Eutypa Dieback of Grapevine in Eastern North America. Plant Dis. 2014, 98, 483–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trouillas, F.P.; Gubler, W.D. Identification and characterization of Eutypa leptoplaca, a new pathogen of grapevine in Northern California. Mycol. Res. 2004, 108, 1195–1204. [Google Scholar] [CrossRef]
- Al-Momany, A. Identification of different Eutypa isolates from grapevine by using polymerase chain reaction. Arab J. Plant Prot. 2009, 27, 49. [Google Scholar]
- Catal, M.; Jordan, S.A.; Butterworth, S.C.; Schilder, A.M.C. Detection of Eutypa lata and Eutypella vitis in grapevine by nested multiplex polymerase chain reaction. Phytopathology 2007, 97, 737–747. [Google Scholar] [CrossRef] [Green Version]
- Octave, S.; Roblin, G.; Vachaud, M.; Fleurat-Lessard, P. Polypeptide metabolites secreted by the fungal pathogen Eutypa lata participate in Vitis vinifera cell structure damage observed in Eutypa dieback. Funct. Plant Biol. 2006, 33, 297–307. [Google Scholar] [CrossRef]
- Dubos, B. Les Maladies Cryptogamiques De La Vigne: Les Champignons Parasites Des Organes Herbacés Et Du Bois De La Vigne; Éditions Féret: Bordeaux, France, 2002; ISBN 2902416776. [Google Scholar]
- Amborabé, B.E.; Octave, S.; Roblin, G. Influence of temperature and nutritional requirements for mycelial growth of Eutypa lata, a vineyard pathogenic fungus. Comptes Rendus Biol. 2005, 328, 263–270. [Google Scholar] [CrossRef]
- Renaud, J.; Tsoupras, G.; Tabacchi, R. Biologically active natural acetylenic compounds from Eutypa lata (Pers: F.) TUL. Helv. Chim. Acta 1989, 72, 929–932. [Google Scholar] [CrossRef]
- Fallot, J.; Deswarte, C.; Dalmayrac, S.; Colrat, S.; Roustan, J.-P. L’Eutypiose de la vigne: Isolement d’une molécule synthétisée par Eutypa lata et toxique pour la vigne. Comptes Rendus L’académie Des Sci. III-Sci. La Vie 1997, 320, 149–158. [Google Scholar] [CrossRef]
- Deswarte, C.; Canut, H.; Klaebe, A.; Roustan, J.-P.; Fallot, J. Transport, cytoplasmic accumulation and mechanism of action of the toxin eutypine in Vitis vinifera cells. J. Plant Physiol. 1996, 149, 336–342. [Google Scholar] [CrossRef]
- Lardner, R.; Scott, E.S.; Stummer, B.E. Genetic variation in Australian isolates of the grapevine pathogen Eutypa lata Australas. Plant Pathol. 2007, 36, 149–156. [Google Scholar]
- Živković, S.; Vasić, T.; Ivanović, M.; Jevremović, D.; Marković, J.; Trkulja, V. Morphological and molecular identification of Eutypa lata on grapevine in Serbia. J. Plant Dis. Prot. 2019, 126, 479–483. [Google Scholar] [CrossRef]
- Lecomte, P.; Péros, J.-P.; Blancard, D.; Bastien, N.; Délye, C. PCR Assays that identify the grapevine dieback Funguseutypa Lata. Appl. Environ. Microbiol. 2000, 66, 4475–4480. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.; Fragoeiro, S.; Oliveira, H.; Phillips, A. Response of Vitis vinifera L. plants inoculated with Phaeoacremonium angustius and Phaeomoniella chlamydospora to thiabendazole, resveratrol and sodium arsenite. Sci. Hortic. 2006, 107, 131–136. [Google Scholar] [CrossRef]
- Mugnai, L.; Graniti, A.; Surico, G. Esca (Black measles) and brown wood-streaking: Two old and elusive diseases of grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viala, P. L’apoplexie. Bulletin de la Société de Patologie Vegétalé de France IX PV des Scences de Janvier-Mars; Gallica: Aussois, France, 1922. [Google Scholar]
- Pacottet-Brin, P. Folletage. Rev. Vit 1899, 11, 672–673. [Google Scholar]
- Ravaz, L. Sur le Folletage. Rev. Vitic. 1898, 10, 184–186. [Google Scholar]
- Galet, A. Précis De Pathologie Viticole, 3rd ed.; Pierre Galet: Montpellier, France, 1999. [Google Scholar]
- Harms, M. Aktueller stand zur esca-situation. Weinbautage 2008, 61, 49–51. [Google Scholar]
- Nikolova, E. Esca disease of grapevine-symptoms, causal organisms and methods for control. Rast. Nauk. 2010, 47, 195–205. [Google Scholar]
- Cvjetković, B.; Ivić, D. Esca-apoplektično venuće starih i mladih čokota vinove loze. Glas. Biljn. Zaštite 2004, 6, 343–349. [Google Scholar]
- Sánchez-Torres, P.; Hinarejos, R.; González, V.; Tuset, J.J. Identification and characterization of fungi associated with esca in vineyards of the comunidad Valenciana (Spain). Span. J. Agric. Res. 2008, 6, 650–660. [Google Scholar] [CrossRef] [Green Version]
- Péros, J.; Berger, G.; Jamaux-Despréaux, I. Symptoms, wood lesions and fungi associated with Esca in organic vineyards in Languedoc-Roussillon (France). J. Phytopathol. 2008, 156, 297–303. [Google Scholar] [CrossRef]
- Rumbos, I.; Rumbou, A. Fungi associated with esca and young grapevine decline in Greece. Phytopathol. Mediterr. 2001, 40, S330–S335. [Google Scholar] [CrossRef]
- Rábai, A.; Dula, T.; Mugnai, L. Distribution of esca disease in Hungary and the pathogens causing the syndrome. Acta Phytopathol. Entomol. Hung. 2008, 43, 45–54. [Google Scholar] [CrossRef]
