
Molecular Phylogeny and Morphology Reveal Four Novel Species of Corynespora and Kirschsteiniothelia (Dothideomycetes, Ascomycota) from China: A Checklist for Corynespora Reported Worldwide

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
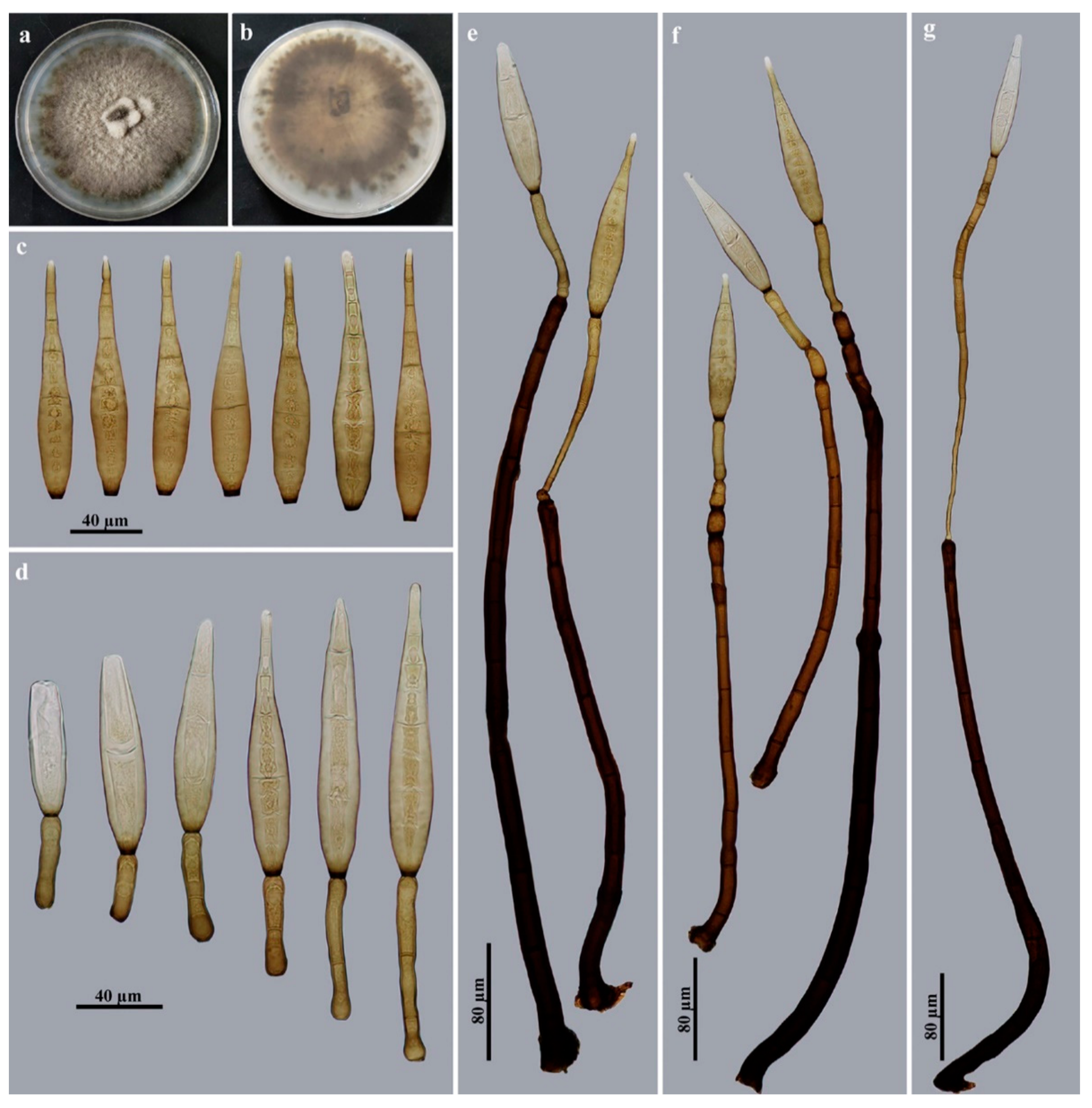
2.1. Sample Collection, Isolation and Morphology
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
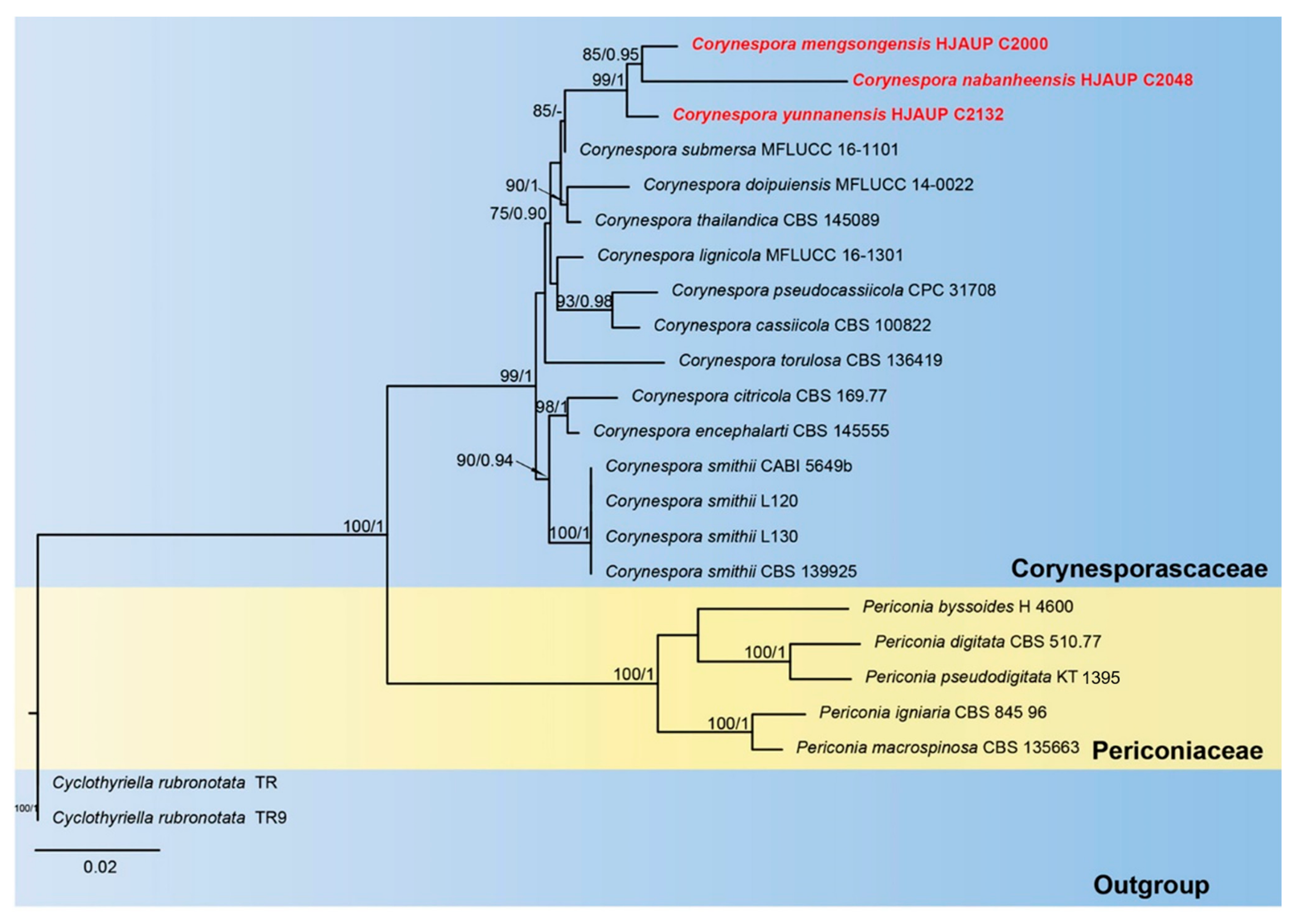
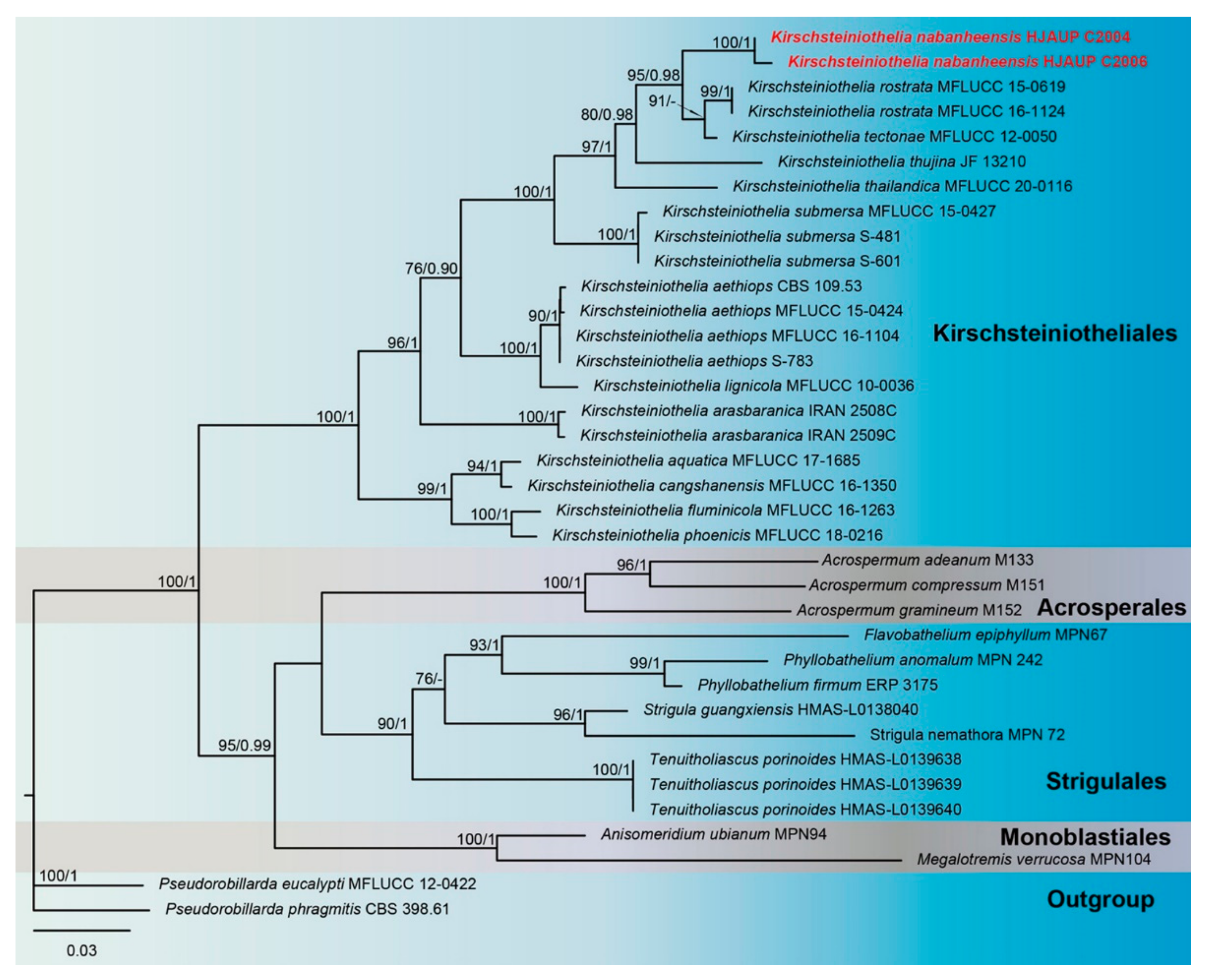
3. Results
3.1. Molecular Phylogeny
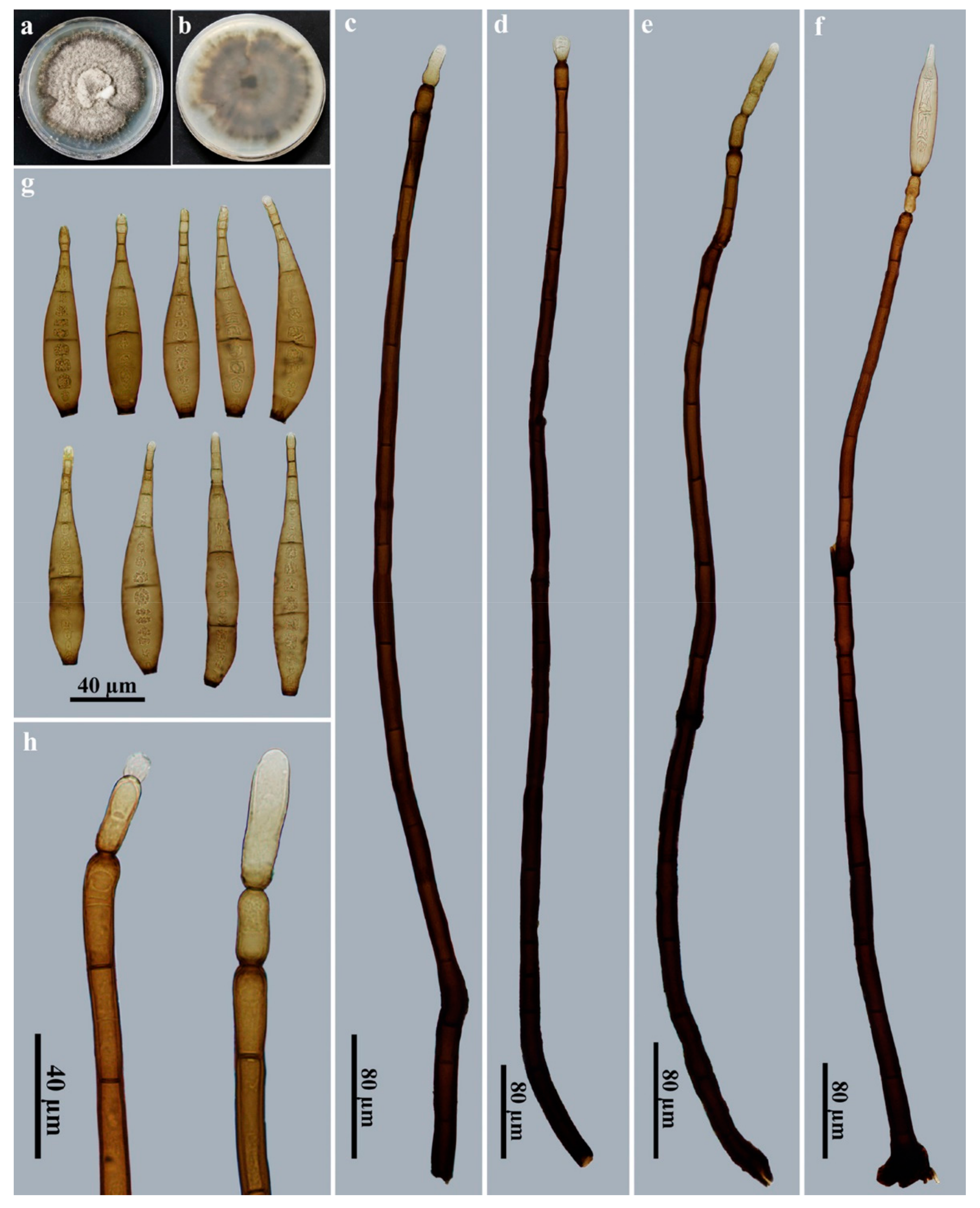
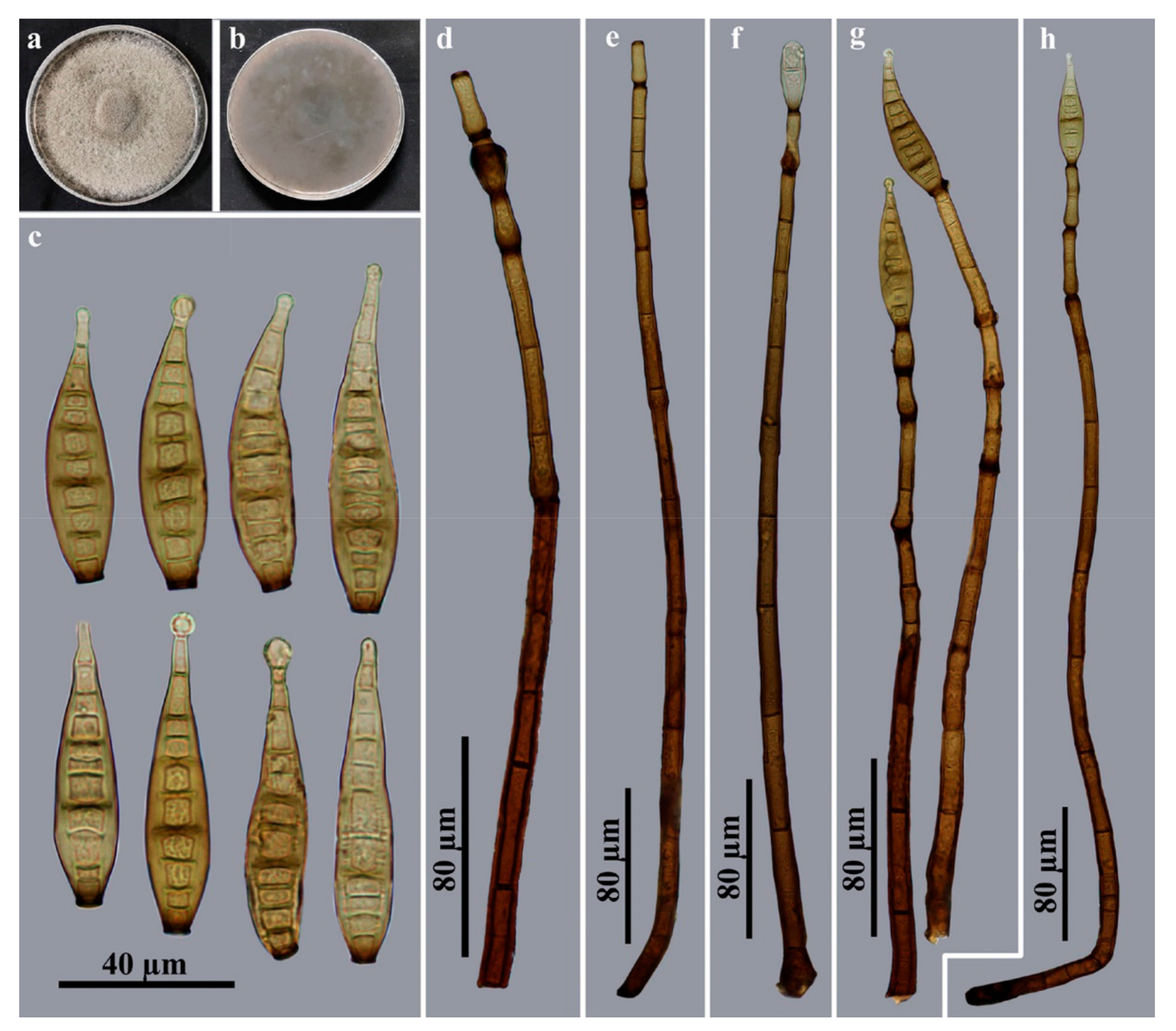
3.2. Taxonomy
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Conida | Host/Locality | References | ||||
|---|---|---|---|---|---|---|---|
| Production | Morphology | Color | Size (µm) | Septation | |||
| Corynespora acaciae | Solitary | Obclavate | Dark brown | 16–30 × 6–8 | 1–5 | On phyllodes of Acacia pycnantha, Australia | [67] |
| C. acalyphae | Solitary | Obclavate, rostrate | Pale brown to brown | 85–120 × 9–11 | 8–16 | On dead branches of Acalypha hamiltoniana, Indonesia | [68] |
| C. achradis | Solitary or catenate | Obclavate, rostrate | Pale olivaceous brown | 60–95 × 6–7 | 5–10 | On leaves of Achras sapota, Brunei | [69] |
| “C. aeria” | Solitary | Obclavate | Subhyaline to olivaceous | Up to 350 × 2–5 | 1–5 | Isolated from air, India | [29] |
| C. albiziicola | Solitary | Obclavate, ellipsoid or clavate | Pale olivaceous yellow | 20–70.1 × 10–18.5 | 1–6 | On leaves of Albizia lebbek, India | [70] |
| C. alstoniae | Solitary or catenate | Cylindrical to obclavate | Subhyaline to light olivaceous | 48–154 × 8–21.5 | 2–15 | On leaves of Alstonia scholaris, Nepal | [71] |
| C. annonacea | Solitary or catenate | Obclavate to obclavate–cylindrical | Subhyaline to olivaceous brown | 25–135 × 10–18 | 1–10 | On living leaves of Annona squamosa, India | [72] |
| C. aquatica | Solitary | Obclavate to cylindrical | Pale brown | 34–46 × 3–4.5 | (1–)2(–3) | On decaying leaves submerged in stream, Mexico | [24] |
| C. arctespora | Solitary or catenate | Cylindrical to obclavate | Brown to pale brown | 13–63 × 4–7 | 2–20 | On twigs of Vaccinium, USA | [73] |
| C. asclepiadacearum | Mostly solitary | Obclavato-cylindric to cylindrical | Pale olivaceous brown | 44–192 × 10–25 | Up to 26 | On leaves of Cryptolepis buchananii, India | [74] |
| C. azadirachtiana | Solitary or catenate | Obclavate | Pale yellow | 32–303.5 × 7–21.5 | 1–20 | On leaves of Azadirachta indica, India | [75] |
| C. barleriicola | Solitary | Obclavate to cylindrical | Olivaceous yellow | 41–246 × 10–18.5 | 3–14 | On leaves of Barleria cristata, India | [75] |
| C. bdellomorpha | Solitary | Obclavate | Mid to dark-reddish brown | 90–138 × 12–17 | 12–19 | On dead stems of Chusquea valdiviensis, Chile | [26] |
| C. beilschmiediae | Solitary | Obclavate | Pale brown to brown | 52–144.5 × 8.5–11 | 7–19 | On dead branches of Beilschmiedia intermedia, China | [76] |
| C. bombacearum | Solitary or catenate | Obclavato-cylindrical to cylindrical | Pale to mid-olivaceous | 26–206 × 8.5–17 | Up to 15 | On leaves of Bombax malabaricum, India | [77] |
| C. bombacina | Solitary or catenate | Obclavate to cylindrical | Light olivaceous | 45–180 × 10–16 | 5–15 | On living leaves of Bombax ceiba, India | [78] |
| C. calicioidea | Solitary | Obclavate | Subhyaline to pale golden brown | 50–170 × 10–15 | 6–21 | On wood, Sri Lanka | [79] |
| C. carrisae | Solitary | Obclavate to cylindrical | Olivaceous to very light brown | 75–242 × 6–14 | 4–17 | On leaves of Carissa spinarum, India | [80] |
| C. caryotae | Solitary | Obclavate to elongate | Pinkish brown | 45–120 × 6–10 | Up to 18 | On dead rachis of Caryota mitis, Singapore | [81] |
| C. cassiae | Solitary | Obclavate | Pale brown to olivaceous brown | 107.5–214 × 11–14 | 10–21 | On dead branches of Cassia surattensis, China | [76] |
| C. cassiicola | Solitary or catenate | Obclavate to cylindrical | Subhyaline to pale olivaceous brown | 40–220 × 9–22 | 4–20 | On leaves of Cassia, Cuba | [5] |
| C. catenulata | Solitary or catenate | Obclavate to obclavato-cylindrical | Dark olivaceous yellow to pale olivaceous brown | 27.5–225.5 × 11–19 | 1–24 | On leaves of Clerodendrum indicum, India | [75] |
| C. catharanthicola | Solitary or catenate | Cylindrical | Brown | 140–310 × 5.5–11 | 4–25 | On leaves of Catharanthus roseus, China | [82] |
| C. celastri | Solitary | Obclavate to obclavato-cylindrical | Olivaceous to very light brown | 55–120 × 8–15 | 7–17 | On living leaves of Celastrus paniculatus, India | [83] |
