
Reference-Based RADseq Unravels the Evolutionary History of Polar Species in ‘the Crux Lichenologorum’ Genus Usnea (Parmeliaceae, Ascomycota)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Sampling
2.2. Morphological and Chemical Characterization
2.3. DNA Extraction and RAD Library Preparation
2.4. RADseq Assembly
2.5. Phylogenomic Analyses
2.6. Estimates of Divergence Times
3. Results
3.1. Morphological and Chemical Characterization
3.2. Assembly of RAD Sequencing
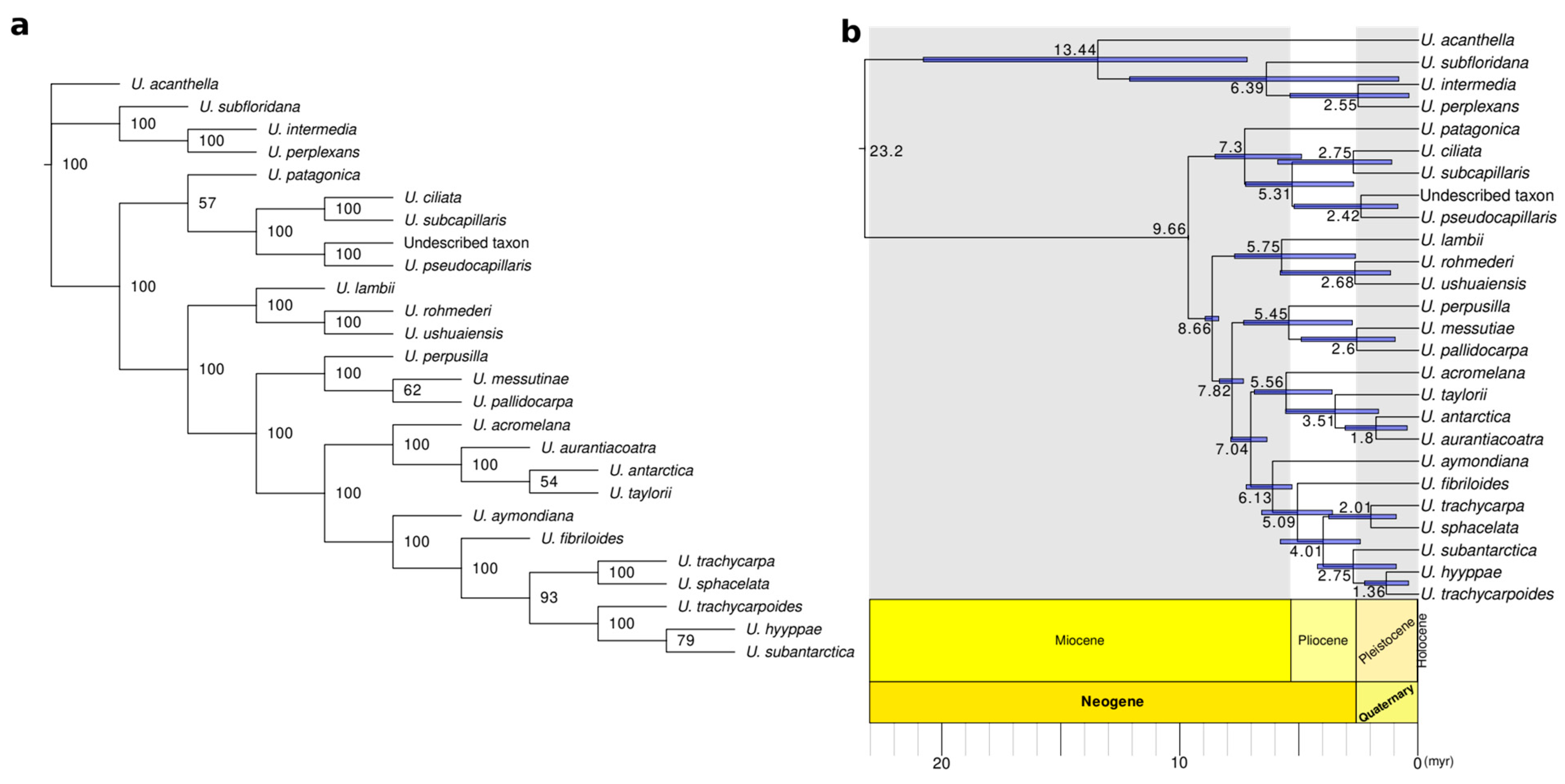
3.3. Phylogenomic Analyses and Divergence Times Estimation
3.4. Taxonomy
3.4.1. Usnea aymondiana A.Otero, Barcenas-Peña, Lumbsch & Grewe sp. nov. (Figure 1r,s)
3.4.2. Usnea fibriloides A.Otero, Barcenas-Peña, Lumbsch & Grewe sp. nov. (Figure 1t,u)
4. Discussion
4.1. Contributions to the Systematics of Neuropogonoid Lichens
4.2. The Usnea Perpusilla Complex
4.3. The Usnea Trachycarpa Complex
4.4. The New Zealand Clade
4.5. A Spatio-Temporal Evolutionary Framework of Neuropogonoid Lichens
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choi, J.; Kim, S.-H. A genome tree of life for the Fungi Kingdom. Proc. Natl. Acad. Sci. USA 2017, 114, 9391–9396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.-W.; May, T.W. Fungal taxonomy: Current status and research agendas for the interdisciplinary and globalisation era. Mycology 2022, 1–8. [Google Scholar] [CrossRef]
- Cheek, M.; Nic Lughadha, E.; Kirk, P.; Lindon, H.; Carretero, J.; Looney, B.; Douglas, B.; Haelewaters, D.; Gaya, E.; Llewellyn, T. New scientific discoveries: Plants and Fungi. Plants People Planet 2020, 2, 371–388. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Lücking, R. Fungal diversity revisited: 2.2 to 3.8 Million species. Microbiol. Spectr. 2017, 5, 5.4.10. [Google Scholar] [CrossRef]
- Nic Lughadha, E.; Bachman, S.P.; Govaerts, R. Plant states and fates: Response to Pimm and Raven. Trends Ecol. Evol. 2017, 32, 887–889. [Google Scholar] [CrossRef]
- Lumbsch, H.T.; Leavitt, S.D. Goodbye morphology? a paradigm shift in the delimitation of species in lichenized Fungi. Fungal Divers. 2011, 50, 59–72. [Google Scholar] [CrossRef]
- Lücking, R.; Leavitt, S.D.; Hawksworth, D.L. Species in lichen-forming fungi: Balancing between conceptual and practical considerations, and between phenotype and phylogenomics. Fungal Divers. 2021, 109, 99–154. [Google Scholar] [CrossRef]
- Barcenas-Peña, A.; Leavitt, S.D.; Grewe, F.; Lumbsch, H.T. Diversity of Xanthoparmelia (Parmeliaceae) species in mexican xerophytic scrub vegetation, evidenced by molecular, morphological and chemistry data. An. Jard. Bot. Madr. 2021, 78, e107. [Google Scholar] [CrossRef]
- Barcenas-Peña, A.; Diaz, R.; Grewe, F.; Widhelm, T.; Lumbsch, H.T. Contributions to the phylogeny of Lepraria (Stereocaulaceae) species from the southern hemisphere, including three new species. Bryologist 2021, 124, 494–505. [Google Scholar] [CrossRef]