- Essakhi, S.; Mugnai, L.; Crous, P.W.; Groenewald, J.Z.; Surico, G. Molecular and phenotypic characterisation of novel Phaeoacremonium species isolated from esca diseased grapevines. Pers. Mol. Phylogeny Evol. Fungi 2008, 21, 119. [Google Scholar] [CrossRef] [Green Version]
- Latinović, N.; Vucinic, Z.; Latinović, J. Phaeoacremonium aleophilum one of the causal agents of esca disease of grapevine in Montenegro Soko Banja. In Proceedings of the 7th Conference on Plant Protection, Soko Banja, Serbia, 15–18 November 2005; pp. 125–126. [Google Scholar]
- Sofia, J.; Oliveira, H.; Gonçalves, M.T. Spatial Distribution of Esca Symptomatic Plants in Dão Vineyards (Centre Portugal) and Isolation of Associated Fungi; Firenze University Press: Florence, Italy, 2006; pp. 1000–1006. [Google Scholar]
- Matei, P.; Iacomi, B.; Dragan, G. Fungi associated with esca decline and their in vitro control by chitosan. Sci. Pap. UASVM Buchar. Ser. A 2010, 53, 448–453. [Google Scholar]
- Srđana, A.; Goran, D.; Schilderc, A.; Brankicad, T. Eutipoza i ESCA bolesti odumiranja i sušenja čokota vinove loze u Srbiji. Biljn. Lek 2008, 36, 94–103. [Google Scholar]
- Kakalíková, L.; Jankura, E. Esca—Disease of trunk on vineyard in Slovakia. Vinohrad 2001, 39, 39–1113. [Google Scholar]
- Bukovec, M.; Celar, F.; Valic, N. Kap vinske trte in Nejni Možni povzročitelji v vinorodni deželi primorska. Zb. Pred. Ref. Slov. Posvetovanja O Varstvu Rastl. Zreče 2005, 7, 362–366. [Google Scholar]
- Hofstetter, V.; Casieri, L.; Viret, O.; Gindro, K. Esca de la vigne et communauté fongique. Rev. Suisse Vitic. Arboric. Hortic. 2009, 41, 247–253. [Google Scholar]
- Ackermann, P. Chřadnutí a odumírání révy (esca, chřadnutí mladých keřů révy). Minist. Zemědělství 2009, 1, 12–13. [Google Scholar]
- Shmatkovskaya, E. Disease of long term timber vineyards in the south of Ukraine and peculiarities of their development. In Proceedings of the International Scientific and Practical Conference, Lviv, Ukraine, 15 August 2013; pp. 228–232. [Google Scholar]
- Rooney-Latham, S.; Eskalen, A.; Gubler, W.D. Occurrence of Togninia minima perithecia in esca-affected vineyards in California. Plant Dis. 2005, 89, 867–871. [Google Scholar] [CrossRef] [Green Version]
- O’Gorman, D.T.; Haag, P.; Sholberg, P.L. News diseases causing decline of wine grapes in the Okanagan valley. Can. Plant Dis. Surv. Can. Phytopathol. Soc. 2009, 90, 140–143. [Google Scholar]
- Morales-Pedraza, G.; Valenzuela-Solano, C.; Hernandez-Martinez, R. Characterization of fungi associated to Esca disease in Baja California, Mexico. In Proceedings of the 8th International Workshop on Grapevine Trunk Diseases, Valencia, Spain, 18–21 June 2012. [Google Scholar]
- Bahrabadi, M.; Karimi Shahri, M.R.; Hashemi, M. Study on genetic variability in fungi associated with esca disease in North Khorasan province vineyards with RAPD-PCR. J. Plant Prot. 2012, 26, 92–100. [Google Scholar]
- Mostert, L.; Groenewald, J.Z.; Summerbell, R.C.; Robert, V.; Sutton, D.A.; Padhye, A.A.; Crous, P.W. Species of Phaeoacremonium associated with infections in humans and environmental reservoirs in infected woody plants. J. Clin. Microbiol. 2005, 43, 1752–1767. [Google Scholar] [CrossRef] [Green Version]
- Poyraz, D.; Onoğur, E. Bağlarda Kav ve Petri Hastalıkları. Anadolu Ege Tarımsal Araştırma Enstitüsü Derg. 2012, 22, 69–81. [Google Scholar]
- White, C.-L.; Halleen, F.; Mostert, L. Symptoms and fungi associated with esca in South African vineyards. Phytopathol. Mediterr. 2011, 50, S236–S246. [Google Scholar]
- Sbaghi, M. Agriculture du Maghreb n50 Mars 2011; Agriculture du Maghreb: Casablanca, Morocco, 2011; pp. 78–84. [Google Scholar]
- Surico, G. Towards a redefinition of the diseases within the esca complex of grapevine. Phytopathol. Mediterr. 2009, 48, 5–10. [Google Scholar]
- Gramaje, D.; Úrbez-Torres, J.R.; Sosnowski, M.R. Managing Grapevine Trunk Diseases With Respect to Etiology and Epidemiology: Current Strategies and Future Prospects. Plant Dis. 2018, 102, 12–39. [Google Scholar] [CrossRef] [Green Version]
- Sidoti, A.; Mugnai, L.; Serges, T.; Buonocore, E. Decline of Young Grapevines Associated with “Phaeoacremonium chlamydosporum” in Sicily (Italy). Phytopathol. Mediterr. 2000, 12, 1000–1005. [Google Scholar]
- Dupont, J.; Laloui, W.; Roquebert, M.F. Partial ribosomal DNA sequences show an important divergence between Phaeoacremonium species isolated from Vitis vinifera. Mycol. Res. 1998, 102, 631–637. [Google Scholar] [CrossRef]
- Larignon, P.; Dupont, J.; Dubos, B. Esca disease. The biological background of two agents of the disease, Phaeoacremonium aleophilum and Phaeomoniella chlamydospora. Phytoma 2000, 527, 30–35. [Google Scholar]
- Larignon, P. Contribution à L’identification et au Mode D’action des Champignons Associés au Syndrome de L’esca de la Vigne 1991. Available online: http://www.theses.fr/1991BOR22015 (accessed on 12 May 2022).