| C. chinensis | Catenate | Obclavate | Pale brown | 31–61 × 5–7.5 | 1–5 | On dead branches of Angiospermae, China | [14] |
| C. citricola | Solitary or catenate | Cylindrical to obclavate | Subhyaline | 48–150 × 4.5–8 | 4–18 | On leaves of Citrus aurantiifolia, Australia | [79] |
| C. clerodendrigena | Solitary or catenate | Obclavate to cylindrical | Light olivaceous | 60–220 × 16–22 | 3–13 | On leaves of Clerodendrum viscosum, India | [84] |
| C. colebrookiana | Solitary or catenate | Obclavate, rarely cylindrical | Pale yellow | 45–330 × 6–22 | 4–16 | On leaves of Colebrookea oppositifolia, India | [75] |
| C. combreli | Solitary | Obclavate, rostrate | Pale olivaceous brown to olivaceous brown | 40–122 × 8–11 | 4–10 | On dead branches of Combretum zeyheri, Zambia | [85] |
| C. cubensis | Solitary or catenate | Cylindrical to obclavate | Pale brown to dark rusty brown | 40–80 × 8–12 | 6–15 | On dead petiole of Coccothrinax, Cuba | [86] |
| C. cucurbiticola | Solitary or catenate | Obclavato-cylindrical | Subhyaline to pale olivaceous | 38.5–230 × 6.5–20 | 6–23 | On leaves of Coccinia grandis, Nepal | [87] |
| C. curvispora | Solitary or catenate | Narrow obclavate | Straw-colored to mid-brown | 40–250 × 10–12 | 5–10 | On fallen herbaceous stems, USA | [88] |
| C. doipuiensis | Solitary | Obclavate to cylindrical | Subhyaline to moderately brown | 136–165 × 5–25.5 | – | On dead herbaceous branches, Thailand | [12] |
| C. donacis | Solitary | Obclavate | Olivaceous brown | 45–70 × 8–12 | 10–14 | On dead branches of Donax, China | [89] |
| C. elaeidicola | Solitary or catenate | Cylindrical to obclavate | Subhyaline or pale olivaceous brown | 43–65 × 4–7 | 3–7 | On dead leaves of Elaeis guineensis, Malaysia | [27] |
| C. encephalarti | Solitary | Obclavate | Medium olivaceous brown | 100–150 × 11–15 | 1–12 | On leaves of Encephalartos, South Africa | [90] |
| C. eranthemi | Solitary | Obclavate | Brown to pale olivaceous brown | 65–176 × 11–14 | 5–25 | On leaves of Eranthemum wattii, Singapore | [32] |
| C. erythropsidis | Solitary | Ellipsoid, doliiform to broad clavate | Pale brown to olivaceous brown | 25–31 × 9–12 | 4 | On dead branches of Erythropsis colorata, China | [91] |
| C. euphorbiacearum | Solitary or catenate | Obclavate | Subhyaline to light olivaceous brown | 59–235 × 11–22.5 | 5–18 | On leaves of Manihot esculenta, India | [71] |
| C. euryae | Solitary | Obclavate | Pale brown to brown | 36–67 × 6–9 | 5–9 | On dead branches of Eurya inaequalis, China | [92] |
| C. fici-altissimae | Solitary | Obclavate, rostrate | Dark brown | 55–85 × 9–12 | 11–18 | On dead branches of Ficus altissima, China | [89] |
| C. fici-benjaminae | Solitary | Obclavate | Pale olivaceous brown | 51.5–71 × 8–11 | 5–10 | On dead branches of Ficus benjamina, China | [76] |
| C. ficigena | Solitary | Obclavate to cylindrical | Light olivaceous brown | 90–165 × 9–20 | 7–13 | On leaves of Ficus religiosa, India | [93] |
| C. flagellata | Solitary | Obclavate, rostrate, smooth or verrucose | Dark brown | 50–100 × 9–11 | 5–10 | On wood of Citrus, Ghana | [94] |
| C. fujianensis | Solitary | Obclavate | Brown | 31–90 × 6.5–10 | 4–10 | On dead branches of Myrioneuron faberi, China | [95] |
| C. gigaspora | Solitary | Obclavate, rostrate | Pale to dark golden brown | 100–270 × 19–28 | 9–52 | On dead wood, Sri Lanka | [79] |
| C. gorakhpurensis | Solitary | Obclavate to ellipsoid | Pale olivaceous yellow | 21–157 × 13–20 | 3–13 | On leaves of Erythrina indica, India | [70] |
| C. gracilis | Solitary | Cylindric to obclavate | Olivaceous | 92–138 × 5–7 | 10–22 | On dead branches of Piper betle, Indonesia | [68] |
| C. gymnocladi | Solitary | Obclavate | Brown to dark brown | 15–40 × 7–10.5 | 2–6 | On dead branches of Gymnocladus chinensis, China | [92] |
| C. hamata | Solitary | Obclavate, hamatate at apex | Pale olivaceous brown | 158–198 × 9–11 | 14–19 | On dead wood, Indonesia | [68] |
| C. hansfordii | Solitary | Obclavate, rostrate | Straw-colored to brown | 70–100 × 9–13 | 7–10 | On dead wood, Uganda | [27] |
| C. hemigraphidis | Solitary | Obclavate | Pale olivaceous brown | 72–218 × 12–15 | 5–25 | On leaves of Hemigraphis alternat, Singapore | [32] |
| C. heterospora | Solitary | Cylindrical to obclavate | Pale olivaceous brown to olivaceous brown | 75–170 × 6–20 | 6–12 | On leaves of Manihot utilissima, Malaysia | [96] |
| C. holopoteleae | Solitary or catenate | Obclavato-cylindrical to cylindrical | Mid olivaceous | 23–234 × 3.6–19.5 | 0–17 | On leaves of Holoptelea integrifolia, India | [77] |
