
Agaricus macrochlamys, a New Species from the (Sub)tropical Cloud Forests of North America and the Caribbean, and Agaricus fiardii, a New Synonym of Agaricus subrufescens

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Materials and Methods
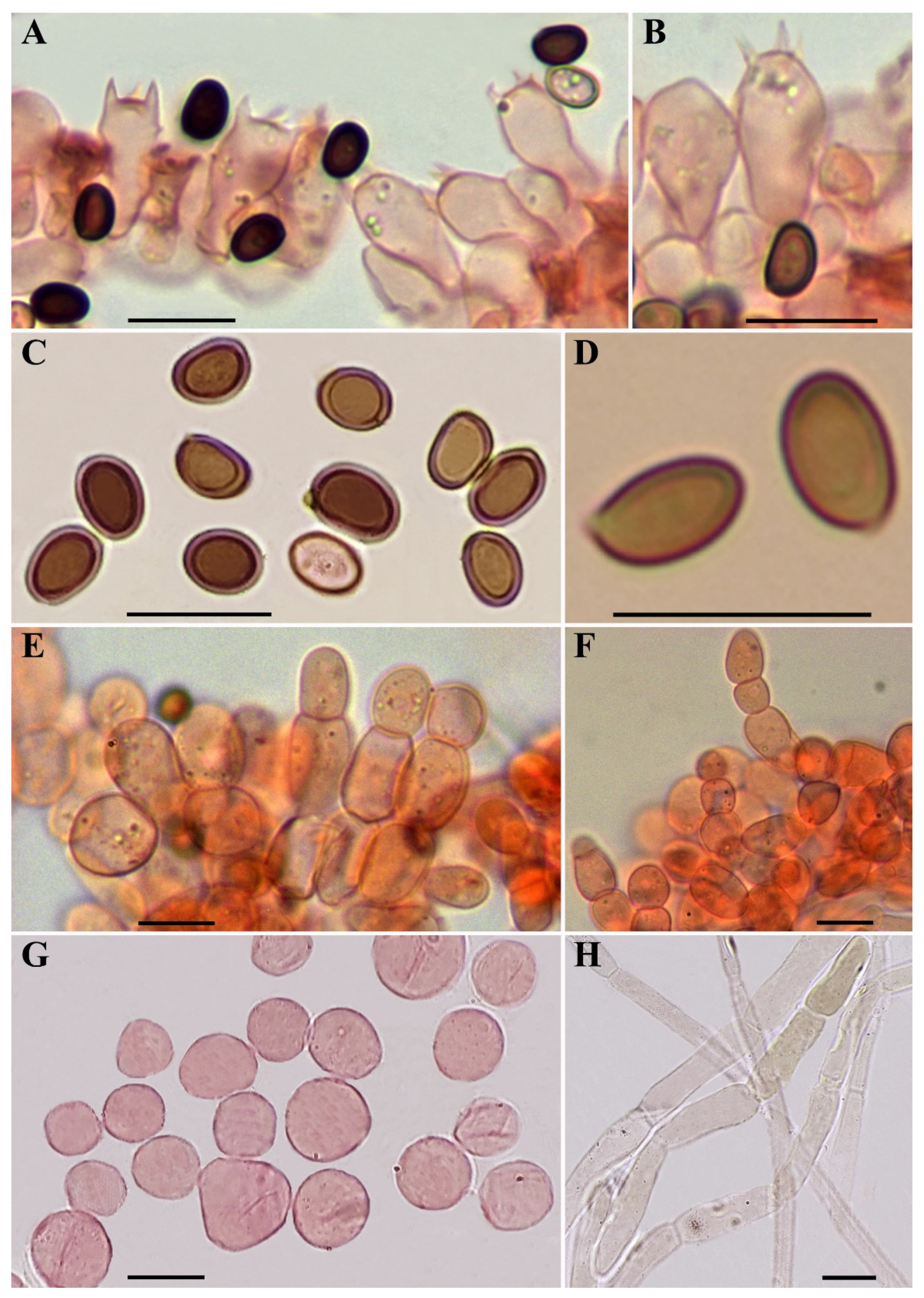
2.1. Sampling and Morphological Description
2.2. DNA Extraction, PCR, and Sequencing
2.3. Sampling for Phylogenetic Analyses
2.4. Phylogenetic Analyses
2.5. Species-Specific ITS Markers
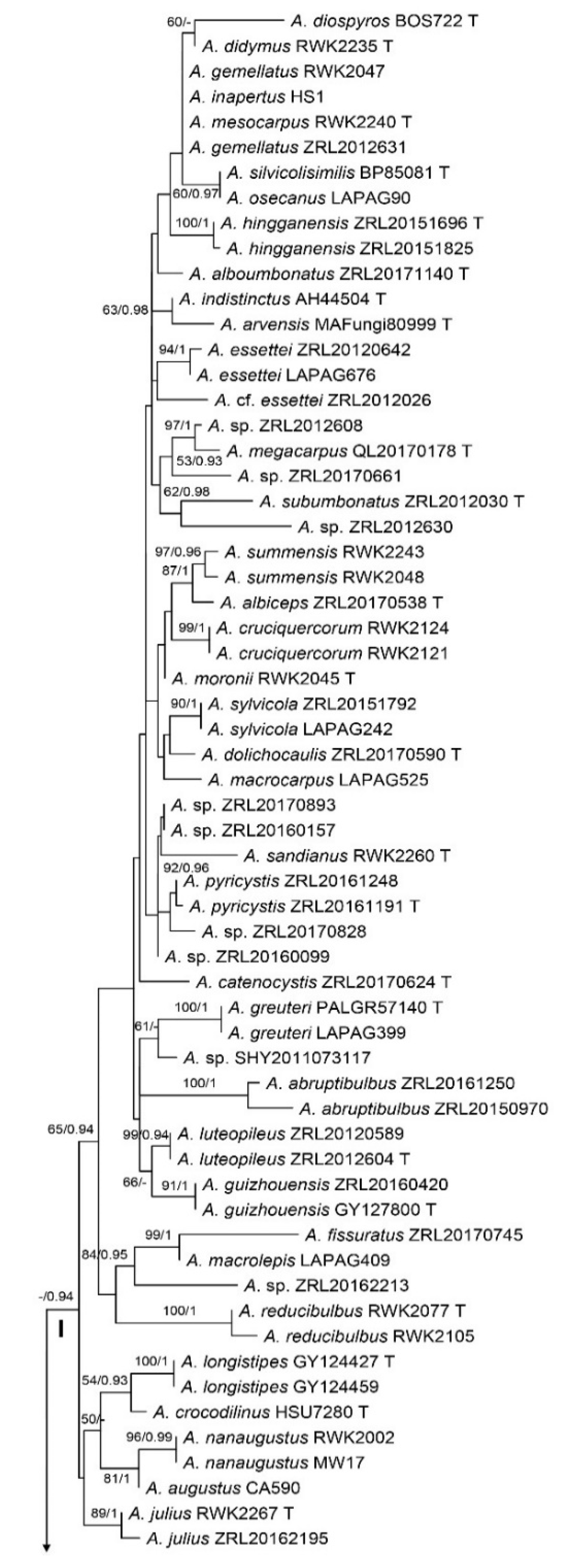
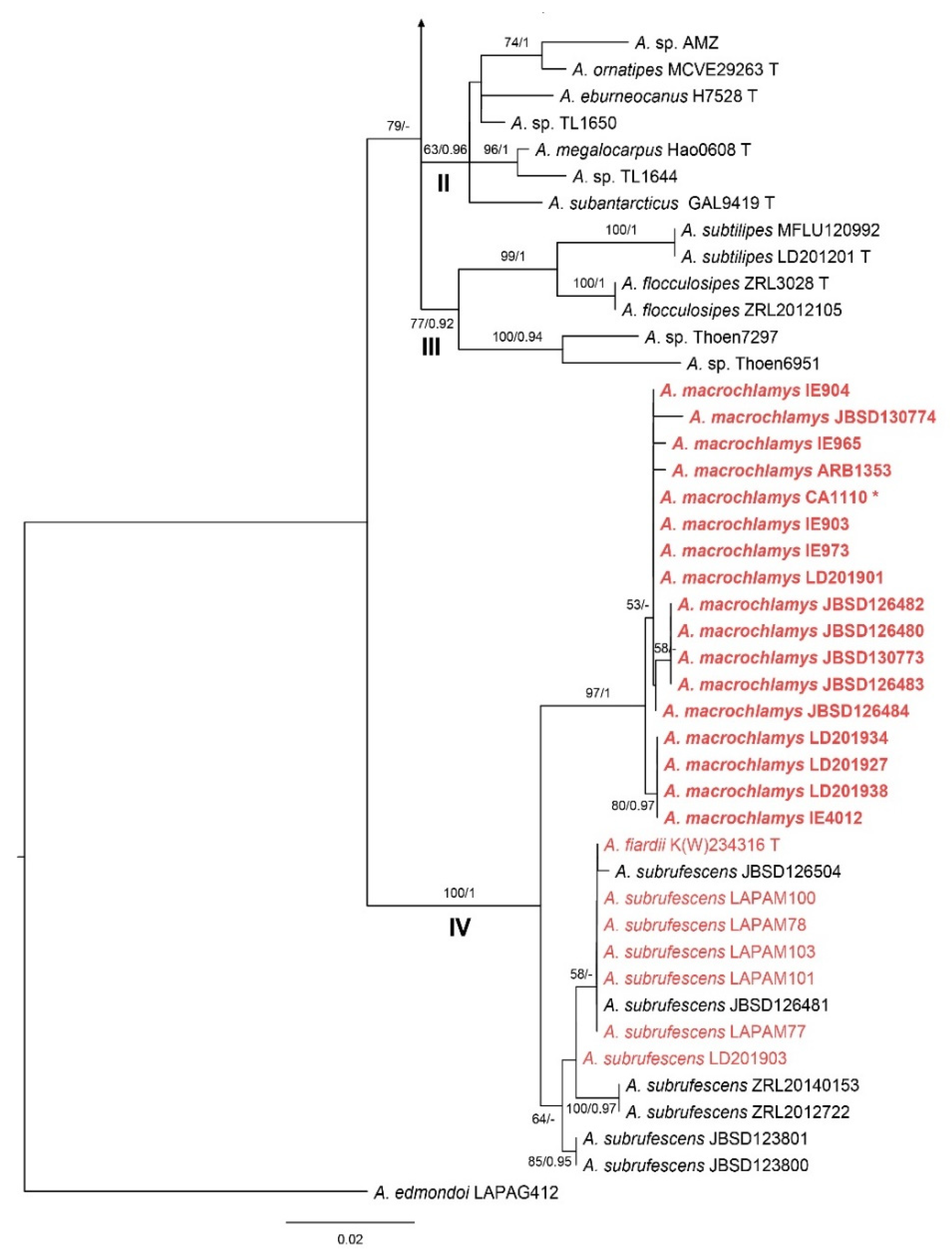
3. Results
3.1. Phylogenetic Analyses
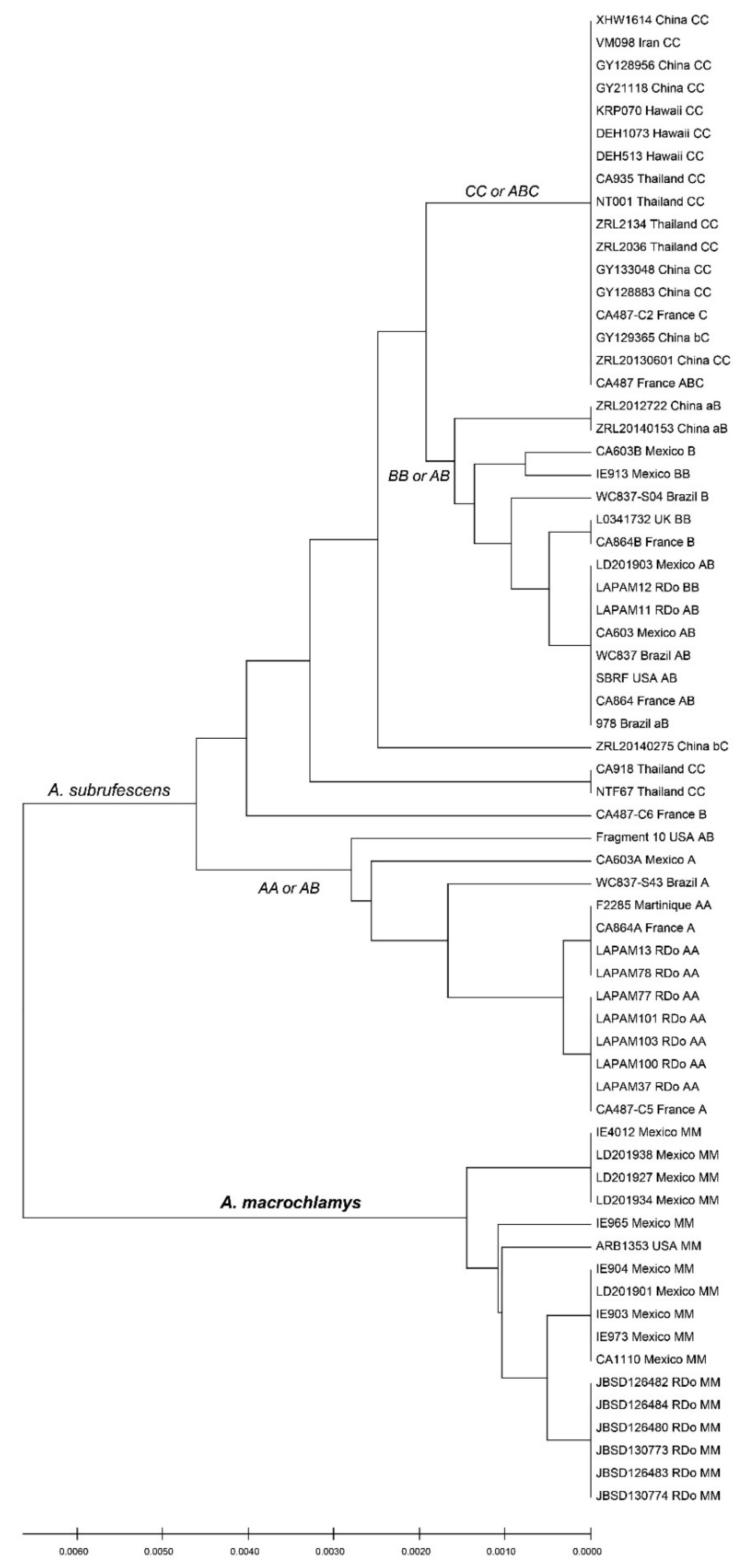
3.2. ITS Types and Characteristic Polymorphisms
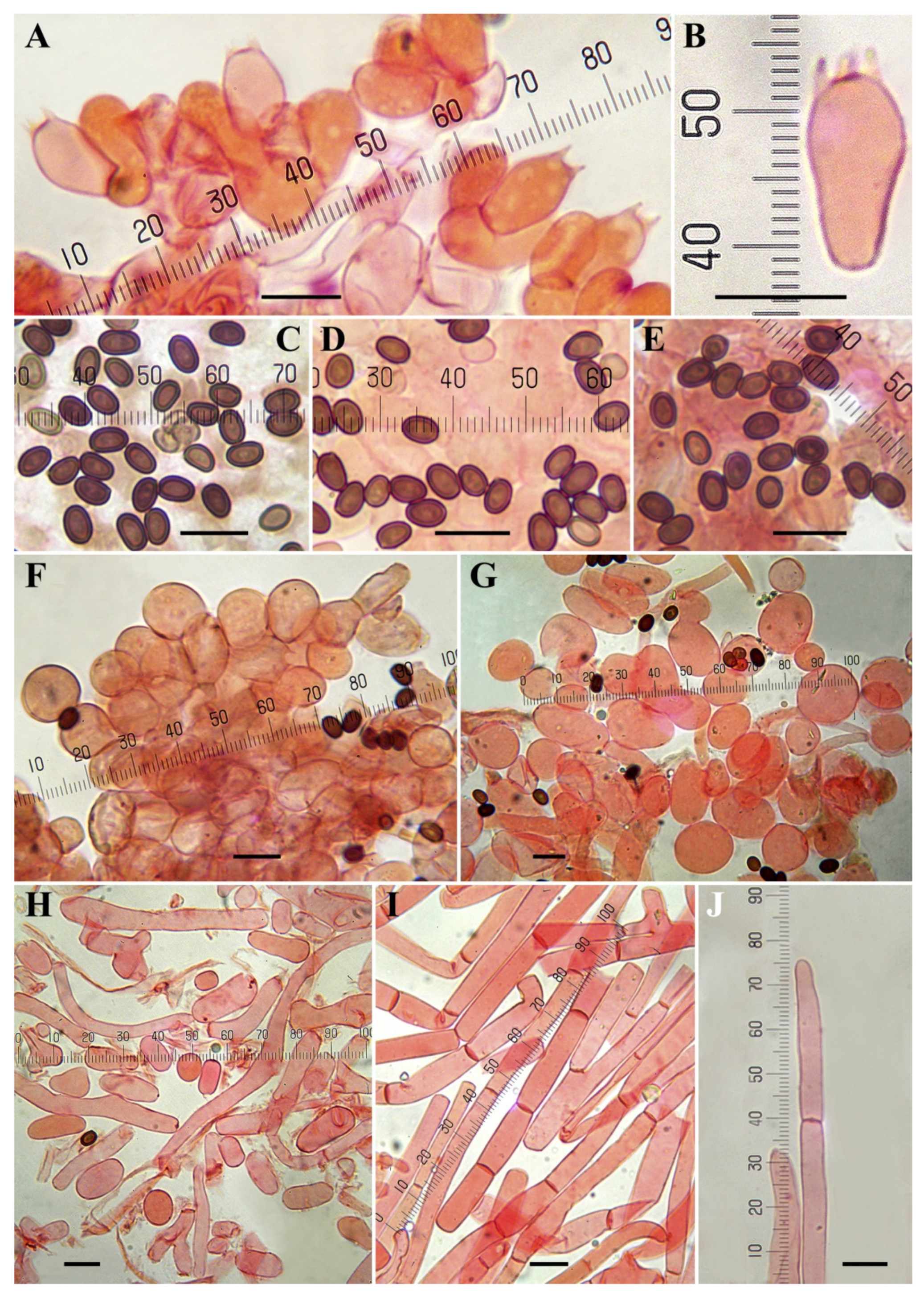
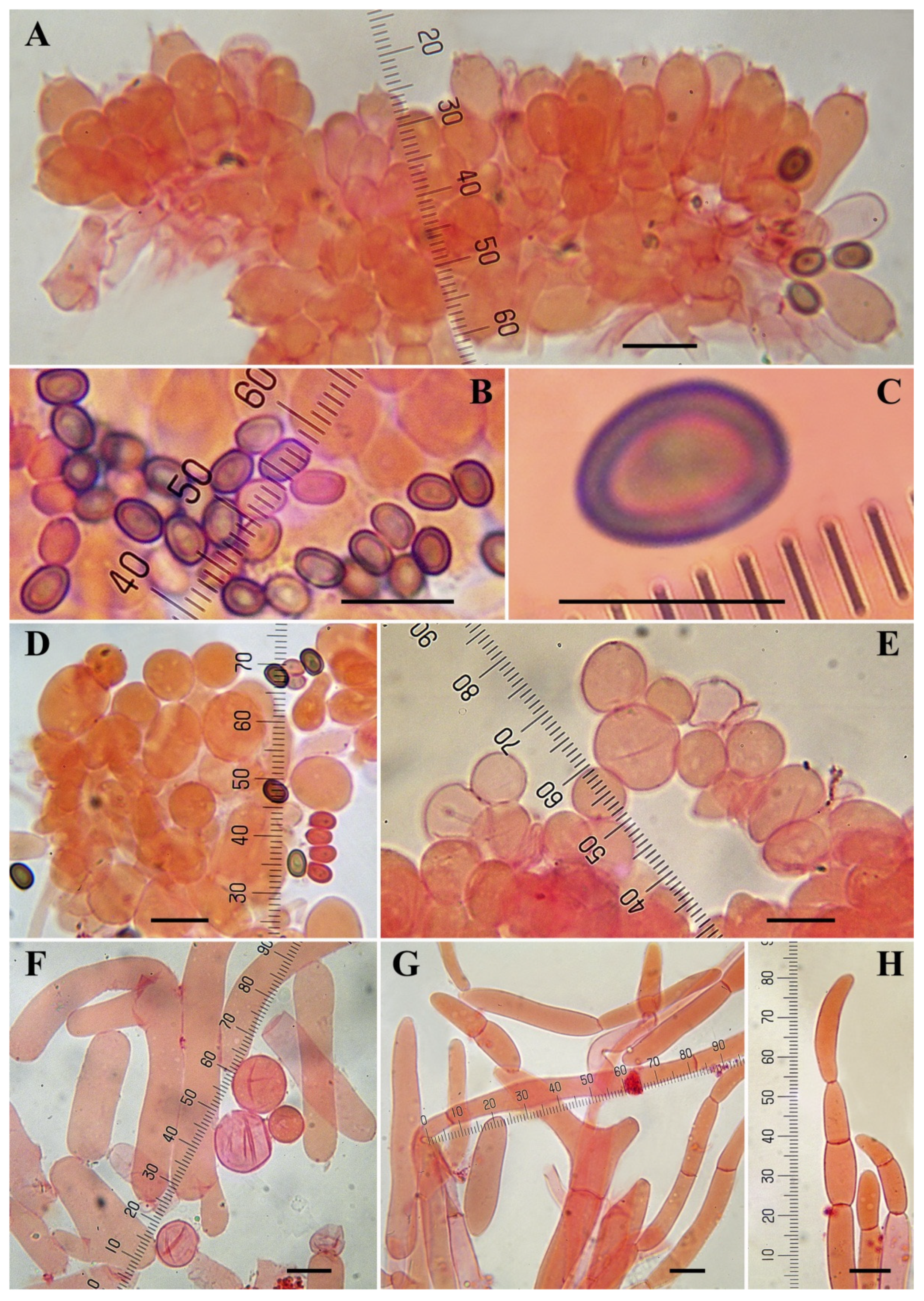
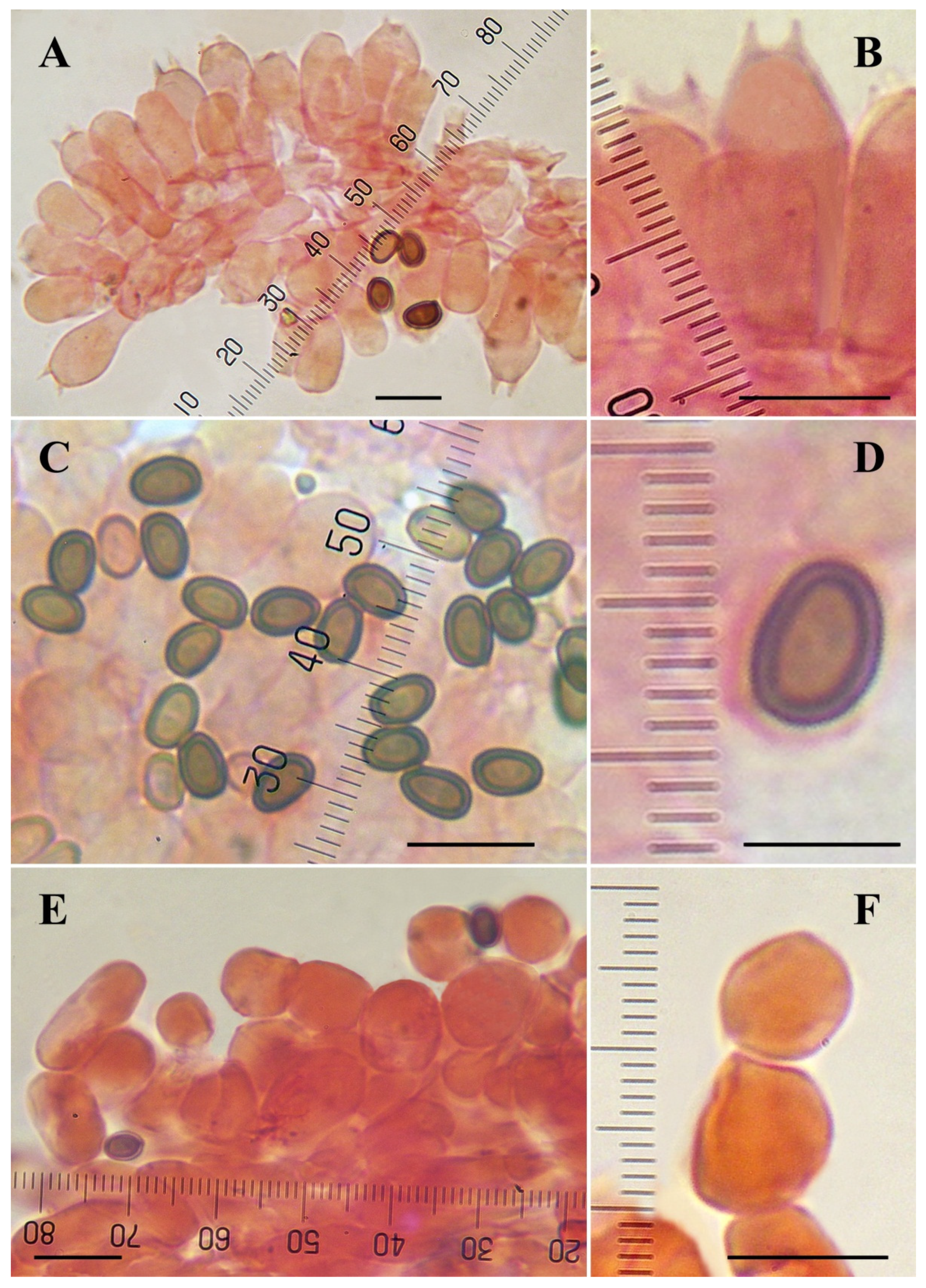
3.3. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karunarathna, S.C.; Chen, J.; Mortimer, P.E.; Xu, J.C.; Zhao, R.L.; Callac, P.; Hyde, K.D. Mycosphere Essay 8: A review of genus Agaricus in tropical and humid subtropical regions of Asia. Mycosphere 2016, 7, 417–439. [Google Scholar] [CrossRef]
- Kerrigan, R.W. Agaricus of North America. Mem. N. Y. Bot. Gard. 2016, 114, 1–573. [Google Scholar]
- Chen, J.; Callac, P.; Parra, L.A.; Karunarathna, S.C.; He, M.Q.; Moinard, M.; De Kesel, A.; Raspé, O.; Wisitrassameewong, K.; Hyde, K.D.; et al. Study in Agaricus subgenus Minores and allied clades reveals a new American subgenus and contrasting phylogenetic patterns in Europe and Greater Mekong Subregion. Persoonia 2017, 38, 170–196. [Google Scholar] [CrossRef] [Green Version]