- Hoffman, J.R.; Lendemer, J.C. A meta-analysis of trends in the application of sanger and next-generation sequencing data in lichenology. Bryologist 2018, 121, 133–147. [Google Scholar] [CrossRef]
- Crespo, A.; Lumbsch, H.T. Cryptic species in lichen-forming fungi. IMA Fungus 2010, 1, 167–170. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lücking, R. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef]
- James, T.Y.; Kauff, F.; Schoch, C.L.; Matheny, P.B.; Hofstetter, V.; Cox, C.J.; Celio, G.; Gueidan, C.; Fraker, E.; Miadlikowska, J.; et al. Reconstructing the early evolution of Fungi using a six-gene phylogeny. Nature 2006, 443, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, N.; Printzen, C.; Lumbsch, H.T. Using haplotype networks, estimation of gene flow and phenotypic characters to understand species delimitation in fungi of a predominantly antarctic Usnea Group (Ascomycota, Parmeliaceae). Org. Divers. Evol. 2012, 12, 17–37. [Google Scholar] [CrossRef]
- Boluda, C.G.; Hawksworth, D.L.; Divakar, P.K.; Crespo, A.; Rico, V.J. Microchemical and molecular investigations reveal Pseudephebe species as cryptic with an environmentally modified morphology. Lichenologist 2016, 48, 527–543. [Google Scholar] [CrossRef] [Green Version]
- Del-Prado, R.; Divakar, P.K.; Lumbsch, H.T.; Crespo, A.M. Hidden genetic diversity in an asexually reproducing lichen forming fungal group. PLoS ONE 2016, 11, e0161031. [Google Scholar] [CrossRef] [Green Version]
- Leavitt, S.D.; Divakar, P.K.; Crespo, A.; Lumbsch, H.T. A matter of time—Understanding the limits of the power of molecular data for delimiting species boundaries. Heia 2016, 29, 479–492. [Google Scholar] [CrossRef]
- Zhao, X.; Fernández-Brime, S.; Wedin, M.; Locke, M.; Leavitt, S.D.; Lumbsch, H.T. Using multi-locus sequence data for addressing species boundaries in commonly accepted lichen-forming fungal species. Org. Divers. Evol. 2017, 17, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Articus, K.; Mattsson, J.-E.; Tibell, L.; Grube, M.; Wedin, M. Ribosomal DNA and β-tubulin data do not support the separation of the Lichens Usnea florida and U. subfloridana as distinct species. Mycol. Res. 2002, 106, 412–418. [Google Scholar] [CrossRef]
- Buschbom, J.; Mueller, G.M. Testing “Species Pair” hypotheses: Evolutionary processes in the lichen-forming species complex Porpidia flavocoerulescens and Porpidia melinodes. Mol. Biol. Evol. 2006, 23, 574–586. [Google Scholar] [CrossRef]
