
Synergism between Streptomyces viridosporus HH1 and Rhizophagus irregularis Effectively Induces Defense Responses to Fusarium Wilt of Pea and Improves Plant Growth and Yield




Abstract
:1. Introduction
2. Materials and Methods
2.1. Pea Cultivar, Pathogen, and Bioagents
2.2. Assessment of Antifungal Activity of S. viridosporus HH1 In Vitro
2.3. Gas Chromatography/Mass Spectrometry (GC/MS) Analysis
2.4. Greenhouse Evaluation
2.4.1. Gene Expression Profiling
2.4.2. Growth and Yield Evaluation
2.4.3. Disease Assessment
2.4.4. Phenolic Content and Activity of Antioxidant Enzymes
2.4.5. Biochemical Analyses
2.4.6. Evaluation of Mycorrhization Level
2.4.7. Ultrastructural Investigation
2.5. Statistical Analyses
3. Results
3.1. Antifungal Activity of S. viridosporus HH1 against F. oxysporum f.sp. pisi In Vitro
3.2. GC-MS
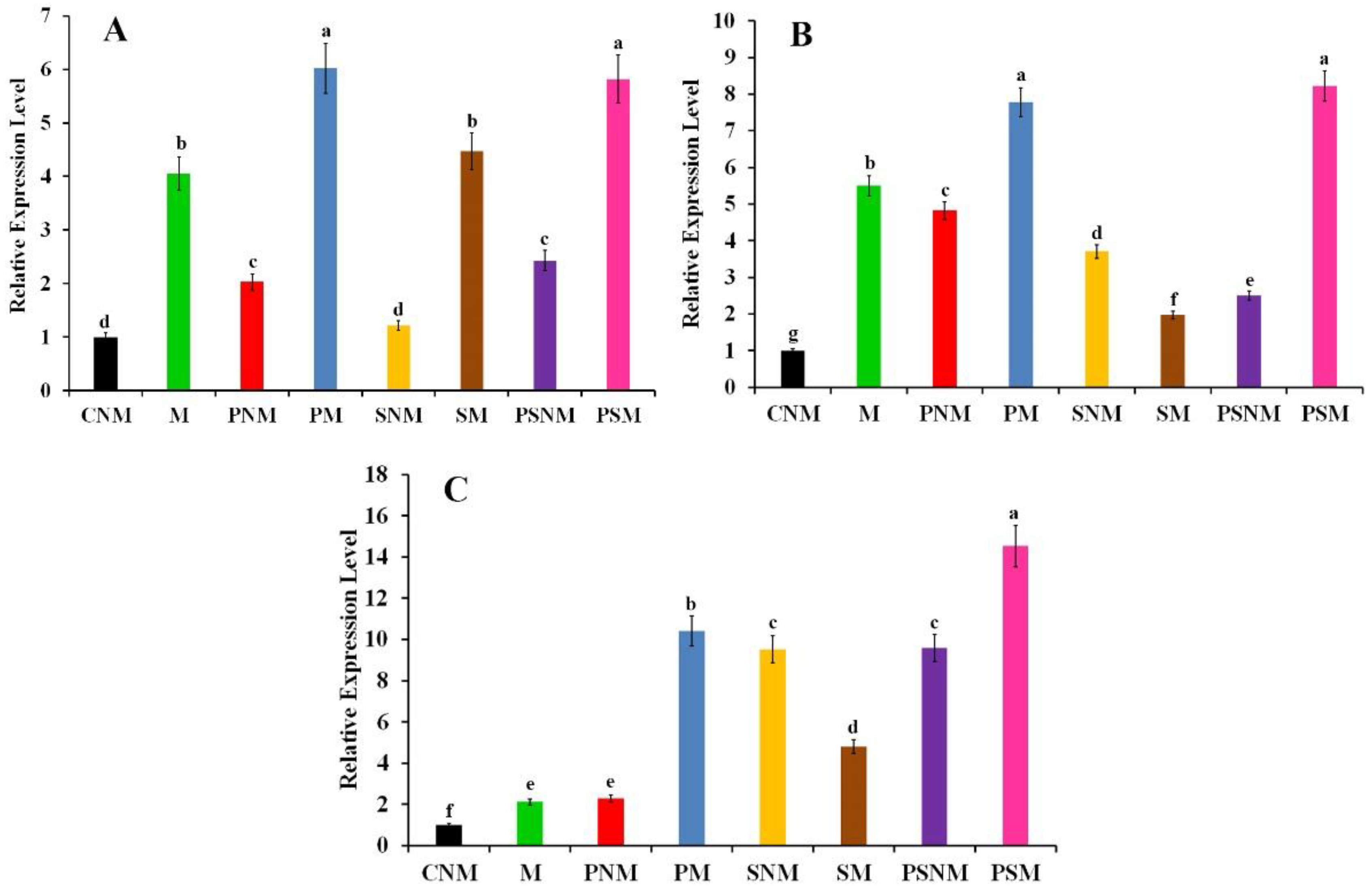
3.3. Gene Expression Profiling
3.4. Growth Parameters
3.5. Yield and Its Components
3.6. Disease Severity
3.7. Total Phenolics, Activity of Antioxidant Enzymes, Cell Electrolyte Leakage, and TSS
3.8. Photosynthetic Pigments
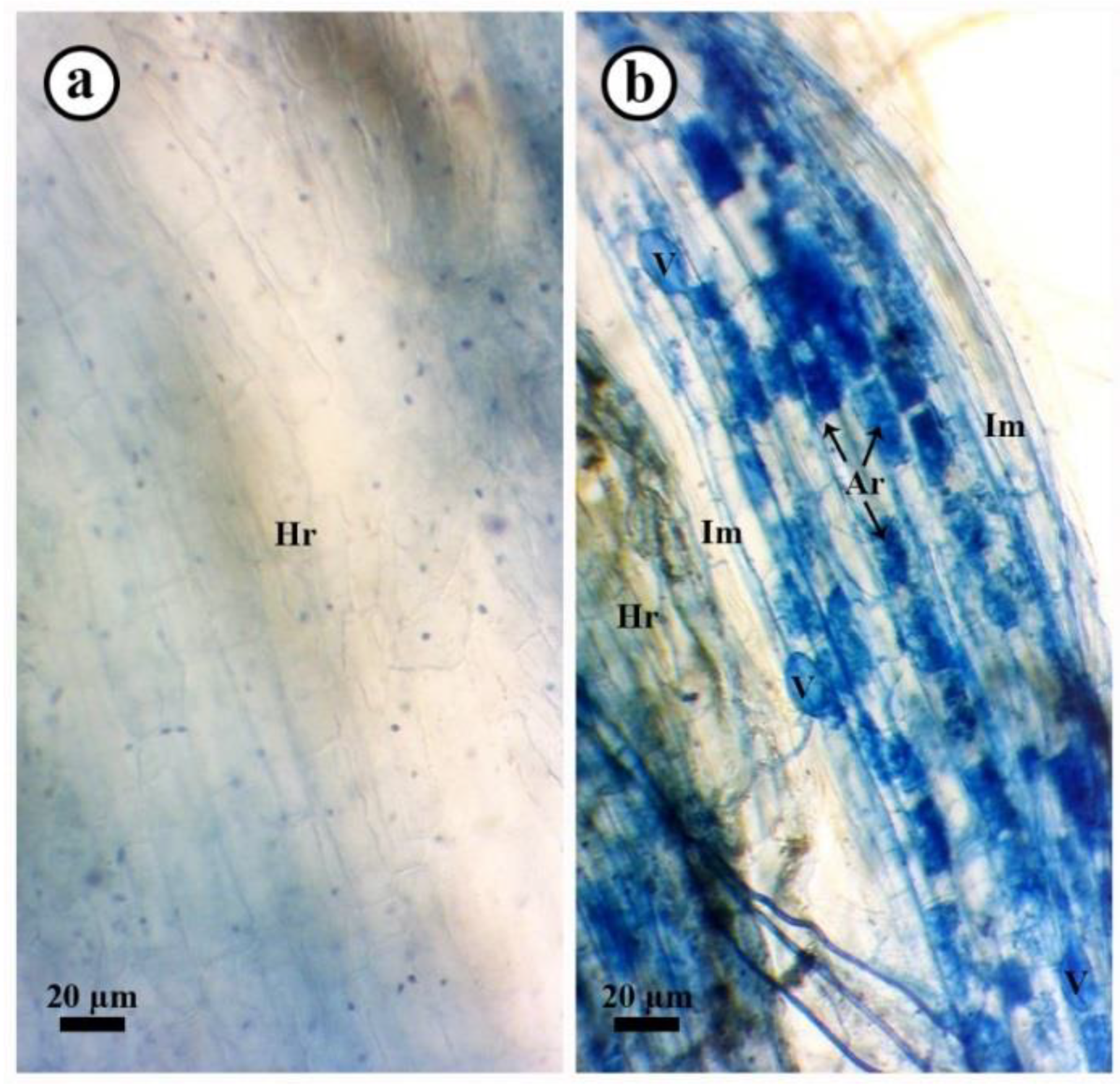
3.9. Mycorrhizal Colonization Level
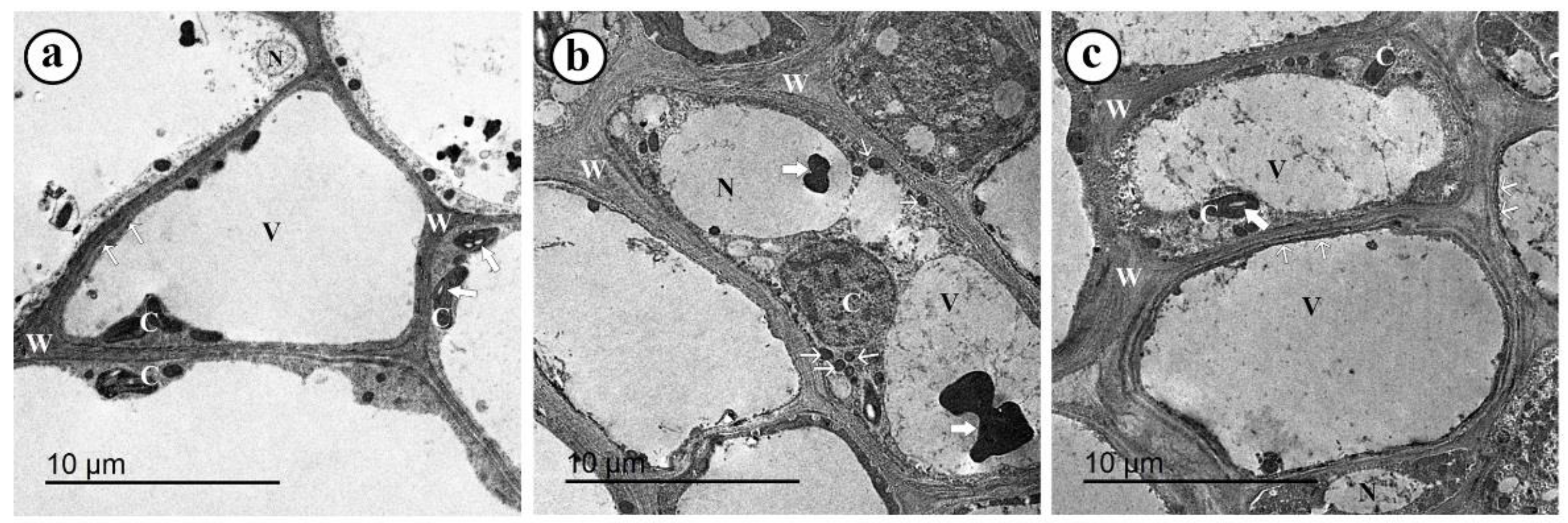
3.10. TEM Observations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT Food and Agriculture Organization of the United Nations. 2022. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 10 May 2022).