- Marchi, G.; Mugnai, L.; Surico, G.; Roberti, S.; D’Ovidio, R. Pectic enzymes production by Phaeomoniella chlamydospora [Vitis vinifera L.-esca-grapevine decline]. Phytopathol. Mediterr. 2001, 40, 1000–1010. [Google Scholar]
- Sparapano, L.; Bruno, G.; Graniti, A. Effects on plants of metabolites produced in culture by Phaeoacremonium chlamydosporum, P. aleophilum and Fomitiporia punctata. Phytopath. Medit. 2000, 12, 169–177. [Google Scholar] [CrossRef]
- Crous, P.W.; Gams, W.; Wingfield, M.J.; Van Wyk, P.S. Phaeoacremonium gen. nov. associated with wilt and decline diseases of woody hosts and human infections. Mycologia 1996, 88, 786–796. [Google Scholar] [CrossRef] [Green Version]
- Gramaje, D.; Armengol, J.; Mohammadi, H.; Banihashemi, Z.; Mostert, L. Novel Phaeoacremonium species associated with Petri disease and esca of grapevine in Iran and Spain. Mycologia 2009, 101, 920–929. [Google Scholar] [CrossRef]
- Gubler, W.D.; Thind, T.; Feliciano, A.; Eskalen, A. Pathogenicity of Phaeoacremonium aleophilum and Phaeomoniella chlamydospora on grape berries in California. Phytopathol. Mediterr. 2004, 43, 4. [Google Scholar]
- Ciccarone, C.; Graniti, A.; Schiaffino, A.; Marras, F. Molecular analysis of Fomitiporia mediterranea isolates from esca-affected grapevines in southern Italy. Phytopathol. Mediterr. 2004, 43, 268–272. [Google Scholar]
- Fisher, K.; Phillips, C. Potential antimicrobial uses of essential oils in food: Is citrus the answer? Trends Food Sci. Technol. 2008, 19, 156–164. [Google Scholar] [CrossRef]
- Tabacchi, R.; Fkyerat, A.; Poliart, C.; Dubin, G.-M. Phytotoxins from fungi of esca of grapevine. Phytopathol. Mediterr. 2000, 39, 156–161. [Google Scholar]
- Pramod, S.; Mesara, S.; Rao, K.S.; Rajput, K.S. Dieback of Gmelina arborea Trees and Structural Alterations Induced in the Wood Cell Walls by Alternaria alternata. Forestist 2021, 72, 2–9. [Google Scholar] [CrossRef]
- Reynier, A. Manuel de Viticulture, 9th ed.; TecDoc: Cachan, France, 2003. [Google Scholar]
- Surico, G. Epidemiology of Esca in Some Vineyards in Tuscany (Italy); Firenze University Press: Florence, Italy, 2000; pp. 190–205. [Google Scholar]
- Valtaud, C.; Thibault, F.; Larignon, P.; Bertsch, C.; Fleurat-Lessard, P.; Bourbouloux, A. Systemic damage in leaf metabolism caused by esca infection in grapevines. Aust. J. Grape Wine Res. 2011, 17, 101–110. [Google Scholar] [CrossRef]
- Levasseur-Garcia, C.; Malaurie, H.; Mailhac, N. An infrared diagnostic system to detect causal agents of grapevine trunk diseases. J. Microbiol. Methods 2016, 131, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbez-Torres, J.R.; Peduto, F.; Striegler, R.K.; Urrea-Romero, K.E.; Rupe, J.C.; Cartwright, R.D.; Gubler, W.D. Characterization of fungal pathogens associated with grapevine trunk diseases in Arkansas and Missouri. Fungal Divers. 2012, 52, 169–189. [Google Scholar] [CrossRef]
- Letousey, P.; Baillieul, F.; Perrot, G.; Rabenoelina, F.; Boulay, M.; Vaillant-Gaveau, N.; Clément, C.; Fontaine, F. Early events prior to visual symptoms in the apoplectic form of grapevine esca disease. Phytopathology 2010, 100, 424–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecomte, P.; Diarra, B.; Carbonneau, A.; Patrice, R.E.Y.; Chevrier, C. Esca of grapevine and training practices in France: Results of a 10-year survey. Phytopathol. Mediterr. 2018, 57, 472–487. [Google Scholar]
- Bruez, E.; Haidar, R.; Alou, M.T.; Vallance, J.; Bertsch, C.; Mazet, F.; Fermaud, M.; Deschamps, A.; Guerin-Dubrana, L.; Compant, S.; et al. Bacteria in a wood fungal disease: Characterization of bacterial communities in wood tissues of esca-foliar symptomatic and asymptomatic grapevines. Front. Microbiol. 2015, 6, 1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, R.C.; Goheen, A.C. Compendium of Grape Diseases; APS Press: St. Paul, MN, USA, 1988. [Google Scholar]
- Cristinzio, G. Gravi attacchi di Botryosphaeria obtusa su vite in provincia di Isernia. Inf. Fitopatol. 1978, 6, 23–25. [Google Scholar]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine trunk diseases: A review of fifteen years of trials for their control with chemicals and biocontrol agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef] [Green Version]
- Auger, J.; Esterio, M.; Ricke, G.; Pérez, I. Black Dead Arm and Basal Canker of Vitis vinifera cv. Red Globe Caused by Botryosphaeria obtusa in Chile. Plant Dis. 2004, 88, 1286. [Google Scholar] [CrossRef]
- Tuset, J.J. Conidial State Activity of Botryosphaeria obtusa on Gravepines in the Southeast of Spain. 1980. Available online: https://scholar.google.co.uk/scholar?hl=zh-TW&as_sdt=0%2C5&q=Conidial+State+Activity+of+Botryosphaeria+obtusa+on+Gravepines+in+the+Southeast+of+Spain.&btnG= (accessed on 12 May 2022).
