
Morphology and Multigene Phylogeny Revealed Three New Species of Helminthosporium (Massarinaceae, Pleosporales) from China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Isolation and Morphology
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
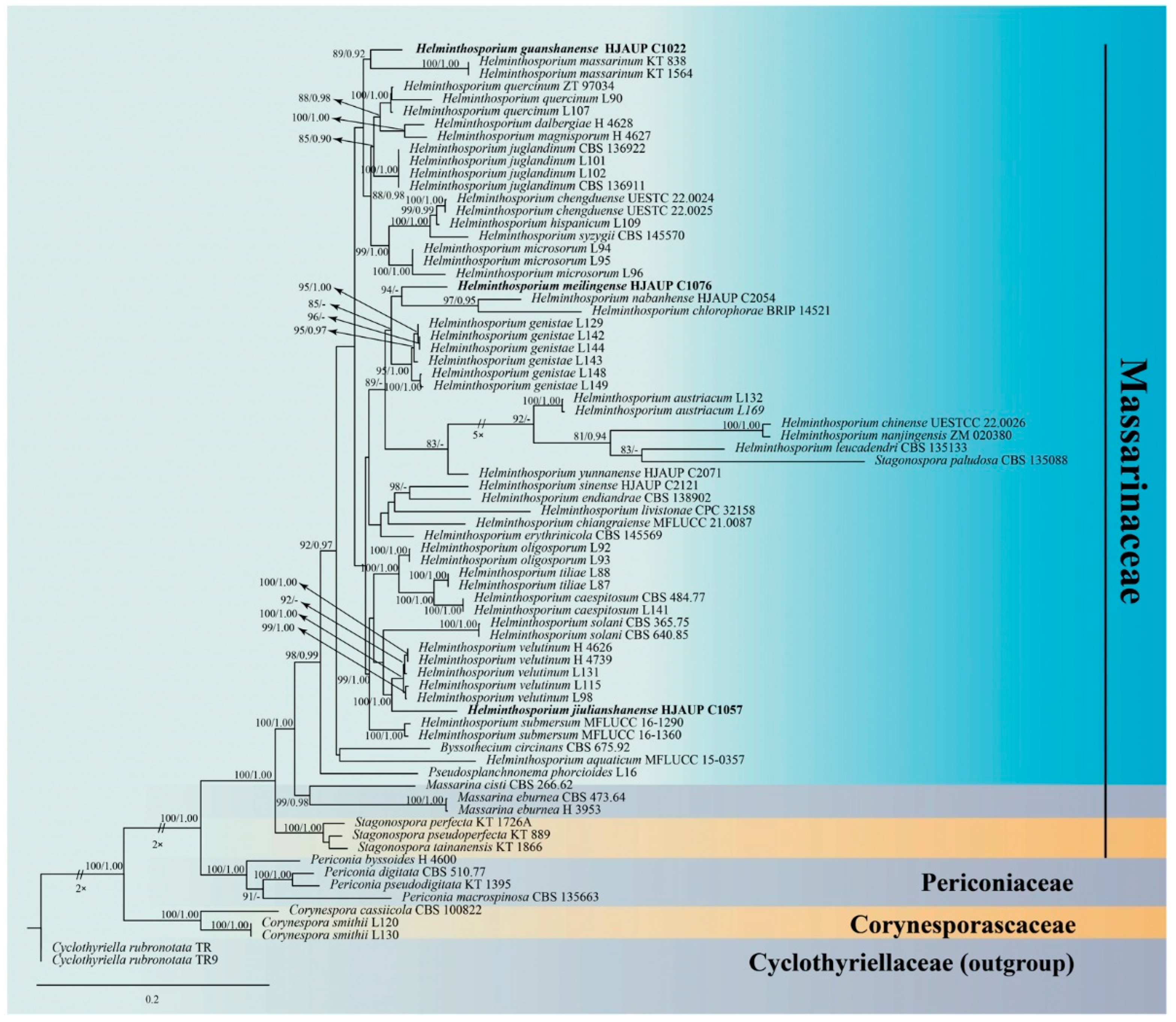
3. Results
3.1. Molecular Phylogeny
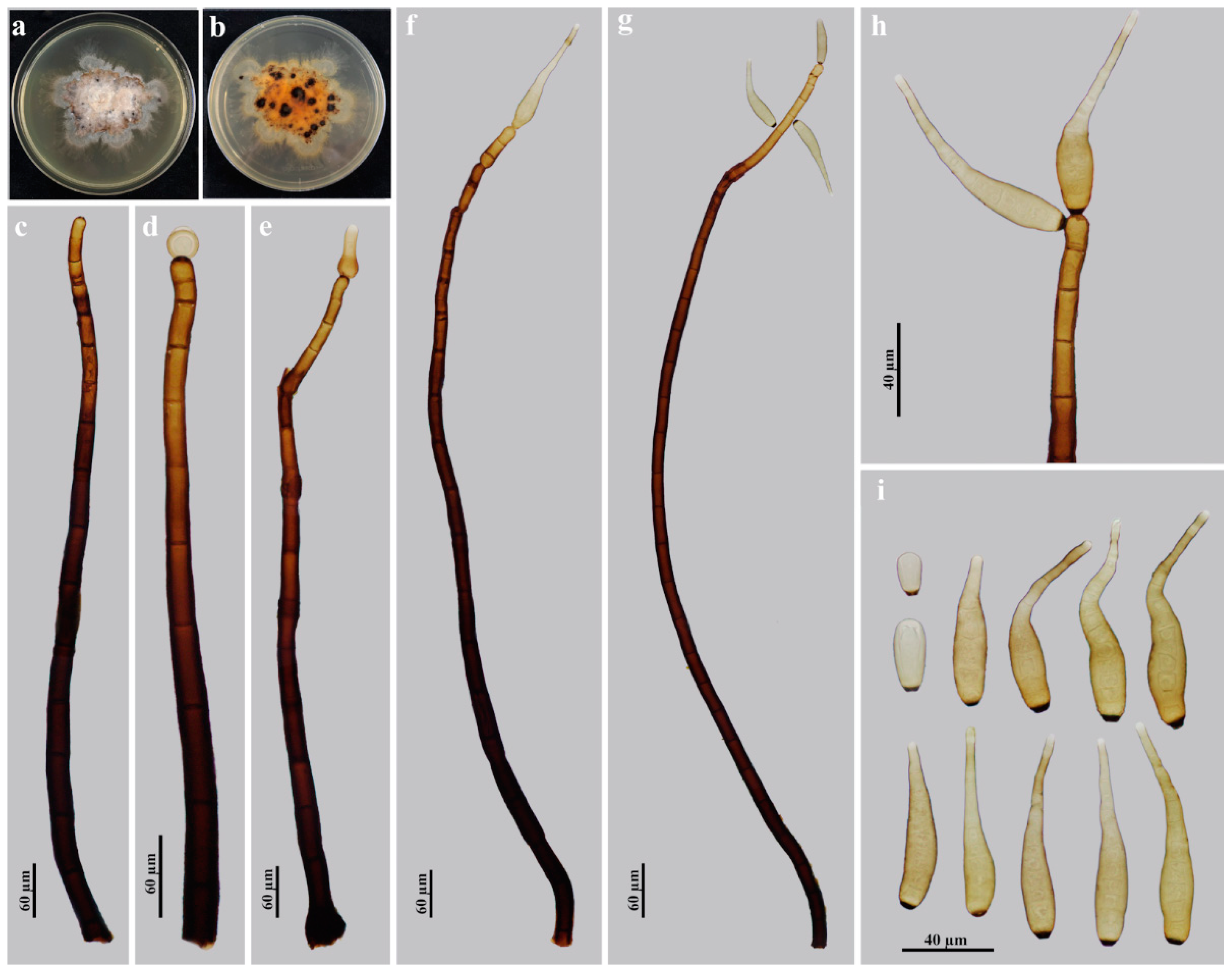
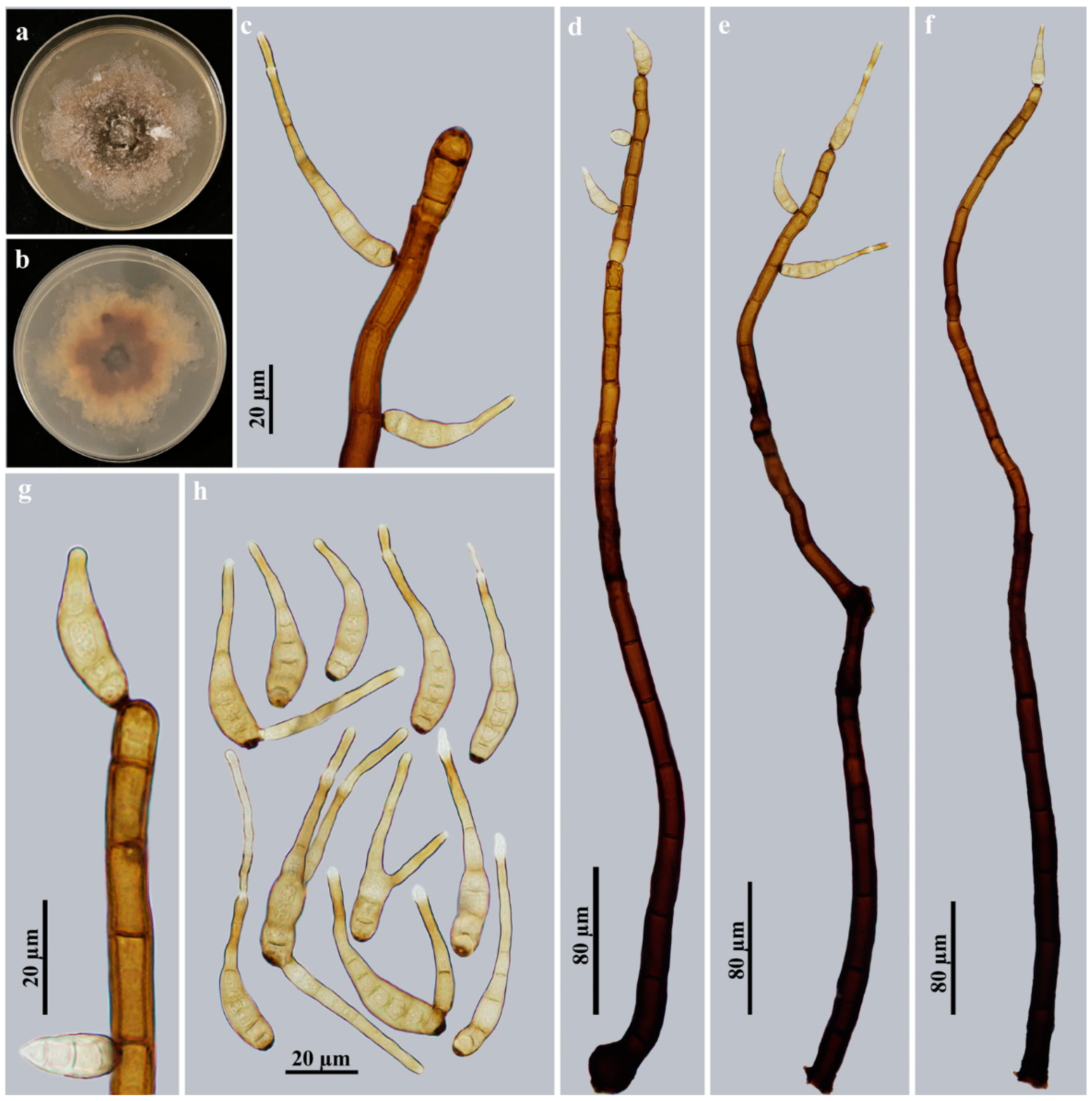
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seifert, K.; Morgan-Jones, G.; Gams, W.; Kendrick, B. The genera of hyphomycetes. CBS Biodivers. Ser. 2011, 9, 1–997. [Google Scholar]
- Wijayawardene, N.N.; Phillips, A.J.L.; Pereira, D.S.; Dai, D.Q.; Aptroot, A.; Monteiro, J.S.; Druzhinina, I.S.; Cai, F.; Fan, X.L.; Selbmann, L.; et al. Forecasting the number of species of asexually reproducing fungi (Ascomycota and Basidiomycota). Fungal Divers. 2022, 114, 463–490. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 6 January 2023).