- Karkanis, A.; Ntatsi, G.; Kontopoulou, C.-K.; Pristeri, A.; Bilalis, D.; Savvas, D. Field Pea in European Cropping Systems: Adaptability, Biological Nitrogen Fixation and Cultivation Practices. Not. Bot. Horti Agrobot. Cluj-Napoca 2016, 44, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Kraft, J. Fusarium wilt of peas (a review). Agronomie 1994, 14, 561–567. [Google Scholar] [CrossRef] [Green Version]
- Harveson, R.M.; Pasche, J.S.; Porter, L.D.; Chen, W.; Burrows, M. Compendium of Pea Diseases and Pests, 3rd ed.; American Phytopathological Society (APS Press): Saint Paul, MI, USA, 2020; ISBN 0890542694. [Google Scholar]
- Aslam, S.; Ghazanfar, M.U.; Munir, N.; Hamid, M.I. Managing fusarium wilt of pea by utilizing different application methods of fungicides. Pakistan J. Phytopathol. 2019, 31, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Gao, X.; Nan, J.; Zhang, T.; Xie, X.; Cai, Q. Fungicides alter the distribution and diversity of bacterial and fungal communities in ginseng fields. Bioengineered 2021, 12, 8043–8056. [Google Scholar] [CrossRef]
- Rashad, Y.M.; Moussa, T.A.A. Biocontrol Agents for Fungal Plant Diseases Management. In Cottage Industry of Biocontrol Agents and Their Applications: Practical Aspects to Deal Biologically with Pests and Stresses Facing Strategic Crops; El-Wakeil, N., Saleh, M., Abu-hashim, M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 337–363. ISBN 978-3-030-33161-0. [Google Scholar]
- de Lima Procópio, R.E.; da Silva, I.R.; Martins, M.K.; de Azevedo, J.L.; de Araújo, J.M. Antibiotics produced by Streptomyces. Brazilian J. Infect. Dis. 2012, 16, 466–471. [Google Scholar] [CrossRef] [Green Version]
- Usha Nandhini, S.; Sudha, S.; Anusha Jeslin, V.; Manisha, S. Isolation, identification and extraction of antimicrobial compounds produced by Streptomyces sps from terrestrial soil. Biocatal. Agric. Biotechnol. 2018, 15, 317–321. [Google Scholar] [CrossRef]
- Viaene, T.; Langendries, S.; Beirinckx, S.; Maes, M.; Goormachtig, S. Streptomyces as a plant’s best friend? FEMS Microbiol. Ecol. 2016, 92, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Pacios-Michelena, S.; Aguilar González, C.N.; Alvarez-Perez, O.B.; Rodriguez-Herrera, R.; Chávez-González, M.; Arredondo Valdés, R.; Ascacio Valdés, J.A.; Govea Salas, M.; Ilyina, A. Application of Streptomyces Antimicrobial Compounds for the Control of Phytopathogens. Front. Sustain. Food Syst. 2021, 5, 1–13. [Google Scholar] [CrossRef]
- Al-Askar, A.A.; Abdulkhair, W.M.; Rashad, Y.M.; Hafez, E.E.; Ghoneem, K.M.; Baka, Z.A. Streptomyces griseorubens E44G: A Potent Antagonist Isolated from Soil in Saudi Arabia. J. Pure Appl. Microbiol. 2014, 8, 221–230. [Google Scholar]
- LeBlanc, N. Bacteria in the genus Streptomyces are effective biological control agents for management of fungal plant pathogens: A meta-analysis. BioControl 2022, 67, 111–121. [Google Scholar] [CrossRef]
- El-Sharkawy, H.H.A.; Abo-El-Wafa, T.S.A.; Ibrahim, S.A.A. Biological control agents improve the productivity and induce the resistance against downy mildew of grapevine. J. Plant Pathol. 2018, 100, 33–42. [Google Scholar] [CrossRef]
- El-Sharkawy, H.H.A.; Tohamey, S.; Khalil, A.A. Combined Effects of Streptomyces viridosporus and Trichoderma harzianum on Controlling Wheat Leaf Rust Caused by Puccinia triticina. Plant Pathol. J. 2015, 14, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Rashad, Y.M.; Al-Askar, A.A.; Ghoneem, K.M.; Saber, W.I.A.; Hafez, E.E. Chitinolytic Streptomyces griseorubens E44G enhances the biocontrol efficacy against Fusarium wilt disease of tomato. Phytoparasitica 2017, 45, 227–237. [Google Scholar] [CrossRef]
- Jones, S.E.; Ho, L.; Rees, C.A.; Hill, J.E.; Nodwell, J.R.; Elliot, M.A. Streptomyces exploration is triggered by fungal interactions and volatile signals. eLife 2017, 6, e21738. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and interactions in plant growth promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, E.M.; Kunova, A.; Cortesi, P.; Saracchi, M.; Pasquali, M. Critical Assessment of Streptomyces spp. Able to Control Toxigenic Fusaria in Cereals: A Literature and Patent Review. Int. J. Mol. Sci. 2019, 20, 6119. [Google Scholar] [CrossRef] [Green Version]
