
Detoxication of Citrinin with Kojic Acid by the Formation of the Citrinin-Kojic Acid Adduct, and the Enhancement of Kojic Acid Production by Citrinin via Oxidative Stress in Aspergillus parasiticus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi | Metabolism | Phenotype | Ref. No. |
|---|---|---|---|
| A. parasiticus SYS-4 | AF+, KA+ | A. parasiticus wild strain (=NRRL2999) | [13] |
| A. parasiticus NFRI-95 | NA+, AFΔ (1), KA+ | a NA-accumulating mutant derived from A. parasiticus SYS-4 by UV irradiation | [13] |
| A. parasiticus NIAH-26 | AF−, KA+ | a mutant of A. parasiticus SYS-4 that produces neither AFs nor any pigmented precursors of AFs | [13] |
| A. parasiticus 11AP, 34C8, 48B11, 34C8, and 59A6 | AF+, KAΔ (1) or AF+, KA− | The isolated mutants with impaired KA productivity from SYS-4 by UV irradiation | This work |
| A. oryzae RIB40 | AF−, KA+ | A. oryzae wild strain (NRIB, Japan) | |
| A. oryzae RIB143, RIB430, and RIB1025 | AF−, KA− | A. oryzae KA-non-producing strains (NRIB) | |
| Penicillium citrinum NFRI-MI190 | CTN+ | The fungus isolated from soil as an inhibitory fungus to aflatoxin production by A. parasiticus | [12] |
2.2. Metabolites
2.3. Media
2.4. Effect of Each Metabolite on Bacterial Growth
2.5. Effect of Each Metabolite on Growth of HeLa Cells
2.6. Tip Culture Method
2.7. Isolation of A. Parasiticus Mutants with Impaired KA Production
2.8. Measurement of KA Concentration in the Fungal Media