- Arzanlou, M.; Moshari, S.; Bakhshi, M.; Khodaie, S. Botryosphaeria dothidea associated with grapevine decline disease in Iran. Australas. Plant Dis. Notes 2012, 7, 197–200. [Google Scholar] [CrossRef] [Green Version]
- Rovesti, L.; Montermini, A. Un deperimento della vite causato da Sphaeropsis malorum diffuso in provincia di Reggio Emilia. Inf. Fitopatol. 1987, 1, 59–61. [Google Scholar]
- Choueiri, E.; Jreijiri, F.; Chlela, P.; Louvet, G.; Lecomte, P. Occurrence of grapevine declines and first report of Black Dead Arm associated with Botryosphaeria obtusa in Lebanon. Plant Dis. 2006, 90, 115. [Google Scholar] [CrossRef]
- Rego, C.; Vaz, A.; Nascimento, T.; Cabral, A.; Oliveira, H. Diseases incited by Botryosphaeriaceae fungi in Portuguese vineyards. In Proceedings of the Phytopathologia Mediterranea; Mediterranean Phytopathological Union C/O Dipartimento Di Biotecnologie: Florence, Italy, 2009; Volume 48, p. 181. [Google Scholar]
- Akgul, D.S.; Savas, N.G.; Eskalen, A. First report of wood canker caused by Botryosphaeria dothidea, Diplodia seriata, Neofusicoccum parvum, and Lasiodiplodia theobromae on grapevine in Turkey. Plant Dis. 2014, 98, 568. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R. The status of Botryosphaeriaceae species infecting grapevines. Phytopathol. Mediterr. 2015, 54, 241–252. [Google Scholar] [CrossRef]
- Copes, W.E.; Fruit, S.; Hendrix, F.F. Effect of Temperature on Sporulation of Botryosphaeria dothidea, B. obtusa, and B. rhodina. Plant Dis. 2014, 88, 292–296. [Google Scholar] [CrossRef] [Green Version]
- Michailides, T.J.; Morgan, D.P. Spore release by Botryosphaeria dothidea in pistachio orchards and disease control by altering the trajectory angle of sprinklers. Phytopathology 1993, 83, 145–152. [Google Scholar] [CrossRef]
- Michailides, T.J. Pathogenicity, distribution, sources of inoculum, and infection courts of Botryosphaeria dothidea on pistachio. Phytopathology 1991, 81, 566–573. [Google Scholar] [CrossRef]
- Pusey, P.L.; Bertrand, P.F. Seasonal infection of nonwounded peach bark by Botryosphaeria dothidea. Phytopathology 1993, 83, 825–829. [Google Scholar] [CrossRef]
- Savocchia, S.; Laurent, E.N.; Stodart, B.J.; Steel, C.C. Botryosphaeria canker and sensitivity to fungicides in vitro. In Proceedings of the 43rd Annual Congress of the South African Society for Plant Pathology, Mossel Bay, South Africa, 23–26 January 2005; p. 88. [Google Scholar]
- Larignon, P.; Dubos, B. The villainy of Black Dead Arm. Wines Vines 2001, 82, 86–89. [Google Scholar]
- Fourie, P.H.; Halleen, F. Occurrence of grapevine trunk disease pathogens in rootstock mother plants in South Africa. Australas. Plant Pathol. 2004, 33, 313–315. [Google Scholar] [CrossRef]
- Aroca, A.; García-Figueres, F.; Bracamonte, L.; Luque, J.; Raposo, R. A survey of trunk disease pathogens within rootstocks of grapevines in Spain. Eur. J. Plant Pathol. 2006, 115, 195–202. [Google Scholar] [CrossRef]
- Baskarathevan, J.; Jaspers, M.V.; Jones, E.E.; Ridgway, H.J. Use of endogenous molecular markers to measure rain water splash dispersal of Neofusicoccum species in the vineyard. Phytopathol. Mediterr. 2010, 49, 107. [Google Scholar]
- Váczy, K.Z.; Németh, M.Z.; Csikós, A.; Kovács, G.M.; Kiss, L. Dothiorella omnivora isolated from grapevine with trunk disease symptoms in Hungary. Eur. J. Plant Pathol. 2018, 150, 817–824. [Google Scholar] [CrossRef]
- Séverine, M.; Coralie, L.; Pascal, L.; Marc, B.; Jean-Philippe, R. Impact of grafting type on Esca foliar symptoms. Oeno One 2017, 51, 221–230. [Google Scholar] [CrossRef]
- Kovács, C.; Balling, P.; Bihari, Z.; Nagy, A.; Sándor, E. Incidence of grapevine trunk diseases is influenced by soil, topology and vineyard age, but not by Diplodia seriata infection rate in the Tokaj Wine Region, Hungary. Phytoparasitica 2017, 45, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Adam, A. La vigne. In In Vino Veritas; Presses de l’Université de Montréal: Montreal, QC, Canada, 2018; pp. 13–14. [Google Scholar]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Masi, M.; Cimmino, A.; Reveglia, P.; Mugnai, L.; Surico, G.; Evidente, A. Advances on Fungal Phytotoxins and Their Role in Grapevine Trunk Diseases. J. Agric. Food Chem. 2018, 66, 5948–5958. [Google Scholar] [CrossRef]
- Venkatasubbaiah, P.; Sutton, T.B.; Chilton, W.S. Effect of phytotoxins produced by Botryosphaeria obtusa, the cause of black rot of apple fruit and frogeye leaf spot. Phytopathology 1991, 81, 243–247. [Google Scholar] [CrossRef]
- Djoukeng, J.D.; Polli, S.; Larignon, P.; Abou-Mansour, E. Identification of phytotoxins from Botryosphaeria obtusa, a pathogen of black dead arm disease of grapevine. Eur. J. Plant Pathol. 2009, 124, 303–308. [Google Scholar] [CrossRef] [Green Version]
- Kuntzmann, P.; Vuillaume, S.; Bertsch, C. Dispersal of Botryosphaeria spp. Conidia in vineyards. In Proceedings of the 6th International Workshop on Grapevine Trunk Diseases, Florence, Italy, 1–3 September 2008. [Google Scholar]
- Rolshausen, P.E.; Trouillas, F.; Gubler, W.D. Identification of Eutypa lata by PCR-RFLP. Plant Dis. 2004, 88, 925–929. [Google Scholar] [CrossRef]
- Lardner, R.; Stummer, B.E.; Sosnowski, M.R.; Scott, E.S. Molecular identification and detection of Eutypa lata in grapevine. Mycol. Res. 2005, 109, 799–808. [Google Scholar] [CrossRef]