| C. holopteleicola | Solitary | Obclavate to obclavato-cylindrical | Olivaceous brown | 33–148 × 5–20 | 0–11 | On living leaves of Holoptelea integrifolia, India | [72] |
| C. homaliicola | Solitary | Obclavate, cylindrical | Subhyaline to straw-colored | 110–220 × 11–22 | 13–28 | On dead branches of Homalium aylmeri, Sierra Leone | [79] |
| “C. ipomoeae” | Solitary or catenate | Obclavate to cylindrical | Subhyaline to pale olivaceous | 40–380 × 5–15 | 2–35 | On leaves of Ipomoea obscura, India | [30] |
| C. jasminicola | Solitary | Obclavate | Pale olivaceous | 39.5–176 × 10–21 | 2–18 | On leaves of Jasminum arborescens, Nepal | [87] |
| C. kamatii | Solitary | Obclavate | Straw-colored | 60–70 × 10–13 | 7–12 | On dead twigs of Vitis, India | [69] |
| C. kenyensis | Solitary | Obclavate to obpyriform, rostrate | Subhyaline to pale brown | 60–125 × 16–25 | 8–15 | On dead stems of Sericostachys scandens, Kenya | [8] |
| C. keskaliicola | Solitary or catenate | Obclavato-cylindrical to cylindrical | Mid olivaceous | 64–164 × 16–28 | Up to 17 | On leaves of Hemidesmus indicus, India | [74] |
| C. laevistipitata | Solitary | Broadly ellipsoid | Red–brown | 17.5–24 × 7–8 | (0–)1–2 (–3) | On Pertusaria ophthalmiza (lichen), USA | [97] |
| C. lanneicola | Solitary | Obclavate | Straw-colored to brown | 40–58 × 10–15 | 4–5 | On dead branches of Lannea afzelii, Sierra Leone | [79] |
| C. lasianthi | Solitary | Obclavate, sometimes rostrate | Pale brown to dark brown | 50–103.5 × 8.5–10 | 4–8 | On dead branches of Lasianthus chinensis, China | [76] |
| C. leptoderridicola | Solitary | Obclavate, rostrate | Subhyaline to straw-colored | 70–120 × 14–17 | 6–16 | On dead branches of Leptoderris fasciculata, Sierra Leone | [79] |
| C. leucaenae | Solitary | Obclavate, obovoid or ellipsoid | Pale yellow | 16–298 × 10–19 | 1–28 | On leaves of Leucaena leucocephala, India | [70] |
| C. lignicola | Solitary or catenate | Cylindrical | Subhyaline to pale brown | 110–156 × 7–9 | – | On submerged decaying wood, China | [12] |
| C. ligustri | Solitary or catenate | Obclavate to cylindrical | Straw-colored to brown | 25–225 × 7.5–30 | 4–20 | On leaves of Ligustrum lucidum, China | [98] |
| C. litseae | Solitary | Obclavate | Pale brown to olivaceous brown | 105–235 × 10–12 | 14–34 | On dead branches of Litsea elongata, China | [99] |
| C. longispora | Solitary | Cylindrical | Subhyaline to pale brown | 120–330 × 5.5–8 | 11–24 | On dead herbaceous stems, India | [100] |
| C. maculiformis | Solitary or catenate | Cylindrical to obclavate | Subhyaline to pale olivaceous brown | 20–86 × 5–10 | 2–8 | On rotten wood, Czech Republic | [101] |
| “C. masseeanum” | Solitary | Elongate to obclavate | Pale olivaceous | 80–120 × 18–20 | 7–11 | On branches of Helicteres Isora, India | [31] |
| C. matuszakii | Solitary or catenate | Cylindrical to obclavate | Pale brown to straw-colored or mid brown | 56–260 × 10–12.5 | 2–10 | On herbaceous stems of Compositae, USA | [102] |
| C. merremiae | Solitary or catenate | Cylindrical to obclavate | Pale olivaceous brown to pale brown | 37–150 × 6–12.5 | 4–22 | On leaves of Merremia hirta, China | [98] |
| C. merrilliopanacis | Solitary | Obclavate, rostrate | Straw-colored to brown | 130–260 × 17–21 | 12–25 | On dead branches of Merrilliopanax listeri, China | [61] |
| C. micheliae | Solitary | Obclavate, rostrate | Subyhaline to brown | 333–360 × 15–19 | 12–28 | On dead branches of Michelia champaca, China | [61] |
| C. millettiae | Solitary or catenate | Obclavate, smooth | Olivaceous brown to mid brown | 30–182 × 7.5–14 | 2–15 | On leaves of Millettia, China | [98] |
| C. moracearum | Solitary | Obclavate to cylindrical | Light olivaceous brown | 27–163 × 12–20 | 5–16 | On living leaves of Ficus hispida, India | [103] |
| C. morindae-tinctoriae | Solitary | Obclavate | Pale olivaceous | 44–127 × 15–26.5 | 6–15 | On leaves of Morinda tinctoria, India | [104] |
| C. myrioneuronis | Solitary | Obclavate | Pale brown to brown | 30–46 × 6.5–8 | 3–4 | On dead branch of Myrioneuron faberi, China | [92] |
| C. mengsongensis | Solitary | Obclavate to cylindrical, rostrate | Brown to golden brown | 96–146 × 16.5–20.5 | 13–18 | On dead branches, China | This study |
| C. nana | Solitary | Obclavate | Subhyaline to pale olivaceous brown | 49.5–110 × 9–18.5 | 4–14 | On leaves of Lantana indica, India | [104] |
| C. nabanheensis | Solitary | Obclavate to cylindrical, expanded to a rounded shape at the apex | Pale brown to brown | 56–84 × 12–14 | 9–13 | On dead branches, China | This study |
| C. occidentalis | Solitary | Ovoid to ellipsoidal | Subyhaline to pale brown | 30–54 × 15–19 | 3–6 | On leaves of Cordia collococca, Cuba | [105] |