- de Jesus Sousa, J.A.; Olivares, F.L. Plant growth promotion by streptomycetes: Ecophysiology, mechanisms and applications. Chem. Biol. Technol. Agric. 2016, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- El-Sharkawy, H.H.A.; Rashad, Y.M.; Ibrahim, S.A. Biocontrol of stem rust disease of wheat using arbuscular mycorrhizal fungi and Trichoderma spp. Physiol. Mol. Plant Pathol. 2018, 103, 84–91. [Google Scholar] [CrossRef]
- Aseel, D.G.; Rashad, Y.M.; Hammad, S.M. Arbuscular Mycorrhizal Fungi Trigger Transcriptional Expression of Flavonoid and Chlorogenic Acid Biosynthetic Pathways Genes in Tomato against Tomato Mosaic Virus. Sci. Rep. 2019, 9, 9692. [Google Scholar] [CrossRef] [Green Version]
- Rashad, Y.M.; El-Sharkawy, H.H.A.; Elazab, N.T. Ascophyllum nodosum Extract and Mycorrhizal Colonization Synergistically Trigger Immune Responses in Pea Plants against Rhizoctonia Root Rot, and Enhance Plant Growth and Productivity. J. Fungi 2022, 8, 268. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A.; et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Fattah, G.M.; El-Haddad, S.A.; Hafez, E.E.; Rashad, Y.M. Induction of defense responses in common bean plants by arbuscular mycorrhizal fungi. Microbiol. Res. 2011, 166, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Rashad, Y.M.; Fekry, W.M.E.; Sleem, M.M.; Elazab, N.T. Effects of Mycorrhizal Colonization on Transcriptional Expression of the Responsive Factor JERF3 and Stress-Responsive Genes in Banana Plantlets in Response to Combined Biotic and Abiotic Stresses. Front. Plant Sci. 2021, 12, 742628. [Google Scholar] [CrossRef]
- Rashad, Y.M.; Abbas, M.A.; Soliman, H.M.; Abdel-Fattah, G.G.; Abdel-Fattah, G.M. Synergy between endophytic Bacillus amyloliquefaciens GGA and arbuscular mycorrhizal fungi induces plant defense responses against white rot of garlic and improves host plant growth. Phytopathol. Mediterr. 2020, 59, 169–186. [Google Scholar] [CrossRef]
- Rashad, Y.; Aseel, D.; Hammad, S.; Elkelish, A. Rhizophagus irregularis and Rhizoctonia solani differentially elicit systemic transcriptional expression of polyphenol biosynthetic pathways genes in sunflower. Biomolecules 2020, 10, 379. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, V.; Vashisht, D.; Cletus, J.; Sakthivel, N. Plant β-1,3-glucanases: Their biological functions and transgenic expression against phytopathogenic fungi. Biotechnol. Lett. 2012, 34, 1983–1990. [Google Scholar] [CrossRef]
- Breen, S.; Williams, S.J.; Outram, M.; Kobe, B.; Solomon, P.S. Emerging Insights into the Functions of Pathogenesis-Related Protein 1. Trends Plant Sci. 2017, 22, 871–879. [Google Scholar] [CrossRef]
- Summerell, B.A.; Salleh, B.; Leslie, J.F. A utilitarian approach to Fusarium identification. Plant Dis. 2003, 87, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Booth, C. The Genus Fusarium; Commonwealth Mycological Institute: Kew, Surrey, UK, 1977. [Google Scholar]
- Fuentes, A.; Ortiz, J.; Saavedra, N.; Salazar, L.A.; Meneses, C.; Arriagada, C. Reference gene selection for quantitative real-time PCR in Solanum lycopersicum L. inoculated with the mycorrhizal fungus Rhizophagus irregularis. Plant Physiol. Biochem. 2016, 101, 124–131. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Bani, M.; Rubiales, D.; Rispail, N. A detailed evaluation method to identify sources of quantitative resistance to Fusarium oxysporum f. sp. pisi race 2 within a Pisum spp. germplasm collection. Plant Pathol. 2012, 61, 532–542. [Google Scholar] [CrossRef] [Green Version]
- Malik, C.P.; Singh, M.B. Estimation of total phenols. In Plant Enzymology and Histo-Enzymology; Kalyani Publishers: New Delhi, India, 1980. [Google Scholar]