- Kotelko, R.; Piercey-Normore, M.D. Cladonia pyxidata and C. Pocillum; genetic evidence to regard them as conspecific. Mycologia 2010, 102, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Printzen, C.; Ekman, S.; Tønsberg, T. Phylogeography of Cavernularia hultenii: Evidence of slow genetic drift in a widely disjunct lichen. Mol. Ecol. 2003, 12, 1473–1486. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Mendoza, F.; Domaschke, S.; García, M.A.; Jordan, P.; Martín, M.P.; Printzen, C. Population structure of mycobionts and photobionts of the widespread lichen Cetraria aculeata. Mol. Ecol. 2011, 20, 1208–1232. [Google Scholar] [CrossRef] [PubMed]
- Kanz, B.; von Brackel, W.; Cezanne, R.; Eichler, M.; Hohmann, M.-L.; Teuber, D.; Printzen, C. DNA Barcodes for the distinction of reindeer lichens: A case study using Cladonia rangiferina and C. stygia. Heia 2015, 28, 445–464. [Google Scholar] [CrossRef]
- Lutsak, T.; Fernández-Mendoza, F.; Kirika, P.; Wondafrash, M.; Printzen, C. Coalescence-based species delimitation using genome-wide data reveals hidden diversity in a cosmopolitan group of lichens. Org. Divers. Evol. 2020, 20, 189–218. [Google Scholar] [CrossRef]
- Widhelm, T.J.; Grewe, F.; Goffinet, B.; Wedin, M.; Goward, T.; Coca, L.F.; Distefano, I.; Košuthová, A.; Lumbsch, H.T. Phylogenomic reconstruction addressing the peltigeralean backbone (Lecanoromycetes, Ascomycota). Fungal Divers. 2021, 110, 59–73. [Google Scholar] [CrossRef]
- Otero, A.; Vargas, P.; Fernández-Mazuecos, M.; Jiménez-Mejías, P.; Valcárcel, V.; Villa-Machío, I.; Hipp, A.L. A snapshot of progenitor–derivative speciation in Iberodes (Boraginaceae). Mol. Ecol. 2022, 31, 3192–3209. [Google Scholar] [CrossRef]
- Grewe, F.; Huang, J.-P.; Leavitt, S.D.; Lumbsch, H.T. Reference-based RADseq resolves robust relationships among closely related species of lichen-forming fungi using metagenomic DNA. Sci. Rep. 2017, 7, 9884. [Google Scholar] [CrossRef] [Green Version]
- Jorna, J.; Linde, J.B.; Searle, P.C.; Jackson, A.C.; Nielsen, M.-E.; Nate, M.S.; Saxton, N.A.; Grewe, F.; Herrera-Campos, M.d.l.A.; Spjut, R.W. Species boundaries in the messy middle—A genome-scale validation of species delimitation in a recently diverged lineage of coastal fog desert lichen fungi. Ecol. Evol. 2021, 11, 18615–18632. [Google Scholar] [CrossRef]
- Widhelm, T.J.; Grewe, F.; Huang, J.-P.; Ramanauskas, K.; Mason-Gamer, R.; Lumbsch, H.T. Using RADseq to understand the circum-antarctic distribution of a lichenized fungus, Pseudocyphellaria glabra. J. Biogeogr. 2021, 48, 78–90. [Google Scholar] [CrossRef]
- Martín-Hernanz, S.; Aparicio, A.; Fernández-Mazuecos, M.; Rubio, E.; Reyes-Betancort, J.A.; Santos-Guerra, A.; Olangua-Corral, M.; Albaladejo, R.G. Maximize resolution or minimize error? using genotyping-by-sequencing to investigate the recent diversification of Helianthemum (Cistaceae). Front. Plant Sci. 2019, 10, 1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hipp, A.L.; Manos, P.S.; Hahn, M.; Avishai, M.; Bodénès, C.; Cavender-Bares, J.; Crowl, A.A.; Deng, M.; Denk, T.; Fitz-Gibbon, S. Genomic landscape of the global oak phylogeny. New Phytol. 2020, 226, 1198–1212. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, Y.X.; Warren, B.H. An overview of current population genomics methods for the analysis of whole-genome resequencing data in eukaryotes. Mol. Ecol. 2021, 30, 6036–6071. [Google Scholar] [CrossRef]