- Callac, P.; Chen, J. Tropical species of Agaricus. In Updates on Tropical Mushrooms; Sanchez, J.E., Mata, G., Royse, D.J., Eds.; El Colegio de la Frontera Sur: Chiapas, Mexico, 2018; pp. 25–38. [Google Scholar]
- Cao, B.; He, M.Q.; Ling, Z.L.; Zhang, M.Z.; Wei, S.L.; Zhao, R.L. A revision of Agaricus section Arvenses with nine new species from china. Mycologia 2020, 113, 191–211. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Santana, B.; Chen, J.; Parra, L.A.; Angelini, C.; Lodge, D.J.; Kerrigan, R.W.; Callac, P. The genus Agaricus in the Caribbean II. Refined phylogeny of Agaricus subg. Spissicaules with description of two new sections and eight new species. Mycol. Prog. 2021, 20, 381–411. [Google Scholar] [CrossRef]
- Zhao, R.L.; Karunarathna, S.; Raspé, O.; Parra, L.A.; Guinberteau, J.; Moinard, M.; De Kesel, A.; Barroso, G.; Courtecuisse, R.; Hyde, K.D.; et al. Major clades in tropical Agaricus. Fungal Divers. 2011, 51, 279–296. [Google Scholar] [CrossRef]
- Mata, G.; Medel, R.; Salmones, D. A preliminary survey of the diversity of the genus Agaricus in Mexico. In Proceedings of the 7th International Conference on Mushroom Biology and Mushroom Products, Arcachon, France, 4–7 October 2011; pp. 134–139. [Google Scholar]