- Maxwell, D.P.; Bateman, D.F. Changes in the activities of some oxidases in extracts of Rhizoctonia-infected bean hypocotyls in relation to lesion maturation. Phytopathology 1967, 57, 132–136. [Google Scholar]
- Galeazzi, M.A.M.; Sgarbieri, V.C.; Constantinides, S.M. Isolation, purification and physicochemical characterization of polyphenol oxidase (PPO) from a dwarf variety of banana (Musa cavendishii, L.). J. Food Sci. 1981, 46, 150–155. [Google Scholar] [CrossRef]
- Harborne, J.B. Phytochemical Methods: A Guide to Modern Techniques of Plant Analysis; Chapman and Hall: London, UK, 1984. [Google Scholar]
- Shi, Q.; Bao, Z.; Zhu, Z.; Ying, Q.; Qian, Q. Effects of different treatments of salicylic acid on heat tolerance, chlorophyll fluorescence, and antioxidant enzyme activity in seedlings of Cucumis sativa L. Plant Growth Regul. 2006, 48, 127–135. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN18. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. du taux de mycorhization VA d’un système radiculaire. Recherche de méthodes d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; INRA: Paris, France, 1986; pp. 217–220. [Google Scholar]
- Qi, D.; Zou, L.; Zhou, D.; Chen, Y.; Gao, Z.; Feng, R.; Zhang, M.; Li, K.; Xie, J.; Wang, W. Taxonomy and broad-spectrum antifungal activity of streptomycessp. SCA3-4 isolated from rhizosphere soil of opuntia stricta. Front. Microbiol. 2019, 10, 1390. [Google Scholar] [CrossRef]
- Qi, D.; Zou, L.; Zhou, D.; Zhang, M.; Wei, Y.; Li, K.; Zhao, Y.; Zhang, L.; Xie, J. Biocontrol potential and antifungal mechanism of a novel Streptomyces sichuanensis against Fusarium oxysporum f. sp. cubense tropical race 4 in vitro and in vivo. Appl. Microbiol. Biotechnol. 2022, 106, 1633–1649. [Google Scholar] [CrossRef]
- Christias, C. Inhibition of Microsclerotium Formation in Verticillium Dahliae by Thioglycolic Acid in Infected Cotton Stems. In Proceedings of the Vascular Wilt Diseases of Plants; Tjamos, E.C., Beckman, C.H., Eds.; Springer: Berlin/Heidelberg, Germany, 1989; pp. 521–527. [Google Scholar]
- Barakat, K.M.; Beltagy, E.A. Bioactive phthalate from marine Streptomyces ruber EKH2 against virulent fish pathogens. Egypt. J. Aquat. Res. 2015, 41, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Law, J.W.-F.; Ser, H.-L.; Khan, T.M.; Chuah, L.-H.; Pusparajah, P.; Chan, K.-G.; Goh, B.-H.; Lee, L.-H. The Potential of Streptomyces as Biocontrol Agents against the Rice Blast Fungus, Magnaporthe oryzae (Pyricularia oryzae). Front. Microbiol. 2017, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Soffan, A.; Alghamdi, S.S.; Aldawood, A.S.; Palumbo, J. Peroxidase and polyphenol oxidase activity in moderate resistant and susceptible Vicia faba induced by Aphis craccivora (Hemiptera: Aphididae) infestation. J. Insect Sci. 2014, 14, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Maruri-López, I.; Figueroa, N.E.; Hernández-Sánchez, I.E.; Chodasiewicz, M. Plant Stress Granules: Trends and Beyond. Front. Plant Sci. 2021, 12, 722643. [Google Scholar] [CrossRef]
- Pontier, D.; del Pozo, O.; Lam, E. Cell Death in Plant Disease: Mechanisms and Molecular Markers. In Plant Cell Death Processes; Noodén, L.D., Ed.; Academic Press: San Diego, MI, USA, 2003; pp. 37–50. ISBN 9780125209151. [Google Scholar]
- Koyano, T.; Kurusu, T.; Hanamata, S.; Kuchitsu, K. Regulation of Vacuole-Mediated Programmed Cell Death During Innate Immunity and Reproductive Development in Plants. In Proceedings of the Sexual Reproduction in Animals and Plants; Sawada, H., Inoue, N., Iwano, M., Eds.; Springer: Tokyo, Japan, 2014; pp. 431–440. [Google Scholar]
- Park, K.Y.; Seo, S.Y.; Oh, B.R.; Seo, J.W.; Kim, Y.J. 2,3-butanediol Induces Systemic Acquired Resistance in the Plant Immune Response. J. Plant Biol. 2018, 61, 424–434. [Google Scholar] [CrossRef]
- Yi, H.S.; Ahn, Y.R.; Song, G.C.; Ghim, S.Y.; Lee, S.; Lee, G.; Ryu, C.M. Impact of a bacterial volatile 2,3-butanediol on Bacillus subtilis rhizosphere robustness. Front. Microbiol. 2016, 7, 993. [Google Scholar] [CrossRef] [Green Version]
