
Multiple Stochastic Parameters Influence Genome Dynamics in a Heterozygous Diploid Eukaryotic Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culturing Conditions
2.2. Blue-Green Collection Strain Construction
2.3. Strain Validation
2.3.1. Basic Phenotyping
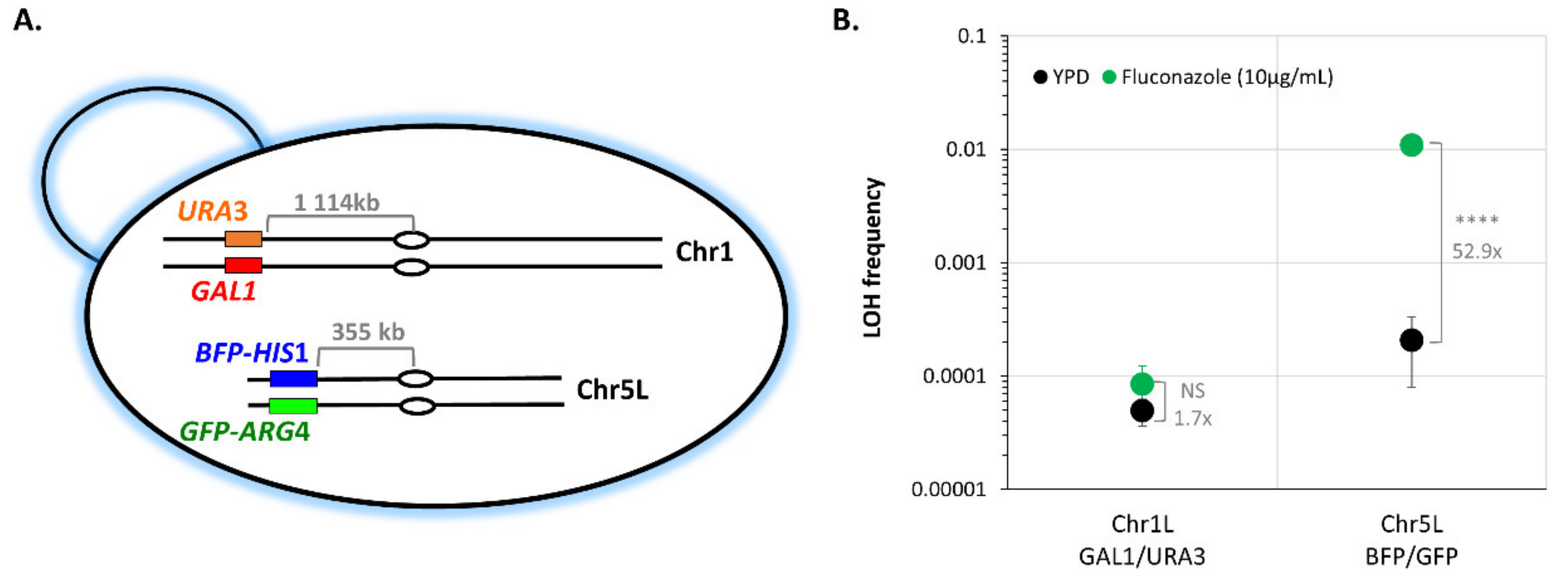
2.3.2. Assessing Functionality of the LOH Reporter System
2.3.3. Identifying the Delimitations of the Mono-Fluorescent Populations on FACS Profiles in Different Genomic Loci
2.4. Fluctuation Assay to Determine LOH Frequency
2.5. Strain Construction for Assessing LOH Frequency Heterogeneity
2.6. Stress Conditions
2.7. Two Reporter System Strain
3. Results
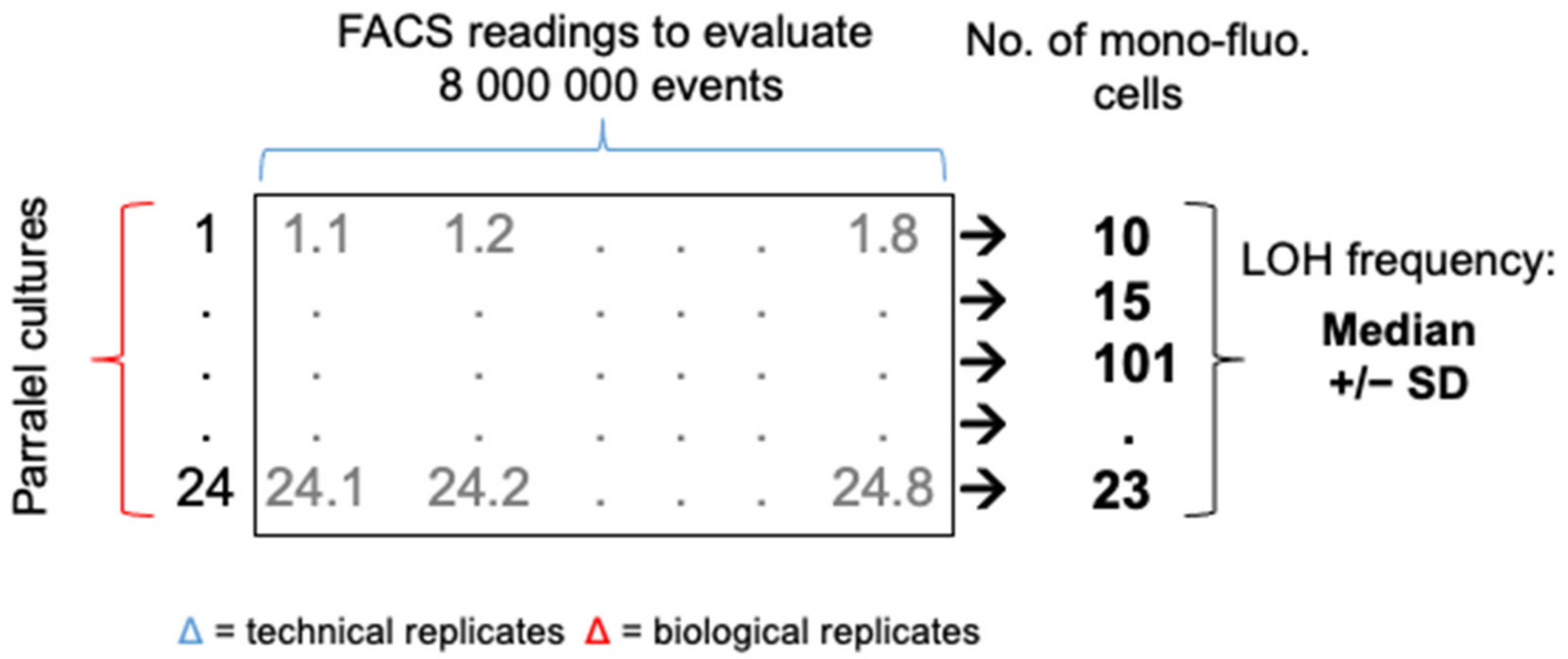
3.1. A Robust Pipeline to Accurately Evaluate Basal LOH
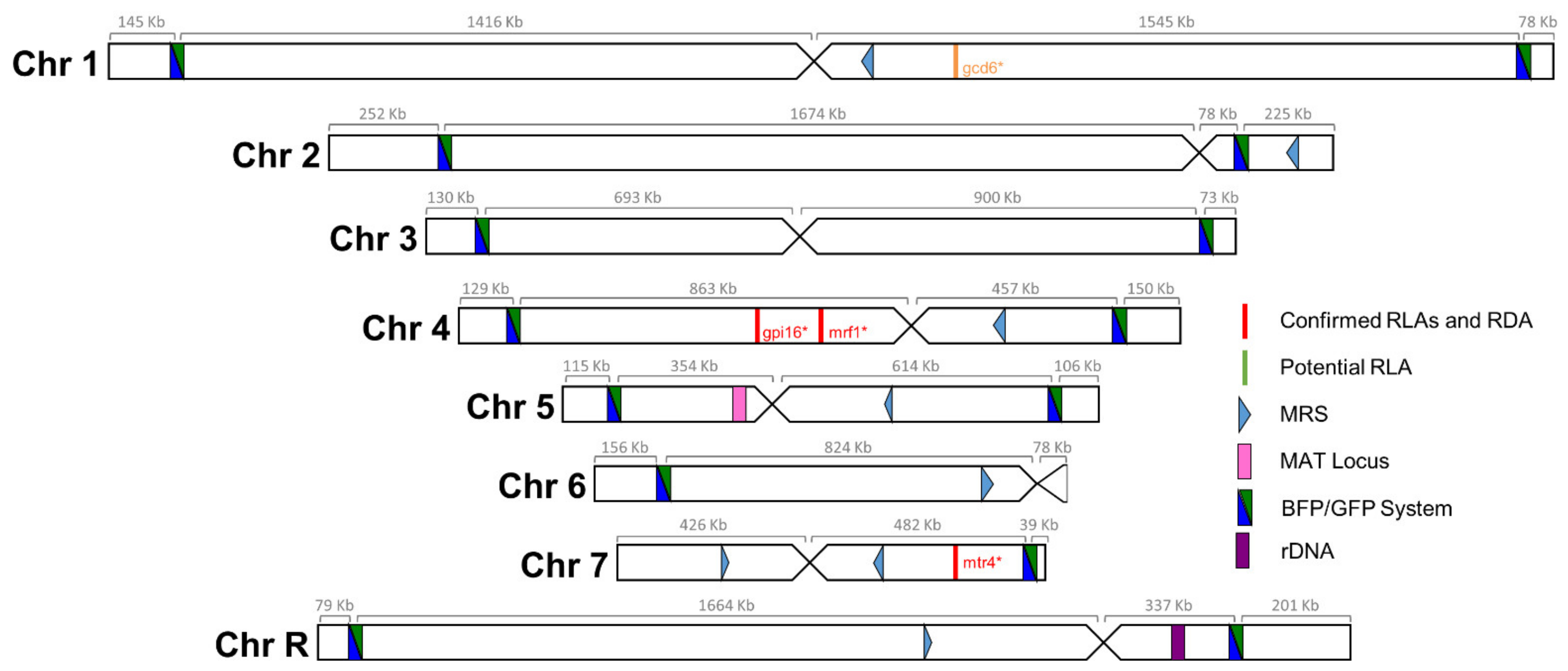
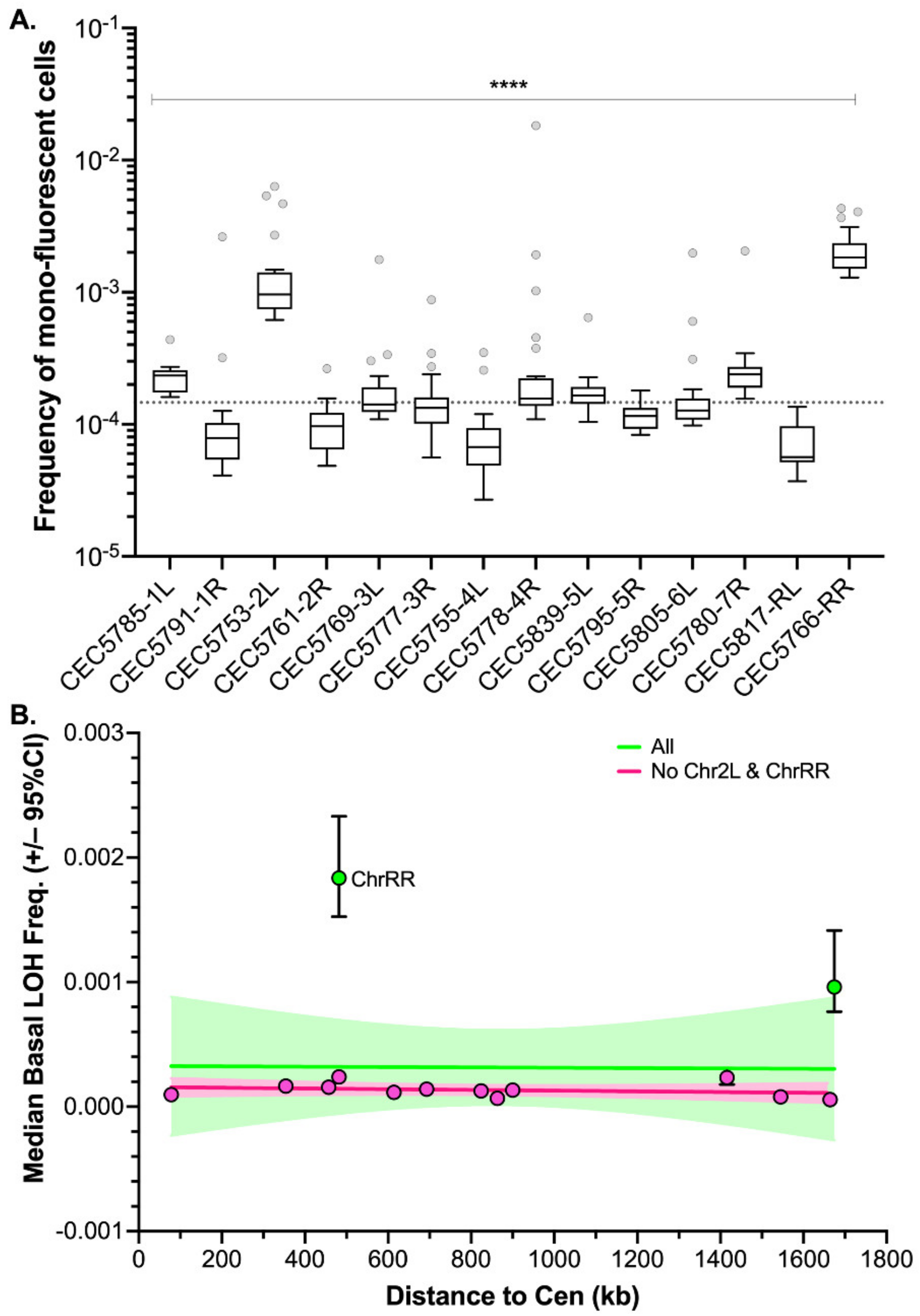
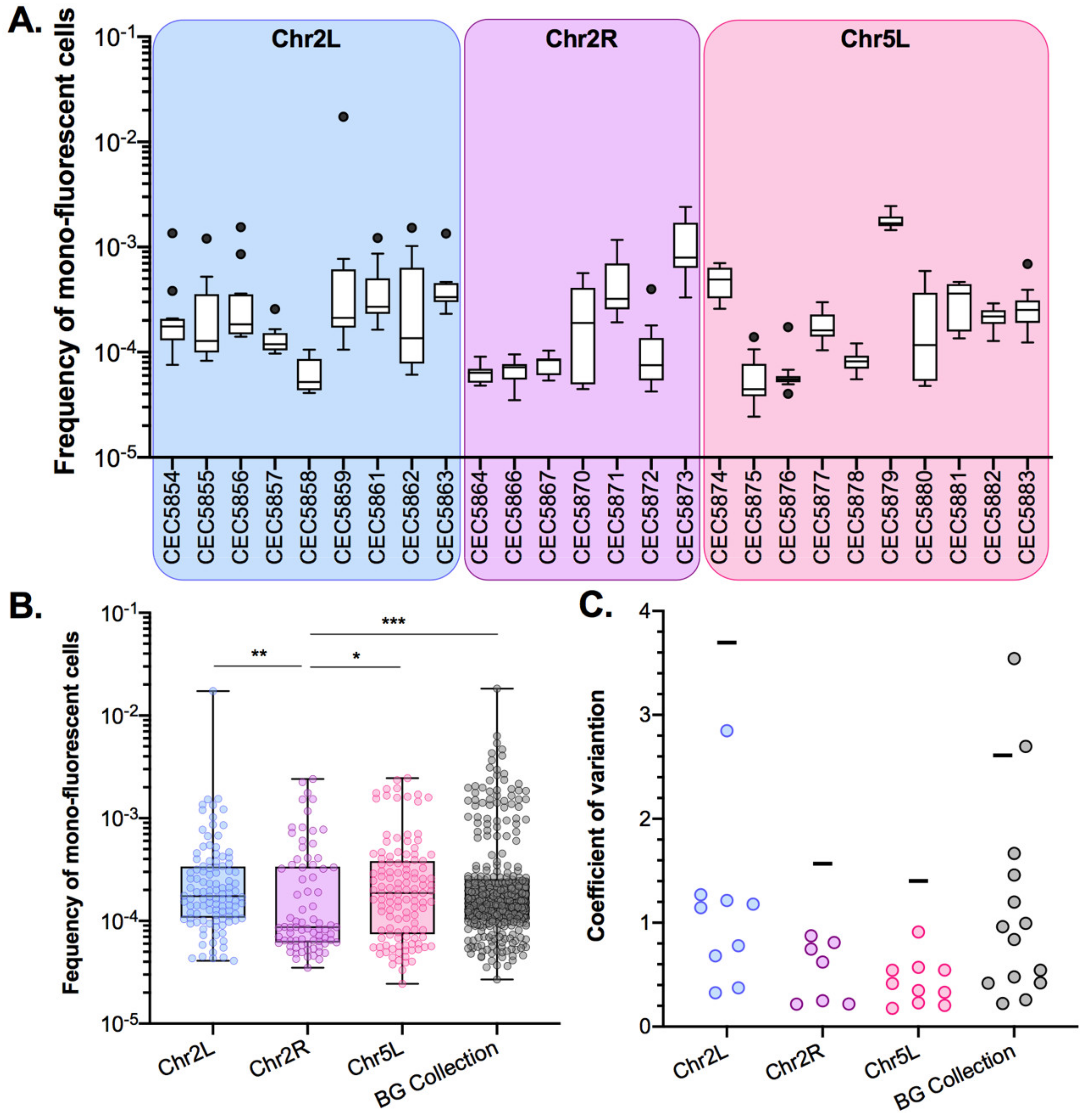
3.2. A Genome-Wide Evaluation of Basal LOH Frequency
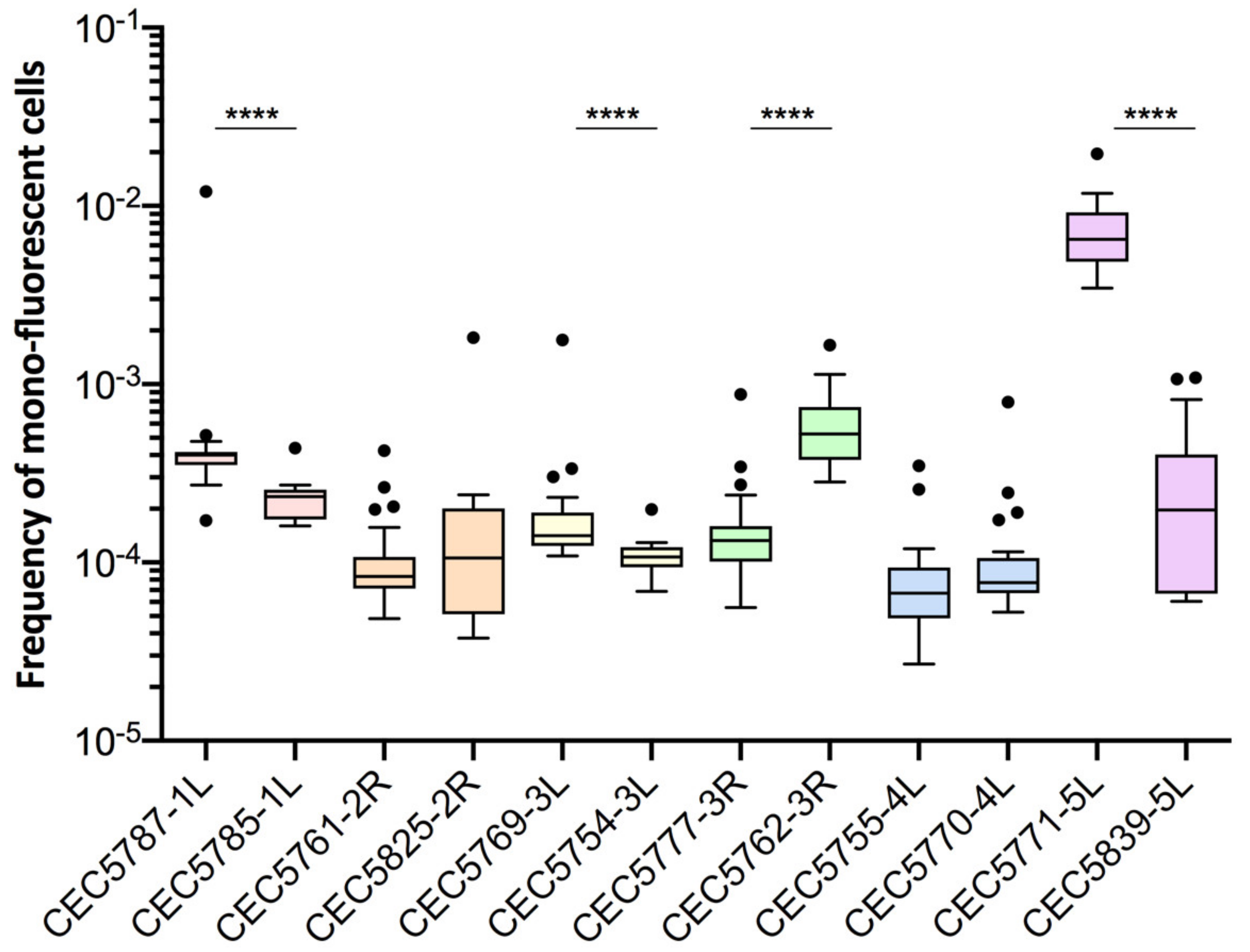
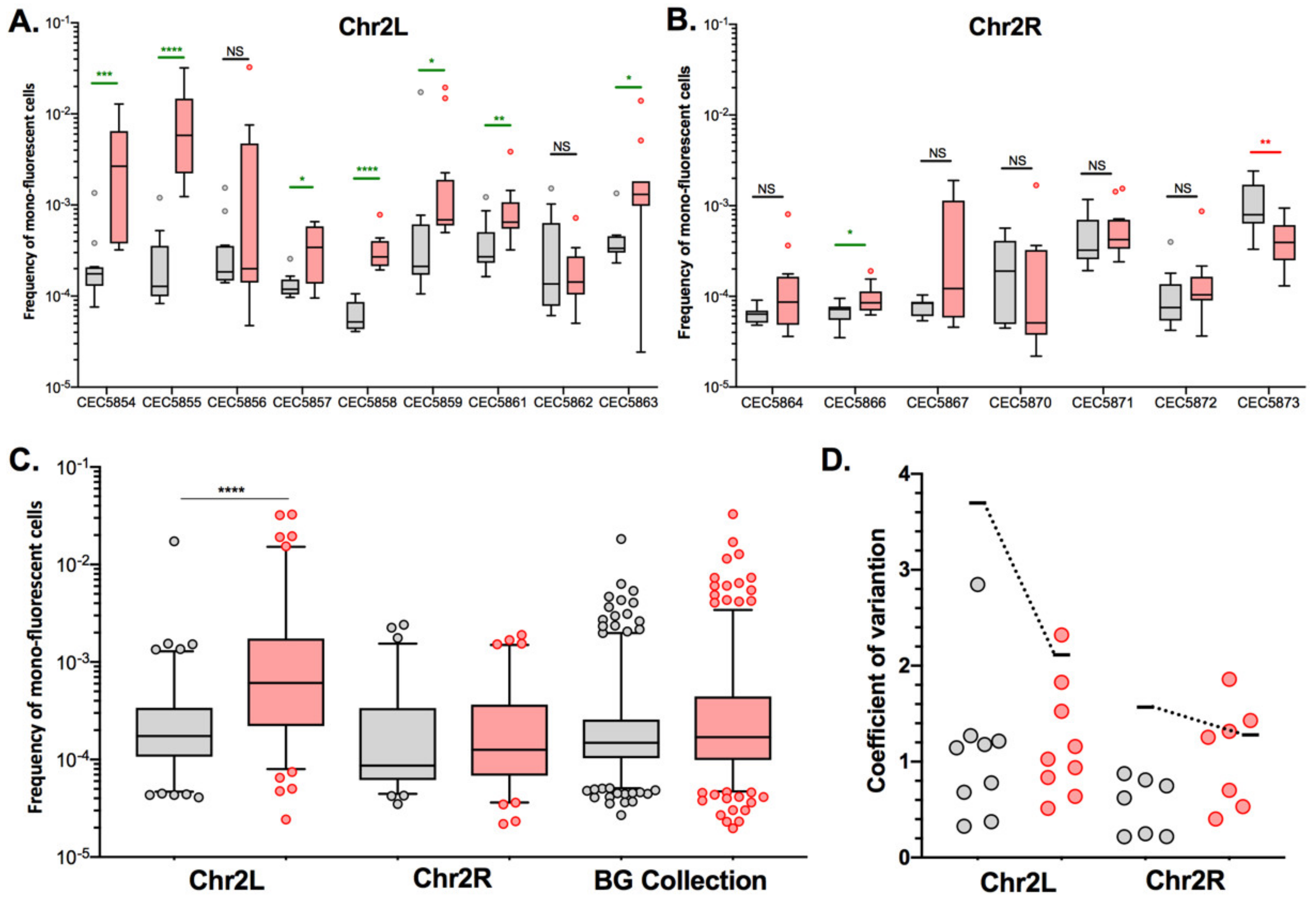
3.3. LOH Frequency Is Influenced by Genetic Background
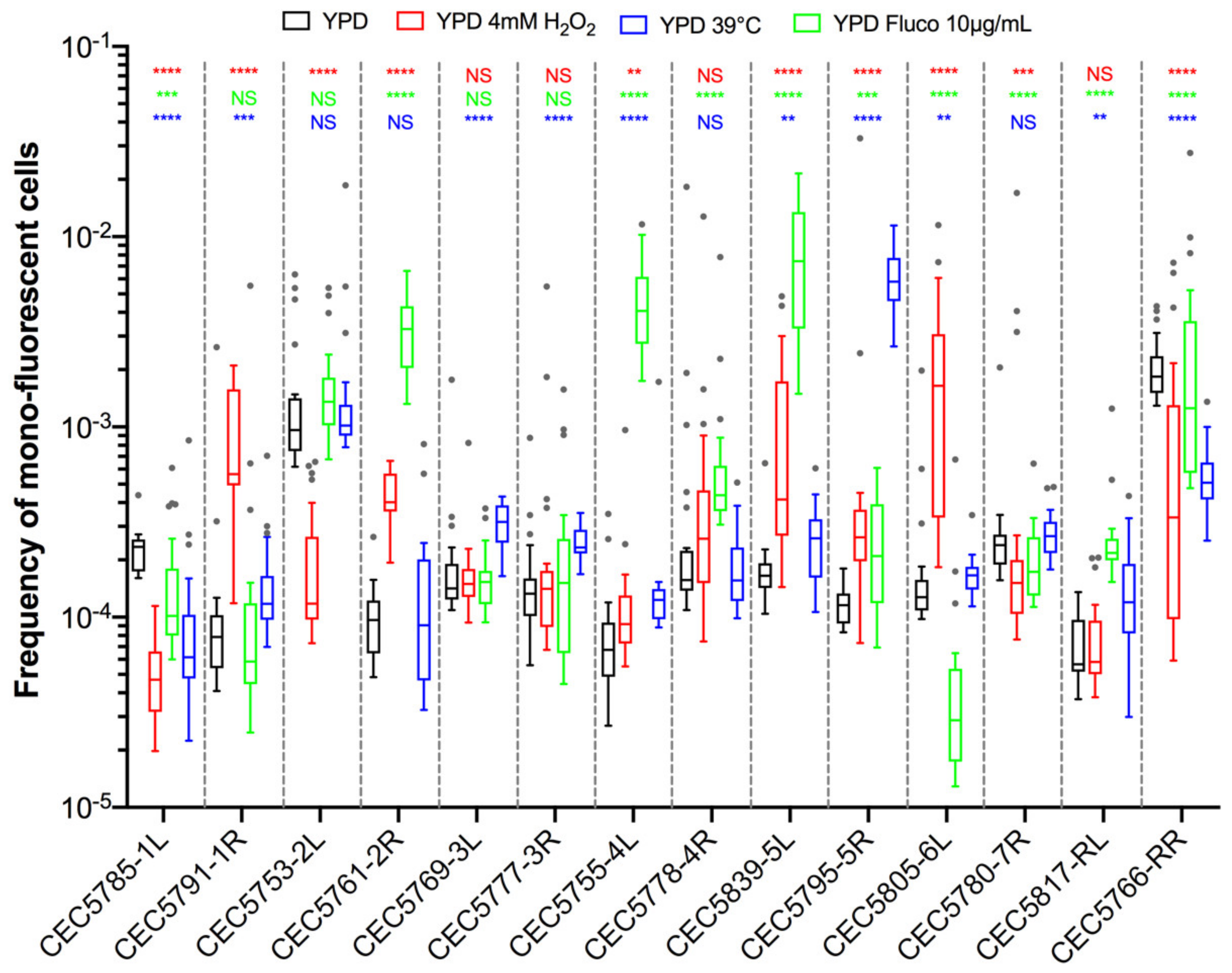
3.4. Stress Induces Different Responses in Terms of LOH Frequency
3.5. Stress May Not Lead to a General Response Triggering Overall Genome Instability
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feri, A.; Loll-Krippleber, R.; Commere, P.-H.; Maufrais, C.; Sertour, N.; Schwartz, K.; Sherlock, G.; Bougnoux, M.