| C. palmicola | Solitary | Obclavate to subcylindrical | Pale brown | 40–70 × 6–9 | 5–7 | On leaves of Cocos australis, Paraguay | [106] |
| C. parapyrenariae | Solitary | Obclavate | Pale brown to brown | 70–100 × 11–14 | 5–9 | On dead branches of Parapyrenaria multisepala, China | [99] |
| C. parvispora | Solitary | Ovoid | Brown | 13–15 × 4.5–7.5 | 1–2 | On dead twigs of Gynotroches axillaris, Singapore | [81] |
| C. pedaliacearum | Solitary or catenate | Obclavato-cylindrical to slightly acicular | Pale olivaceous | 16–163 × 3.2–6 | 3–28 | On leaves of Sesamum indicum, India | [107] |
| C. peristrophicola | Solitary | Obclavate to obclavato-cylindrical | Olivaceous to very light brown | 60–135 × 5–16 | 5–12 | On leaves of Peristrophe bicalyculata, India | [80] |
| C. phylloshureae | Solitary | Obclavate | Brown | 30–50 × 8–10 | 6–10 | On dead branches of Phyllostachys sulphurea, China | [89] |
| C. pogostemonicola | Solitary | Obclavate to obclavato-cylindrical | Olivaceous to olivaceous brown | 77–288 × 8–14 | 5–24 | On leaves of Pogostemon plectrantoides, India | [108] |
| C. polyphragmia | Solitary or catenate | Obclavate | Pale to mid golden brown | 110–280 × 14–17 | 10–25 | On decorticated branches of Camellia japonica, Japan | [109] |
| C. pongamiicola | Solitary | Obclavate, ellipsoidal, clavate or club-shaped | Light olivaceous yellow | 18–65.2 × 8–16.5 | 1–6 | On living leaves of Pongamia pinnata, India | [110] |
| C. premnigena | Solitary or catenate | Obclavate to obclavato-cylindrical | Subhyaline to pale yellow | 52–265 × 10–15 | 1–19 | On leaves of Premna mucronata, India | [75] |
| C. proliferata | Solitary or catenate | Obclavate, rostrate | Pale brown to brown | 30–300 × 9–12 | 3–17 | On wood of Fagus sylvatica, the Netherlands | [111] |
| C. pruni | Solitary or catenate | Obclavate | Olivaceous brown or brown | 50–130 × 10–16 | 4–9 | On bark of Prunus serotina, USA | [27] |
| C. pseudocassiicola | Solitary | Subcylindrical to obclavate | Medium brown | 95–160 × 9–10 | (4–)8–12(–17) | On leaves of Byrsonima, Colombia | [112] |
| C. queenslandica | Solitary | Obclavate | Pale brown | 72–114 × 8–10 | 6–9 | On phyllodes of Acacia leiocalyx, Australia | [15] |
| C. rhapidis-humilis | Solitary | Obclavate, rostrate | Pale brown to olivaceous brown | 90–130 × 6–8 | 12–16 | On dead branches of Rhapis humilis, China | [94] |
| C. rhododendri | Solitary | Obclavate to long rostrate | Pale brown to olivaceous brown | 180–400 × 7.5–11 | 19–36 | On dead branches of Rhododendron hainanense, China | [113] |
| C. ripogoni | Solitary | Obclavate | Brown | 60–160 × 10–13.5 | 7–15 | On dead stems of Ripogonum scandens, New Zealand | [9] |
| C. rosacearum | Solitary or catenate | Obclavate to obclavato-cylindrical | Subhyaline to pale olivaceous brown | 26.5–269 × 9–18.5 | 1–18 | On leaves of Eriobotrya japonica, India | [104] |
| “C. ruelliae” | Solitary | Obclavate | Brown to pale olivaceous brown | 60–150 × 12–15 | 5–16 | On leaves of Ruellia macrophylla and Ruellia dipteracanthus, Singapore | [32] |
| C. sacchari | Solitary | Obclavate, rostrate, verrucose or smooth | Pale brown to olivaceous brown | 80–120 × 8–9 | 10–14 | On dead branches of Saccharum sinense, China | [114] |
| C. salasiae | Solitary | Ellipsoidal, doliiform | Brown | 17–20 × 8–12 | 0–2 | On dead stems of grass, Cuba | [115] |
| C. schleichericola | Solitary or catenate | Obclavate | Pale olivaceous brown | 22.5–66 × 3.8–8.5 | 1–12 | On leaves of Schleichera trijuga, India | [107] |
| C. scolopiae | Solitary | Obclavate | Pale brown to brown | 90–150 × 10–13 | 8–11 | On dead branches of Scolopia chinensis, China | [116] |
| C. sed-acaciae | Solitary | Obclavate | Pale brown to olivaceous brown | 40–70 × 11–13.5 | 8–12 | On dead branches of Acacia confusa, China | [113] |
| C. sidae | Solitary | Obclavate to obclavato-cylindrical | Olivaceous brown to very light brown | 25–220 × 7–17 | 2–24 | On leaves of Sida acuta, India | [117] |
| C. sinensis | Catenate | Obclavate or fusiform, ellipsoid | Brown | 21–42 × 8–9.5 | 3(–4) | On dead branches, China | [13] |
| C. siwalika | Solitary | Obclavate, rostrate | Pale straw-colored to golden brown | 88–140 × 15–20 | 9–19 | On branches of Helicteres isora, India | [109] |
| C. smithii | Solitary or catenate | Cylindrical | Subhyaline to golden brown | 70–410 × 12–19 | 7–45 | On bark of Ilex, UK | [79] |
| C. solani | Solitary or catenate | Obclavate to cylindrical | Olivaceous yellow | 80.6–276 × 8–10 | 1–17 | On leaves of Solanum indicum, India | [118] |