- Pilotti, M.; Gervasi, F.; Brunetti, A. Molecular identification of Fomitiporia mediterranea and Eutypa lata/Libertella blepharis in Platanus× acerifolia. J. Phytopathol. 2005, 153, 193–202. [Google Scholar] [CrossRef]
- Gramaje, D.; Alaniz, S.; Pérez-Sierra, A.; Abad-Campos, P.; García-Jiménez, J.; Armengol, J. First Report of Phaeoacremonium mortoniae Causing Petri Disease of Grapevine in Spain. Plant Dis. 2007, 91, 1206. [Google Scholar] [CrossRef]
- Makaradze, E.; Meparishvili, G.; Varshanidze, N.; Diasamidze, I.; Dolidze, K.; Jakeli, E.; Zarnadze, N. RAPD-analysis of Cyclamen spp. genome polymorphism. CBU Int. Conf. Proc. 2019, 7, 949–953. [Google Scholar] [CrossRef] [Green Version]
- Pollastro, S.; Dongiovanni, C.; Abbatecola, A.; De Guido, M.A.; De Miccolis Angelini, R.M.; Faretra, F. Specific SCAR primers for fungi associated with wood decay of grapevine. Phytopathol. Mediterr. 2001, 40, 362–368. [Google Scholar] [CrossRef]
- Martini, C.; Mari, M. Monilinia Fructicola, Monilinia Laxa (Monilinia Rot, Brown Rot); Elsevier: Amsterdam, The Netherlands, 2014; ISBN 9780124115682. [Google Scholar]
- Moisy, C.; Berger, G.; Flutre, T.; Le Cunff, L.; Péros, J.P. Quantitative assessment of grapevine wood colonization by the dieback fungus Eutypa lata. J. Fungi 2017, 3, 21. [Google Scholar] [CrossRef] [Green Version]
- Tegli, S.; Surico, G.; Bertelli, E. Sequence Analysis of ITS Ribosomal DNA in Five “Phaeoacremonium” Species and Development of a PCR-Based Assay for the Detection of “P. chlamydosporum” and “P. aleophilum” in Grapevine Tissue. Phytopathol. Mediterr. 2000, 1000–1016. [Google Scholar] [CrossRef]
- Fischer, M. A new wood-decaying basidiomycete species associated with esca of grapevine: Fomitiporia mediterranea (Hymenochaetales). Mycol. Prog. 2002, 1, 315–324. [Google Scholar] [CrossRef]
- Billones-Baaijens, R.; Úrbez-Torres, J.R.; Liu, M.; Ayres, M.; Sosnowski, M.; Savocchia, S. Molecular methods to detect and quantify botryosphaeriaceae inocula associated with grapevine dieback in Australia. Plant Dis. 2018, 102, 1489–1499. [Google Scholar] [CrossRef] [Green Version]
- Mackay, I.M. Real-time PCR in the microbiology laboratory. Clin. Microbiol. Infect. 2004, 10, 190–212. [Google Scholar] [CrossRef] [Green Version]
- Espy, M.J.; Uhl, J.R.; Sloan, L.M.; Buckwalter, S.P.; Jones, M.F.; Vetter, E.A.; Yao, J.D.C.; Wengenack, N.L.; Rosenblatt, J.E.; Cockerill, F.R. 3 Real-time PCR in clinical microbiology: Applications for routine laboratory testing. Clin. Microbiol. Rev. 2006, 19, 165–256. [Google Scholar] [CrossRef] [Green Version]
- Lau, A.; Chen, S.; Sleiman, S.; Sorrell, T. Current status and Future perspectives on molecular and serological methods in diagnostic mycology. Future Microbiol. 2009, 4, 1185–1222. [Google Scholar] [CrossRef]
- Pouzoulet, J.; Rolshausen, P.E.; Schiavon, M.; Bol, S.; Travadon, R.; Lawrence, D.P.; Baumgartner, K.; Ashworth, V.E.; Comont, G.; Corio-Costet, M.-F.; et al. A method to detect and quantify Eutypa lata and Diplodia seriata -Complex DNA in grapevine pruning wounds. Plant Dis. 2017, 101, 1470–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armijo, G.; Schlechter, R.; Agurto, M.; Muñoz, D.; Nuñez, C.; Arce-johnson, P. Grapevine pathogenic microorganisms: Understanding infection strategies and host response scenarios. Front. Plant Sci. 2016, 7, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brederode, F.T.; Linthorst, H.J.M.; Bol, J.F. Differential induction of acquired resistance and PR gene expression in tobacco by virus infection, ethephon treatment, UV light and wounding. Plant Mol. Biol. 1991, 17, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C. Occurrence and properties of plant pathogenesis-related proteins. In Pathogenesis-Related Proteins in Plants; CRC: Boca Raton, FL, USA, 1999; pp. 1–19. [Google Scholar]
- Kan, J.A.L. Van Licensed to kill: The lifestyle of a necrotrophic plant pathogen. Trends Plant Sci. 2006, 11, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef]
- Turner, J.G.; Ellis, C.; Devoto, A. The jasmonate signal pathway. Plant Cell 2002, 14, S153–S164. [Google Scholar] [CrossRef] [Green Version]
- Glazebrook, J. Against biotrophic and necrotrophic. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Yalpani, N.; Silverman, P.; Wilson, T.M.; Kleier, D.A.; Raskin, I. Salicylic acid is a systemic signal and an inducer of pathogenesis-related proteins in virus-infected tobacco. Plant Cell 1991, 3, 809–818. [Google Scholar]
- Sinha, M.; Singh, R.P.; Kushwaha, G.S.; Iqbal, N.; Singh, A.; Kaushik, S.; Kaur, P.; Sharma, S.; Singh, T.P. Current overview of allergens of plant pathogenesis related protein families. Sci. World J. 2014, 2014, 543195. [Google Scholar] [CrossRef] [Green Version]
- Chamnongpol, S.; Willekens, H.; Moeder, W.; Langebartels, C.; Sandermann, H.; Van Montagu, M.; Inzé, D.; Van Camp, W. Defense activation and enhanced pathogen tolerance induced by H2O2 in transgenic tobacco. Proc. Natl. Acad. Sci. USA 1998, 95, 5818–5823. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.I.; Chang, P.-F.L.; Liu, D.; Narasimhan, M.L.; Raghothama, K.G.; Hasegawa, P.M.; Bressan, R.A. Plant defense genes are synergistically induced by ethylene and methyl jasmonate. Plant Cell 1994, 6, 1077–1085. [Google Scholar] [CrossRef] [Green Version]
- Monaghan, J.; Zipfel, C. Plant pattern recognition receptor complexes at the plasma membrane. Curr. Opin. Plant Biol. 2012, 15, 349–357. [Google Scholar] [CrossRef]