-E.; d’Enfert, C.; Legrand, M. Analysis of Repair Mechanisms Following an Induced Double-Strand Break Uncovers Recessive Deleterious Alleles in the Candida albicans Diploid Genome. Mbio 2016, 7, e01109-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legrand, M.; Chan, C.L.; Jauert, P.A.; Kirkpatrick, D.T. Role of DNA Mismatch Repair and Double-Strand Break Repair in Genome Stability and Antifungal Drug Resistance in Candida albicans. Eukaryot Cell 2007, 6, 2194–2205. [Google Scholar] [CrossRef] [Green Version]
- Vyas, V.K.; Bushkin, G.G.; Bernstein, D.A.; Getz, M.A.; Sewastianik, M.; Barrasa, M.I.; Bartel, D.P.; Fink, G.R. New CRISPR Mutagenesis Strategies Reveal Variation in Repair Mechanisms among Fungi. Msphere 2018, 3, e00154-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malkova, A.; Klein, F.; Leung, W.-Y.; Haber, J.E. HO Endonuclease-Induced Recombination in Yeast Meiosis Resembles Spo11-Induced Events. Proc. Natl. Acad. Sci. USA 2000, 97, 14500–14505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forche, A.; Abbey, D.; Pisithkul, T.; Weinzierl, M.A.; Ringstrom, T.; Bruck, D.; Petersen, K.; Berman, J. Stress Alters Rates and Types of Loss of Heterozygosity in Candida albicans. Mbio 2011, 2, e00129-11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciudad, T.; Andaluz, E.; Steinberg-Neifach, O.; Lue, N.F.; Gow, N.A.R.; Calderone, R.A.; Larriba, G. Homologous Recombination in Candida albicans: Role of CaRad52p in DNA Repair, Integration of Linear DNA Fragments and Telomere Length. Mol. Microbiol. 2004, 53, 1177–1194. [Google Scholar] [CrossRef]
- Ciudad, T.; Bellido, A.; Hermosa, B.; Andaluz, E.; Larriba, G. DLH1, the Candida albicans Homologue of the Meiosis-specific DMC1, Is Not Involved in DNA Repair but Catalyses Spontaneous Interhomologue Recombination and Might Promote Non-crossover Events. Cell Microbiol. 2020, 22, e13137. [Google Scholar] [CrossRef]