- Link, H.F. Observationes in Ordines plantarum naturales. Dissertatio prima complectens Anandrarum ordines Epiphytas, Mucedines, Gastromycos et Fungos. Ges. Natur. 1809, 3, 3–42. [Google Scholar]
- Hughes, S.J. Revision es hyphomycetum aliquot cum appendice de nominibusrejiciendis. Can. J. Bot. 1958, 36, 727–836. [Google Scholar] [CrossRef]
- Shoemaker, R.A. Nomenclature of Drechslera and Bipolaris, grass parasites segregated from ‘Helminthosporium’. Can. J. Bot. 1959, 37, 879–887, Erratum in Can. J. Plant Pathol. 2006, 28, S212–S220. [Google Scholar] [CrossRef]
- Ellis, M.B. Dematiaceous hyphomycetes. III. Mycol. Pap. 1961, 82, 1–55. [Google Scholar]
- Tsuda, M.; Ueyama, A.; Nishihara, N. Pseudocochliobolus nisikadoi, the perfect state of Helminthosporium coicis. Mycologia 1977, 69, 1109–1120. [Google Scholar] [CrossRef]
- Sivanesan, A. Graminicolous species of Bipolaris, Curvularia, Exserohilum and their teleomorphs. Mycol. Pap. 1987, 158, 1–261. [Google Scholar]
- Siboe, G.M.; Kirk, P.M.; Cannon, P.F. New dematiaceous hyphomycetes from Kenyan rare plants. Mycotaxon 1999, 73, 283–302. [Google Scholar]
- Castañeda-Ruíz, R.F.; Li, D.W.; Zhang, X.G.; Kendrick, B.; Ramos-García, B.; Pérez-Martínez, S.; Sosa, D. Ellismarsporium gen. nov. and Stanhughesiella gen. nov. to accommodate atypical Helminthosporium and Corynesporella species. Mycotaxon 2017, 132, 759–766. [Google Scholar] [CrossRef]
- Xu, Z.H.; Qiu, L.; Kuang, W.G.; Shi, X.G.; Zhang, X.G.; Castañeda-Ruíz, R.F.; Ma, J. Varioseptispora chinensis gen. & sp. nov., V. apicalis nom. nov., V. hodgkissii comb. nov., and V. versiseptatis comb. nov. Mycotaxon 2020, 135, 753–759. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, H.; Li, D.W.; Castañeda-Ruíz, R.F. Mirohelminthosporium gen. nov. for an atypical Helminthosporium species and H. matsushimae nom. nov. Mycotaxon 2020, 135, 777–783. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, T.Y.; Wu, W.P. Taxonomic studies of Helminthosporium from China II. Two new species in Sichuan Province. Mycosystema 2004, 23, 179–182. [Google Scholar]
- Zhang, M.; Zhang, T.Y.; Wu, W.P. Taxonomic studies of Helminthosporium from China III. Three new species in Guangdong Province. Mycotaxon 2007, 99, 137–142. [Google Scholar]
- Zhang, M.; Wu, H.Y.; Wang, Z.Y. Taxonomic studies of Helminthosporium from China 5. Two new species in Hunan and Sichuan Province. Mycotaxon 2010, 113, 95–99. [Google Scholar] [CrossRef] [Green Version]
- Shirouzu, T.; Harada, Y. Lignicolous dematiaceous hyphomycetes in Japan: Five new records for Japanese mycoflora, and proposals of a new name, Helminthosporium magnisporum, and a new combination, Solicorynespora foveolata. Mycoscience 2008, 49, 126–131. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, T.Y. Taxonomic studies of Helminthosporium from China 4. Six new species and a key to Helminthosporium from China. Mycotaxon 2009, 109, 399–413. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Sun, G.Y. Flora Fungorum Sinicorum, Vol. 30; Helminthosporioid Hyphomycetes; Science Press: Beijing, China, 2010; pp. 1–272. [Google Scholar]
- Zhao, G.C.; Zhao, R.L. The Higher Microfungi from Forests of Yunnan Province; Yunnan Science and Technology Press: Kunming, China, 2012; pp. 1–564. [Google Scholar]
- Wang, X.J.; Wu, H.Y.; Zhang, M. A new species of Helminthosporium from Jiangsu, China. Mycotaxon 2014, 127, 1–4. [Google Scholar] [CrossRef]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Kudo, H.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y.; et al. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Luo, Z.L.; Baht, D.J.; Mckenzie, E.H.C.; Bahkali, A.H.; Zhou, D.Q.; Su, H.Y.; Hyde, K.D. Helminthosporium velutinum and H. aquaticum sp. nov. from aquatic habitats in Yunnan Province, China. Phytotaxa 2016, 253, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Voglmayr, H.; Jaklitsch, W.M. Corynespora, Exosporium and Helminthosporium revisited—New species and generic reclassification. Stud. Mycol. 2017, 87, 43–76. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.S.J.; Gené, J.; Guarro, J.; Baseia, I.G.; García, D.; Gusmão, L.F.P.; Souza-Motta, C.M.; et al. Fungal Planet description sheets: 716–784. Persoonia 2018, 40, 240–393. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Li, W.J.; Hyde, K.D.; Camporesi, E.; Bhat, D.J.; Chomnunti, P.; Xu, J.C. Molecular taxonomy of five species of microfungi on Alnus spp. from Italy. Mycol. Prog. 2018, 17, 255–274. [Google Scholar] [CrossRef]
- Zhao, N.; Luo, Z.L.; Hyde, K.D.; Su, H.Y.; Bhat, D.J.; Liu, J.K.; Bao, D.F.; Hao, Y.E. Helminthosporium submersum sp. nov. (Massarinaceae) from submerged wood in north-western Yunnan Province. China. Phytotaxa 2018, 348, 269–278. [Google Scholar] [CrossRef]
- Boonmee, S.; Wanasinghe, D.N.; Calabon, M.S.; Huanraluek, N.; Chandrasiri, S.K.U.; Jones, G.E.B.; Rossi, W.; Leonardi, M.; Singh, S.K.; Rana, S.; et al. Fungal diversity notes 1387–1511: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2021, 111, 1–335. [Google Scholar] [CrossRef]