- Medel, R.; Palestina-Villa, E.N.; Mata, G.; Parra-Sánchez, L.A. An overview of tropical and subtropical Agaricus in Mexico. In Updates on Tropical Mushrooms; Sanchez, J.E., Mata, G., Royse, D.J., Eds.; El Colegio de la Frontera Sur: Chiapas, Mexico, 2018; pp. 39–47. [Google Scholar]
- Palestina-Villa, E.N.; Parra-Sánchez, L.A.; Villegas, M.; Garibay-Orijel, R.; Medel-Ortiz, R. Las especies conocidas de Agaricus (Agaricales, Agaricaceae) en méxico, una actualización y revisión nomenclatural. Sci. Fungorum 2020, 50, e1269. [Google Scholar] [CrossRef]
- Parra, L.A.; Angelini, C.; Ortiz-Santana, B.; Mata, G.; Rojo, C.; Chen, J.; Callac, P. The genus Agaricus in the Caribbean. Nine new taxa mostly based on collections from the Dominican Republic. Phytotaxa 2018, 345, 219–271. [Google Scholar] [CrossRef]
- Peck, C.H. Report of the botanist (1892). Annu. Rep. N. Y. State Mus. Nat. Hist. 1893, 46, 85–149. [Google Scholar]
- Velázquez-Narváez, A.C.; Medel, R.; Mata, G.; Espinosa-de-los-Monteros, A. Agaricus subrufescens en México: Un recurso forestal no maderable, comestible, medicinal y potencialmente cultivable. Madera Bosques 2018, 24, 1–3. [Google Scholar] [CrossRef]
- Kerrigan, R.W. Agaricus subrufescens, a cultivated edible and medicinal mushroom, and its synonyms. Mycologia 2005, 97, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Parra, L.A. Agaricus L. Allopsalliota Nauta & Bas, Part 2; Fungi Europaei; Candusso Edizioni s.a.s.: Alassio, Italy, 2013; pp. 1–1168. [Google Scholar]