- Jonathan, D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323–329. [Google Scholar]
- Hayward, A.P.; Tsao, J.; Dinesh-Kumar, S.P. Autophagy and plant innate immunity: Defense through degradation. In Proceedings of the Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2009; Volume 20, pp. 1041–1047. [Google Scholar]
- Van Loon, L.C.; Van Strien, E.A. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Nielsen, K.K.; Mikkelsen, J.D.; Kragh, K.M.; Bojsen, K. An acidic class III chitinase in sugar beet: Induction by Cercospora beticola, characterization, and expression in transgenic tobacco plants. Mol. Plant Microbe Interact. 1993, 6, 495. [Google Scholar] [CrossRef]
- Anguelova-Merhar, V.S.; VanDer Westhuizen, A.J.; Pretorius, Z.A. β-1, 3-glucanase and chitinase activities and the resistance response of wheat to leaf rust. J. Phytopathol. 2001, 149, 381–384. [Google Scholar] [CrossRef]
- Brogue, K.; Chet, I.; Holliday, M.; Cressman, R.; Biddle, P.; Knowlton, S.; Mauvais, C.J.; Broglie, R. Transgenic plants with enhanced resistance to the fungal pathogen Rhizoctonia solani. Science 1991, 254, 1194–1197. [Google Scholar] [CrossRef]
- Yamamoto, T.; Iketani, H.; Ieki, H.; Nishizawa, Y.; Notsuka, K.; Hibi, T.; Hayashi, T.; Matsuta, N. Transgenic grapevine plants expressing a rice chitinase with enhanced resistance to fungal pathogens. Plant Cell Rep. 2000, 19, 639–646. [Google Scholar] [CrossRef]
- Datta, K.; Velazhahan, R.; Oliva, N.; Ona, I.; Mew, T.; Khush, G.S.; Muthukrishnan, S.; Datta, S.K. Over-expression of the cloned rice thaumatin-like protein (PR-5) gene in transgenic rice plants enhances environmental friendly resistance to Rhizoctonia solani causing sheath blight disease. Theor. Appl. Genet. 1999, 98, 1138–1145. [Google Scholar] [CrossRef]
- Rohini, V.K.; Rao, K.S. Transformation of peanut (Arachis hypogaea L.) with tobacco chitinase gene: Variable response of transformants to leaf spot disease. Plant Sci. 2001, 160, 889–898. [Google Scholar] [CrossRef]
- Pan, S.-Q.; Ye, X.-S.; Kuć, J. Direct detection of β-1, 3-glucanase isozymes on polyacrylamide electrophoresis and isoelectrofocusing gels. Anal. Biochem. 1989, 182, 136–140. [Google Scholar] [CrossRef]
- Adams, D.J. Fungal cell wall chitinases and glucanases. Microbiology 2004, 150, 2029–2035. [Google Scholar] [CrossRef]
- Klarzynski, O.; Plesse, B.; Joubert, J.-M.; Yvin, J.-C.; Kopp, M.; Kloareg, B.; Fritig, B. Linear β-1, 3 glucans are elicitors of defense responses in tobacco. Plant Physiol. 2000, 124, 1027–1038. [Google Scholar] [CrossRef] [Green Version]
- Camps, C.; Kappel, C.; Lecomte, P.; Léon, C.; Gomès, E.; Coutos-Thévenot, P.; Delrot, S. A transcriptomic study of grapevine (Vitis vinifera cv. Cabernet-Sauvignon) interaction with the vascular ascomycete fungus Eutypa lata. J. Exp. Bot. 2010, 61, 1719–1737. [Google Scholar] [CrossRef]
- Mutawila, C.; Stander, C.; Halleen, F.; Vivier, M.A.; Mostert, L. Response of Vitis vinifera cell cultures to Eutypa lata and Trichoderma atroviride culture filtrates: Expression of defence-related genes and phenotypes. Protoplasma 2017, 254, 863–879. [Google Scholar] [CrossRef]
- La Camera, S.; Gouzerh, G.; Dhondt, S.; Hoffmann, L.; Fritig, B.; Legrand, M.; Heitz, T. Metabolic reprogramming in plant innate immunity: The contributions of phenylpropanoid and oxylipin pathways. Immunol. Rev. 2004, 198, 267–284. [Google Scholar] [CrossRef]
- Piasecka, A.; Jedrzejczak-Rey, N.; Bednarek, P. Secondary metabolites in plant innate immunity: Conserved function of divergent chemicals. New Phytol. 2015, 206, 948–964. [Google Scholar] [CrossRef] [PubMed]
- Langcake, P.; Pryce, R.J. The production of resveratrol by Vitis vinifera and other members of the Vitaceae as a response to infection or injury. Physiol. Plant Pathol. 1976, 9, 77–86. [Google Scholar] [CrossRef]
- Melchior, F.; Kindl, H. Coordinate-and elicitor-dependent expression of stilbene synthase and phenylalanine ammonia-lyase genes in Vitis cv. Optima. Arch. Biochem. Biophys. 1991, 288, 552–557. [Google Scholar] [CrossRef]
- Fritzemeier, K.-H.; Kindl, H. Coordinate induction by UV light of stilbene synthase, phenylalanine ammonia-lyase and cinnamate 4-hydroxylase in leaves of Vitaceae. Planta 1981, 151, 48–52. [Google Scholar] [CrossRef]
- Adrian, M.; Jeandet, P.; Veneau, J.; Weston, L.A.; Bessis, R. Biological activity of resveratrol, a stilbenic compound from grapevines, against Botrytis cinerea, the causal agent for gray mold. J. Chem. Ecol. 1997, 23, 1689–1702. [Google Scholar] [CrossRef]
- Coutos-Thévenot, P.; Poinssot, B.; Bonomelli, A.; Yean, H.; Breda, C.; Buffard, D.; Esnault, R.; Hain, R.; Boulay, M. In vitro tolerance to Botrytis cinerea of grapevine 41B rootstock in transgenic plants expressing the stilbene synthase Vst 1 gene under the control of a pathogen-inducible PR 10 promoter. J. Exp. Bot. 2001, 52, 901–910. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, F.; Pinto, C.; Vallet, J.; Clément, C.; Gomes, A.C.; Spagnolo, A. The effects of grapevine trunk diseases (GTDs) on vine physiology. Eur. J. Plant Pathol. 2016, 144, 707–721. [Google Scholar] [CrossRef]
- Gomès, E.; Coutos-Thévenot, P. Molecular aspects of grapevine-pathogenic fungi interactions. In Grapevine Molecular Physiology & Biotechnology; Springer: Berlin/Heidelberg, Germany, 2009; pp. 407–428. [Google Scholar]