- Chen, Y.P.; Tian, W.H.; Guo, Y.B.; Madrid, H.; Maharachchikumbura, S.S.N. Synhelminthosporium gen. et sp. nov. and two new species of Helminthosporium (Massarinaceae, Pleosporales) from Sichuan Province, China. J. Fungi 2022, 8, 712. [Google Scholar] [CrossRef]
- Liu, J.; Hu, Y.; Luo, X.; Castaneda-Ruíz, R.F.; Ma, J. Three novel species of Helminthosporium (Massarinaceae, Pleosporales) from China. MycoKeys 2022, 94, 73–89. [Google Scholar] [CrossRef]
- Matsushima, T. Matsushima Mycological Memoirs No. 7; Matsushima Fungus: Kobe, Japan, 1993; pp. 1–141. [Google Scholar]
- Shirouzu, T.; Harada, Y. Notes on species of Helminthosporium and its allied genera in Japan. Mycoscience 2004, 45, 17–23. [Google Scholar] [CrossRef]
- Konta, S.; Hyde, K.; Karunarathna, S.C.; Mapook, A.; Senwanna, C.; Dauner, L.A.P.; Nanayakkara, C.M.; Xu, J.; Tibpromma, S.; Lumyong, S. Multi-gene phylogeny and morphology reveal Haplohelminthosporium gen. nov. and Helminthosporiella gen. nov. associated with palms in Thailand and a checklist for Helminthosporium reported worldwide. Life 2021, 11, 454. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, Y.; Ma, L.G.; Zhang, Y.D.; Castañeda-Ruíz, R.F.; Zhang, X.G. Three new species of Neosporidesmium from Hainan, China. Mycol. Prog. 2011, 10, 157–162. [Google Scholar] [CrossRef]
- Goh, T.K. Single-spore isolation using a hand-made glass needle. Fungal Divers. 1999, 2, 47–63. [Google Scholar]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute, Kew, Surrey & British Mycological Society: Kew, UK, 1970. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Xia, J.W.; Ma, Y.R.; Li, Z.; Zhang, X.G. Acrodictys-like wood decay fungi from southern China, with two new families Acrodictyaceae and Junewangiaceae. Sci. Rep. 2017, 7, 7888. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S. Primers for Elongation Factor 1-α (EF1-α). 2001. Available online: http://ocid.nacse.org/research/deephyphae/EF1primer.pdf (accessed on 15 December 2022).
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, F.L.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- De Bory, S.V.; De Durieu, M. Exploration Scientifique de l’Algérie Pendant les Années 1840, 1841, 1842 Publiée par Ordre du Gouvernement et Avec le Concours D’une Commission AcadéMique; Sciences Physiques; Botanique par MM Bory de St-.Vincent et Durieu de Maisonneuve membres de la Commission Scientifique D’Algérie; Imprimerie nationale: Paris, France, 1846–1869. [Google Scholar]
- Luttrell, E.S. Systematics of Helminthosporium and related genera. Mycologia 1964, 56, 119–132. [Google Scholar] [CrossRef]
- Ellis, M.B. Dematiaceous Hyphomycetes; Kew, Commonwealth Mycological Institute: Surrey, UK, 1971. [Google Scholar]
- Drechsler, C. Phytopathological and taxonomical aspects of Ophiobolus, Pyrenophora, Helminthosporium and a new genus Cochliobolus. Phytopathology 1934, 24, 953–981. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. I. Mycol. Pap. 1960, 76, 1–36. [Google Scholar]
- Manamgoda, D.S.; Cai, L.; Bahkali, A.H.; Chukeatirote, E.; Hyde, K.D. Cochliobolus: An overview and current status of species. Fungal Divers. 2011, 51, 3–42. [Google Scholar] [CrossRef]
- Manamgoda, D.S.; Cai, L.; McKenzie, E.H.C.; Crous, P.W.; Madrid, H.; Chukeatirote, E.; Shivas, R.G.; Tan, Y.P.; Hyde, K. A phylogenetic and taxonomic re-evaluation of the Bipolaris-Cochliobolus-Curvularia complex. Fungal Divers. 2012, 56, 131–144. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.E.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Wijayawardene, N.N.; Hyde, K.D.; Rajeshkumar, K.C.; Hawksworth, D.L.; Madrid, H.; Kirk, K.M.; Braun, U.; Singh, R.V.; Crous, P.W.; Kukwa, M.; et al. Notes for genera: Ascomycota. Fungal Divers 2017, 86, 1–594. [Google Scholar] [CrossRef] [Green Version]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Subramanian, C.V.; Bhat, D.J. Hyphomycetes from South India I. Some new taxa. Kavaka 1987, 15, 41–74. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs No. 5; Matsushima Fungus: Kobe, Japan, 1987; pp. 1–100. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Schumacher, R.K.; Summerell, B.A.; Giraldo, A.; Gené, J.; Guarro, J.; Wanasinghe, D.N.; Hyde, K.D.; Camporesi, E.; et al. Fungal Planet description sheets: 281–319. Persoonia 2014, 33, 212–289. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, G.V.; Sekar, G. Three bitunicate ascomycetes and their tretic anamorphs. Kavaka 1987, 15, 87–97. [Google Scholar]
- Gornostai, V.I. New and uncommon fungal species in the maritime area. Mikol. Fitopatol. 1972, 6, 154–156. [Google Scholar]
- Quaedvlieg, W.; Verkley, G.J.M.; Shin, H.D.; Barreto, R.W.; Alfenas, A.C.; Swart, W.J.; Groenewald, J.Z.; Crous, P.W. Sizing up Septoria. Stud. Mycol. 2013, 75, 307–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooke, M.C. Some exotic fungi. Grevillea 1883, 12, 37–39. [Google Scholar]