2.9. Co-Culture of Two Different Fungi
2.10. RT-PCR
3. Results
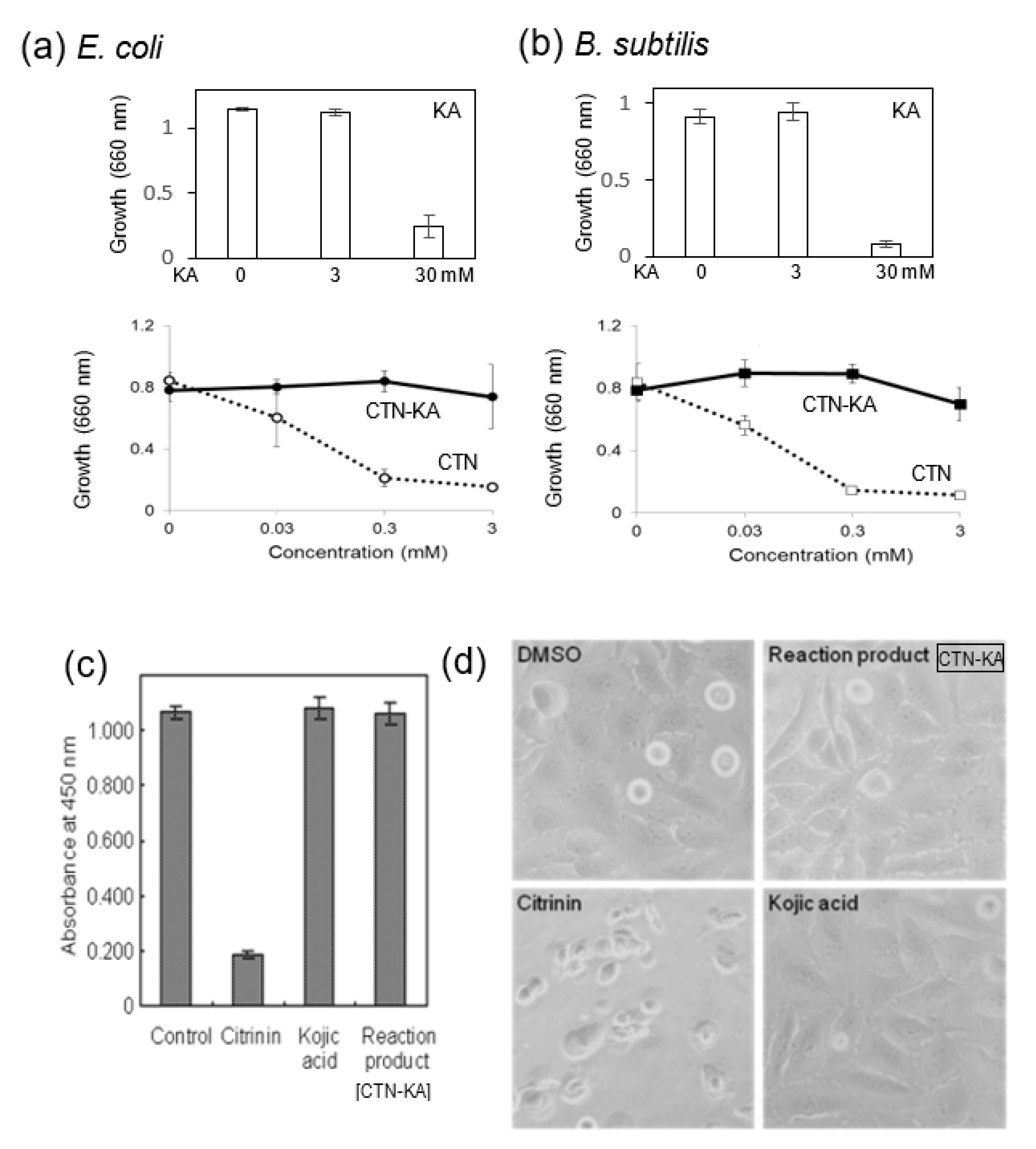
3.1. Toxicity of the CTN-KA Adduct to Bacteria as Well as HeLa Cells
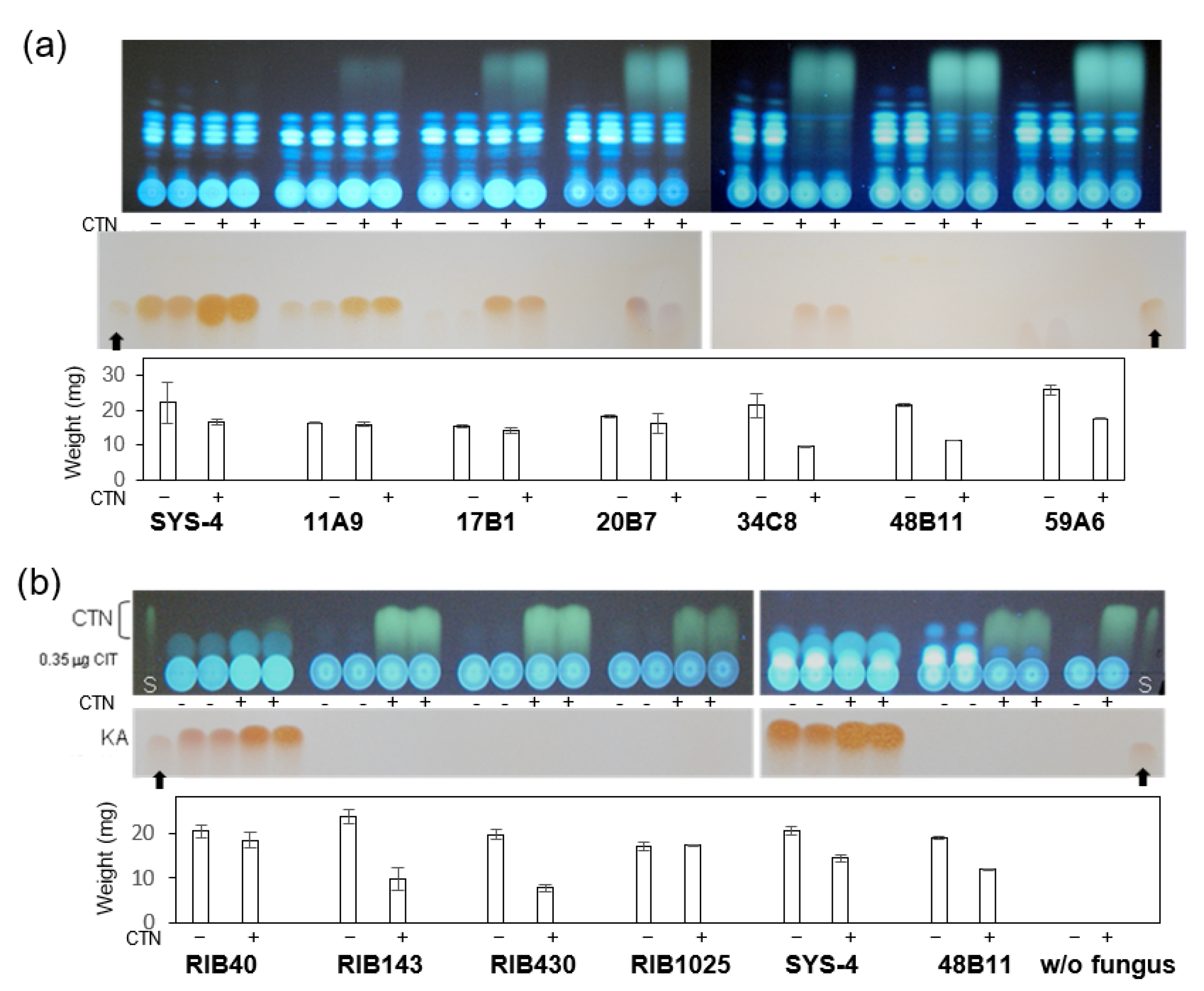
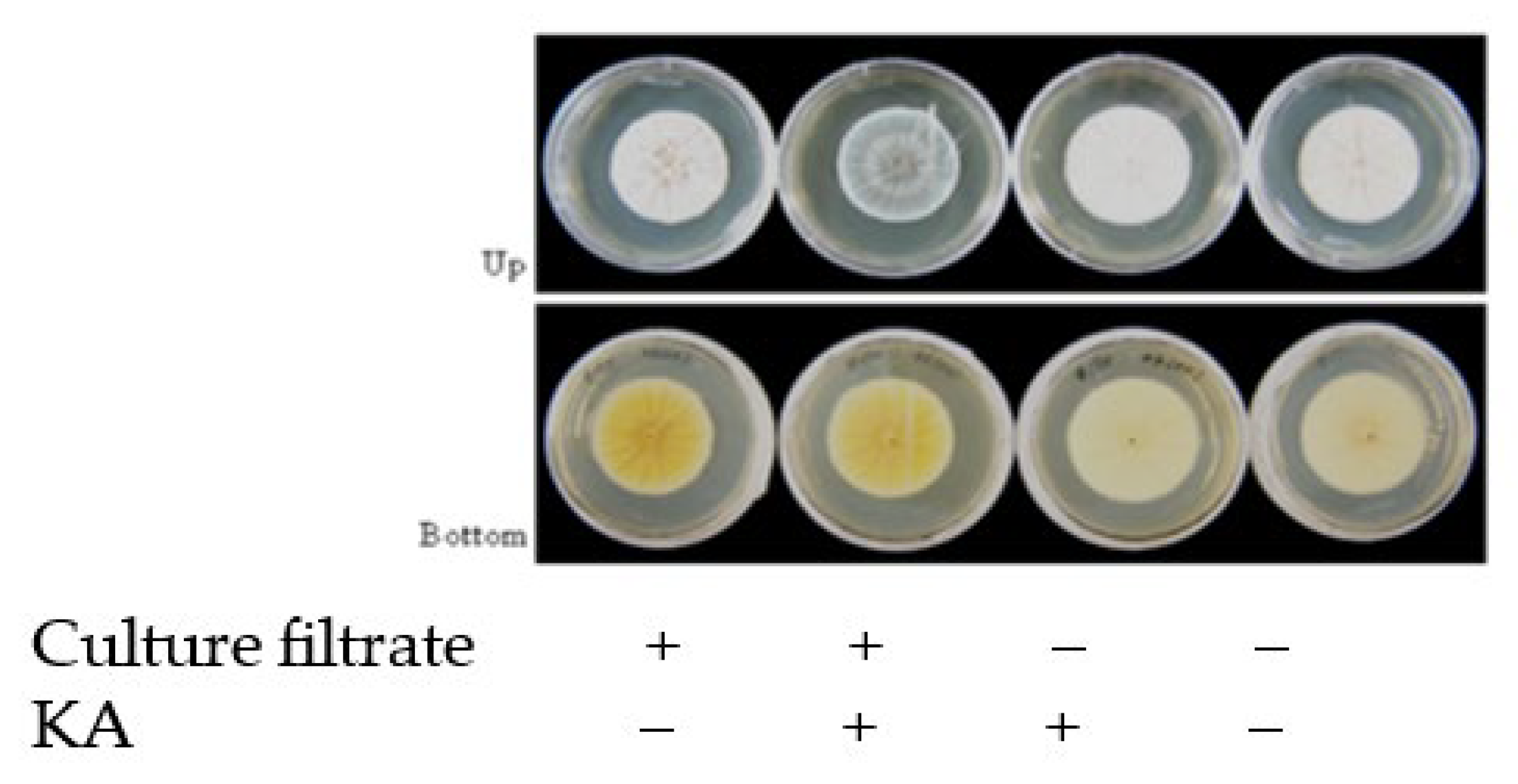
3.2. Effect of CTN on KA-Producing or KA-Non-Producing Fungi by the Tip Culture Method
3.3. Effects of CTN on KA Producers and KA Non-Producers in Agar Medium
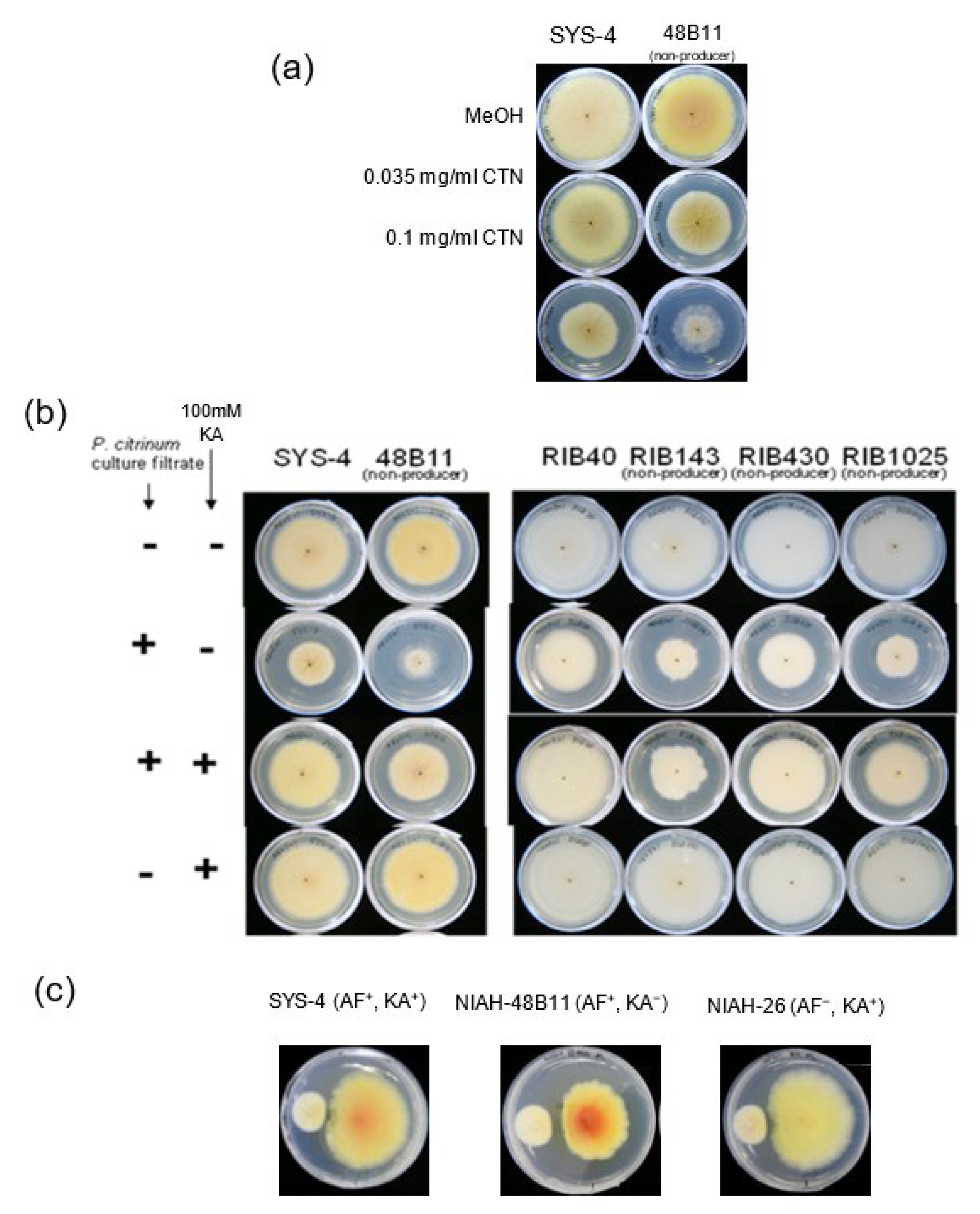
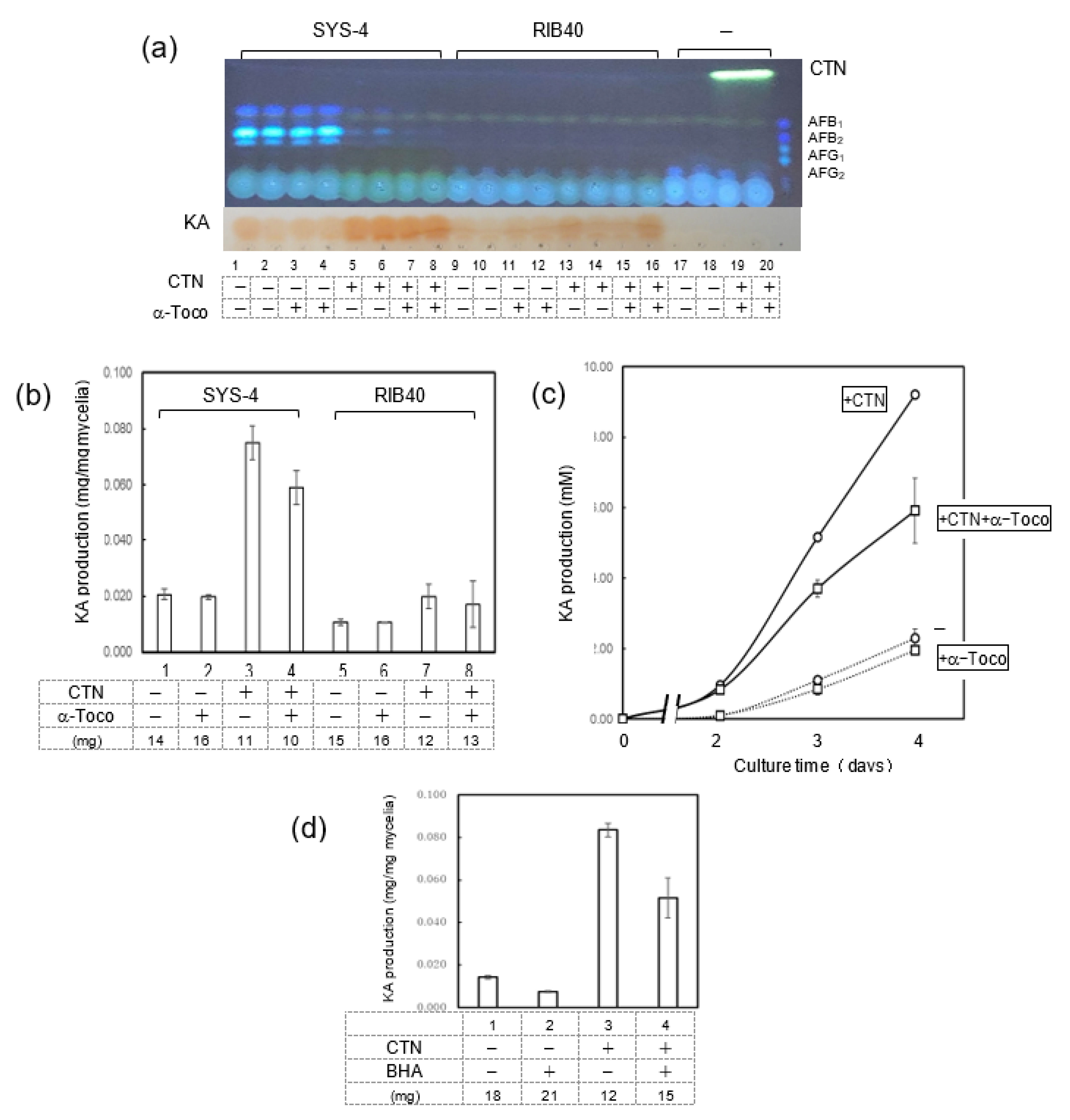
3.4. Effects of CTN and α-Tocopherol on KA Production by Fungi
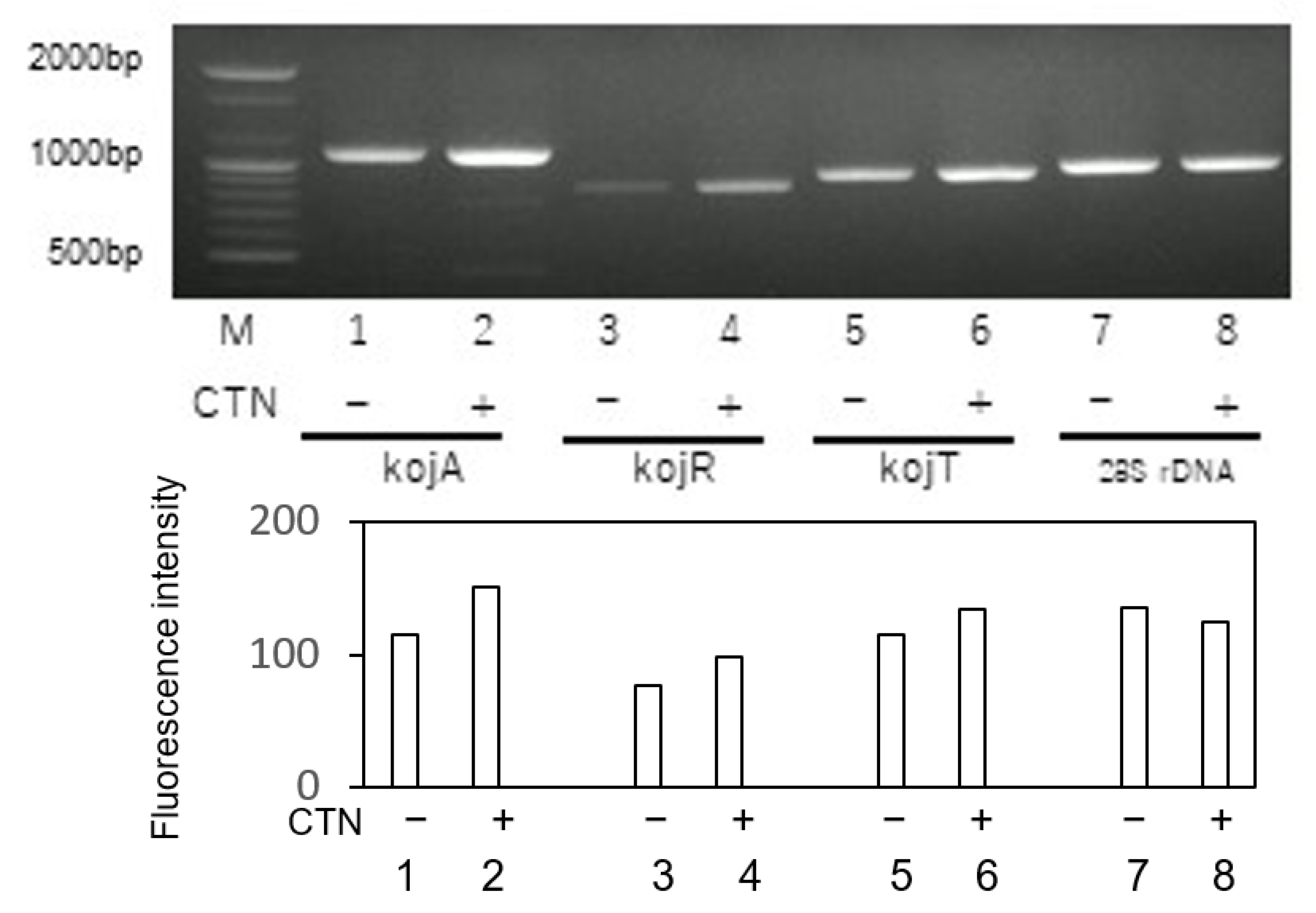
3.5. Effect of CTN on the Gene Expressions of KA Biosynthesis Genes
3.6. Effect of α-Tocopherol on Aflatoxin Production as Well as Fungal Growth
3.7. Effect of KA on the Growth of P. Citrinum
4. Discussion
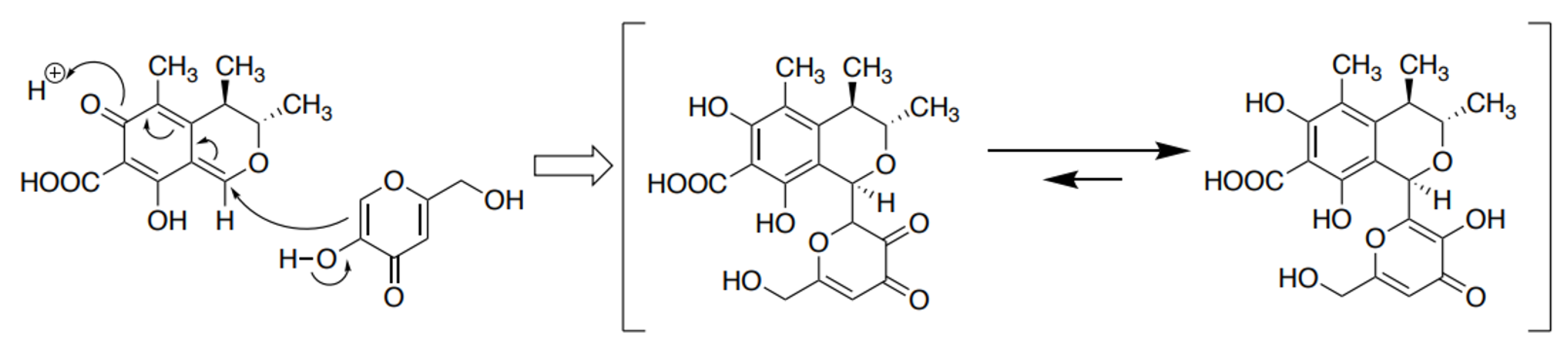
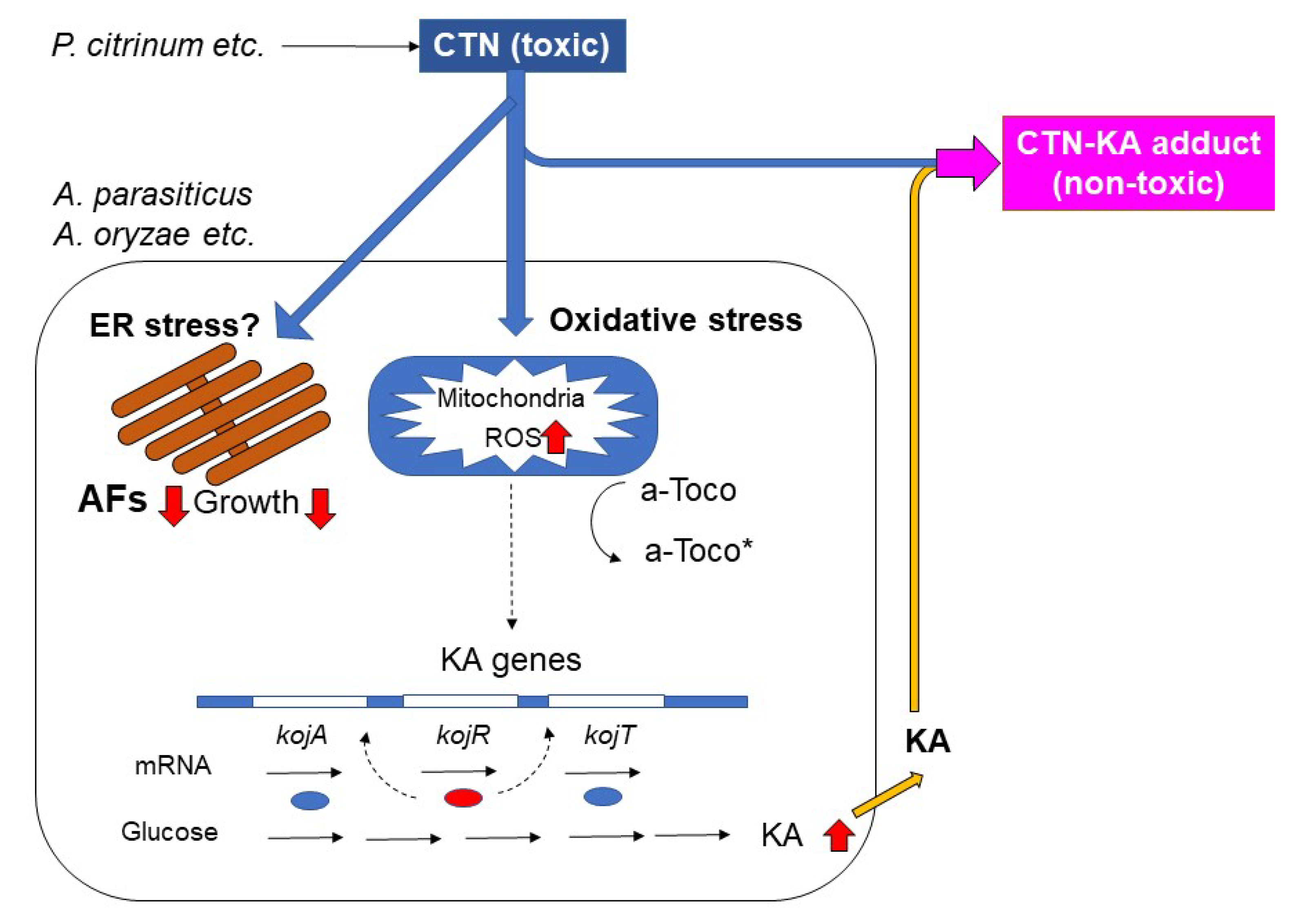
4.1. The Detoxication of CTN with KA to Form the CTN-KA Adduct
4.2. Effects of CTN on KA-Producing Fungi
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tola, M.; Kebede, B. Occurrence, Importance and Control of Mycotoxins: A Review. Cogent Food Agric. 2016, 2, 1191103. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [Green Version]
- International Agency for Research on Cancer. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; WHO Press: Lyon, France, 1993; Volume 56, pp. 41–487. [Google Scholar]