- El-Sharkawy, H.H.A.; Abbas, M.S.; Soliman, A.S.; Ibrahim, S.A.; El-Nady, I.A.I. Synergistic effect of growth-promoting microorganisms on bio-control of Fusarium oxysporum f. sp. pisi, growth, yield, physiological and anatomical characteristics of pea plants. Pestic. Biochem. Physiol. 2021, 178, 104939. [Google Scholar] [CrossRef]
- Figueiredo, A.F.; Boy, J.; Guggenberger, G. Common Mycorrhizae Network: A Review of the Theories and Mechanisms Behind Underground Interactions. Front. Fungal Biol. 2021, 2, 735299. [Google Scholar] [CrossRef]
- Sato, T.; Hachiya, S.; Inamura, N.; Ezawa, T.; Cheng, W.; Tawaraya, K. Secretion of acid phosphatase from extraradical hyphae of the arbuscular mycorrhizal fungus Rhizophagus clarus is regulated in response to phosphate availability. Mycorrhiza 2019, 29, 599–605. [Google Scholar] [CrossRef]
- Balestrini, R.; Brunetti, C.; Chitarra, W.; Nerva, L. Photosynthetic traits and nitrogen uptake in crops: Which is the role of arbuscular mycorrhizal fungi? Plants 2020, 9, 1105. [Google Scholar] [CrossRef]
- Pons, S.; Fournier, S.; Chervin, C.; Bécard, G.; Rochange, S.; Dit Frey, N.F.; Pagès, V.P. Phytohormone production by the arbuscular mycorrhizal fungus Rhizophagus irregularis. PLoS ONE 2020, 15, e0240886. [Google Scholar] [CrossRef] [PubMed]
- Amaresan, N.; Kumar, K.; Naik, J.H.; Bapatla, K.G.; Mishra, R.K. Streptomyces in Plant Growth Promotion: Mechanisms and Role. In New and Future Developments in Microbial Biotechnology and Bioengineering: Actinobacteria: Diversity and Biotechnological Applications; Singh, B.P., Gupta, V.K., Passari, A.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 125–135. ISBN 9780444639950. [Google Scholar]
- Myo, E.M.; Ge, B.; Ma, J.; Cui, H.; Liu, B.; Shi, L.; Jiang, M.; Zhang, K. Indole-3-acetic acid production by Streptomyces fradiae NKZ-259 and its formulation to enhance plant growth. BMC Microbiol. 2019, 19, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maglangit, F.; Tong, M.H.; Jaspars, M.; Kyeremeh, K.; Deng, H. Legonoxamines A-B, two new hydroxamate siderophores from the soil bacterium, Streptomyces sp. MA37. Tetrahedron Lett. 2019, 60, 75–79. [Google Scholar] [CrossRef]
- Dahal, B.; NandaKafle, G.; Perkins, L.; Brözel, V.S. Diversity of free-Living nitrogen fixing Streptomyces in soils of the badlands of South Dakota. Microbiol. Res. 2017, 195, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Jaemsaeng, R.; Jantasuriyarat, C.; Thamchaipenet, A. Molecular interaction of 1-aminocyclopropane-1-carboxylate deaminase (ACCD)-producing endophytic Streptomyces sp. GMKU 336 towards salt-stress resistance of Oryza sativa L. cv. KDML105. Sci. Rep. 2018, 8, 1950. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Khanna, K.; Manhas, R.K.; Bhardwaj, R.; Ohri, P.; Alkahtani, J.; Alwahibi, M.S.; Ahmad, P. Insights into the role of streptomyces hydrogenans as the plant growth promoter, photosynthetic pigment enhancer and biocontrol agent against meloidogyne incognita in solanum lycopersicum seedlings. Plants 2020, 9, 1109. [Google Scholar] [CrossRef]
- Passari, A.K.; Upadhyaya, K.; Singh, G.; Abdel-Azeem, A.M.; Thankappan, S.; Uthandi, S.; Hashem, A.; Abd-Allah, E.F.; Malik, J.A.; As, A.; et al. Enhancement of disease resistance, growth potential, and photosynthesis in tomato (Solanum lycopersicum) by inoculation with an endophytic actinobacterium, Streptomyces thermocarboxydus strain BPSAC147. PLoS ONE 2019, 14, e0219014. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.E.; Pham, C.A.; Zambri, M.P.; McKillip, J.; Carlson, E.E.; Elliot, M.A. Streptomyces volatile compounds influence exploration and microbial community dynamics by altering iron availability. MBio 2019, 10, e00171-19. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhou, J.; Li, C.; Ma, Y. Antifungal and plant growth promotion activity of volatile organic compounds produced by Bacillus amyloliquefaciens. Microbiologyopen 2019, 8, e00813. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Description | Abbrev. | Accession No. | Sequence (5′-3′) |