- Thongklang, N.; Hoang, E.; Rodriguez-Estrada, A.E.; Sysouphanthong, P.; Moinard, M.; Hyde, K.D.; Kerrigan, R.W.; Foulongne-Oriol, M.; Callac, P. Evidence for amphithallism and broad geographical hybridization potential among Agaricus subrufescens isolates from Brazil, France, and Thailand. Fungal Biol. 2014, 118, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Moinard, M.; Xu, J.; Wang, S.; Foulongne-Oriol, M.; Zhao, R.; Hyde, K.D.; Callac, P. Genetic Analyses of the Internal Transcribed Spacer Sequences Suggest Introgression and Duplication in the Medicinal Mushroom Agaricus subrufescens. PLoS ONE 2016, 11, e0157927. [Google Scholar] [CrossRef]
- Pegler, D.N. Agaric Flora of the Lesser Antilles. Kew Bull. Addit. Ser. 1983, 9, 1–667. [Google Scholar]
- Heinemann, P.; Rammeloo, J. De la mesure des spores et son expression. Agarica 1985, 6, 366–380. [Google Scholar]
- Parra, L.A. Agaricus L. Allopsalliota Nauta & Bas, Part 1; Fungi Europaei; Edizioni Candusso: Alassio, Italy, 2008; pp. 1–823. [Google Scholar]
- Holmgren, P.K.; Holmgren, N.H. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium. Available online: http://sweetgum.nybg.org/ih (accessed on 31 January 2022).
- Zolan, M.E.; Pukkila, P.J. Inheritance of DNA methylation in Coprinus cinereus. Mol. Cell. Biol. 1986, 6, 195–200. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.M.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–321. [Google Scholar]
- Rehner, S.A.; Buckley, E. A beauveria phylogeny inferred from nuclear ITS and EF1-a sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Ihrmark, K.; Bödeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandström-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2 region--evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.L.; Desjardin, D.E.; Soytong, K.; Perry, B.A.; Hyde, K.D. A monograph of Micropsalliota in Northern Thailand based on morphological and molecular data. Fungal Divers. 2010, 45, 33–79. [Google Scholar] [CrossRef]
- Mullis, K.B.; Faloona, F.A. [21] Specific synthesis of dna in vitro via a polymerase-catalyzed chain reaction. Methods Enzymol. 1987, 155, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Izzo, A.; Agbowo, J.; Bruns, T.D. Detection of plot-level changes in ectomycorrhizal communities across years in an old-growth mixed-conifer forest. New Phytol. 2005, 166, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Gui, Y.; Zhu, G.S.; Callac, P.; Hyde, K.D.; Parra, L.A.; Chen, J.; Yang, T.J.; Huang, W.B.; Gong, G.L.; Liu, Z.Y. Agaricus section Arvenses: Three new species inhighland subtropical southwest china. Fungal Biol. 2015, 119, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest Version 2; Program distributed by the author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer v1.6. 2014. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 5 April 2022).