- Juzlovâ, P.; Martïnkovâ, L.; Kren, V. Secondary Metabolites of the Fungus Monascus: A Review. J. Ind. Microbiol. 1996, 16, 163–170. [Google Scholar] [CrossRef]
- Wu, D.; Yang, C.; Yang, M.; Wu, Y.; Mao, Y.; Zhou, X.; Wang, J.; Yuan, Z.; Wu, J. Citrinin-Induced Hepatotoxicity in Mice Is Regulated by the Ca2+ /Endoplasmic Reticulum Stress Signaling Pathway. Toxins 2022, 14, 259. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Song, Y.; Sakuno, E.; Nakajima, H. Cyclo (l-Leucyl-l-Prolyl) Produced by Achromobacter xylosoxidans Inhibits Aflatoxin Production by Aspergillus parasiticus. Appl. Environ. Microbiol. 2004, 70, 7466–7473. [Google Scholar] [CrossRef] [Green Version]
- Yoshinari, T.; Akiyama, T.; Nakamura, K.; Kondo, T.; Takahashi, Y.; Muraoka, Y.; Nonomura, Y.; Nagasawa, H.; Sakuda, S. Dioctatin A Is a Strong Inhibitor of Aflatoxin Production by Aspergillus parasiticus. Microbiology 2007, 153, 2774–2780. [Google Scholar] [CrossRef] [Green Version]
- Rajani, P.; Sridevi, V.; Lakshmi, M.V.V.C. A Review on Biological Control of Aflatoxin Crop Contamination. Int. J. Chem. Environ. Pharm. Res. 2012, 3, 83–86. [Google Scholar] [CrossRef]
- Mossini, S.A.G.; Kemmelmeier, C. Inhibition of Citrinin Production in Penicillium citrinum Cultures by Neem [Azadirachta indica A. Juss (Meliaceae)]. Int. J. Mol. Sci. 2008, 9, 1676–1684. [Google Scholar] [CrossRef] [Green Version]