- Lücking, R.; Nadel, M.R.A.; Araujo, E.; Gerlach, A. Two decades of DNA barcoding in the genus Usnea (Parmeliaceae): How useful and reliable is the ITS? Plant Fungal Syst. 2020, 65, 303–357. [Google Scholar] [CrossRef]
- Walker, F.J. Lichen Genus Usnea Subgenus Neuropogon; British Museum (Natural History): London, UK, 1985; ISBN 0-565-08004-0. [Google Scholar]
- Chown, S.L.; Gaston, K.J. Areas, cradles and museums: The latitudinal gradient in species richness. Trends Ecol. Evol. 2000, 15, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.I. Vascular plants as bioindicators of regional warming in Antarctica. Oecologia 1994, 99, 322–328. [Google Scholar] [CrossRef]
- Convey, P. Antarctic climate change and its influences on terrestrial ecosystems. In Trends in Antarctic Terrestrial and Limnetic Ecosystems: Antarctica as a Global Indicator; Bergstrom, D.M., Convey, P., Huiskes, A.H.L., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 253–272. ISBN 978-1-4020-5277-4. [Google Scholar]
- Bokhorst, S.; Convey, P.; Huiskes, A.; Aerts, R. Usnea antarctica, an important antarctic lichen, is vulnerable to aspects of regional environmental change. Polar Biol. 2016, 39, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Ellis, C.J.; Yahr, R.; Hodkinson, T.R.; Jones, M.B.; Waldren, S.; Parnell, J.A.N. An interdisciplinary review of climate change trends and uncertainties: Lichen biodiversity, arctic-alpine ecosystems and habitat loss. In Climate Change, Ecology and Systematics; Cambridge University Press: Cambridge, UK, 2011; pp. 457–489. [Google Scholar]
- Convey, P.; Bindschadler, R.; Di Prisco, G.; Fahrbach, E.; Gutt, J.; Hodgson, D.A.; Mayewski, P.A.; Summerhayes, C.P.; Turner, J.; Consortium, A. Antarctic Climate Change and the Environment. Antarct. Sci. 2009, 21, 541–563. [Google Scholar]
- Lumbsch, H.T.; Wirtz, N. Phylogenetic relationships of the neuropogonoid core group in the genus Usnea (Ascomycota: Parmeliaceae). Lichenologist 2011, 43, 553–559. [Google Scholar] [CrossRef]
- Jatta, A. Sylloge Lichenum Italicorum; Typis V. Vecchi: Trani, Italy, 1990. [Google Scholar]
- Du Rietz, G.E. Om Släktena Evernia Ach., Letharia (Th. Fr.) Zahlbr. Emend DR. Och Usnea Ach. Subgenus Neuropogon (Nees et Flot.) Jatta; Almqvist & Wiksells Boktr, 1926. [Google Scholar]
- Motyka, J. Lichenum Generis Usnea Studium Monographicum. 1936–1938. Pars systematica 1.