- Callac, P.; Imbernon, M.; Guinberteau, J.; Desmerger, C. A novel homothallic variety of Agaricus bisporus comprises rare tetrasporic isolates from europe. Mycologia 2003, 95, 222–231. [Google Scholar] [CrossRef]
- Challen, M.P.; Kerrigan, R.W.; Callac, P. A phylogenetic reconstruction and emendation of Agaricus section Duploannulatae. Mycologia 2003, 95, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Parra, L.A.; De Kesel, A.; Khalid, A.N.; Qasim, T.; Ashraf, A.; Bahkali, A.H.; Hyde, K.D.; Zhao, R.; Callac, P. Inter- and intraspecific diversity in Agaricus endoxanthus and allied species reveals a new taxon, A. punjabensis. Phytotaxa 2016, 252, 1–16. [Google Scholar] [CrossRef]
- Rocha de Brito, M.; Foulongne-Oriol, M.; Moinard, M.; Souza Dias, E.; Savoie, J.M.; Callac, P. Spore behaviors reveal a category of mating-competent infertile heterokaryons in the offspring of the medicinal fungus Agaricus subrufescens. Appl. Microbiol. Biotechnol. 2016, 100, 781–796. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Specimen | ITS | tef1 | Geographic Origin |
|---|---|---|---|---|
| A. abruptibulbus | ZRL20150970 | MK617880 | MK614388 | Yunnan, China |
| A. abruptibulbus | ZRL20161250 | MK617909 | MK614417 | Jilin, China |
| A. albiceps | ZRL20170538 T | MK617917 | MK614424 | Inner Mongolia, China |
| A. alboumbonatus | ZRL20171140 T | MK617943 | MK614449 | Heilongjiang, China |
| A. arvensis | LAPAG450 T | KF114474 | KX198047 | Spain |
| A. augustus | CA590 | JF797193 | MK614368 | France |
| A. catenocystis | ZRL20170624 T | MK617925 | MK614432 | Inner Mongolia, China |
| A. cf. essettei | ZRL2012026 | KT951363 | KT951630 | Yunnan, China |
| A. crocodilinus | HSU7280 T | NR144989 | – | California, USA |
| A. cruciquercorum | RWK2121 | KJ859120 | – | California, USA |
| A. cruciquercorum | RWK2124 | KJ859127 | – | California, USA |
| A. didymus | RWK2235 T | NR144990 | – | New Mexico, USA |
| A. diospyros | BOS722 T | NR144993 | – | Wisconsin, USA |
| A. dolichocaulis | ZRL20170590 T | MK617923 | MK614430 | Inner Mongolia, China |
| A. eburneocanus | H7528 T | JF495062 | – | Australia |
| A. essettei | LAPAG676 | KJ548131 | – | Savona, Italy |
| A. essettei | ZRL20120642 | MK617870 | MK614379 | Tibet, China |
| A. fissuratus | ZRL20170745 | MK617935 | MK614441 | Inner Mongolia, China |
| A. flocculosipes | ZRL2012105 | KT951365 | KT951618 | Yunnan, China |
| A. flocculosipes | ZRL3028 T | JN664954 | – | Doi Inthanon, Thailand |
| A. gemellatus | ZRL2012631 | KT951380 | KT951623 | Tibet, China |
| A. gemellatus | RWK2047 T | KJ859080 | – | Utah, USA |
| A. greuteri | LAPAG399 | MK617859 | – | Palencia, Spain |
| A. greuteri | PAL-GR57140 T | KF114473 | – | Emilia-Romagna, Italy |
| A. guizhouensis | ZRL20160420 | MK617906 | MK614414 | Heilongjiang, China |
| A. guizhouensis | GY127800 T | KJ755658 | – | Guizhou, China |
| A. hingganensis | ZRL20151825 | MK617898 | MK614406 | Inner Mongolia, China |
| A. hingganensis | ZRL20151696 T | MK617882 | MK614390 | Heilongjiang, China |
| A. inapertus | HS1 | KJ859088 | – | California, USA |
| A. indistinctus | AH44504 T | KF114475 | – | Burgos, Spain |
| A. julius | ZRL20162195 | MK617915 | MK614422 | Gansu, China |
| A. julius | RWK2267 T | NR144991 | – | New Mexico, USA |
| A. longistipes | GY124459 | KJ755642 | – | Guizhou, China |
| A. longistipes | GY124427 T | KJ755643 | – | Guizhou, China |
| A. luteopileus | ZRL20120589 | MK617868 | MK614377 | Tibet, China |
| A. luteopileus | ZRL2012604 T | KT951375 | KT951620 | Tibet, China |
| A. macrocarpus | LAPAG525 | MK617860 | MK614369 | Bohemia, Czech Republic |
| A. macrochlamys | ARB1353 | KY704305 | – | Georgia, USA |
| A. macrochlamys | CA1110 * | KY704307 | – | Veracruz, Mexico |
| A. macrochlamys | IE903 | ON332803 | – | Veracruz, Mexico |
| A. macrochlamys | IE904 | ON332804 | – | Veracruz, Mexico |
| A. macrochlamys | IE965 | KY114573 | – | Veracruz, Mexico |
| A. macrochlamys | IE973 | KY114574 | – | Veracruz, Mexico |
| A. macrochlamys | IE4012 | ON332805 | ON376243 | Veracruz, Mexico |
| A. macrochlamys | JBSD126480 | ON332793 | ON376239 | Puerto Plata, Dominican Republic |
| A. macrochlamys | JBSD126482 | ON332794 | – | Puerto Plata, Dominican Republic |
| A. macrochlamys | JBSD126483 | ON332795 | ON376240 | Puerto Plata, Dominican Republic |
| A. macrochlamys | JBSD126484 | ON332796 | – | Puerto Plata, Dominican Republic |
| A. macrochlamys | JBSD130773 | ON332797 | ON376241 | Puerto Plata, Dominican Republic |
| A. macrochlamys | JBSD130774 | ON332798 | ON376242 | Puerto Plata, Dominican Republic |
| A. macrochlamys | LD201901 | ON332799 | ON376244 | Veracruz, Mexico |
| A. macrochlamys | LD201927 | ON332800 | – | Veracruz, Mexico |
| A. macrochlamys | LD201934 | ON332801 | – | Veracruz, Mexico |
| A. macrochlamys | LD201938 | ON332802 | ON376245 | Veracruz, Mexico |
| A. macrolepis | LAPAG409 | KJ548128 | – | Burgos, Spain |
| A. megacarpus | QL20170178 T | MK617861 | MK614371 | Gansu, China |
| A. megalocarpus | Hao0608 T | KJ755638 | – | Yunnan, China |
| A. mesocarpus | RWK2240 T | KJ859094 | – | Colorado, USA |
| A. moronii | RWK2045 T | NR144992 | – | Utah, USA |
| A. nanaugustus | MW17 | KJ859117 | – | Pennsylvania, USA |
| A. nanaugustus | RWK2002 | KJ859116 | – | Pennsylvania, USA |
| A. ornatipes | MCVE29263 T | MG664795 | – | Italy |