- Bentley, R. From Miso, Sake and Shoyu to Cosmetics: A Century of Science for Kojic Acid. Nat. Prod. Rep. 2007, 23, 1046–1062. [Google Scholar] [CrossRef]
- Bentley, R. Preparation and Analysis of Kojic Acid. Methods Enzymol. 1957, 3, 238–241. [Google Scholar] [CrossRef]
- Ichinomiya, M.; Kawamoto, A.; Sakuno-Fukushima, E.; Nakagawa, H.; Nakajima, H.; Yabe, K. Inhibition of Aflatoxin Production by Citrinin and Non-Enzymatic Formation of a Novel Citrinin-Kojic Acid Adduct. J. Fungi 2023, 9, 29. [Google Scholar] [CrossRef]
- Yabe, K.; Nakamura, H.; Ando, Y.; Terakado, N.; Nakajima, H.; Hamasaki, T. Isolation and Characterization of Aspergillus parasiticus Mutants with Impaired Aflatoxin Production by a Novel Tip Culture Method. Appl. Environ. Microbiol. 1988, 54, 2096–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yabe, K.; Hatabayashi, H.; Ikehata, A.; Zheng, Y.; Kushiro, M. Development of the Dichlorvos-Ammonia (DV-AM) Method for the Visual Detection of Aflatoxigenic Fungi. Appl. Microbiol. Biotechnol. 2015, 99, 10681–10694. [Google Scholar] [CrossRef] [PubMed]
- Yabe, K.; Ando, Y.; Hamasaki, T. Biosynthetic Relationship among Aflatoxins B1, B2, G1, and G2. Appl. Environ. Microbiol. 1988, 54, 2101–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terabayashi, Y.; Sano, M.; Yamane, N.; Marui, J.; Tamano, K.; Sagara, J.; Dohmoto, M.; Oda, K.; Ohshima, E.; Tachibana, K.; et al. Identification and Characterization of Genes Responsible for Biosynthesis of Kojic Acid, an Industrially Important Compound from Aspergillus oryzae. Fungal Genet Biol. 2010, 47, 953–961. [Google Scholar] [CrossRef]