- Lamb, I.M. Antarctic Lichens: I. The genera Usnea, Ramalina, Himantormia, Alectoria, Cornicularia. Repub. Br. Antarct. Surv. 1964, 38, 1–34. [Google Scholar]
- Ohmura, Y.; Kanda, H. Taxonomic status of section Neuropogon in the genus Usnea elucidated by morphological comparisons and ITS DNA sequences. Lichenologist 2004, 36, 217–225. [Google Scholar] [CrossRef]
- Lamb, I.M. A Review of the genus Neuropogon (Nees & Flot.) Nyl., with special reference to the Antarctic species. J. Linn. Soc. Lond. Bot. 1939, 52, 199–237. [Google Scholar]
- Lamb, I.M. Further data on the genus Neuropogon. Lilloa 1948, 14, 138–168. [Google Scholar]
- Krog, H. Lethariella and Protousnea, two new lichen genera in Parmeliaceae. Nor. J. Bot. 1976, 3, 83–106. [Google Scholar]
- Krog, H. Evolutionary trends in foliose and fruticose lichens of the Parmeliaceae. J. Hattori Bot. Lab. 1982, 52, 303–311. [Google Scholar]
- Rogers, R.W. Lichens of arid Australia. J. Hattori Bot. Lab. 1982, 53, 351–355. [Google Scholar] [CrossRef]
- Galloway, D.J. New taxa in the New Zealand lichen Flora. N. Z. J. Bot. 1983, 21, 191–199. [Google Scholar] [CrossRef]
- Articus, K. Neuropogon and the phylogeny of Usnea s.l. (Parmeliaceae, Lichenized Ascomycetes). Taxon 2004, 53, 925–934. [Google Scholar] [CrossRef] [Green Version]
- Stevens, G.N. A revision of the lichen family Usneaceae in Australia. Bibl. Lichenol. 1999, 72, 1–28. [Google Scholar]
- Wirtz, N.; Printzen, C.; Sancho, L.G.; Lumbsch, T.H. The phylogeny and classification of Neuropogon and Usnea (Parmeliaceae, Ascomycota) revisited. Taxon 2006, 55, 367–376. [Google Scholar] [CrossRef]
- Clerc, P. Species concepts in the genus Usnea (Lichenized Ascomycetes). Lichenologist 1998, 30, 321–340. [Google Scholar] [CrossRef] [Green Version]
- Wirtz, N.; Printzen, C.; Lumbsch, H.T. The delimitation of antarctic and bipolar species of neuropogonoid Usnea (Ascomycota, Lecanorales): A cohesion approach of species recognition for the Usnea perpusilla complex. Mycol. Res. 2008, 112, 472–484. [Google Scholar] [CrossRef] [PubMed]
- Lumbsch, H.T.; Ahti, T.; Altermann, S.; De Paz, G.A.; Aptroot, A.; Arup, U.; Peña, A.B.; Bawingan, P.A.; Benatti, M.N.; Betancourt, L. One hundred new species of lichenized fungi: A signature of undiscovered global diversity. Phytotaxa 2011, 18, 1–127. [Google Scholar] [CrossRef] [Green Version]
- Grewe, F.; Lagostina, E.; Wu, H.; Printzen, C.; Lumbsch, H.T. Population genomic analyses of RAD sequences resolves the phylogenetic relationship of the lichen-forming fungal species Usnea antarctica and Usnea aurantiacoatra. MycoKeys 2018, 43, 91. [Google Scholar] [CrossRef] [Green Version]
- Culberson, C.F.; Johnson, A. Substitution of methyl tert.-butyl ether for diethyl ether in the standardized thin-layer chromatographic method for lichen products. J. Chromatogr. A 1982, 238, 483–487. [Google Scholar] [CrossRef]
- Arup, U.; Ekman, S.; Lindblom, L.; Mattsson, J.-E. High performance thin layer chromatography (HPTLC), an improved technique for screening lichen substances. Lichenologist 1993, 25, 61–71. [Google Scholar] [CrossRef]
- Lumbsch, H.T. Analysis of phenolic products in lichens for identification and taxonomy. In Protocols in Lichenology; Springer: Berlin/Heidelberg, Germany, 2002; pp. 281–295. [Google Scholar]
- Orange, A.; James, P.W.; White, F.J. Microchemical Methods for the Identification of Lichens, 2nd ed.British Lichen Society: London, UK, 2010. [Google Scholar]
- Elix, J.A.; Wirtz, N.; Lumbsch, H.T. Studies on the chemistry of some Usnea species of the Neuropogon group (Lecanorales, Ascomycota). Nova Hedwig. 2007, 85, 491–502. [Google Scholar] [CrossRef]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef]
- Eaton, D.A.; Overcast, I. Ipyrad: Interactive assembly and analysis of RADseq datasets. Bioinformatics 2020, 36, 2592–2594. [Google Scholar] [CrossRef]