- Saccardo, D. Contribuzione alla micologia veneta e modenense. Malpighia 1898, 12, 201–228. [Google Scholar]
- Hughes, S.J. New Zealand Fungi 27. New species of Guedea, Hadrosporium, and Helminthosporium. N. Z. J. Bot. 1980, 18, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Matsushima, T. Microfungi of the Solomon Islands and Papua-New Guinea; Matsushima Fungus: Kobe, Japan, 1971; pp. 1–78. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Culture/Specimen No. | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| SSU | LSU | ITS | RPB2 | TEF1 | ||
| Byssothecium circinans | CBS 675.92 | GU205235 | GU205217 | OM337536 | DQ767646 | GU349061 |
| Corynespora cassiicola | CBS 100822 | GU296144 | GU301808 | – | GU371742 | GU349052 |
| C. smithii | L120 | – | KY984297 | KY984297 | KY984361 | KY984435 |
| C. smithii | L130 | KY984419 | KY984298 | KY984298 | KY984362 | KY984436 |
| Cyclothyriella rubronotata | TR = CBS 121892 | – | KX650541 | KX650541 | KX650571 | KX650516 |
| C. rubronotata | TR9 ET = CBS 141486 | KX650507 | KX650544 | KX650544 | KX650574 | KX650519 |
| Helminthosporium aquaticum | S-096 HT = MFLUCC 15-0357 | KU697310 | KU697306 | KU697302 | – | – |
| H. austriacum | L132 HT = CBS 139924 | KY984420 | KY984301 | KY984301 | KY984365 | KY984437 |
| H. austriacum | L169 = CBS 142388 | – | KY984303 | KY984303 | KY984367 | KY984439 |
| H. caespitosum | L99 ET = CBS 484.77 | KY984421 | JQ044448 | JQ044429 | KY984370 | KY984440 |
| H. caespitosum | L141 | – | KY984305 | KY984305 | KY984368 | – |
| H. chengduense | UESTC 22.0024 = CGMCC 3.23575 HT | ON557757 | ON557745 | ON557751 | ON563073 | ON600598 |
| H. chengduense | UESTC 22.0025 | ON557756 | ON557744 | ON557750 | ON563072 | ON600597 |
| H. chiangraiense | MFLUCC 21-0087 HT | – | MZ538538 | MZ538504 | – | – |
| H. chinense | UESTCC 22.0026 = CGMCC 3.23570 HT | ON557760 | ON557748 | ON557754 | – | ON600601 |
| H. chlorophorae | BRIP 14521 | – | – | AF120259 | – | – |
| H. dalbergiae | H4628 = MAFF 243853 | AB797231 | AB807521 | LC014555 | – | AB808497 |
| H. endiandrae | CBS 138902 = CPC 22194 HT | – | KP004478 | KP004450 | – | – |
| H. erythrinicola | CBS 145569 HT = CPC 35291 | – | MK876432 | NR_165563 | MK876486 | – |
| H. genistae | L129 = CBS 139922 | KY984423 | KY984309 | KY984309 | KY984373 | – |
| H. genistae | L142 ET = CBS142597 | – | KY984310 | KY984310 | KY984374 | – |
| H. genistae | L143 = CBS 139927 | – | KY984311 | KY984311 | KY984375 | – |
| H. genistae | L144 = CBS 139928 | – | KY984312 | KY984312 | KY984376 | – |
| H. genistae | L148 = CBS 139929 | – | KY984315 | KY984315 | KY984379 | – |
| H. genistae | L149 = CBS 139930 | – | KY984316 | KY984316 | KY984380 | – |
| H. guanshanense | HJAUP C1022 ET | OQ172247 | OQ172239 | OQ172249 | OQ234978 | OQ256247 |
| H. hispanicum | L109 HT = CBS 136917 | KY984424 | KY984318 | KY984318 | KY984381 | KY984441 |
| H. jiulianshanense | HJAUP C1057 ET | – | OQ172253 | OQ172245 | OQ234979 | – |
| H. juglandinum | L101 = CBS 136912 | – | KY984319 | KY984319 | KY984382 | KY984442 |
| H. juglandinum | L102 = CBS 136913 | – | KY984320 | KY984320 | – | – |
| H. juglandinum | L118 HT = CBS 136922 | – | KY984321 | KY984321 | KY984383 | KY984443 |
| H. juglandinum | L97 = CBS 136911 | KY984425 | KY984322 | KY984322 | – | – |
| H. leucadendri | CBS 135133 = CPC19345 HT | – | KF251654 | KF251150 | KF252159 | KF253110 |
| H. livistonae | CPC 32158 = CBS144413 HT | – | NG_064539 | NR_160348 | – | – |
| H. magnisporum | H4627 = MAFF 239278 = TS 33 HT | AB797232 | AB807522 | AB811452 | – | AB808498 |
| H. massarinum | KT 838 ET = MAFF239604 | AB797233 | AB807523 | AB809628 | – | AB808499 |
| H. massarinum | KT 1564 HT = MAFF 239605 = CBS 139690 | AB797234 | AB807524 | AB809629 | – | AB808500 |
| H. meilingense | HJAUP C1076 ET | OQ172246 | OQ172238 | OQ172244 | OQ234980 | OQ234981 |
| H. microsorum | L94 | KY984426 | KY984327 | KY984327 | KY984388 | KY984446 |
| H. microsorum | L95 | – | KY984328 | KY984328 | KY984389 | KY984447 |
| H. microsorum | L96 ET = CBS 136910 | KY984427 | KY984329 | KY984329 | KY984390 | KY984448 |
| H. nabanhense | HJAUP C2054 ET | OP555400 | OP555398 | OP555394 | – | OP961931 |
| H. nanjingense | ZM020380 = HHAUF020380 | – | – | KF192322 | – | – |
| H. oligosporum | L92 = CBS 136908 | KY984428 | KY984332 | KY984332 | KY984393 | KY984450 |
| H. oligosporum | L93 ET = CBS 136909 | – | KY984333 | KY984333 | KY984394 | KY984451 |
| H. quercinum | ZT-97034 = CBS 112393 | – | KY984334 | KY984334 | KY984395 | KY984452 |
| H. quercinum | L107 = CBS 136915 | – | KY984336 | KY984336 | KY984397 | – |
| H. quercinum | L90 HT = CBS 136921 | KY984429 | KY984339 | KY984339 | KY984400 | KY984453 |
| H. sinense | HJAUP C2121 ET | OP555399 | OP555397 | OP555393 | – | OP961932 |
| H. solani | CBS 365.75 | KY984430 | KY984341 | KY984341 | KY984402 | KY984455 |
| H. solani | CBS 640.85 | – | KY984342 | KY984342 | KY984403 | – |
