
The Lichen Genus Sticta (Lobariaceae, Peltigerales) in East African Montane Ecosystems
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Locations and Sampling
2.2. Morphological Inspection
2.3. DNA Sequencing
2.4. Phylogenetic Analyses
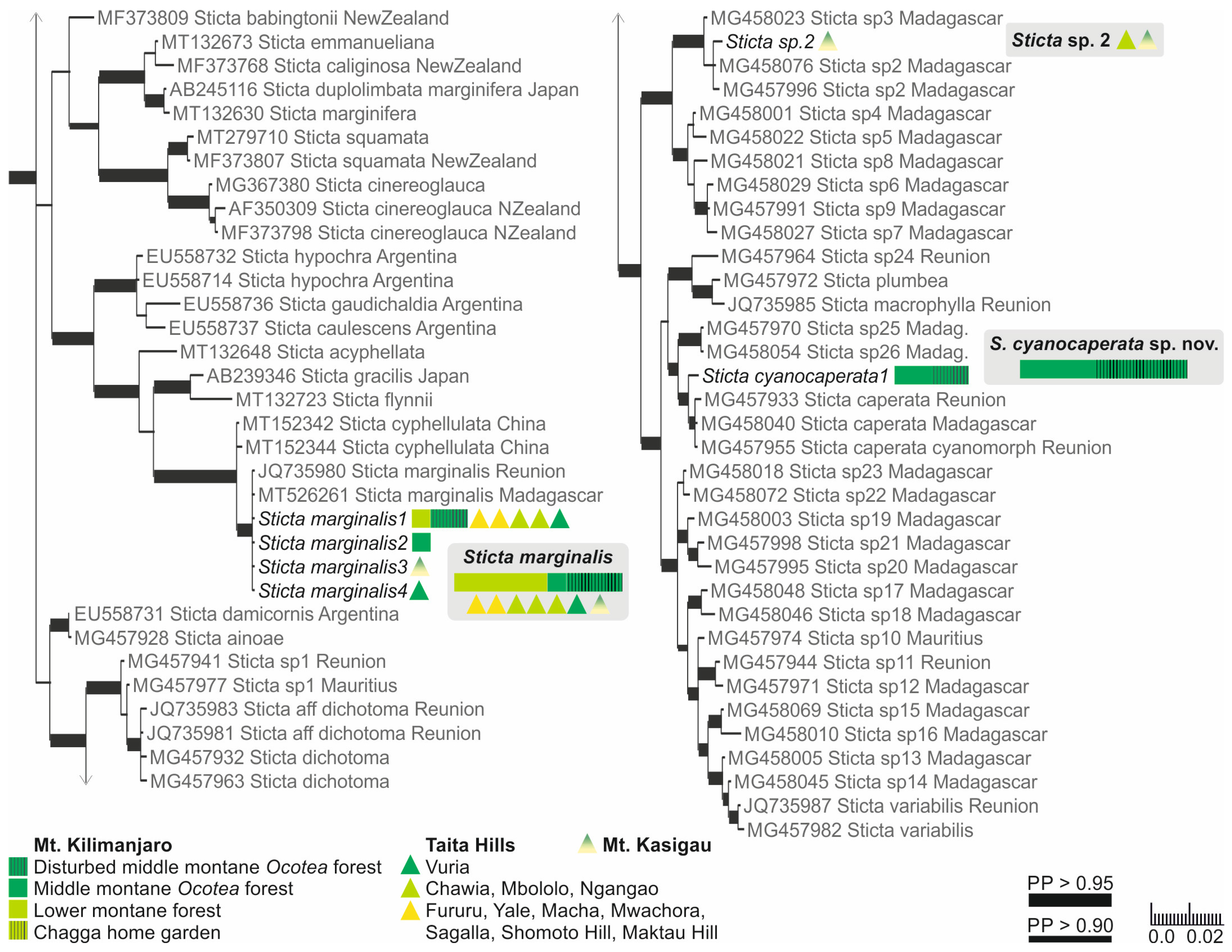
3. Results
3.1. Phylogenetic Analyses of the Specimens
3.1.1. Specimens with Soredia or Pustules
3.1.2. Fertile Specimens without Symbiotic Propagules
3.1.3. Specimens with Laminal Isidia (Sticta fuliginosa Morphodeme)
3.1.4. Specimens of the Sticta weigelii Morphodeme
3.1.5. Specimens with Marginal Isidia
3.1.6. Sticta with Green Algae
3.2. Species of Sticta Identified
3.2.1. Key to Sticta Species in East Africa
- 1a.