- Loll-Krippleber, R.; Feri, A.; Nguyen, M.; Maufrais, C.; Yansouni, J.; d’Enfert, C.; Legrand, M. A FACS-Optimized Screen Identifies Regulators of Genome Stability in Candida albicans. Eukaryot Cell 2015, 14, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Andersen, M.P.; Nelson, Z.W.; Hetrick, E.D.; Gottschling, D.E. A Genetic Screen for Increased Loss of Heterozygosity in Saccharomyces Cerevisiae. Genetics 2008, 179, 1179–1195. [Google Scholar] [CrossRef] [Green Version]
- Tutaj, H.; Pogoda, E.; Tomala, K.; Korona, R. Gene Overexpression Screen for Chromosome Instability in Yeast Primarily Identifies Cell Cycle Progression Genes. Curr. Genet. 2019, 65, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Forche, A.; Magee, P.T.; Selmecki, A.; Berman, J.; May, G. Evolution in Candida albicans Populations During a Single Passage Through a Mouse Host. Genetics 2009, 182, 799–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ene, I.V.; Farrer, R.A.; Hirakawa, M.P.; Agwamba, K.; Cuomo, C.A.; Bennett, R.J. Global Analysis of Mutations Driving Microevolution of a Heterozygous Diploid Fungal Pathogen. Proc. Natl. Acad. Sci. USA 2018, 115, E8688–E8697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forche, A.; Cromie, G.; Gerstein, A.C.; Solis, N.V.; Pisithkul, T.; Srifa, W.; Jeffery, E.; Abbey, D.; Filler, S.G.; Dudley, A.M.; et al. Rapid Phenotypic and Genotypic Diversification After Exposure to the Oral Host Niche in Candida albicans. Genetics 2018, 209, 725–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coste, A.; Selmecki, A.; Forche, A.; Diogo, D.; Bougnoux, M.-E.; d’Enfert, C.; Berman, J.; Sanglard, D. Genotypic Evolution of Azole Resistance Mechanisms in Sequential Candida albicans Isolates. Eukaryot Cell 2007, 6, 1889–1904. [Google Scholar] [CrossRef] [Green Version]