- Loon, L.C.V.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C. French-Italian public consortium for grapevine genome characterization The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar]
- Robert, N.; Roche, K.; Lebeau, Y.; Breda, C.; Boulay, M.; Esnault, R.; Buffard, D. Expression of grapevine chitinase genes in berries and leaves infected by fungal or bacterial pathogens. Plant Sci. 2002, 162, 389–400. [Google Scholar] [CrossRef]
- Jacobs, A.K.; Dry, I.B.; Robinson, S.P. Induction of different pathogenesis-related cDNAs in grapevine infected with powdery mildew and treated with ethephon. Plant Pathol. 1999, 48, 325–336. [Google Scholar] [CrossRef]
- Robert, N.; Ferran, J.; Breda, C.; Coutos-Thévenot, P.; Boulay, M.; Buffard, D.; Esnault, R. Molecular characterization of the incompatible interaction of Vitis vinifera leaves with Pseudomonas syringae pv. pisi: Expression of genes coding for stilbene synthase and class 10 PR protein. Eur. J. Plant Pathol. 2001, 107, 249–261. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.D.C.; Santoyo, G. Plant-microbial endophytes interactions: Scrutinizing their beneficial mechanisms from genomic explorations. Curr. Plant Biol. 2021, 25, 100189. [Google Scholar] [CrossRef]
- Pawlowski, M.L.; Hartman, G.L. Infection mechanisms and colonization patterns of fungi associated with soybean. Fungal Pathog. 2016, 12, 25–42. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Wei, T.; Lou, H.; Shu, X. A Critical review on communication mechanism within plant-endophytic fungi interactions to cope with biotic and abiotic stresses. J. Fungi 2021, 7, 719. [Google Scholar] [CrossRef]
- Pradhan, A.; Ghosh, S.; Sahoo, D.; Jha, G. Fungal effectors, the double edge sword of phytopathogens. Curr. Genet. 2021, 67, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Marhadour, S.; Wu, H.; Yang, W.; Marivingt-Mounir, C.; Bonnemain, J.L.; Chollet, J.F. Vectorisation of agrochemicals via amino acid carriers: Influence of the spacer arm structure on the phloem mobility of phenylpyrrole conjugates in the Ricinus system. Pest Manag. Sci. 2017, 73, 1972–1982. [Google Scholar] [CrossRef]
- Xiao, Y.; Li, J.; Xiao, Y.; Li, J. Research progress on molecular design and conduction distribution mechanism of guided pesticide. Chin. J. Pestic. Sci. 2021, 23, 438–445. [Google Scholar] [CrossRef]
- Leal, C.; Richet, N.; Guise, J.F.; Gramaje, D.; Armengol, J.; Fontaine, F.; Trotel-Aziz, P. Cultivar contributes to the beneficial effects of Bacillus subtilis PTA-271 and Trichoderma atroviride SC1 to protect grapevine against Neofusicoccum parvum. Front. Microbiol. 2021, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Surico, G.; Marchi, G.; Mugnai, L. Older and more recent observations on esca: A critical overview. Phytopathol. Mediterr. 2006, 1000–1019. [Google Scholar] [CrossRef]
- Sosnowski, M.R.; Creaser, M.L.; Wicks, T.J.; Lardner, R.; Scott, E.S. Protection of grapevine pruning wounds from infection by Eutypa lata. Aust. J. Grape Wine Res. 2008, 14, 134–142. [Google Scholar] [CrossRef]
- Martín, L.; Martín, M.T. Characterization of fungicide resistant isolates of Phaeoacremonium aleophilum infecting grapevines in Spain. Crop Prot. 2013, 52, 141–150. [Google Scholar] [CrossRef]
- Fourie, P.H.; Halleen, F. Chemical and biological protection of grapevine propagation material from trunk disease pathogens. Eur. J. Plant Pathol. 2006, 116, 255–265. [Google Scholar] [CrossRef]
- Gramaje, D.; Aroca, Á.; Raposo, R.; García-Jiménez, J.; Armengol, J. Evaluation of fungicides to control Petri disease pathogens in the grapevine propagation process. Crop Prot. 2009, 28, 1091–1097. [Google Scholar] [CrossRef]
- Amponsah, N.T.; Jones, E.; Ridgway, H.J.; Jaspers, M.V. Evaluation of fungicides for the management of Botryosphaeria dieback diseases of grapevines. Pest Manag. Sci. 2012, 68, 676–683. [Google Scholar] [CrossRef]
- Longa, C.M.O.; Savazzini, F.; Tosi, S.; Elad, Y.; Pertot, I. Evaluating the survival and environmental fate of the biocontrol agent Trichoderma atroviride SC1 in vineyards in northern Italy. J. Appl. Microbiol. 2009, 106, 1549–1557. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Bissett, J.; Druzhinina, I.; Kullnig-Gradinger, C.; Szakacs, G. Genetic and metabolic diversity of Trichoderma: A case study on South-East Asian isolates. Fungal Genet. Biol. 2003, 38, 310–319. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Gaigole, A.H.; Wagh, G.N.; Khadse, A.C. Antifungal activity of Trichoderma species against soil borne pathogen. Asiat. J. Biotechnol. Resour. 2011, 2, 461–465. [Google Scholar]
- Sandhu, S.S.; Sharma, A.K.; Beniwal, V.; Goel, G.; Batra, P.; Kumar, A.; Jaglan, S.; Sharma, A.K.; Malhotra, S. Myco-biocontrol of insect pests: Factors involved, mechanism, and regulation. J. Pathog. 2012, 2012. [Google Scholar] [CrossRef]
- Singh, A.; Srivastava, S.; Singh, H.B. Effect of substrates on growth and shelf life of Trichoderma harzianum and its use in biocontrol of diseases. Bioresour. Technol. 2007, 98, 470–473. [Google Scholar] [CrossRef]
- Locatelli, G.O.; Dos Santos, G.F.; Botelho, P.S.; Finkler, C.L.L.; Bueno, L.A. Development of Trichoderma sp. formulations in encapsulated granules (CG) and evaluation of conidia shelf-life. Biol. Control 2018, 117, 21–29. [Google Scholar] [CrossRef]
- Peil, S.; Beckers, S.J.; Fischer, J.; Wurm, F.R. Biodegradable, lignin-based encapsulation enables delivery of Trichoderma reesei with programmed enzymatic release against grapevine trunk diseases. Mater. Today Bio 2020, 7, 100061. [Google Scholar] [CrossRef] [PubMed]