- Main photobiont green alga—2.
- 1b.
- Main photobiont cyanobacterium—3.
- 2a.
- Thallus thin and fragile, with marginal lobules—Sticta sp. 2 (Figure 6a)
- 2b.
- Thallus robust, without marginal lobules—Sticta dichotoma
- 3a.
- Thallus with marginal to submarginal soralia (may occasionally form coralloid, isidia-resembling structures) and/or pustules—4.
- 3b.
- Thallus without soredia or pustules—5.
- 4a.
- With true, mainly marginal soredia (found in sub/lower montane habitats, mainly <2000 masl)—Sticta sublimbata
- 4b.
- With pustules that may appear sorediate (found in upper montane and subalpine habitats, >3500 masl)—Sticta umbilicariiformis
- 5a.
- Thallus without symbiotic propagules, often fertile—6.
- 5b.
- Thallus with marginal or laminal isidia and/or phyllidia—9.
- 6a.
- Apothecial margin with abundant white hairs, not nodulous; lobe margins often with projecting tufts of hair/tomentum; cyphellae urceolate with a small pore; ascospores large (>40 × 8 µm in diam.)—Sticta tomentosa
- 6b.
- Apothecial margin slightly pubescent at most, most often nodulous; cyphellae not strongly urceolate with a small pore; ascospores shorter—7.
- 7a.
- Ascospores 3-septate; lobes usually <3 cm long and wide—Sticta munda sp. nov.
- 7b.
- Ascospores 1-septate; lobes often larger—8.
- 8a.
- Ascospores 33–40 µm long—Sticta umbilicariiformis
- 8b.
- Ascospores usually shorter—Sticta aspratilis sp. nov.
- 9a.
- Dark isidia marginal, submarginal, and on scrobiculate ridges present at least near the thallus margins of the brown thallus—Sticta cellulosa sp. nov.
- 9b.
- Thallus without scrobiculate isidiate ridges—10.
- 10a.
- Isidia scattered over thallus lamina; without apothecia—11.
- 10b.
- Isidia predominantly on thallus margins (in fertile specimens often also on lamina)—14.
- 11a.
- Lobes elongate-obovate, fan-shaped, ascending from one attachment point with a funnel-like base; older thalli commonly with stalked lobules; lower side with pronounced, often clearly dome-like cyphellae (in middle montane to subalpine habitats, >2500 masl)—Sticta fuliginoides agg. (Sticta fuliginoides and Sticta sp. A)
- 11b.
- Lobes rounded and/or palmate, without a funnel-shaped base, stalked lobules not present—12.
- 12a.
- Thallus small (up to 2 cm in diam.), lobes widely rounded (wider than long) and revolute (often complete lobes becoming convex); lower side marginally etomentose with widely different sized and often flat cyphellae (in lower and middle montane habitats, <2600 masl)—Sticta ciliata agg. (Sticta ciliata and Sticta sp. B, Figure 6b)
- 12b.
- Thallus usually larger, lower side fully tomentose, cyphellae cupuliform to slightly urceolate—13.
- 13a.
- Lower side usually with abundant, arachnoid, moniliform secondary tomentum—Sticta aspratilis sp. nov.
- 13b.
- Lower side usually without arachnoid, moniliform secondary tomentum—Sticta fuliginosa
- 14a.
- Lobes palmate, clearly stipitate and ascending from one attachment point; lower side largely etomentose, often with a yellow hue; marginal isidia developing into lobules especially in older thalli—Sticta marginalis
- 14b.
- Lobes not clearly stipitate, palmate, and ascending; lower side tomentose—15.
- 15a.
- Lobes elongate, with rounded apices, often with dark marginal cilia; cyphellae distinctly raised and urceolate with a small opening (dome-like)—Sticta duplolimbata
- 15b.
- Lobe margins without dark marginal cilia; cyphellae not distinctly dome-like—16.
- 16a.
- Isidia cylindrical to coralloid, in congested dark heaps, occasionally developing into stalked lobules—Sticta sp. C (weigelii agg.) (Figure 6c)
- 16b.
- Isidia flattened and extending horizontally from the margins, usually not in congested heaps—17.
- 17a.
- Lower surface color is usually cream to light brown with light to brown tomentum (or lower surface brown with white tomentum); upper surface grey to chocolate brown—18.
- 17b.
- Lower surface dark brown (at least centrally) with dark tomentum; upper surface color is usually fawn to yellow–brown (occasionally light grey or dark brown)—19.