| C. subcylindrica | Catenate | Broadly ellipsoid, subcylindrical | Pale brown | 18–60(–90) × 5–13 | 0–3(–6) | On leaves of Lippia sidoides, Brazil | [63] |
| C. submersa | Solitary or catenate | Obclavate, rostrate | Subhyaline to golden brown | 100–150 × 16–24 | 9–13 | On submerged decaying wood, China | [12] |
| C. supkharii | Solitary | Obclavate | Pale olivaceous brown | 22.5–142.5 × 10–17.5 | 2–11 | On leaves of Phyllanthus parvifolius, India | [119] |
| C. tanaceti | Solitary | Obclavate, smooth or verruculose | Pale brown to olivaceous brown | 60–104 × 12–16 | 7–12 | On dead branches of Tanacetum vulgare, China | [116] |
| C. tectonae | Solitary | Obclavate, rostrate, verrucose or smooth | Pale brown to olivaceous brown | 110–160 × 10–12 | 12–18 | On dead branches of Tectona grandis, China | [114] |
| C. thailandica | Mostly solitary | Obclavate | Brown | 80–110 × 10–12 | 4–8 | On wood, Thailand | [120] |
| C. thorii | Catenate | Subcylindrical, broadly ellipsoid to almost obovoid | Pale brown to medium olivaceous brown | 20–30 × 5–7 | (0–)1(–3) | On thallus, apothecia of Lecanora, Japan | [121] |
| C. titarpaniensis | Solitary | Obclavate to cylindrical | Olivaceous brown to light brown | 50–340 × 5–20 | 5–35 | On living leaves of Lepidagathis, India | [122] |
| C. tomenticola | Solitary | Cylindrical | Olivaceous brown to brown | 50–230 × 10.5–20.5 | 3–6 | On living leaves of Terminalia tomentosa, India | [110] |
| C. toonae | Solitary | Obclavate, rostrate | Pale brown to dark brown | 65–144 × 7–9 | 4–14 | On dead branches of Toona sinensis, China | [114] |
| C. torulosa | Solitary | Clavate | Dark olivaceous brown | 35–60 × 13–20 | 3–5 | On dead leaves of Musa sapientum Brazil | [123] |
| C. tremae | Solitary | Obclavate to obclavato-cylindrical | Light brown to brown | 50–160 × 4–12 | 5–20 | On dead petiole of Trema orientalis, India | [124] |
| C. trematicola | Solitary | Obclavate to ellipsoid | Pale olivaceous brown | 104–296 × 11–16 | 1–12 | On leaves of Trema orientalis, India | [118] |
| C. trichiliae | Solitary | Obclavate, rostrate | Subhyaline to straw-colored | 53–74 × 9–11 | 4–6 | On branches of Trichilia heudelotii, Sierra Leone | [27] |
| C. trichoides | Solitary | Obclavato-cylindrical or obclavate | Pale olivaceous brown | 29–170 × 10–15 | 3–14 | On leaves of Triumfetta rhomboidea, Nepal | [87] |
| C. ulmacearum | Solitary | Obclavate | Subhyaline to pale olivaceous brown | 15–106 × 3.5–10 | 2–16 | On leaves of Trema orientalis, India | [107] |
| C. vismiae | Solitary or catenate | Obclavate, rostrate | Pale olivaceous brown or straw-colored | 55–107 × 6–9 | 3–5 | On leaves of Vismia guineensis, Sierra Leone | [85] |
| C. viticis | Solitary or catenate | Cylindrical | Pale brown | 80–383 × 6–9 | – | On leaves of Vitex rotundifolia, China | [98] |
| C. viticola | Solitary or catenate | Obclavate, cylindrical to obovoid | Pale olivaceous brown | 34–170 × 7–17.5 | 1–14 | On leaves of Cayratia carnosa, India | [118] |
| C. woodfordiana | Solitary or catenate | Obclavate, rostrate | Light olivaceous brown | 40–170 × 9.5–17 | 4–14 | On leaves of Woodfordia fruticosa, India | [71] |
| C. yerbae | Solitary or catenate | Obclavate | Subhyaline to pale golden brown | 72–170 × 16–18 | 8–19 | On dead branches of Ilex paraguayensis, Argentina | [26] |
| C. yunnanensis | Solitary | Obclavato-cylindrical, rostrate | Brown to golden brown | 80–128 × 16–19 | 3–16 | On dead branches, China | This study |
| C. ziziphae | Solitary | Obclavato-cylindrical, cylindrical or clavate | Mid olivaceous brown to straw-colored | 33–215 × 10–27 | Up to 15 | On leaves of Ziziphus giraldii, India | [77] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seifert, K.; Morgan-Jones, G.; Gams, W.; Kendrick, B. The genera of hyphomycetes. CBS Biodivers. Ser. 2011, 9, 1–997. [Google Scholar]
- Index Fungorum. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 6 January 2023).
- Celio, G.J.; Padamsee, M.; Dentinger, B.T.M.; Bauer, R.; McLaughlin, D.J. Assembling the Fungal Tree of Life: Constructing the Structural and Biochemical Database. Mycologia 2006, 98, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Güssow, H.T. Notes on a disease of cucumbers. II. J. Royal Agric. Soc. Engl. 1905, 65, 271–272. [Google Scholar]
- Wei, C.T. Notes on Corynespora. Mycol. Pap. 1950, 34, 1–10. [Google Scholar]
- Ellis, M.B. Dematiaceous Hyphomycetes; Kew, Commonwealth Mycological Institute: Surrey, UK, 1971. [Google Scholar]
- Species Fungorum. 2023. Available online: http://www.speciesfungorum.org/Names/Names.asp (accessed on 6 January 2023).