| A. osecanus | LAPAG90 | KJ548133 | – | Madrid, Spain |
| A. pyricystis | ZRL20161248 | MK617908 | MK614416 | Jilin, China |
| A. pyricystis | ZRL20161191 T | MK617907 | MK614415 | Jilin, China |
| A. reducibulbus | RWK2105 | KJ859130 | – | Pennsylvania, USA |
| A. reducibulbus | RWK2077 T | NR144994 | – | Pennsylvania, USA |
| A. sandianus | RWK2260 T | NR144995 | – | New Mexico, USA |
| A. silvicolisimilis | BP85081 T | KF114472 | – | Budapest, Hungary |
| A. sp. | AMZ | AY484687 | – | Hungary |
| A. sp. | SHY2011073117 | KT951407 | KT951622 | Yunnan, China |
| A. sp. | Thoen6951 | JN204433 | – | Senegal |
| A. sp. | Thoen7297 | JF514542 | – | Senegal |
| A. sp. | TL1644 | JF495060 | – | Australia |
| A. sp. | TL1650 | JF495061 | – | Australia |
| A. sp. | ZRL2012608 | KT951377 | KT951627 | Tibet, China |
| A. sp. | ZRL2012630 | KT951379 | KT951621 | Tibet, China |
| A. sp. | ZRL20160099 | MK617903 | MK614411 | Heilongjiang, China |
| A. sp. | ZRL20160157 | MK617904 | MK614412 | Heilongjiang, China |
| A. sp. | ZRL20162213 | MK617916 | MK614423 | Beijing, China |
| A. sp. | ZRL20170661 | MK617927 | MK614434 | Inner Mongolia, China |
| A. sp. | ZRL20170828 | MK617941 | MK614447 | Heilongjiang, China |
| A. sp. | ZRL20170893 | MK617942 | MK614448 | Heilongjiang, China |
| A. subantarcticus | GAL9419 T | DQ232642 | – | New Zealand |
| A. subrufescens | ZRL2012722 | KT951383 | KT951632 | Yunnan, China |
| A. subrufescens | ZRL20140153 | MK617874 | MK614382 | Yunnan, China |
| A. subrufescens ** | K(W)234316 | ON561779 | – | Martinique |
| A. subrufescens | LD201903 | ON332806 | ON376236 | Veracruz, Mexico |
| A. subrufescens | LAPAM11 | MF511113 | ON376237 | Puerto Plata, Dominican Republic |
| A. subrufescens | LAPAM12 | MF511114 | ON376238 | Puerto Plata, Dominican Republic |
| A. subrufescens | LAPAM13 | MF511115 | – | Puerto Plata, Dominican Republic |
| A. subrufescens | LAPAM37 | MF511128 | – | Puerto Plata, Dominican Republic |
| A. subrufescens | LAPAM77 | ON332807 | ON568203 | Puerto Plata, Dominican Republic |
| A. subrufescens | LAPAM78 | ON332808 | – | Puerto Plata, Dominican Republic |
| A. subrufescens | LAPAM100 | ON332809 | – | Puerto Plata, Dominican Republic |
| A. subrufescens | LAPAM101 | ON332810 | – | Puerto Plata, Dominican Republic |
| A. subrufescens | LAPAM103 | ON332811 | – | Puerto Plata, Dominican Republic |
| A. subtilipes | MFLU120992 | KP705079 | – | Thailand |
| A. subtilipes | LD201201 T | KP705078 | MK614370 | Thailand |
| A. subumbonatus | ZRL2012030 T | KT951364 | KT951628 | Yunnan, China |
| A. summensis | RWK2048 | KJ859105 | – | Utah, USA |
| A. summensis | RWK2243 | KJ859104 | – | Colorado, USA |
| A. sylvicola | LAPAG242 | KJ548130 | – | Burgos, Spain |
| A. sylvicola | ZRL20151792 | MK617894 | MK614402 | Inner Mongolia, China |
| A. edmondoi (outgroup) | LAPAG412 | KT951326 | KT951590 | Segovia, Spain |
| Sample | Country | ITS | Position | GenBank | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Type | 39 | 122 | 130 | 145 | 146 | 166 | 196 | 200 | 210 | 269 | 466 | 475 | 494 | 518 | |||
| IE965 | Mexico | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | KY114573 |
| IE973 | Mexico | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | KY114574 |
| CA1110 | Mexico | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | KY704307 |
| IE903 | Mexico | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332803 |
| IE904 | Mexico | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332804 |
| IE4012 | Mexico | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332805 |
| LD201901 | Mexico | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332799 |
| LD201927 | Mexico | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332800 |
| LD201934 | Mexico | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332801 |
| LD201938 | Mexico | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332802 |
| JBSD126480 | Dominican | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332793 |
| JBSD126482 | Dominican | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332794 |
| JBSD126483 | Dominican | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332795 |
| JBSD126484 | Dominican | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332796 |
| JBSD130773 | Dominican | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332797 |
| JBSD130774 | Dominican | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | ON332798 |
| ARB1353 | USA | MM | - | G | A | A | T | G | G | C | G | G | A | T | A | G | KY704305 |
| F2285 | Martinique | AA | T | A | A | G | A | A | A | T | A | G | A | T | Y | T | JF797201 |
| LAPAM13 | Dominican | AA | T | A | A | G | A | A | A | T | A | G | A | T | T | T | MF511115 |
| LAPAM37 | Dominican | AA | T | A | A | G | A | A | A | T | A | G | A | T | C | T | MF511128 |
| LAPAM77 | Dominican | AA | T | A | A | G | A | A | A | T | A | G | A | T | C | T | ON332807 |
| LAPAM78 | Dominican | AA | T | A | A | G | A | A | A | T | A | G | A | T | Y | T | ON332808 |
| LAPAM100 | Dominican | AA | T | A | A | G | A | A | A | T | A | G | A | T | Y | T | ON332809 |
| LAPAM101 | Dominican | AA | T | A | A | G | A | A | A | T | A | G | A | T | C | T | ON332810 |
| LAPAM103 | Dominican | AA | T | A | A | G | A | A | A | T | A | G | A | T | C | T | ON332811 |
| CA603A | Mexico | A | T | A | A | G | A | A | A | T | A | G | A | T | C | T | KY704306 1 |