- Kitabatake, N.; Trivedi, A.B.; Doi, E. Thermal Decomposition and Detoxication of Citrinin under Various Moisture Conditions. J. Agric. Food Chem. 1991, 39, 2240–2244. [Google Scholar] [CrossRef]
- Trivedi, A.; Doi, E.; Kitabatake, N. Toxic Compounds Formed on Prolonged Heating of Citrinin under Watery Conditions. J. Food Sci. 1993, 58, 229–232. [Google Scholar] [CrossRef]
- Hirota, M.; Menta, A.B.; Yoneyama, K.; Kitabatake, N. A Major Decomposition Product, Citrinin H2, from Citrinin on Heating with Moisture. Biosci. Biotechnol. Biochem. 2002, 66, 206–210. [Google Scholar] [CrossRef]
- Lee, C.; Chen, W.; Wang, J.; Pan, T.-M. A Simple and Rapid Approach for Removing Citrinin While Retaining Monacolin K in Red Mold Rice. J. Agric. Food Chem. 2007, 55, 11101–11108. [Google Scholar] [CrossRef]
- Burnett, C.L.; Bergfeld, W.F.; Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.C.; Marks, J.G.; Shank, R.C.; Slaga, T.J.; Snyder, P.W.; et al. Final Report of the Safety Assessment of Kojic Acid as Used in Cosmetics. Int. J. Toxicol. 2010, 29, 244S–273S. [Google Scholar] [CrossRef]
- LeBlanc, D.T.; Akers, H.A. Maltol and Ethyl Maltol: From the Larch Tree to Successful Food Additive. Synthesis of a Natural Compound Led To. Food Technol. 1989, 43, 78–84. [Google Scholar]