- Siboe, G.M.; Kirk, P.M.; Cannon, P.F. New dematiaceous hyphomycetes from Kenyan rare plants. Mycotaxon 1999, 73, 283–302. [Google Scholar]
- McKenzie, E.H.C. Three new phragmosporous hyphomycetes on Ripogonum from an ‘ecological island’ in New Zealand. Mycotaxon 2010, 111, 183–196. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Verkley, G.J.M.; Shin, H.D.; Barreto, R.W.; Alfenas, A.C.; Swart, W.J.; Groenewald, J.Z.; Crous, P.W. Sizing up Septoria. Stud. Mycol. 2013, 75, 307–390. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Wingfield, M.J.; Schumacher, R.K.; Summerell, B.A.; Giraldo, A.; Gené, J.; Guarro, J.; Wanasinghe, D.N.; Hyde, K.D.; Camporesi, E.; et al. Fungal Planet description sheets: 281–319. Persoonia 2014, 33, 212–289. [Google Scholar] [CrossRef]
- Hyde, K.D.; de Silva, N.I.; Jeewon, R.; Bhat, D.J.; Phookamsak, R.; Doilom, M.; Boonmee, S.; Jayawardena, R.S.; Maharachchikumbura, S.S.N.; Senanayake, I.C.; et al. AJOM new records and collections of fungi: 1–100. Asian J. Mycol. 2020, 3, 22–294. [Google Scholar] [CrossRef]
- Xu, Z.H.; Kuang, W.G.; Qiu, L.; Zhang, X.G.; Castañeda-Ruíz, R.F.; Ma, J. Corynespora sinensis sp. nov. from Jiangxi, China. Mycotaxon 2020, 135, 803–809. [Google Scholar] [CrossRef]
- Liu, J.W.; Zhang, X.G.; Castañeda-Ruíz, R.F.; Ma, J. Corynespora chinensis sp. nov. from Hainan, China. Mycotaxon 2022, 137, 203–207. [Google Scholar] [CrossRef]
- Sutton, B.C.; Pascoe, I.G. Some dematiaceous hyphomycetes from branches and phyllodes on Acacia in Australia. Aust. Syst. Bot. 1988, 1, 127–138. [Google Scholar] [CrossRef]
- Castañeda-Ruíz, R.F. Deuteromycotina de Cuba. Hyphomycetes III; Instituto de Investigaciones Fundamentales en Agricultura Tropical “Alejandro de Humboldt”: Habana, Cuba, 1985; pp. 1–42. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. II. Mycol. Pap. 1961, 79, 1–23. [Google Scholar]
- Deighton, F.C. Observations on Phaeoisariopsis. Mycol. Res. 1990, 94, 1096–1102. [Google Scholar] [CrossRef]
- Sutton, B.C. Mitosporic fungi from Malawi. Mycol. Pap. 1993, 167, 1–93. [Google Scholar]
- Dyko, B.J.; Sutton, B.C. New and interesting dematiaceous hyphomycetes from Florida. Mycotaxon 1979, 8, 119–124. [Google Scholar]
- Castañeda-Ruíz, R.F.; Zhang, X.G.; Li, D.W.; Gusmão, L.F.P.; Pérez-Martínez, S.; Sosa, D. Notes on Vamsapriya and V. camagueyensis comb. nov. Mycotaxon 2017, 132, 553–557. [Google Scholar] [CrossRef]
- Delgado-Rodríguez, G.; Mena-Portales, J.; Calduch, M.; Decock, C. Hyphomycetes (Hongos Mitospóricos) del area protegida Mil Cumbres, Cuba Occidental. Cryptog. Mycol. 2002, 23, 277–293. [Google Scholar]
- Heredia, G.; Li, D.W.; Wendt, L.; Reblová, M.; Arias, R.M.; Gamboa-Angulo, M.; Štěpánek, V.; Stadler, M.; Castañeda-Ruíz, R.F. Natonodosa speciosa gen. et sp. nov. and rediscovery of Poroisariopsis inornata: Neotropical anamorphic fungi in Xylariales. Mycol. Prog. 2020, 19, 15–30. [Google Scholar] [CrossRef]
- Castañeda-Ruíz, R.F.; Heredia, G.P.; Arias, R.M.; Saikawa, M.; Minter, D.W.; Stadler, M.; Guarro, J.; Decock, C. Two new hyphomycetes from rainforests of Mexico, and Briansuttonia, a new genus to accommodate Corynespora alternarioides. Mycotaxon 2004, 89, 297–305. [Google Scholar]
- Castañeda-Ruíz, R.F.; Kendrick, B. Conidial fungi from Cuba: II. Univ. Waterloo. Biol. Ser. 1990, 33, 1–61. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. IV. Mycol. Pap. 1963, 87, 1–42. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. I. Mycol. Pap. 1960, 76, 1–36. [Google Scholar]
- Voglmayr, H.; Jaklitsch, W.M. Corynespora, Exosporium and Helminthosporium revisited—New species and generic reclassification. Stud. Mycol. 2017, 87, 43–76. [Google Scholar] [CrossRef]
- Swapana, S.; Nair, N.N. Corynespora aeria: A new species recorded amongst aero mycoflora of Kerala, India. World J. Pharm. Pharm. Sci. 2015, 4, 487–492. [Google Scholar]
- Verma, N.K.; Surywanshi, J.S.; Rai, A.N. Corynespora ipomoeae, a novel taxon of dematiaceous hyphomycetes. J. Mycol. Pl. Pathol. 2014, 44, 466–469. [Google Scholar]
- Kirk, P.M. Nomenclatural novelties. Index Fungorum 2014, 120, 1. [Google Scholar]
- Yen, J.M.; Lim, G. Étude sur les champignons parasites du Sud-Est asiatique. 39. Les Corynespora de Malaisie. Cryptog. Mycol. 1980, 1, 83–90. [Google Scholar]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.H.; Li, D.Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress, Shenzhen, China, July 2017; Regnum Vegetabile 159; Koeltz Botanical Books: Glashütten, Germany, 2018. [Google Scholar] [CrossRef]
- Sivanesan, A. Corynesporasca caryotae gen. et sp. nov. with a Corynespora anamorph, and the family Corynesporascaceae. Mycol. Res. 1996, 100, 783–788. [Google Scholar] [CrossRef]
- Rossman, A.Y.; Crous, P.W.; Hyde, K.D.; Hawksworth, D.L.; Aptroot, A.; Bezerra, J.L.; Bhat, J.D.; Boehm, E.; Braun, U.; Boonmee, S.; et al. Recommended names for pleomorphic genera in Dothideomycetes. IMA Fungus 2015, 6, 507–523. [Google Scholar] [CrossRef]
- Hawksworth, D.L. Kirschsteiniothelia, a new genus for the Microthelia incrustans-group (Dothideales). Bot. J. Linn. Soc. 1985, 91, 181–202. [Google Scholar] [CrossRef]
- Boonmee, S.; Ko, T.W.K.; Chukeatirote, E.; Hyde, K.D.; Chen, H.; Cai, L.; McKenzie, E.H.C.; Jones, E.B.G.; Kodsueb, R.; Hassan, B.A. Two new Kirschsteiniothelia species with Dendryphiopsis anamorphs cluster in Kirschsteiniotheliaceae fam. nov. Mycologia 2012, 104, 698–714. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.R.; Jayawardena, R.S.; Hyde, K.D.; Wang, Y. Kirschsteiniothelia thailandica sp. nov. (Kirschsteiniotheliaceae) from Thailand. Phytotaxa 2021, 490, 172–182. [Google Scholar] [CrossRef]
- Schoch, C.; Crous, P.W.; Groenewald, J.Z.; Boehm, E.; Burgess, T.I.; De Gruyter, J.; De Hoog, G.S.; Dixon, L.; Grube, M.; Gueidan, C. A class-wide phylogenetic assessment of Dothideomycetes. Stud. Mycol. 2009, 64, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wijayawardene, N.N.; Crous, P.W.; Kirk, P.M.; Hawksworth, D.L.; Boonmee, S.; Braun, U.; Dai, D.Q.; D’souza, M.J.; Diederich, P.; Dissanayake, A.; et al. Naming and outline of Dothideomycetes–2014 including proposals for the protection or suppression of generic names. Fungal Divers. 2014, 69, 1–55. [Google Scholar] [CrossRef]
- Bao, D.F.; Luo, Z.L.; Liu, J.K.; Bhat, D.J.; Sarunya, N.; Li, W.L.; Su, H.Y.; Hyde, K.D. Lignicolous freshwater fungi in China III: Three new species and a new record of Kirschsteiniothelia from northwestern Yunnan Province. Mycosphere 2018, 9, 755–768. [Google Scholar] [CrossRef]
- Su, H.Y.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Ariyawansa, H.A.; Luo, Z.L.; Promputtha, I.; Tian, Q.; Lin, C.G.; Shang, Q.J.; Zhao, Y.C.; et al. The families Distoseptisporaceae fam. nov.; Kirschsteiniotheliaceae, Sporormiaceae and Torulaceae, with new species from freshwater in Yunnan Province, China. Fungal Divers. 2016, 80, 375–409. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Chethana, K.W.T.; Clericuzio, M.; Dayarathne, M.C.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.Q.; et al. Fungal diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Li, G.J.; Hyde, K.D.; Zhao, R.L.; Hongsanan, S.; Abdel-Aziz, F.A.; Abdel-Wahab, M.A.; Alvarado, P.; Alves-Silva, G.; Ammirati, J.F.; Ariyawansa, H.A.; et al. Fungal diversity notes 253–366: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 78, 1–237. [Google Scholar] [CrossRef]
- Barr, M.E. Prodromus to Class Loculoascomycetes; Lubrecht & Cramer, Limited: Amherst, MA, USA, 1987; pp. 1–168. [Google Scholar]
- Barr, M.E. Notes on the Pleomassariaceae. Mycotaxon 1993, 49, 129–142. [Google Scholar]
- Schoch, C.L.; Shoemaker, R.A.; Seifert, K.A.; Hambleton, S.; Spatafora, J.W.; Crous, P.W. A multigene phylogeny of the Dothideomycetes using four nuclear loci. Mycologia 2006, 98, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Suetrong, S.; Schoch, C.L.; Spatafora, J.W.; Kohlmeyer, J.; Volkmann-Kohlmeyer, B.; Sakayaroj, J.; Phongpaichit, S.; Tanaka, K.; Hirayama, K.; Jones, E.B.G. Molecular systematics of the marine Dothideomycetes. Stud. Mycol. 2009, 64, 155–173. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Restrepo, M.; Gené, J.; Castañeda-Ruíz, R.F.; Mena-Portales, J.; Crous, P.W.; Guarro, J. Phylogeny of saprobic microfungi from Southern Europe. Stud. Mycol. 2017, 86, 53–97. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, Y.; Ma, L.G.; Zhang, Y.D.; Castañeda-Ruíz, R.F.; Zhang, X.G. Three new species of Neosporidesmium from Hainan, China. Mycol. Prog. 2011, 10, 157–162. [Google Scholar] [CrossRef]