- Tso, G.H.W.; Reales-Calderon, J.A.; Tan, A.S.M.; Sem, X.; Le, G.T.T.; Tan, T.G.; Lai, G.C.; Srinivasan, K.G.; Yurieva, M.; Liao, W.; et al. Experimental Evolution of a Fungal Pathogen into a Gut Symbiont. Science 2018, 362, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, S.-H.; Anderson, M.Z.; Hirakawa, M.P.; Wang, J.M.; Frazer, C.; Alaalm, L.M.; Thomson, G.J.; Ene, I.V.; Bennett, R.J. Hemizygosity Enables a Mutational Transition Governing Fungal Virulence and Commensalism. Cell Host Microbe 2019, 25, 418–431.e6. [Google Scholar] [CrossRef] [Green Version]
- Gerstein, A.C.; Kuzmin, A.; Otto, S.P. Loss-of-Heterozygosity Facilitates Passage through Haldane’s Sieve for Saccharomyces Cerevisiae Undergoing Adaptation. Nat. Commun. 2014, 5, 3819. [Google Scholar] [CrossRef] [Green Version]
- James, T.Y.; Michelotti, L.A.; Glasco, A.D.; Clemons, R.A.; Powers, R.A.; James, E.S.; Simmons, D.R.; Bai, F.; Ge, S. Adaptation by Loss of Heterozygosity in Saccharomyces Cerevisiae Clones Under Divergent Selection. Genetics 2019, 213, 665–683. [Google Scholar] [CrossRef]
- Marton, T.; Feri, A.; Commere, P.-H.; Maufrais, C.; d’Enfert, C.; Legrand, M. Identification of Recessive Lethal Alleles in the Diploid Genome of a Candida albicans Laboratory Strain Unveils a Potential Role of Repetitive Sequences in Buffering Their Deleterious Impact. Msphere 2019, 4, e00709-18. [Google Scholar] [CrossRef] [Green Version]