- Chaudhary, J. Production Technology and Applications of Kojic Acid. Annu. Res. Rev. Biol. 2014, 4, 3165–3196. [Google Scholar] [CrossRef]
- Rodrigues, A.P.D.; Farias, L.H.S.; Carvalho, A.S.C.; Santos, A.S.; Do Nascimento, J.L.M.; Silva, E.O. A Novel Function for Kojic Acid, a Secondary Metabolite from Aspergillus Fungi, as Antileishmanial Agent. PLoS ONE 2014, 9, e91259. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.-Y.; Shi, X.-C.; Wang, S.-Y.; Wang, B.; Laborda, P. Antifungal Mechanism and Efficacy of Kojic Acid for the Control of Sclerotinia sclerotiorum in Soybean. Front. Plant Sci. 2022, 13, 845698. [Google Scholar] [CrossRef]
- Morton, H.E.; Kocholaty, W.; Junowicz-, R.; Kelner, A. Toxicity and Antibiotic Activity of Kojic Acid Produced. J. Bacteriol. 1945, 50, 579–584. [Google Scholar] [CrossRef] [Green Version]
- El-Kady, I.A.; Zohri, A.N.A.; Hamed, S.R. Kojic Acid Production from Agro-Industrial By-Products Using Fungi. Biotechnol. Res. Int. 2014, 2014, 642385. [Google Scholar] [CrossRef] [Green Version]
- Brtko, J.; Rondahl, L.; Ficková, M.; Hudecová, D.; Eybl, V.; Uher, M. Kojic Acid and Its Derivatives: History and Present State of Art. Cent. Eur. J. Public Health 2004, 12, S16–S18. [Google Scholar] [CrossRef]