- Kono, M.; Kon, Y.; Ohmura, Y.; Satta, Y.; Terai, Y. In vitro resynthesis of lichenization reveals the genetic background of symbiosis-specific fungal-algal interaction in Usnea hakonensis. BMC Genom. 2020, 21, 671. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A Fast, Scalable and User-Friendly Tool for Maximum Likelihood Phylogenetic Inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aberer, A.J.; Kobert, K.; Stamatakis, A. ExaBayes: Massively parallel bayesian tree inference for the whole-genome era. Mol. Biol. Evol. 2014, 31, 2553–2556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chifman, J.; Kubatko, L. Quartet inference from snp data under the coalescent model. Bioinformatics 2014, 30, 3317–3324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swofford, D.L. Paup 4.0 for Unix/Vms: Phylogenetic Analysis Using Parsimony; Sinauer Associates: Sunderland, MA, USA, 1999; ISBN 0-87893-804-4. [Google Scholar]
- Durand, E.Y.; Patterson, N.; Reich, D.; Slatkin, M. Testing for ancient admixture between closely related populations. Mol. Biol. Evol. 2011, 28, 2239–2252. [Google Scholar] [CrossRef] [Green Version]
- Eaton, D.A.; Ree, R.H. Inferring phylogeny and introgression using RADseq Data: An example from flowering plants (Pedicularis: Orobanchaceae). Syst. Biol. 2013, 62, 689–706. [Google Scholar] [CrossRef] [Green Version]
- Vargas, O.M.; Ortiz, E.M.; Simpson, B.B. Conflicting phylogenomic signals reveal a pattern of reticulate evolution in a recent high-andean diversification (Asteraceae: Astereae: Diplostephium). New Phytol. 2017, 214, 1736–1750. [Google Scholar] [CrossRef]
- Smith, S.A.; O’Meara, B.C. TreePL: Divergence time estimation using penalized likelihood for large phylogenies. Bioinformatics 2012, 28, 2689–2690. [Google Scholar] [CrossRef] [Green Version]
- Sanderson, M.J. R8s: Inferring absolute rates of molecular evolution and divergence times in the absence of a molecular clock. Bioinform. Appl. Note 2003, 19, 301–302. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E. Molecular dating of phylogenies by likelihood methods: A comparison of models and a new information criterion. Mol. Phylogenetics Evol. 2013, 67, 436–444. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Wiens, J.J. Combining phylogenomic and supermatrix approaches, and a time-calibrated phylogeny for squamate reptiles (lizards and snakes) based on 52 genes and 4162 species. Mol. Phylogenetics Evol. 2016, 94, 537–547. [Google Scholar] [CrossRef]
- Zheng, Y.; Wiens, J.J. Do missing data influence the accuracy of divergence-time estimation with BEAST? Mol. Phylogenetics Evol. 2015, 85, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Mazuecos, M.; Vargas, P.; McCauley, R.A.; Monjas, D.; Otero, A.; Chaves, J.A.; Andino, J.E.G.; Rivas-Torres, G. The radiation of Darwin’s giant daisies in the Galapagos Islands. Curr. Biol. 2020, 30, 4989–4998. [Google Scholar] [CrossRef]
- Kraichak, E.; Divakar, P.K.; Crespo, A.; Leavitt, S.D.; Nelsen, M.P.; Lücking, R.; Lumbsch, H.T. A tale of two hyper-diversities: Diversification dynamics of the two largest families of lichenized fungi. Sci. Rep. 2015, 5, 10028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villaverde, T.; Maguilla, E.; Luceño, M.; Hipp, A.L. Assessing the sensitivity of divergence time estimates to locus sampling, calibration points, and model priors in a RAD-seq Phylogeny of Carex Section Schoenoxiphium. J. Syst. Evol. 2021, 59, 687–697. [Google Scholar] [CrossRef]
- Lagostina, E.; Andreev, M.; Dal Grande, F.; Grewe, F.; Lorenz, A.; Lumbsch, H.T.; Rozzi, R.; Ruprecht, U.; Sancho, L.G.; Søchting, U. Effects of dispersal strategy and migration history on genetic diversity and population structure of antarctic lichens. J. Biogeogr. 2021, 48, 1635–1653. [Google Scholar] [CrossRef]
- Shen, X.-X.; Hittinger, C.T.; Rokas, A. Contentious relationships in phylogenomic studies can be driven by a handful of genes. Nat. Ecol. Evol. 2017, 1, 0126. [Google Scholar] [CrossRef] [Green Version]