- Ciudad, T.; Hickman, M.; Bellido, A.; Berman, J.; Larriba, G. Phenotypic Consequences of a Spontaneous Loss of Heterozygosity in a Common Laboratory Strain of Candida albicans. Genetics 2016, 203, 1161–1176. [Google Scholar] [CrossRef] [Green Version]
- Dunkel, N.; Morschhäuser, J. Loss of Heterozygosity at an Unlinked Genomic Locus Is Responsible for the Phenotype of a Candida albicans Sap4Δ Sap5Δ Sap6Δ Mutant. Eukaryot Cell 2011, 10, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boeke, J.D.; Croute, F.L.; Fink, G.R. A Positive Selection for Mutants Lacking Orotidine-5′-Phosphate Decarboxylase Activity in Yeast: 5-Fluoro-Orotic Acid Resistance. Mol. Genet. Genom. 1984, 197, 345–346. [Google Scholar] [CrossRef] [PubMed]
- Gorman, J.A.; Gorman, J.W.; Koltin, Y. Direct Selection of Galactokinase-Negative Mutants of Candida albicans Using 2-Deoxy-Galactose. Curr. Genet. 1992, 21, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.M.; Bennett, R.J.; Anderson, M.Z. The Genome of the Human Pathogen Candida albicans Is Shaped by Mutation and Cryptic Sexual Recombination. Mbio 2018, 9, e01205-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, R.T.; Wikoff, T.D.; Forche, A.; Selmecki, A. Genome Plasticity in Candida albicans Is Driven by Long Repeat Sequences. Elife 2019, 8, e45954. [Google Scholar] [CrossRef]
- Hirakawa, M.P.; Martinez, D.A.; Sakthikumar, S.; Anderson, M.Z.; Berlin, A.; Gujja, S.; Zeng, Q.; Zisson, E.; Wang, J.M.; Greenberg, J.M.; et al. Genetic and Phenotypic Intra-Species Variation in Candida albicans. Genome Res. 2015, 25, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Ropars, J.; Maufrais, C.; Diogo, D.; Marcet-Houben, M.; Perin, A.; Sertour, N.; Mosca, K.; Permal, E.; Laval, G.; Bouchier, C.; et al. Gene Flow Contributes to Diversification of the Major Fungal Pathogen Candida albicans. Nat. Commun. 