- Goh, T.K. Single-spore isolation using a hand-made glass needle. Fungal Divers. 1999, 2, 47–63. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Xia, J.W.; Ma, Y.R.; Li, Z.; Zhang, X.G. Acrodictys-like wood decay fungi from southern China, with two new families Acrodictyaceae and Junewangiaceae. Sci. Rep. 2017, 7, 7888. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, F.L.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Shang, Z.Q.; Zhang, X.G. Two new Corynespora species from Jiangsu, China. Mycotaxon 2007, 100, 155–158. [Google Scholar]
- Verma, R.K.; Prasher, I.B.; Sushma; Gautam, A.K.; Rajeshkumar, K.C.; Castañeda-Ruíz, R.F. Kirschsteiniothelia shimlaensis sp. nov. from Himachal Pradesh, India. Mycotaxon 2021, 136, 401–407. [Google Scholar] [CrossRef]
- Siqueira, M.V.; Braun, U.; Souza-Motta, C.M. Corynespora subcylindrica sp. nov., a new hyphomycete species from Brazil and a discussion on the taxonomy of corynespora-like genera. Sydowia 2008, 60, 113–122. [Google Scholar]
- Farr, D.F.; Rossman, A.Y. Fungal Databases; U.S. National Fungus Collections, USDA: Washington, DC, USA, 2022. [Google Scholar]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Rodríguez-Andrade, E.; Stchigel, A.M.; Guarro, J.; Cano-Lira, J.F. Fungal diversity of deteriorated sparkling wine and cork stoppers in Catalonia, Spain. Microorganisms 2020, 8, 12. [Google Scholar] [CrossRef]
- Swart, H.J. Australian leaf-inhabiting fungi. XVIII. Corynespora acaciae sp. nov. Trans. Br. Mycol. Soc. 1985, 84, 175–177. [Google Scholar] [CrossRef]
- Wulandari, N.F. Three new species of Corynespora from Indonesia. Mycotaxon 2006, 97, 21–27. [Google Scholar]
- Ellis, M.B. More Dematiaceous Hyphomycetes; Kew, Commonwealth Mycological Institute: Surrey, UK, 1976. [Google Scholar]
- Sharma, N.; Singh, P.N.; Kamal. Three new taxa of Corynespora causing foliar blight in forest plants of North Eastern Uttar Pradesh. J. Mycol. Pl. Pathol. 2003, 33, 26–32. [Google Scholar]
- Meenu, K.; Singh, A.; Singh, S.K. Some new species of Corynespora. Indian Phytopath. 1997, 50, 17–24. [Google Scholar]
- Kumar, S.; Singh, R.; Gond, D.K.; Saini, D.C. Two new species of Corynespora from Uttar Pradesh, India. Mycosphere 2012, 3, 864–869. [Google Scholar] [CrossRef]
- Carris, L.M. Vaccinium fungi: Corynespora arctespora, comb. nov. Mycotaxon 1987, 30, 127–132. [Google Scholar]
- Dubey, R.K.; Rai, A.N. Two new hyphomycetous fungi from India. Indian Phytopath. 2003, 56, 486–490. [Google Scholar]
- Sharma, N.; Chaudhary, R.K.; Kamal. Five undescribed species of Corynespora. Indian Phytopath. 2002, 55, 458–463. [Google Scholar]
- Zhang, K.; Fu, H.B.; Zhang, X.G. Taxonomic studies of Corynespora from Hainan, China. Mycotaxon 2009, 109, 85–93. [Google Scholar] [CrossRef]
- Jain, S.L.; Rai, A.N.; Mehta, P. Additions to the genus Corynespora from India. Indian Phytopath. 2002, 55, 51–56. [Google Scholar]
- Kumar, S.; Singh, A.; Singh, R.; Dubey, N.K. Corynespora bombacina causing foliar disease on Bombax ceiba from Sonebhadra forest of Uttar Pradesh, India. Can. J. Plant Prot. 2013, 1, 76–77. [Google Scholar]
- Ellis, M.B. Some species of Corynespora. Mycol. Pap. 1957, 65, 1–15. [Google Scholar]
- Singh, R.; Kamal. Two new species of Corynespora from northeastern Uttar Pradesh, India. Mycotaxon 2011, 118, 123–129. [Google Scholar] [CrossRef]
- Subramanian, C.V. Hyphomycetes from South East Asia—Novelties from Singapore and Malaysia. Kavaka 1994, 22, 52–76. [Google Scholar]
- Chi, P.K. Fungal Diseases of Cultivated Medicinal Plants in Guangdong Province; Guangdong Science and Technology Press: Guangzhou, China, 1994; pp. 1–275. [Google Scholar]
- Kumar, S.; Singh, R. Corynespora celastri sp. nov. on Celastraceae from India. Stud. Fung. 2016, 1, 125–129. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, S.; Singh, R.; Dubey, N.K. Corynespora clerodendrigena sp. nov. causing foliar disease on Clerodendrum viscosum from Sonebhadra forest of Uttar Pradesh, India. Plant Pathol. Quar. 2013, 3, 15–17. [Google Scholar] [CrossRef]
- Ellis, M.B. Dematiaceous hyphomycetes. V. Mycol. Pap. 1963, 93, 1–33. [Google Scholar]
- Holubová-Jechová, V.; Mercado Sierra, A. Studies on hyphomycetes from Cuba II. Hyphomycetes from the Isla de la Juventud. Ceská Mykol. 1984, 38, 96–120. [Google Scholar]
- Meenu; Kharwar, R.N.; Bhartiya, H.D. Some new forms of genus Corynespora from Kathmandu Valley of Nepal. Indian Phytopath. 1998, 51, 146–151. [Google Scholar]
- Raja, H.A.; Stchigel, A.M.; Miller, A.N.; Crane, J.L.; Shearer, C.A. Hyphomycetes from the great Smoky mountains national park, including three new species. Fungal Divers. 2007, 26, 271–286. [Google Scholar]
- Zhang, X.G.; Xu, J.J. Taxonomic studies of Corynespora from Guangxi, China. Mycotaxon 2005, 92, 431–436. [Google Scholar]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef]
- Wang, X.M.; Zhang, X.G. A new species of Corynespora from Yunnan, China. Mycotaxon 2007, 101, 77–79. [Google Scholar]
- Ma, J.; Zhang, X.G. Three new species of Corynespora from China. Mycotaxon 2007, 99, 353–358. [Google Scholar]
- Singh, A.; Kumar, S.; Singh, R.; Dubey, N.K. A new species of Corynespora causing foliar disease on Ficus religiosa from forest of Sonebhadra, Uttar Pradesh, India. Mycosphere 2012, 3, 890–892. [Google Scholar] [CrossRef]
- Zhang, X.G.; Ji, M. Taxonomic studies of Corynespora from Yunnan, China. Mycotaxon 2005, 92, 425–429. [Google Scholar]
- Ma, L.G.; Ma, J.; Zhang, Y.D.; Zhang, X.G. Craspedodidymum and Corynespora spp. nov. and a new anamorph recorded from southern China. Mycotaxon 2011, 117, 351–358. [Google Scholar] [CrossRef]
- Yen, J.M. Étude sur les champignons parasites du Sud-Est asiatique. 37. Champignons Parasites de Malaisie. 20. Bull. Soc. myc. Fr. 1980, 96, 18–25. [Google Scholar]
- Heuchert, B.; Braun, U. On some dematiaceous lichenicolous hyphomycetes. Herzogia 2006, 19, 11–21. [Google Scholar]
- Guo, Y.L. Four new species of Corynespora. Acta Mycol. Sin. 1984, 3, 161–169. [Google Scholar]
- Ma, J.; Zhang, K.; Zhang, X.G. Taxonomic studies of Corynespora from Hainan, China. Mycotaxon 2008, 104, 153–157. [Google Scholar]
- Saikia, U.N.; Sarbhoy, A.K. Hyphomycetes of North-East India II. Indian Phytopath. 1980, 33, 466–470. [Google Scholar]
- Holubová-Jechová, V. Revisiones generum obscurorum hyphomycetum: Four genera described by A.C.J. Corda. Sydowia 1994, 46, 238–246. [Google Scholar]
- Morgan-Jones, G. Notes on hyphomycetes. LX. Corynespora matuszakii, an undescribed species with narrow, cylindrical, catenate conidia and highly-reduced conidial cell lumina. Mycotaxon 1988, 33, 483–487. [Google Scholar]
- Singh, A.; Kumar, S.; Singh, R.; Dubey, N.K. A new species of Corynespora from Sonebhadra forest of Uttar Pradesh, India. Curr. Res. Envir. App. Myc. 2014, 4, 149–151. [Google Scholar] [CrossRef]
- Meenu, K.; Kamal. New species of Corynespora. Mycol. Res. 1998, 102, 344–346. [Google Scholar] [CrossRef]
- Castañeda-Ruíz, R.F. Fungi Cubenses III; Instituto de Investigaciones Fundamentales en Agricultura Tropical “Alejandro de Humboldt”: Habana, Cuba, 1988; pp. 1–27. [Google Scholar]
- Braun, U.; Crous, P.W.; Nakashima, C. Cercosporoid fungi (Mycosphaerellaceae) 2. Species on monocots (Acoraceae to Xyridaceae, excluding Poaceae). IMA Fung. 2014, 5, 203–390. [Google Scholar] [CrossRef]
- Singh, A.; Singh, S.K.; Kamal. Three new species of Corynespora from India. J. Mycol. Pl. Pathol. 2000, 30, 44–49. [Google Scholar]
- Kumar, S.; Singh, R.; Saini, D.C.; Kamal. A new species of Corynespora Güssow from terai forest of northeastern Uttar Pradesh, India. Mycosphere 2012, 3, 410–412. [Google Scholar] [CrossRef]