| WC837-S43 | Brazil | A | T | A | A | G | A | A | A | T | A | G | A | T | C | T | KJ541797 |
| CA864A | France | A | T | A | A | G | A | A | A | T | A | G | A | T | T | T | KU557349 1 |
| CA487-C5 | France | A | T | A | A | G | A | A | A | T | A | G | A | T | C | T | KU557352 |
| CA603B | Mexico | B | - | G | G | A | T | A | A | C | A | G | A | T | C | T | KY704306 1 |
| L0341732 | UK | BB | - | G | G | A | T | A | A | C | A | G | A | T | C | T | AY818649 |
| IE913 | Mexico | BB | - | G | G | A | T | A | A | C | A | G | A | T | C | T | KY114572 |
| LAPAM12 | Dominican | BB | - | G | G | A | T | A | A | C | A | G | A | T | C | T | MF511114 |
| WC837-S04 | Brazil | B | - | G | G | A | T | A | A | C | A | G | A | T | C | T | KJ541796 |
| CA864B | France | B | - | G | G | A | T | A | A | C | A | G | A | T | C | T | KU557349 1 |
| CA487-C6 | France | B | - | G | G | A | T | A | A | C | A | G | A | T | C | T | KU557353 |
| CA487-C2 | France | C | - | G | G | A | T | A | A | C | A | A | C | - | C | T | KU557351 |
| GY128883 | China | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | KJ755634 |
| GY133048 | China | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | KJ755635 |
| GY121118 | China | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | KJ755633 |
| GY128956 | China | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | KJ755632 |
| XHW1614 | China | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | KJ755636 |
| ZRL2036 | Thailand | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | KU557345 |
| ZRL2134 | Thailand | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | KU557346 |
| ZRL20130601 | China | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | MK617873 |
| VM098 | Iran | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | KT983412 |
| NT001 | Thailand | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | KU557347 |
| CA935 | Thailand | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | KU557348 |
| CA918 | Thailand | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | KJ541798 |
| NTF67 | Thailand | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | JF514529 |
| DEH513 | Hawaii | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | AY818646 |
| DEH1073 | Hawaii | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | AY818647 |
| KRP070 | Hawaii | CC | - | G | G | A | T | A | A | C | A | A | C | - | C | T | AY818648 |
| CA487 | France | ABC | T/- | R | R | R | W | A | A | Y | A | R | M | T/- | C | T | KU557351 2 KU557352 2 KU557353 2 |
| CA603 | Mexico | AB | T/- | R | R | R | W | A | A | Y | A | G | A | T | C | T | KY704306 |
| LD201903 | Mexico | AB | T | R | R | A | T | A | A | Y | A | G | A | T | C | T | ON332806 |
| LAPAM11 | Dominican | AB | T | R | R | R | W | A | A | Y | A | G | A | T | Y | T | MF511113 |
| WC837 | Brazil | AB | T/- | R | R | R | W | A | A | Y | A | G | A | T | C | T | KU557350 |
| CA864 | France | AB | T/- | R | R | R | W | A | A | Y | A | G | A | T | Y | T | KU557349 |
| Fragment 10 | USA | (AB) | T/- | R | R | R | W | A | A | Y | A | G | A | T | C | T | EU071699 |
| SBRF | USA | (AB) | T/- | R | R | R | W | A | A | Y | A | G | A | T | C | T | AY818656 |
| ZRL2012722 | China | (aB) | - | A | G | A | T | A | A | C | A | G | A | - | C | T | KT951383 |
| ZRL20140153 | China | (aB) | - | A | G | A | T | A | A | C | A | G | A | - | C | T | MK617874 |
| 978 | Brazil | (aB) | - | G | G | R | T | A | A | C | A | G | A | T | C | T | AY818650 |
| GY129365 | China | (bC) | - | G | G | A | T | A | A | C | A | R | M | - | C | T | KJ755631 |
| ZRL20140275 | China | (bC) | - | G | G | A | T | A | A | C | A | G | A | - | C | T | MK617876 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medel-Ortiz, R.; Garibay-Orijel, R.; Argüelles-Moyao, A.; Mata, G.; Kerrigan, R.W.; Bessette, A.E.; Geml, J.; Angelini, C.; Parra, L.A.; Chen, J. Agaricus macrochlamys, a New Species from the (Sub)tropical Cloud Forests of North America and the Caribbean, and Agaricus fiardii, a New Synonym of Agaricus subrufescens. J. Fungi 2022, 8, 664. https://doi.org/10.3390/jof8070664
Medel-Ortiz R, Garibay-Orijel R, Argüelles-Moyao A, Mata G, Kerrigan RW, Bessette AE, Geml J, Angelini C, Parra LA, Chen J. Agaricus macrochlamys, a New Species from the (Sub)tropical Cloud Forests of North America and the Caribbean, and Agaricus fiardii, a New Synonym of Agaricus subrufescens. Journal of Fungi. 2022; 8(7):664. https://doi.org/10.3390/jof8070664
Chicago/Turabian StyleMedel-Ortiz, Rosario, Roberto Garibay-Orijel, Andrés Argüelles-Moyao, Gerardo Mata, Richard W. Kerrigan, Alan E. Bessette, József Geml, Claudio Angelini, Luis A. Parra, and Jie Chen. 2022. "Agaricus macrochlamys, a New Species from the (Sub)tropical Cloud Forests of North America and the Caribbean, and Agaricus fiardii, a New Synonym of Agaricus subrufescens" Journal of Fungi 8, no. 7: 664. https://doi.org/10.3390/jof8070664
APA StyleMedel-Ortiz, R., Garibay-Orijel, R., Argüelles-Moyao, A., Mata, G., Kerrigan, R. W., Bessette, A. E., Geml, J., Angelini, C., Parra, L. A., & Chen, J. (2022). Agaricus macrochlamys, a New Species from the (Sub)tropical Cloud Forests of North America and the Caribbean, and Agaricus fiardii, a New Synonym of Agaricus subrufescens. Journal of Fungi, 8(7), 664. https://doi.org/10.3390/jof8070664