2018, 9, 2253. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yan, Z.; Xu, J. Quantitative Variation of Biofilms among Strains in Natural Populations of Candida albicans. Microbiology + 2003, 149, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.Y.; Woolford, C.A.; May, G.; McManus, C.J.; Mitchell, A.P. Circuit Diversification in a Biofilm Regulatory Network. PLoS Pathog. 2019, 15, e1007787. [Google Scholar] [CrossRef]
- Rosenberg, A.; Ene, I.V.; Bibi, M.; Zakin, S.; Segal, E.S.; Ziv, N.; Dahan, A.M.; Colombo, A.L.; Bennett, R.J.; Berman, J. Antifungal Tolerance Is a Subpopulation Effect Distinct from Resistance and Is Associated with Persistent Candidemia. Nat. Commun 2018, 9, 2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, T.; Schacherer, J. Genetic Backgrounds and Hidden Trait Complexity in Natural Populations. Curr. Opin. Genet. Dev. 2017, 47, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Gerstein, A.C.; Berman, J. Candida albicans Genetic Background Influences Mean and Heterogeneity of Drug Responses and Genome Stability during Evolution in Fluconazole. Msphere 2020, 5, e00480-20. [Google Scholar] [CrossRef]
- Tutaj, H.; Pirog, A.; Tomala, K.; Korona, R. Genome-Scale Patterns in the Loss of Heterozygosity Incidence in Saccharomyces Cerevisiae. Genetics 2022, 221, iyac032. [Google Scholar] [CrossRef] [PubMed]