- Ammad, F.; Moumen, O.; Gasem, A.; Othmane, S.; Hisashi, K.N.; Zebib, B.; Merah, O. The potency of lemon (Citrus limon L.) essential oil to control some fungal diseases of grapevine wood. Comptes Rendus Biol. 2018, 341, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Tripathi, A. Effects of Citrus sinensis (L.) Osbeck epicarp essential oil on growth and morphogenesis of Aspergillus niger (L.) Van Tieghem. Microbiol. Res. 2008, 163, 337–344. [Google Scholar] [CrossRef]
- Phillips, C.A.; Laird, K.; Allen, S.C. The use of Citri-VTM®—An antimicrobial citrus essential oil vapour for the control of Penicillium chrysogenum, Aspergillus niger and Alternaria alternata in vitro and on food. Food Res. Int. 2012, 47, 310–314. [Google Scholar] [CrossRef]
- Van Hung, P.; Chi, P.T.L.; Phi, N.T.L. Comparison of antifungal activities of Vietnamese citrus essential oils. Nat. Prod. Res. 2013, 27, 506–508. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.K.; Dubey, N. Evaluation of some essential oils for their toxicity against fungi causing deterioration of stored food commodities. Appl. Environ. Microbiol. 1994, 60, 1101–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, N.; Batra, S.; Mishra, D. Fungitoxic properties of the essential oil of Citrus limon (L.) Burm. Against a few Dermatophytes: Fungitoxische eigenschaften des ätherischen Öls von Citrus limon (L.) Burm. gegenüber einigen Dermatophyten. Mycoses 1988, 31, 380–382. [Google Scholar] [CrossRef]
- Sawamura, M. Citrus Essential Oils: Flavor and Fragrance; John Wiley & Sons: Hoboken, NJ, USA, 2011; ISBN 1118074386. [Google Scholar]
- Gramaje, D.; Armengol, J. Fungal trunk pathogens in the grapevine propagation process: Potential inoculum sources, detection, identification, and management strategies. Plant Dis. 2011, 95, 1040–1055. [Google Scholar] [CrossRef] [Green Version]
- Bruez, E.; Larignon, P.; Compant, S.; Rey, P. Investigating the durable effect of the hot water treatment used in nurseries on pathogenic fungi inhabiting grapevine wood and involved in Grapevine Trunk Diseases. Crop Prot. 2017, 100, 203–210. [Google Scholar] [CrossRef]
- Gramaje, D.; Agustí-Brisach, C.; Pérez-Sierra, A.; Moralejo, E.; Olmo, D.; Mostert, L.; Damm, U.; Armengol, J. Fungal trunk pathogens associated with wood decay of almond trees on Mallorca (Spain). Pers. Mol. Phylogeny Evol. Fungi 2012, 28, 1–13. [Google Scholar] [CrossRef]
- Pierron, R.J.G.; Pages, M.; Couderc, C.; Compant, S.; Jacques, A.; Violleau, F. In vitro and in planta fungicide properties of ozonated water against the esca-associated fungus Phaeoacremonium aleophilum. Sci. Hortic. 2015, 189, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Smilanick, J.L.; Margosan, D.M.; Mlikota Gabler, F. Impact of ozonated water on the quality and shelf-life of fresh citrus fruit, stone fruit, and table grapes. Ozone Sci. Eng. 2002, 24, 343–356. [Google Scholar] [CrossRef]
- Fujiwara, K.; Fujii, T.; Park, J.-S. Comparison of foliar spray efficacy of electrolytically ozonated water and acidic electrolyzed oxidizing water for controlling powdery mildew infection on cucumber leaves. Ozone Sci. Eng. 2009, 31, 10–14. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungus | Primer | Sequence | Length | Reference |
|---|---|---|---|---|
| Eutypa lata | Eut02 Eut02 | 5′TGGTGGACGGGTAGGGTTAG3′ 5′GGCCTTACCGAAATAGACCAA3′ | 643 bp | [162,168] |
| Phaeoacremonium minimum | PAL1 PAL2 | 5′-AGGTCGGGGGCCAAC-3′ 5′-AGGTGTAAACTACTGCGC-3′ | 415 bp | [169] |
| Phaeomoniella chlamydospora | Pch1 Pch2 | 5′-CTCCAACCCTTTGTTTATC-3′ 5′-TGAAAGTTGATATGGACCC-3′ | 360 bp | [169] |
| Fomitiporia mediterranea | Fmed1 Fmed2 | 5′-GCAGTAGTAATAATAACAATC-3′ 5′-GGTCAAAGGAGTCAAATGGT-3′ | 550 bp | [170] |
| Botryosphaeriaceae spp. | Bot-BtF1 Bot-BtR1 | 5′-GTATGGCAATCTTCTGAACG-3′ 5′-CAGTTGTTACCGGCRCCAGA-3′ | 410 bp | [171] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kenfaoui, J.; Radouane, N.; Mennani, M.; Tahiri, A.; El Ghadraoui, L.; Belabess, Z.; Fontaine, F.; El Hamss, H.; Amiri, S.; Lahlali, R.; et al. A Panoramic View on Grapevine Trunk Diseases Threats: Case of Eutypa Dieback, Botryosphaeria Dieback, and Esca Disease. J. Fungi 2022, 8, 595. https://doi.org/10.3390/jof8060595
Kenfaoui J, Radouane N, Mennani M, Tahiri A, El Ghadraoui L, Belabess Z, Fontaine F, El Hamss H, Amiri S, Lahlali R, et al. A Panoramic View on Grapevine Trunk Diseases Threats: Case of Eutypa Dieback, Botryosphaeria Dieback, and Esca Disease. Journal of Fungi. 2022; 8(6):595. https://doi.org/10.3390/jof8060595
Chicago/Turabian StyleKenfaoui, Jihane, Nabil Radouane, Mohammed Mennani, Abdessalem Tahiri, Lahsen El Ghadraoui, Zineb Belabess, Florence Fontaine, Hajar El Hamss, Said Amiri, Rachid Lahlali, and et al. 2022. "A Panoramic View on Grapevine Trunk Diseases Threats: Case of Eutypa Dieback, Botryosphaeria Dieback, and Esca Disease" Journal of Fungi 8, no. 6: 595. https://doi.org/10.3390/jof8060595
APA StyleKenfaoui, J., Radouane, N., Mennani, M., Tahiri, A., El Ghadraoui, L., Belabess, Z., Fontaine, F., El Hamss, H., Amiri, S., Lahlali, R., & Barka, E. A. (2022). A Panoramic View on Grapevine Trunk Diseases Threats: Case of Eutypa Dieback, Botryosphaeria Dieback, and Esca Disease. Journal of Fungi, 8(6), 595. https://doi.org/10.3390/jof8060595