- MacGuigan, D.J.; Near, T.J. Phylogenomic signatures of ancient introgression in a rogue lineage of darters (Teleostei: Percidae). Syst. Biol. 2019, 68, 329–346. [Google Scholar] [CrossRef]
- Paetzold, C.; Wood, K.R.; Eaton, D.A.; Wagner, W.L.; Appelhans, M.S. Phylogeny of hawaiian Melicope (Rutaceae): RAD-Seq resolves species relationships and reveals ancient introgression. Front. Plant Sci. 2019, 10, 1074. [Google Scholar] [CrossRef] [Green Version]
- Long, C.; Kubatko, L. The effect of gene flow on coalescent-based species-tree inference. Syst. Biol. 2018, 67, 770–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donoghue, M.J.; Eaton, D.A.; Maya-Lastra, C.A.; Landis, M.J.; Sweeney, P.W.; Olson, M.E.; Cachu, N.I.; Moeglein, M.K.; Gardner, J.R.; Heaphy, N.M.; et al. Replicated radiation of a plant clade along a cloud forest archipelago. Nat. Ecol. Evol. 2022, 6, 1318–1329. [Google Scholar] [CrossRef] [PubMed]
- Lamb, I.M. La Vegetación liquénica de los parques nacionales patagónicos. An. Parq. Nac. 1958, 7, 1–188. [Google Scholar]
- Seymour, F.A.; Crittenden, P.D.; Wirtz, N.; Øvstedal, D.O.; Dyer, P.S.; Lumbsch, H.T. Phylogenetic and morphological analysis of antarctic lichen-forming Usnea species in the group Neuropogon. Antarct. Sci. 2007, 19, 71–82. [Google Scholar] [CrossRef]
- Räsänen, V. Zur Kenntnis Der Flechtenflora Feuerlands, Sowie Der Prov. de Magallanes, Prov. de Chiloé Und Prov. de Ñuble in Chile. Ann. Bot. Soc. Zool.-Bot. Fenn. Vanamo 1932, 2, 1–68. [Google Scholar]
- Dodge, C.W. Lichen Flora of the Antarctic Continents and the Adjacent Islands; Phoenix Publishing: Canaan, NH, USA, 1973; ISBN 0-914016-01-6. [Google Scholar]
- Müller-Argoviensis, J. Usnea trachycarpa . Nuovo Giorn. Bot. Ital. 1889, 21, 37. [Google Scholar]
- Galloway, D.J. Flora of New Zealand: Lichens, Including Lichen-Forming and Lichenicolous Fungi, Vol 1 and 2; Manaaki Whenua Press, Landcare Research: Lincoln, New Zealand, 2007; ISBN 0-478-09376-4. [Google Scholar]
- Del-Prado, R.; Blanco, O.; Lumbsch, H.T.; Divakar, P.K.; Elix, J.A.; Molina, M.C.; Crespo, A. Molecular phylogeny and historical biogeography of the lichen-forming fungal genus Flavoparmelia (Ascomycota: Parmeliaceae). Taxon 2013, 62, 928–939. [Google Scholar] [CrossRef] [Green Version]
- Leavitt, S.D.; Esslinger, T.L.; Divakar, P.K.; Lumbsch, H.T. Miocene and pliocene dominated diversification of the lichen-forming fungal genus Melanohalea (Parmeliaceae, Ascomycota) and pleistocene population expansions. BMC Evol. Biol. 2012, 12, 176. [Google Scholar] [CrossRef] [Green Version]
- Divakar, P.K.; Del-Prado, R.; Lumbsch, H.T.; Wedin, M.; Esslinger, T.L.; Leavitt, S.D.; Crespo, A. Diversification of the newly recognized lichen-forming fungal lineage Montanelia (Parmeliaceae, Ascomycota) and its relation to key geological and climatic events. Am. J. Bot. 2012, 99, 2014–2026. [Google Scholar] [CrossRef]
- Leavitt, S.D.; Kirika, P.M.; De Paz, G.A.; Huang, J.-P.; Jae-Seoun, H.U.R.; Grewe, F.; Divakar, P.K.; Lumbsch, H.T. Assessing phylogeny and historical biogeography of the largest genus of lichen-forming fungi, Xanthoparmelia (Parmeliaceae, Ascomycota). Lichenologist 2018, 50, 299–312. [Google Scholar] [CrossRef]
- Divakar, P.K.; Crespo, A.; Kraichak, E.; Leavitt, S.D.; Singh, G.; Schmitt, I.; Lumbsch, H.T. Using a temporal phylogenetic method to harmonize family-and genus-level classification in the largest clade of lichen-forming fungi. Fungal Divers. 2017, 84, 101–117. [Google Scholar] [CrossRef]
- Zachos, J.C.; Dickens, G.R.; Zeebe, R.E. An early cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 2008, 451, 279–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sepulchre, P.; Ramstein, G.; Fluteau, F.; Schuster, M.; Tiercelin, J.-J.; Brunet, M. Tectonic uplift and eastern Africa aridification. Science 2006, 313, 1419–1423. [Google Scholar] [CrossRef] [Green Version]