- Noble, S.M.; Johnson, A.D. Strains and Strategies for Large-Scale Gene Deletion Studies of the Diploid Human Fungal Pathogen Candida albicans. Eukaryot Cell 2005, 4, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Marton, T.; Maufrais, C.; d’Enfert, C.; Legrand, M. Use of CRISPR-Cas9 to Target Homologous Recombination Limits Transformation-Induced Genomic Changes in Candida albicans. Msphere 2020, 5, e00620-20. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Ichikawa, Y.; Woolford, C.A.; Mitchell, A.P. Candida albicans Gene Deletion with a Transient CRISPR-Cas9 System. Msphere 2016, 1, e00130-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walther, A.; Wendland, J. An Improved Transformation Protocol for the Human Fungal Pathogen Candida albicans. Curr Genet. 2003, 42, 339–343. [Google Scholar] [CrossRef]
- Parfrey, L.W.; Lahr, D.J.G.; Katz, L.A. The Dynamic Nature of Eukaryotic Genomes. Mol. Biol. Evol. 2008, 25, 787–794. [Google Scholar] [CrossRef]
- Rustchenko, E.P.; Curran, T.M.; Sherman, F. Variations in the Number of Ribosomal DNA Units in Morphological Mutants and Normal Strains of Candida albicans and in Normal Strains of Saccharomyces Cerevisiae. J. Bacteriol. 1993, 175, 7189–7199. [Google Scholar] [CrossRef] [Green Version]
- Marton, T. Loss of Heterozygosity: Its Impact on Generating and Shaping Genetic Variations in the Human Fungal Pathogen Candida albicans. Ph.D. Thesis, Université de Paris-Ecole Doctorale BioSPC, Paris, France, 2020. [Google Scholar]
- Heil, C.S.S.; DeSevo, C.G.; Pai, D.A.; Tucker, C.M.; Hoang, M.L.; Dunham, M.J. Loss of Heterozygosity Drives Adaptation in Hybrid Yeast. Mol. Biol. Evol. 2017, 34, msx098. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; François, J.-M.; Capp, J.