| H. submersum | MFLUCC 16-1360 HT | MG098796 | MG098787 | – | – | MG098586 |
| H. submersum | MFLUCC 16-1290 PT | MG098797 | MG098788 | MG098780 | MG098592 | MG098587 |
| H. syzygii | CPC35312 = CBS 145570 HT | – | MK876433 | NR_165564 | MK876487 | – |
| H. tiliae | L87 = CBS 136906 | – | KY984344 | KY984344 | KY984405 | – |
| H. tiliae | L88 ET = CBS 136907 | KY984431 | KY984345 | KY984345 | KY984406 | KY984457 |
| H. velutinum | H4626 | AB797240 | AB807530 | LC014556 | – | AB808505 |
| H. velutinum | H4739 | AB797235 | AB807525 | LC014557 | – | AB808501 |
| H. velutinum | L115 = CBS 136924 | – | KY984347 | KY984347 | KY984408 | KY984458 |
| H. velutinum | L131 ET = CBS 139923 | KY984432 | KY984352 | KY984352 | KY984413 | KY984463 |
| H. velutinum | L98 | KY984433 | KY984359 | KY984359 | KY984417 | KY984466 |
| H. yunnanense | HJAUP C2071 ET | OP555392 | OP555396 | OP555395 | OP961934 | OP961933 |
| Massarina cisti | CBS 266.62 = JCM 14140 HT | AB797249 | AB807539 | LC014568 | – | AB808514 |
| M. eburnea | CBS 473.64 | AF164367 | GU301840 | AF383959 | GU371732 | GU349040 |
| M. eburnea | H3953 = CBS 139697 | AB521718 | AB521735 | LC014569 | – | AB808517 |
| Periconia byssoides | H4600 = MAFF243872 | AB797280 | AB807570 | LC014581 | – | AB808546 |
| P. digitata | CBS 510.77 | AB797271 | AB807561 | LC014584 | – | AB808537 |
| P. macrospinosa | CBS 135663 | KP184080 | KP184038 | KP183999 | – | – |
| P. pseudodigitata | KT 1395 = CBS139699 = MAFF 239,676 HT | NG_064850 | NG_059396 | NR_153490 | – | AB808540 |
| Pseudosplanchnonema phorcioides | L16 = CBS 122935 | KY984434 | KY984360 | KY984360 | KY984418 | KY984467 |
| Stagonospora paludosa | S 601 NT = CBS 135088 | – | KF251760 | KF251257 | KF252262 | KF253207 |
| S. perfecta | KT 1726A = MAFF 239609 | AB797289 | AB807579 | AB809642 | – | AB808555 |
| S. pseudoperfecta | KT 889 = CBS 120,236 = MAFF 239607 HT | AB797287 | AB807577 | AB809641 | – | AB808553 |
| S. tainanensis | KT 1866 = MAFF 243860 | AB797290 | AB807580 | AB809643 | – | AB808556 |
| Species | Conidiophores (μm) | Conida | Sequence Data | Host/Locality | References | |||
|---|---|---|---|---|---|---|---|---|
| Shape | Colour | Size (µm) | Septa | |||||
| Helminthosporium acaciae | 140–280 × 7–11 | Obclavate | Subhyaline to pale brown | 31–49 × 10–14 | 3–6 | Absent | On dead branches of Acacia farnesiana/Sierra Leone | [7] |
| H. ahmadii | 220–650 × 12–15 | Obclavate, sometimes rostrate | Brown to dark brown | 95–150 × 25–30 | 5–15 | Absent | On dead branches of Quercus sp./Pakistan | [7] |
| H. aquaticum | 410–580 × 13–17 | Obclavate | Pale brown to brown | 70–80 × 16–18 | 8–10 | Present | On submerged decaying wood/China | [23] |
| H. austriacum | 275–700(–920) × 11.5–19 | Obpyriform to lageniform | Pale brown, | (30–)35–48(–97) × (10.0–)13.7–16.5(–19.8) | (4–)5–7(–10) | Present | On dead corticated twigs of Fagus sylvatica/Austria | [24] |
| H. bambusicola | 55–247 × 4–6 | Obclavate | Pale brown, paler towards the apex | 36–66 × 6–11 | 5–8 | Absent | On dead Bambusa sp. culm/China | [16] |
| H. bauhiniae | 350–1100 × 10–15 | Obclavate, rostrate | Subhyaline to pale brown | 55–145 × 16–18 | 7–18 | Absent | On dead twigs of Bauhinia tomentosa/Sierra Leone | [7] |
| H. belgaumense | 260–455 × 6.6–10 | Ellipsoidal to subspherical | Brown to dark brown | 18–24 × 14.9–18.2 | 1 | Absent | On dead twigs of unidentified plant/India | [59] |
| H.caespitosum | (21–)27–37(–44) × (11.2–)12.2–14.5(–16.5) | Medium to dark reddish brown, paler toward the apex | Broadly ellipsoid to obclavate, sometimes rostrate | (67–)82–109(–119) × (22.0–)27.3–35.5(–40.5) | (3–)6–10 | Present | On dead corticated branches of Betula sp./Canada | [24] |
| H. chengduense | 133–391 × 7–15 | Obclavate, sigmoid, lunate or uncinate | Grey-white to pale brown | 41–251 × 8–13 | 3–16 | Present | On decaying branch of unidentified host/China | [30] |
| H. chiangraiense | 168–304.5 × 5.5–12 | Obclavate, rostrate | Pale brown | 141–207 × 14–22 | 9–13 | Present | On dead twigs of unidentified plant/Thailand | [29] |
| H. chinense | 214–461 × 6–16 | Obclavate | Pale gray to brown | 42–109 × 5–11 | 4–10 | Present | On decaying branch of palm trees/China | [30] |
| H. chlorophorae | 120–270 × 7–10 | Obclavate | Subhyaline to pale brown | 52–102 × 8–11 | 6–9 | Present | On dead twigs of Chlorophora regia/Sierra Leone | [7] |
| H. claviphorum | (200–)350–900 × 8.5–11 | Obclavate | Pale yellowish brown | 45–95 × 12–15 | 5–14 | Absent | On rotten branch/Peru | [32] |
| H. conidiophorellum | 60–280 × 7–8.5 | Subulate | Pale brown | 100–147.5 × 9.5–11 | 11–17 | Absent | On dead branches of an unidentified tree/China | [18] |
| H. constrictum | 88–205 × 5–8 | Obclavate | Pale brown, paler towards the apex | 57–120 × 9–12 | 9–15 | Absent | On dead branches of Trachycarpus fortunei/China | [15] |
| H. cubense | (25–)50–150 × 4–5 | Obclavate or cylindrical | Brown, paler towards the apex | 18–62.5 × 6–18 | 3–7 | Absent | On rachis of Roystonea regia/Cuba | [60] |