- Flower, B.P.; Kennett, J.P. The middle miocene climatic transition: East antarctic ice sheet development, deep ocean circulation and global carbon cycling. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994, 108, 537–555. [Google Scholar] [CrossRef]
- Janis, C.M. Tertiary mammal evolution in the context of changing climates, vegetation, and tectonic events. Annu. Rev. Ecol. Syst. 1993, 24, 467–500. [Google Scholar] [CrossRef]
- Hinojosa, L.F.; Villagrán, C. Historia de los bosques del sur de Sudamérica, I: Antecedentes paleobotánicos, geológicos y climáticos del terciario del cono sur de América. Rev. Chil. Hist. Nat. 1997, 70, 225–239. [Google Scholar]
- Galloway, D.J. Plate tectonics and the distribution of cool temperate southern hemisphere macrolichens. Bot. J. Linn. Soc. 1988, 96, 45–55. [Google Scholar] [CrossRef]
- Nylinder, S.; Swenson, U.; Persson, C.; Janssens, S.B.; Oxelman, B. A dated species–tree approach to the trans–pacific disjunction of the genus Jovellana (Calceolariaceae, Lamiales). Taxon 2012, 61, 381–391. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Phylogenetic Clade | Species | Hypostrepsilic Acid | Fumarprotocetraric Acid | Psoromic Acid | Salazinic Acid | Neuropogolic Acid |
|---|---|---|---|---|---|---|
| A | U. ciliata | ± | ± | + | ||
| A | U. subcapillaris | ± | ± | + | ||
| A | Undescribed taxon | + | + | |||
| A | U. pseudocapillaris | ± | + | |||
| B | U. rohmederi | |||||
| C | U. perpusilla | ± | ||||
| D | U. aymondiana | ± | ± | |||
| D | U. fibriloides | + | ||||
| D | U. sphacelata | ± | ||||
| D | U. subantarctica | + | ||||
| D | U. trachycarpa | ± | ± | ± | ± | |
| D | U. trachycarpoides | + | ||||
| D | U. hyyppae | ± |
| Dataset | N Samples | N Loci Filtered | Mean Loci per Sample | SD Loci per Sample | Min Loci | Max Loci | N snps | % Missing Data |
|---|---|---|---|---|---|---|---|---|
| m4 | 126 | 21,831 | 5917.376 | 3619.83 | 938 | 15,073 | 381,659 | 82.91 |
| m25 | 126 | 8698 | 4705.896 | 2340.84 | 804 | 8182 | 271,485 | 76.22 |
| m50 | 126 | 4504 | 3180.304 | 1141.65 | 706 | 4442 | 166,242 | 72.411 |
| Species | Node Age | BV |
|---|---|---|
| U. acromelana | 5.56 | 3.64–6.89 |
| U. antarctica | 1.8 | 0.49–3.09 |
| U. aurantiacoatra | 1.8 | 0.49–3.09 |
| U. aymondiana | 6.13 | 5.31–7.23 |
| U. ciliata | 2.75 | 1.14–5.91 |
| U. fibriloides | 5.09 | 3.62–6.58 |
| U. hyyppae | 1.36 | 0.43–2.27 |
| U. lambii | 5.75 | 2.66–7.72 |
| U. messutiae | 2.60 | 1–4.93 |
| U. pallidocarpa | 2.60 | 1–4.93 |
| U. patagonica | 7.3 | 4.91–8.54 |
| U. perpusilla | 5.45 | 2.79–7.34 |
| U. pseudocapillaris | 2.42 | 0.88–5.22 |
| U. rohmederi | 2.68 | 1.19–5.81 |
| U. sphacelata | 2.01 | 0.95–3.77 |
| U. subantarctica | 2.75 | 0.95–4.25 |
| U. subcapillaris | 2.75 | 1.14–5.91 |
| U. taylorii | 3.51 | 1.69–5.58 |
| U. trachycarpa | 2.01 | 0.95–3.77 |
| U. trachycarpoides | 1.36 | 0.43–2.27 |
| U. ushuaiensis | 2.68 | 1.19–5.81 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otero, A.; Barcenas-Peña, A.; Lumbsch, H.T.; Grewe, F. Reference-Based RADseq Unravels the Evolutionary History of Polar Species in ‘the Crux Lichenologorum’ Genus Usnea (Parmeliaceae, Ascomycota). J. Fungi 2023, 9, 99. https://doi.org/10.3390/jof9010099
Otero A, Barcenas-Peña A, Lumbsch HT, Grewe F. Reference-Based RADseq Unravels the Evolutionary History of Polar Species in ‘the Crux Lichenologorum’ Genus Usnea (Parmeliaceae, Ascomycota). Journal of Fungi. 2023; 9(1):99. https://doi.org/10.3390/jof9010099
Chicago/Turabian StyleOtero, Ana, Alejandrina Barcenas-Peña, H. Thorsten Lumbsch, and Felix Grewe. 2023. "Reference-Based RADseq Unravels the Evolutionary History of Polar Species in ‘the Crux Lichenologorum’ Genus Usnea (Parmeliaceae, Ascomycota)" Journal of Fungi 9, no. 1: 99. https://doi.org/10.3390/jof9010099
APA StyleOtero, A., Barcenas-Peña, A., Lumbsch, H. T., & Grewe, F. (2023). Reference-Based RADseq Unravels the Evolutionary History of Polar Species in ‘the Crux Lichenologorum’ Genus Usnea (Parmeliaceae, Ascomycota). Journal of Fungi, 9(1), 99. https://doi.org/10.3390/jof9010099