-P. Gene Expression Noise Produces Cell-to-Cell Heterogeneity in Eukaryotic Homologous Recombination Rate. Front. Genet. 2019, 10, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buscaino, A. Chromatin-Mediated Regulation of Genome Plasticity in Human Fungal Pathogens. Genes 2019, 10, 855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, B.S.; De, S. Loss of Heterozygosity Preferentially Occurs in Early Replicating Regions in Cancer Genomes. Nucleic Acids Res. 2013, 41, 7615–7624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koren, A.; Handsaker, R.E.; Kamitaki, N.; Karlić, R.; Ghosh, S.; Polak, P.; Eggan, K.; McCarroll, S.A. Genetic Variation in Human DNA Replication Timing. Cell 2014, 159, 1015–1026. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marton, T.; d’Enfert, C.; Legrand, M. Multiple Stochastic Parameters Influence Genome Dynamics in a Heterozygous Diploid Eukaryotic Model. J. Fungi 2022, 8, 650. https://doi.org/10.3390/jof8070650
Marton T, d’Enfert C, Legrand M. Multiple Stochastic Parameters Influence Genome Dynamics in a Heterozygous Diploid Eukaryotic Model. Journal of Fungi. 2022; 8(7):650. https://doi.org/10.3390/jof8070650
Chicago/Turabian StyleMarton, Timea, Christophe d’Enfert, and Melanie Legrand. 2022. "Multiple Stochastic Parameters Influence Genome Dynamics in a Heterozygous Diploid Eukaryotic Model" Journal of Fungi 8, no. 7: 650. https://doi.org/10.3390/jof8070650
APA StyleMarton, T., d’Enfert, C., & Legrand, M. (2022). Multiple Stochastic Parameters Influence Genome Dynamics in a Heterozygous Diploid Eukaryotic Model. Journal of Fungi, 8(7), 650. https://doi.org/10.3390/jof8070650