- Ellis, M.B. Dematiaceous hyphomycetes. III. Mycol. Pap. 1961, 82, 1–55. [Google Scholar]
- Singh, D.P.; Mall, T.P. Two novel additions to Corynespora Güssow from India. Int. J. Plant Sci. 2011, 6, 321–324. [Google Scholar]
- Loerakker, W.M. A new species of Corynespora. Persoonia 1975, 8, 220–222. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.S.t.J.; Gené, J.; Guarro, J.; Baseia, I.G.; García, D.; Gusmão, L.F.P.; Souza-Motta, C.M.; et al. Fungal Planet description sheets: 716–784. Persoonia 2018, 40, 240–393. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Ma, J.; Zhang, X.G. Two new species of Corynespora from Hainan, China. Mycotaxon 2008, 104, 159–163. [Google Scholar]
- Zhang, X.G.; Shi, C.H. Taxonomic studies of Corynespora from China. Mycotaxon 2005, 92, 417–423. [Google Scholar]
- Castañeda-Ruíz, R.F.; Guarro, J.; Cano, J. Notes on conidial fungi. I. A new species of Corynespora. Mycologia 1995, 87, 271–272. [Google Scholar] [CrossRef]
- Zhang, G.M.; Zhang, X.G. Two new species of Corynespora from Guangdong, China. Mycotaxon 2007, 99, 347–352. [Google Scholar]
- Kumar, S.; Singh, R. Biodiversity, distribution and taxonomy of conidial fungus Corynespora (Corynesporascaceae) associated with Malvaceae. J. Biodivers. Endanger. Species 2016, 4, 166. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.; Chaudhary, S.; Kamal. Three new species of genus Corynespora. Indian Phytopathol. 2002, 55, 178–181. [Google Scholar]
- Sharma, N.; Soni, K.K.; Jamaluddin; Verma, R.K. A new species of Corynespora from central India. Indian Phytopathol. 2005, 58, 503–504. [Google Scholar]
- Crous, P.W.; Luangsa-ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal Planet description sheets: 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef]
- Zhurbenko, M.P.; Braun, U.; Heuchert, B.; Kobzeva, A.A. New lichenicolous hyphomycetes from Eurasia. Herzogia 2015, 28, 584–598. [Google Scholar] [CrossRef]
- Kushwaha, P.; Singh, R.; Kumar, S. Corynespora titarpaniensis sp. nov., on Lepidagathis from Central India. Mycotaxon 2017, 132, 271–279. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Cheewangkoon, R.; van der Bank, M.; Swart, W.J.; Stchigel, A.M.; Cano-Lira, J.F.; Roux, J.; Madrid, H.; et al. Fungal Planet description sheets: 154–213. Persoonia 2013, 31, 188–296. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Singh, R. Diversity, distribution and taxonomy of Corynespora associated with Cannabaceae and Ulmaceae. Plant Pathol. Quar. 2016, 6, 225–231. [Google Scholar] [CrossRef]






| Taxon | Strain Number | GenBank Accession Numbers | |||
|---|---|---|---|---|---|
| SSU | LSU | ITS | tef1-α | ||
| Corynespora cassiicola | CBS 100822 | GU296144 | GU301808 | – | GU349052 |
| C. citricola | CBS 169.77 | – | – | FJ852594 | – |
| C. doipuiensis | MFLUCC 14–0022 | MN648318 | MN648326 | MN648322 | – |
| C. encephalarti | CBS 145555 | – | MK876424 | MK876383 | – |
| C. lignicola | MFLUCC 16–1301 | – | MN860554 | MN860549 | – |
| C. mengsongensis | HJAUP C2000T | OQ060575 | OQ060578 | OQ060574 | – |
| C. nabanheensis | HJAUP C2048T | OQ060576 | OQ060580 | OQ060577 | OQ067526 |
| C. pseudocassiicola | CPC 31708 | – | MH327830 | MH327794 | MH327877 |
| C. smithii | L120 | – | KY984297 | KY984297 | KY984435 |
| C. smithii | L130 | KY984419 | KY984298 | KY984298 | KY984436 |
| C. smithii | CABI 5649b | – | GU323201 | FJ852597 | GU349018 |
| C. smithii | CBS 139925 | – | KY984299 | KY984299 | – |
| C. submersa | MFLUCC 16–1101 | – | MN860553 | MN860548 | – |
| C. torulosa | CBS 136419 | – | MH877634 | MH866095 | – |
| C. thailandica | CBS 145089 | – | MK047505 | MK047455 | MK047567 |
| C. yunnanensis | HJAUP C2132T | OQ060584 | OQ060583 | OQ060579 | – |
| Periconia byssoides | H 4600 | AB797280 | AB807570 | LC014581 | AB808546 |
| P. digitata | CBS 510.77 | AB797271 | AB807561 | LC014584 | AB808537 |
| P. igniaria | CBS 845.96 | AB797277 | AB807567 | LC014586 | AB808543 |
| P. macrospinosa | CBS 135663 | KP184080 | KP184038 | KP183999 | – |
| P. pseudodigitata | KT 1395 | NG_064850 | NG_059396 | NR_153490 | AB808540 |
| Cyclothyriella rubronotata | TR, CBS 121892 | – | KX650541 | KX650541 | KX650516 |
| C. rubronotata | TR9, CBS 141486 | KX650507 | KX650544 | KX650544 | KX650519 |
| Taxon | Strain | Genbank Accession Numbers | ||
|---|---|---|---|---|
| ITS | LSU | SSU | ||
| Acrospermum adeanum | M133 | EU940180 | EU940104 | EU940031 |
| A. compressum | M151 | EU940161 | EU940084 | EU940012 |
| A. gramineum | M152 | EU940162 | EU940085 | EU940013 |
| Anisomeridium ubianum | MPN94 | – | GU327709 | JN887379 |
| Flavobathelium epiphyllum | MPN67 | – | GU327717 | JN887382 |
| Kirschsteiniothelia aethiops | CBS 109.53 | – | AY016361 | AY016344 |
| K. aethiops | MFLUCC 16–1104 | MH182583 | MH182589 | MH182615 |
| K. aethiops | S–783 | MH182586 | MH182595 | MH182617 |
| K. aethiops | MFLUCC 15–0424 | KU500571 | KU500578 | KU500585 |
| K. aquaticaT | MFLUCC 17–1685 | MH182587 | MH182594 | MH182618 |
| K. arasbaranica | IRAN 2509C | KX621986 | KX621987 | KX621988 |
| K. arasbaranicaT | IRAN 2508C | KX621983 | KX621984 | KX621985 |
| K. cangshanensisT | MFLUCC 16–1350 | MH182584 | MH182592 | – |
| K. fluminicolaT | MFLUCC 16–1263 | MH182582 | MH182588 | – |
| K. lignicolaT | MFLUCC 10–0036 | HQ441567 | HQ441568 | HQ441569 |
| K. nabanheensisT | HJAUP C2004 | OQ023197 | OQ023273 | OQ023038 |
| K. nabanheensis | HJAUP C2006 | OQ023274 | OQ023275 | OQ023037 |
| K. phoenicisT | MFLUCC 18–0216 | MG859978 | MG860484 | MG859979 |
| K. rostrataT | MFLUCC 15–0619 | KY697280 | KY697276 | KY697278 |
| K. rostrata | MFLUCC 16–1124 | – | MH182590 | – |
| K. submersaT | MFLUCC 15–0427 | KU500570 | KU500577 | KU500584 |
| K. submersa | S–481 | – | MH182591 | MH182616 |
| K. submersa | S–601 | MH182585 | MH182593 | – |
| K. tectonaeT | MFLUCC 12–0050 | KU144916 | KU764707 | – |
| K. thailandicaT | MFLUCC 20–0116 | MT985633 | MT984443 | MT984280 |
| K. thujina | JF 13210 | KM982716 | KM982718 | KM982717 |
| Megalotremis verrucosa | MPN104 | – | GU327718 | JN887383 |
| Phyllobathelium anomalum | MPN 242 | – | GU327722 | JN887386 |
| P. firmum | ERP 3175 | – | GU327723 | – |
| Pseudorobillarda eucalypti | MFLUCC 12–0422 | KF827451 | KF827457 | KF827463 |
| P. phragmitis | CBS 398.61 | MH858101 | EU754203 | EU754104 |
| Strigula guangxiensisT | HMAS-L0138040 | NR_146255 | MK206256 | – |
| S. nemathora | MPN 72 | – | JN887405 | JN887389 |
| Tenuitholiascus porinoidesT | HMAS-L0139638 | – | MK206259 | MK352441 |
| T. porinoides | HMAS-L0139639 | – | MK206258 | MK352442 |
| T. porinoides | HMAS-L0139640 | – | MK206260 | MK352443 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Hu, Y.; Luo, X.; Castañeda-Ruíz, R.F.; Xia, J.; Xu, Z.; Cui, R.; Shi, X.; Zhang, L.; Ma, J. Molecular Phylogeny and Morphology Reveal Four Novel Species of Corynespora and Kirschsteiniothelia (Dothideomycetes, Ascomycota) from China: A Checklist for Corynespora Reported Worldwide. J. Fungi 2023, 9, 107. https://doi.org/10.3390/jof9010107
Liu J, Hu Y, Luo X, Castañeda-Ruíz RF, Xia J, Xu Z, Cui R, Shi X, Zhang L, Ma J. Molecular Phylogeny and Morphology Reveal Four Novel Species of Corynespora and Kirschsteiniothelia (Dothideomycetes, Ascomycota) from China: A Checklist for Corynespora Reported Worldwide. Journal of Fungi. 2023; 9(1):107. https://doi.org/10.3390/jof9010107
Chicago/Turabian StyleLiu, Jingwen, Yafen Hu, Xingxing Luo, Rafael F. Castañeda-Ruíz, Jiwen Xia, Zhaohuan Xu, Ruqiang Cui, Xugen Shi, Lianhu Zhang, and Jian Ma. 2023. "Molecular Phylogeny and Morphology Reveal Four Novel Species of Corynespora and Kirschsteiniothelia (Dothideomycetes, Ascomycota) from China: A Checklist for Corynespora Reported Worldwide" Journal of Fungi 9, no. 1: 107. https://doi.org/10.3390/jof9010107
APA StyleLiu, J., Hu, Y., Luo, X., Castañeda-Ruíz, R. F., Xia, J., Xu, Z., Cui, R., Shi, X., Zhang, L., & Ma, J. (2023). Molecular Phylogeny and Morphology Reveal Four Novel Species of Corynespora and Kirschsteiniothelia (Dothideomycetes, Ascomycota) from China: A Checklist for Corynespora Reported Worldwide. Journal of Fungi, 9(1), 107. https://doi.org/10.3390/jof9010107