- Haraguchi, H.; Tanaka, T.; Taniguchi, M.; Oi, S.; Hashimoto, K. Inhibitory Effects of Citrinin on Mitochondrial Function. Agric. Biol. Chem. 1986, 51, 1453–1454. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Dwivedi, P.D.; Dhawan, A.; Das, M.; Ansari, K.M. Citrinin-Generated Reactive Oxygen Species Cause Cell Cycle Arrest Leading to Apoptosis via the Intrinsic Mitochondrial Pathway in Mouse Skin. Toxicol. Sci. 2011, 122, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Doi, K.; Uetsuka, K. Mechanisms of Mycotoxin-Induced Dermal Toxicity and Tumorigenesis through Oxidative Stress-Related Pathways. J. Toxicol. Pathol. 2014, 27, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Pascual-Ahuir, A.; Vanacloig-Pedros, E.; Proft, M. Toxicity Mechanisms of the Food Contaminant Citrinin: Application of a Quantitative Yeast Model. Nutrients 2014, 6, 2077–2087. [Google Scholar] [CrossRef] [PubMed]
- Chagas, G.M.; Campello, A.P.; Klüppel, M.L.W. Mechanism of Citrinin-Induced Dysfunction of Mitochondria. I. Effects on Respiration, Enzyme Activities and Membrane Potential of Renal Cortical Mitochondria. J. Appl. Toxicol. 1992, 12, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.V.; Ellerby, H.M.; Bredesen, D.E. Coupling Endoplasmic Reticulum Stress to the Cell Death Program. Cell Death Differ. 2004, 11, 372–380. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ichinomiya, M.; Kawamoto, A.; Yamaguchi, T.; Iwashita, K.; Nagashima, H.; Hatabayashi, H.; Nakajima, H.; Yabe, K. Detoxication of Citrinin with Kojic Acid by the Formation of the Citrinin-Kojic Acid Adduct, and the Enhancement of Kojic Acid Production by Citrinin via Oxidative Stress in Aspergillus parasiticus. J. Fungi 2023, 9, 51. https://doi.org/10.3390/jof9010051
Ichinomiya M, Kawamoto A, Yamaguchi T, Iwashita K, Nagashima H, Hatabayashi H, Nakajima H, Yabe K. Detoxication of Citrinin with Kojic Acid by the Formation of the Citrinin-Kojic Acid Adduct, and the Enhancement of Kojic Acid Production by Citrinin via Oxidative Stress in Aspergillus parasiticus. Journal of Fungi. 2023; 9(1):51. https://doi.org/10.3390/jof9010051
Chicago/Turabian StyleIchinomiya, Masayuki, Ayaka Kawamoto, Takahiro Yamaguchi, Keiko Iwashita, Hitoshi Nagashima, Hidemi Hatabayashi, Hiromitsu Nakajima, and Kimiko Yabe. 2023. "Detoxication of Citrinin with Kojic Acid by the Formation of the Citrinin-Kojic Acid Adduct, and the Enhancement of Kojic Acid Production by Citrinin via Oxidative Stress in Aspergillus parasiticus" Journal of Fungi 9, no. 1: 51. https://doi.org/10.3390/jof9010051
APA StyleIchinomiya, M., Kawamoto, A., Yamaguchi, T., Iwashita, K., Nagashima, H., Hatabayashi, H., Nakajima, H., & Yabe, K. (2023). Detoxication of Citrinin with Kojic Acid by the Formation of the Citrinin-Kojic Acid Adduct, and the Enhancement of Kojic Acid Production by Citrinin via Oxidative Stress in Aspergillus parasiticus. Journal of Fungi, 9(1), 51. https://doi.org/10.3390/jof9010051