|---|---|---|---|
| Jasmonate and ethylene-responsive factor 3 | JERF3-F JERF3-R | AY383630 | GCCATTTGCCTTCTCTGCTTC GCAGCAGCATCCTTGTCTGA |
| β-1,3-glucanase | GLU-F GLU-R | M80604 | TTTCGATGCCCTTGTGGATT CGGCCAACCACTTTCCGATAC |
| Pathogenesis-related protein 1 | PR1-F PR1-R | M69247 | ACTTGGCATCCCGAGCACAA CTCGGACACCCACAATTGCA |
| Elongation factor 1-α | EF1-α-F EF1-α-R | EC959059 | GAACTGGGTGCTTGATAGGC AACCAAAATATCCGGAGTAAAAGA |
| Peak # | Retention Time (min) | Peak Area (%) | Compound Name |
|---|---|---|---|
| 1 | 1.592 | 13.35 | Thioglycolic acid |
| 2 | 3.139 | 17.26 | 2,3-Butanediol |
| 3 | 3.266 | 8.90 | d-2,3-Butanediol |
| 4 | 3.647 | 9.21 | 2-Ethoxypropane |
| 5 | 5.003 | 0.09 | 1-Methoxy-2-butanol |
| 6 | 5.891 | 0.05 | Acetal |
| 7 | 6.895 | 0.05 | Isopropyl isobutyrate |
| 8 | 17.558 | 4.88 | Cyclohexasiloxane, dodecamethyl |
| 9 | 22.661 | 11.74 | Cycloheptasiloxane, tetradecamethyl- |
| 10 | 24.470 | 0.32 | Isopropoxy-3,3,3-trimethyl-[(trimethylsilyl)oxy] |
| 11 | 26.910 | 6.65 | Hexasiloxane, tetradecamethyl- |
| 12 | 27.424 | 0.26 | Homogentisic acid, bis(tert-butyldimethylsilyl) |
| 13 | 28.520 | 0.11 | Tetradecamethylhexasiloxane |
| 14 | 29.643 | 13.78 | Phthalic acid |
| 15 | 30.535 | 1.73 | Hexadecamethylheptasiloxane |
| 16 | 31.900 | 6.94 | Phloroglucinol, tris(trimethylsilyl ether) |
| 17 | 33.761 | 0.55 | Glaucine |
| 18 | 34.659 | 1.52 | Phenyltrimethylsilane |
| 19 | 40.785 | 2.33 | 1,2-Diphenyltetramethyldisilane |
| 20 | 42.271 | 0.27 | Pyrrolidino[1,2-a]piperazine-3,6-dione |
| Treatment | Mycorrhizal Status | Shoot Height (cm) | Root Length (cm) | Shoot Dry Weight (g) | Root Dry Weight (g) | Number of Leaves/Plant | Leaf Area (cm2) |
|---|---|---|---|---|---|---|---|
| C | NM | 46.0 ± 2.6 | 5.7 ± 0.7 | 1.7 ± 0.3 | 0.13 ± 0.04 | 5.3 ± 0.4 | 38.7 ± 3.0 |
| M | 58.7 ± 3.1 | 16.1 ± 0.9 | 2.6 ± 0.5 | 0.18 ± 0.03 | 7.7 ± 0.5 | 52.4 ± 2.7 | |
| P | NM | 36.1 ± 1.9 | 4.2 ± 0.6 | 1.0 ± 0.2 | 0.10 ± 0.04 | 4.0 ± 0.5 | 34.0 ± 2.3 |
| M | 54.3 ± 2.4 | 12.3 ± 0.4 | 2.4 ± 0.3 | 0.15 ± 0.03 | 8.0 ± 0.4 | 44.3 ± 2.8 | |
| S | NM | 55.5 ± 2.7 | 12.0 ± 0.5 | 2.6 ± 0.2 | 0.16 ± 0.02 | 7.3 ± 0.6 | 51.5 ± 3.1 |
| M | 62.7 ± 2.5 | 18.5 ± 0.9 | 2.8 ± 0.6 | 0.21 ± 0.04 | 7.3 ± 0.6 | 61.4 ± 3.6 | |
| PS | NM | 51.2 ± 1.9 | 10.3 ± 0.7 | 1.6 ± 0.2 | 0.13 ± 0.02 | 6.7 ± 0.9 | 51.1 ± 2.9 |
| M | 58.8 ± 2.1 | 16.3 ± 0.6 | 2.4 ± 0.3 | 0.19 ± 0.03 | 6.7 ± 0.3 | 55.1 ± 3.2 | |
| PF | NM | 44.8 ± 2.0 | 5.7 ± 0.3 | 1.7 ± 0.2 | 0.13 ± 0.04 | 5.3 ± 0.3 | 35.5 ± 2.5 |
| M | 49.5 ± 1.8 | 7.3 ± 0.6 | 2.1 ± 0.4 | 0.14 ± 0.05 | 7.3 ± 0.4 | 36.4 ± 2.0 | |
| LSD (p < 0.05) | 3.5 | 1.5 | 0.7 | 0.03 | 1.28 | 4.4 | |
| Treatment | *** | *** | ** | * | * | *** | |
| Mycorrhiza | ** | ** | *** | ** | * | *** | |
| Treatment × Mycorrhiza | *** | ** | ** | * | * | *** | |
| Treatment | Mycorrhizal Status | No. Of Pods/Plant | Pod Weight (g) | Pod Length (cm) | Pod Width (cm) | Yield/Plant (g) | No. of Seeds/Pod |
|---|---|---|---|---|---|---|---|
| C | NM | 2.0 ± 0.6 | 4.6 ± 0.8 | 5.4 ± 0.3 | 1.5 ± 0.05 | 9.1 ± 0.8 | 4.7 ± 0.6 |
| M | 3.3 ± 0.3 | 5.3 ± 0.9 | 8.0 ± 0.6 | 1.6 ± 0.04 | 17.2 ± 1.1 | 7.0 ± 0.8 | |
| P | NM | 2.0 ± 0.3 | 2.3 ± 0.3 | 4.6 ± 0.3 | 1.17 ± 0.05 | 4.6 ± 0.6 | 3.0 ± 0.5 |
| M | 3.0 ± 0.4 | 4.9 ± 0.4 | 7.3 ± 0.3 | 1.16 ± 0.04 | 14.9 ± 1.3 | 6.7 ± 0.7 | |
| S | NM | 3.3 ± 0.3 | 4.9 ± 0.5 | 7.6 ± 0.6 | 1.53 ± 0.03 | 16.4 ± 1.8 | 4.4 ± 0.3 |
| M | 3.7 ± 0.6 | 5.5 ± 0.6 | 9.5 ± 0.2 | 1.63 ± 0.03 | 19.4 ± 2.0 | 5.1 ± 0.5 | |
| PS | NM | 3.0 ± 0.3 | 4.7 ± 0.2 | 7.3 ± 0.2 | 1.47 ± 0.04 | 14.0 ± 0.5 | 6.7 ± 0.3 |
| M | 3.7 ± 0.9 | 5.0 ± 0.7 | 9.0 ± 0.3 | 1.63 ± 0.02 | 16.6 ± 0.9 | 6.0 ± 0.6 | |
| PF | NM | 2.0 ± 0.3 | 3.8 ± 0.6 | 5.6 ± 0.4 | 1.47 ± 0.02 | 8.8 ± 0.6 | 4.0 ± 0.3 |
| M | 3.0 ± 0.2 | 4.0 ± 0.4 | 6.9 ± 0.3 | 1.53 ± 0.03 | 10.8 ± 0.8 | 4.4 ± 0.2 | |
| LSD (p < 0.05) | 0.98 | 1.5 | 0.8 | 0.2 | 4.1 | 1.4 | |
| Treatment | ** | * | *** | Ns | ** | * | |
| Mycorrhiza | ** | * | * | Ns | ** | * | |