| H. dalbergiae | 300–1300 × 10–12(–15) | Obclavate | Straw-coloured to pale brown | 58–125 × 12–14 | 5–17 | Present | On dead branches of Dalbergia sissoo/Pakistan | [7] |
| H. dongxingense | 340–650 × 16–25 | Ovoid or obpyriform | Middle brown to brown, paler towards the apex | 50–78 × 17–25 | 6–10 | Absent | On dead branches of Rhododendron sp./China | [19] |
| H. endiandrae | 200–300 × 5–7 | Obclavate | Brown | (35–)37–45(–57) × (7–)8(–9) | 3(–4) | Present | On leaves of Endiandra introrsa/Australia | [24,61] |
| H. erythrinicola | 500–1200 × 6–10 | Obclavate | Medium brown | (70–)80–90(–110) × (9–)10–11(–12) | (6–)7–8(–12) | Present | On leaves of Erythrina humeana/South Africa | [26] |
| H.genistae | (155–)280–460(–560) × 15–23 | Obclavate to rostrate | Pale golden brown to brown | (41–)51–73(–93) × (10.5–)12.7–15.8(–17.5) | 5–12 | Present | On dead corticated twigs of Cytisus scoparius/France | [24] |
| H. guangxiense | 330–850 × 15–20 | Obclavate | Middle brown, paler towards the apex | 76–110 × 16–22 | 9–17 | Absent | On dead branches of an unidentified tree/China | [18] |
| H. guanshanense | 282.5–977.5 × 15–20 | Obclavate, rostrate | Pale brown | 55–107.5 × 10–15 | 3–14 | Present | On dead branches of an unidentified broadleaf tree/China | This study |
| H. hispanicum | 130–540 × 13–22.5 | Obclavate | Pale brown | 69–99(–130) × (17–)18–21(–24) | (4–)6–11(–14) | Present | On dead corticated twigs of Juglans regia/Spain | [24] |
| H. hunanense | 70–226 × 5–7 | Obclavate | Middle brown, paler towards the apex | 56–127 × 10–14 | 4–12 | Absent | On dead branches of an unidentified tree/China | [16] |
| H. italicum | (190–)330–600 × (12–)16–18(–20) | Obclavate | Pale brown to brown, with apical cell paler than other cells, | 58–78 × 15–19(–23) | 6–11 | Present | On dead branch of Alnus glutinosa/Italy | [27] |
| H. jiulianshanense | (290–)531–712 × 10–15 | Obclavate, rostrate | Pale brown to brown | (57–)78–120 × 13–17.5 | 6–13 | Present | On dead branches of an unidentified broadleaf tree/China | This study |
| H. juglandinum | (175–)215–325(–455) × 11–23 | Obclavate, rostrate | Pale brown | (69–)89–145(–205) × (15.0–)16.5–20.0(–25.0) | (5–)9–17(–20) | Present | On dead corticated twigs of Juglans regia/Austria | [24] |
| H. juglandis | 619–1030 × 10.5–14 | Clavate | Brown | 50–119 × 10–12.7 | 4–15 | Absent | On living branches of Juglans regia/China | [20] |
| H. kakamegense | 250–550 × 8–12 | Obclavate, rostrate | Subhyaline | 30–90 × 8–10 | 4–15 | Absent | On dead attached twig of Uvariopsis congensis/Kenya | [10] |
| H. kalakadense | 1000–2000 × 17–25 | Obclavate | – | 45–60 × 13–16 | 8 | Absent | On dead unidentified twig/India | [24,62] |
| H. kalopanacis | 63.1–207.5 × 8.3–13.3 | Subcylindrical | Pale dark brown | 33.2–59.8 × 10–16.6 | 2–5 | Absent | On dead wood of Kalopanax septemlobus/Russia | [63] |
| H. leucadendri | 100–300 × 4–6(–7) | Obclavate to subcylindrical | Medium brown | (35–)70–110(–170) × (6–)7–8(–11) | (3–)4–6(–10) | Present | On leaves of Leucadendron sp./South Africa | [24,64] |
| H. ligustri | 127–700 × 9.5–18 | Obclavate, rostrate or pseudorostrate | Pale brown, subhyaline towards the apex | 24–38.5 × 9.5–13 | 4–6 | Absent | On dead branches of an unidentified tree/China | [18] |
| H. livistonae | Up to 500 × 4–6 | subcylindrical | Medium brown | (25–)40–55(–65) × (7–)8–9 | (3–)4–6(–7) | Present | On leaves of Livistona australis/Australia | [25] |
| H. longisinuatum | 20–75 × 3.5–5 | Narrowly obclavate | Middle brown, paler toward the apex | 65–220(–1000) × 8–10.5 | 9–22 | Absent | On rotten trunk of Palmae/Peru | [32] |
| H. magnisporum | 150–270 × 8.5–13.5 | Obclavate, rostrate | Pale olive-brown to pale brown, paler toward the apex | 100–203 × 12.5–22.5 | 7–18 | Present | On dead fallen branches of an unknown woody plant/Japan | [17,33] |
| H. massarinum | 380–810 × 7–9 | Obclavate, rostrate | Pale brown | 17–56.5 × 5–9 | 1–8 | Present | On vines of Berchemia racemosa/Japan | [22] |
| H. matsushimae | 20–65 × 3–4.5 | Cylindrical | Medium to dark brown | (20–) 50–100 × 6–8.5 | (3–) 6–14 | Absent | On rotten petiole of Palmae/Peru | [13,32] |
| H. mauritianum | 250–750 × 14–20 | Obclavate | Subhyaline to rather pale brown | 27–55 × 8–13 | 3–7 | Absent | On twigs and stems/Mauritius | [7,65] |
| H. meilingense | 544–712.5 × 12.5–17 | Obclavate, rostrate | Pale brown | (20.7–)41.5–82.8 × 6.9–10.4 | 6–13 | Present | On dead branches of an unidentified broadleaf tree/China | This study |
| H. microsorum | 100–550 × 8–14 | Obclavate | Pale to mid golden-brown | 60–160 × 12–22 | 9–17 | Present | On twigs of Quercus ilex/Italy | [6,66] |
| H. multiseptatum | 390–650 × 10–14 | Thinly obclavate or nearly whip-like | Pale brown, paler towards the apex | 78–190 × 11–16 | 13–25 | Absent | On dead branches of an unidentified tree/China | [15] |