| Treatment × Mycorrhiza | * | * | * | Ns | ** | * | |
| Treatment | Mycorrhizal Status | Phenolic Content (mg g−1 fwt) | Peroxidase (∆A470 min−1 g−1 fwt) | Polyphenol Oxidase (∆A420 min−1 g−1 fwt) | Electrolyte Leakage (%) | Soluble Solids Content (°Brix) |
|---|---|---|---|---|---|---|
| C | NM | 394.6 ± 8.5 | 0.9 ± 0.08 | 1.0 ± 0.04 | 55.3 ± 1.02 | 14.1 ± 0.7 |
| M | 570.2 ± 7.4 | 1.8 ± 0.05 | 1.5 ± 0.03 | 55.9 ± 1.1 | 16.2 ± 0.5 | |
| P | NM | 568.9 ± 5.6 | 1.4 ± 0.03 | 1.3 ± 0.04 | 118.4 ± 1.7 | 8.1 ± 1.0 |
| M | 571.0 ± 6.8 | 2.3 ± 0.09 | 1.9 ± 0.06 | 84.5 ± 1.3 | 15.9 ± 0.6 | |
| S | NM | 500.2 ± 9.2 | 1.7 ± 0.04 | 1.5 ± 0.04 | 56.9 ± 1.2 | 15.0 ± 0.4 |
| M | 604.4 ± 10.1 | 2.0 ± 0.05 | 1.4 ± 0.04 | 55.4 ± 1.4 | 17.3 ± 0.3 | |
| PS | NM | 763.1 ± 12.3 | 2.0 ± 0.03 | 1.6 ± 0.03 | 84.9 ± 1.3 | 14.8 ± 0.4 |
| M | 1135.2 ± 20.4 | 2.3 ± 0.05 | 1.9 ± 0.03 | 75.4 ± 1.1 | 16.4 ± 0.2 | |
| PF | NM | 537.1 ± 6.3 | 1.3 ± 0.09 | 1.3 ± 0.07 | 83.1 ± 1.5 | 12.3 ± 0.8 |
| M | 667.7 ± 8.4 | 1.9 ± 0.04 | 1.8 ± 0.08 | 84.5 ± 1.4 | 12.9 ± 0.6 | |
| LSD (p < 0.05) | 163.1 | 0.2 | 0.2 | 3.5 | 1.7 | |
| Treatment | *** | *** | *** | *** | *** | |
| Mycorrhiza | ** | ** | *** | * | ** | |
| Treatment × Mycorrhiza | * | *** | *** | *** | *** | |
| Treatment | Mycorrhizal Status | Chl. a (mg g−1 fwt) | Chl. b (mg g−1 fwt) | Carotenoids (mg g−1 fwt) | Total Pigments (mg g−1 fwt) |
|---|---|---|---|---|---|
| C | NM | 2.5 ± 0.5 | 0.8 ± 0.06 | 0.4 ± 0.04 | 3.7 ± 0.9 |
| M | 3.0 ± 0.4 | 0.7 ± 0.05 | 0.8 ± 0.05 | 4.6 ± 0.4 | |
| P | NM | 0.6 ± 0.1 | 0.1 ± 0.02 | 0.1 ± 0.01 | 0.9 ± 0.09 |
| M | 2.4 ± 0.6 | 0.5 ± 0.04 | 0.6 ± 0.02 | 3.6 ± 0.5 | |
| S | NM | 2.7 ± 0.3 | 1.0 ± 0.02 | 0.6 ± 0.02 | 4.3 ± 0.3 |
| M | 3.8 ± 0.6 | 1.2 ± 0.07 | 0.7 ± 0.06 | 5.7 ± 0.6 | |
| PS | NM | 2.2 ± 0.4 | 0.4 ± 0.03 | 0.5 ± 0.04 | 3.1 ± 0.4 |
| M | 2.7 ± 0.2 | 1.2 ± 0.05 | 0.3 ± 0.05 | 4.2 ± 0.3 | |
| PF | NM | 2.1 ± 0.5 | 0.5 ± 0.01 | 0.4 ± 0.03 | 2.9 ± 0.3 |
| M | 2.3 ± 0.4 | 0.4 ± 0.02 | 0.6 ± 0.06 | 3.3 ± 0.4 | |
| LSD (p < 0.05) | 0.2 | 0.2 | 0.1 | 0.3 | |
| Treatment | *** | ** | * | *** | |
| Mycorrhiza | *** | ** | *** | *** | |
| Treatment × Mycorrhiza | *** | * | ** | *** | |
| Treatment | Mycorrhizal Status | FC (%) | IC (%) | FA (%) |
|---|---|---|---|---|
| C | NM | 0 | 0 | 0 |
| M | 78.3 ± 1.2 | 35.4 ± 1.0 | 11.6 ± 0.9 | |
| P | NM | 0 | 0 | 0 |
| M | 54.6 ± 0.9 | 17.2 ± 0.7 | 8.4 ± 0.5 | |
| S | NM | 0 | 0 | 0 |
| M | 85.3 ± 1.3 | 44.3 ± 1.1 | 24.4 ± 1.0 | |
| PS | NM | 0 | 0 | 0 |
| M | 80.5 ± 0.8 | 40.7 ± 1.1 | 20.7 ± 0.6 | |
| PF | NM | 0 | 0 | 0 |
| M | 75.4 ± 0.7 | 24.8 ± 0.8 | 10.0 ± 0.5 | |
| LSD (p < 0.05) | 2.4 | 3.3 | 1.5 | |
| Treatment | *** | *** | *** | |
| Mycorrhiza | *** | *** | *** | |
| Treatment × Mycorrhiza | *** | ** | ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Sharkawy, H.H.A.; Rashad, Y.M.; Elazab, N.T. Synergism between Streptomyces viridosporus HH1 and Rhizophagus irregularis Effectively Induces Defense Responses to Fusarium Wilt of Pea and Improves Plant Growth and Yield. J. Fungi 2022, 8, 683. https://doi.org/10.3390/jof8070683
El-Sharkawy HHA, Rashad YM, Elazab NT. Synergism between Streptomyces viridosporus HH1 and Rhizophagus irregularis Effectively Induces Defense Responses to Fusarium Wilt of Pea and Improves Plant Growth and Yield. Journal of Fungi. 2022; 8(7):683. https://doi.org/10.3390/jof8070683
Chicago/Turabian StyleEl-Sharkawy, Hany H. A., Younes M. Rashad, and Nahla T. Elazab. 2022. "Synergism between Streptomyces viridosporus HH1 and Rhizophagus irregularis Effectively Induces Defense Responses to Fusarium Wilt of Pea and Improves Plant Growth and Yield" Journal of Fungi 8, no. 7: 683. https://doi.org/10.3390/jof8070683
APA StyleEl-Sharkawy, H. H. A., Rashad, Y. M., & Elazab, N. T. (2022). Synergism between Streptomyces viridosporus HH1 and Rhizophagus irregularis Effectively Induces Defense Responses to Fusarium Wilt of Pea and Improves Plant Growth and Yield. Journal of Fungi, 8(7), 683. https://doi.org/10.3390/jof8070683