| H. nabanhense | 365–557 × 6.5–13.5 | Obclavate, rostrate | Pale brown to brown | 26.5–46.5 × 6.5–10 | 3–6 | Present | On dead branches of an unidentified broadleaf tree/China | [31] |
| H. nanjingense | 250–470 × 6.9–7.7 | Subulate or nearly whip-like | Pale brown | 64.5–170.5 × 7.3–10.3 | 6–17 | Present | On dead branches of an unidentified tree/China | [21] |
| H. novae-zelandiae | 165–330 × 12.5–14.5 | Obclavate to fusiform, sometimes shortly rostrate | Golden brown to dark brown, paler toward the apex | 56–103 × 16–21.5 | (5–)6–7(–8) | Absent | On dead wood and bark of Vitex lucens/New Zealand | [67] |
| H. obpyriforme | 225–460 × 9.5–13 | Obpyriform | Middle brown, paler towards the apex | 47–74 × 14–19 | 5–9 | Absent | On dead branches of an unidentified tree/China | [18] |
| H. oligosporum | (17–)22–35(–46) × (8.0–)8.5–10.5(–11.5) | Obclavate, sometimes rostrate, smooth but occasionally wrinkled with age | Pale brown to brown, paler toward the apex | (37–)59–80(–124) × (14.8–)15.8–18.0(–20.0) | 6–12(–16) | Present | On dead corticated twigs of Tilia cordata/Austria | [24] |
| H. ovoideum | 380–510 × 15–25 | Ovoid to ellipsoidal | Moderately brown, paler towards the apex | 27–61 × 13–21 | 3–8 | Absent | On dead branches of an unidentified tree/China | [18] |
| H. palmigenum | 70–180(–250) × 7–10 | Obclavate | Below pale brown, upper subhyaline | 27–47 × 6.5–9 | 4–9 | Absent | On rotten fruit of Cocos nucifera/Papua New Guinea | [68] |
| H. pseudomicrosorum | 155–288 × 11–15 | Obclavate | Brown, paler towards the apex | 82–142 × 17–27 | 7–16 | Absent | On dead branches of an unidentified tree/China | [18] |
| H. quercicola | (115–)133–226(–300) × 14–20 | Obclavate | Brown | 60–100 × 15–22 | 8–10 | Absent | On dead corticated branches of Quercus cf. reticulata/USA | [24] |
| H. quercinum | (40–)74–199(–332) × 11–18 | Obclavate, rostrate | Brown | (47–)78–130(–201) × (13.2–)15.3–18.0(–20.5) | 8–13(–20) | Present | On dead corticated twigs of Quercus petraea/Austria | [24] |
| H. sichuanense | 300–550 × 14–25 | Obclavate, | Middle brown, paler towards the apex | 41–86 × 10–14 | 5–11 | Absent | On dead branches of an unidentified plant/China | [14] |
| H. sinense | 220–370 × 6–8.5 | Obclavate | Pale brown | 37–60 × 5.5–8.5 | 2–7 | Present | On dead branches of an unidentified broadleaf tree/China | [31] |
| H. solani | 120–600 × 9–15 | Obclavate | Subhyaline to brown | 24–85 × 7–11 | 2–8 | Present | On stem of Solanum nigrum/England | [7,49] |
| H. spurirostrum | 200–600 × 18–23 | Obclavate, sometimes pseudorostrate | Moderately brown to brown, paler to the apex | 27–73 × 7–15.5 | 4–7 | Absent | On dead branches of an unidentified plant/China | [14] |
| H. subhyalinum | 120–200 × 6–8.5 | Thinly obclavate | Subhyaline | 72–125 × 9–11.5 | 6–9 | Absent | On living leaves of Phoenix hanceana/China | [15] |
| H. submersum | 239–423 × 8.5–15.5 | Obclavate, rostrate | Pale brown to mid-brown | 41–55 × 14.5–18.5 | 6–10 | Present | On submerged decaying wood/China | [28] |
| H. syzygii | 150–400 × 10–15 | Obclavate | Medium brown | (70–)80–100(–150) × (19–)22–23(–25) | (7–)9–12 | Present | On bark canker of Syzygium sp./South Africa | [26] |
| H.tiliae | (68–)79–133(–150) × 9–15 | Obclavate to rostrate | Pale to golden brown | (57–)74–111(–122) × (13.5–)13.7–19.0(–24.5) | 7–18(–25) | Present | On dead corticated branches of Tilia platyphyllos/Austria | [24] |
| H. velutinum | (163–)340–698(–960) × 14–26 | Obclavate to rostrate | Pale golden brown to brown | (42–)56–89(–142) × (11–)14.3–18.5(–24.7) | 6–18 | Present | On dead corticated twigs of Fagus sylvatica/Austria | [4,24] |
| H. yunnanense | 560–680 × 12.5–15.5 | Obclavate, sigmoid, lunate or uncinate | Pale brown | 30.5–55.5 × 9–11 | 4–7 | Present | On dead branches of an unidentified broadleaf tree/China | [31] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.-F.; Liu, J.-W.; Xu, Z.-H.; Castañeda-Ruíz, R.F.; Zhang, K.; Ma, J. Morphology and Multigene Phylogeny Revealed Three New Species of Helminthosporium (Massarinaceae, Pleosporales) from China. J. Fungi 2023, 9, 280. https://doi.org/10.3390/jof9020280
Hu Y-F, Liu J-W, Xu Z-H, Castañeda-Ruíz RF, Zhang K, Ma J. Morphology and Multigene Phylogeny Revealed Three New Species of Helminthosporium (Massarinaceae, Pleosporales) from China. Journal of Fungi. 2023; 9(2):280. https://doi.org/10.3390/jof9020280
Chicago/Turabian StyleHu, Ya-Fen, Jing-Wen Liu, Zhao-Huan Xu, Rafael F. Castañeda-Ruíz, Kai Zhang, and Jian Ma. 2023. "Morphology and Multigene Phylogeny Revealed Three New Species of Helminthosporium (Massarinaceae, Pleosporales) from China" Journal of Fungi 9, no. 2: 280. https://doi.org/10.3390/jof9020280
APA StyleHu, Y. -F., Liu, J. -W., Xu, Z. -H., Castañeda-Ruíz, R. F., Zhang, K., & Ma, J. (2023). Morphology and Multigene Phylogeny Revealed Three New Species of Helminthosporium (Massarinaceae, Pleosporales) from China. Journal of Fungi, 9(2), 280. https://doi.org/10.3390/jof9020280