- 18a.
- Thallus thick and large; with marginal flattened isidia; primary tomentum of agglutinated hyphae—Sticta afromontana sp. nov.
- 18b.
- Rounded lobes with marginal and submarginal isidia and stalked lobules; lower surface at least centrally dark brown and glossy, tomentum white-light, only weakly agglutinated and hair-like, often entangled—Sticta sp. D
- 19a.
- K+ yellow; with moniliform secondary tomentum—Sticta andina
- 19b.
- K−; without moniliform secondary tomentum—Sticta cyanocaperata sp. nov.

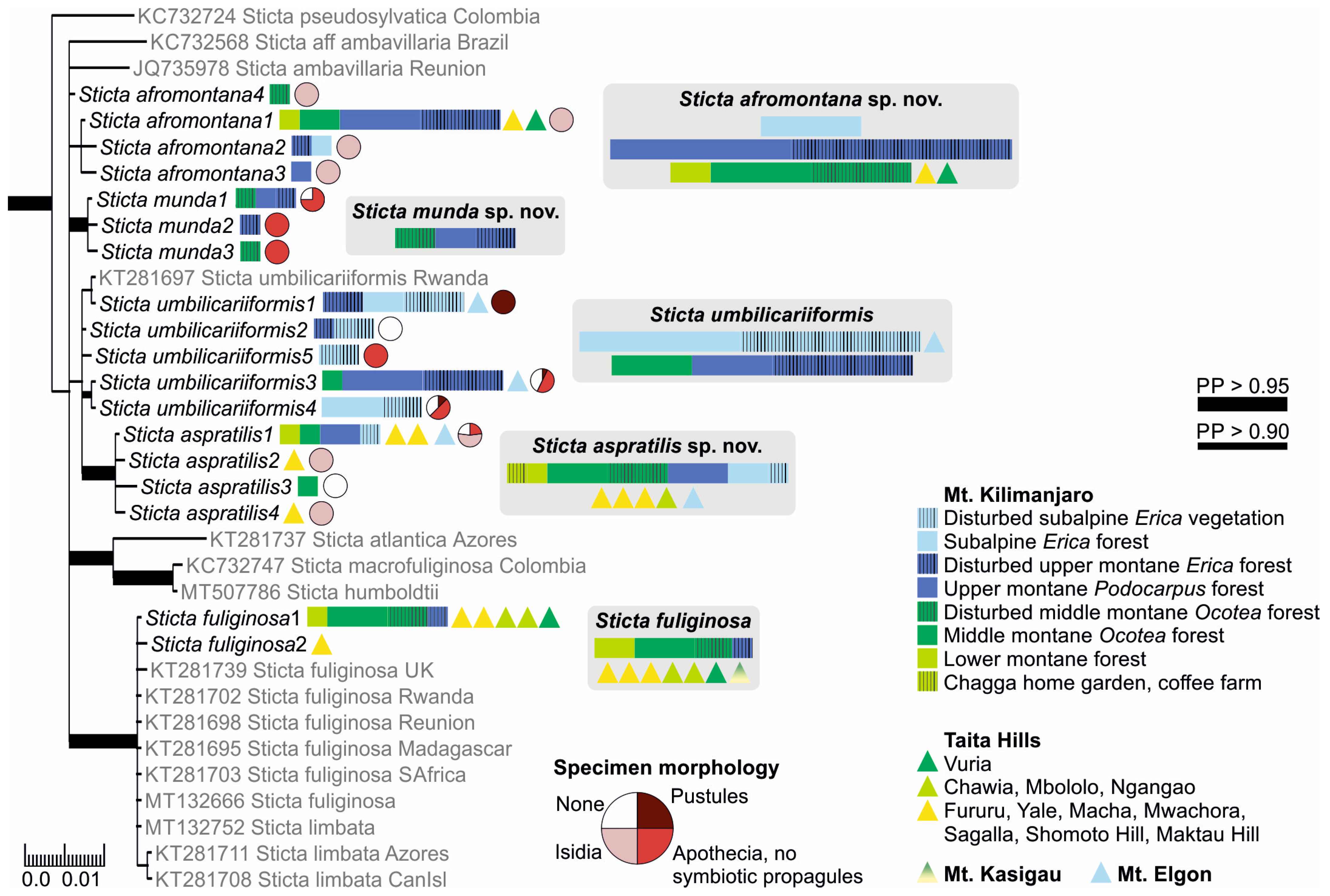
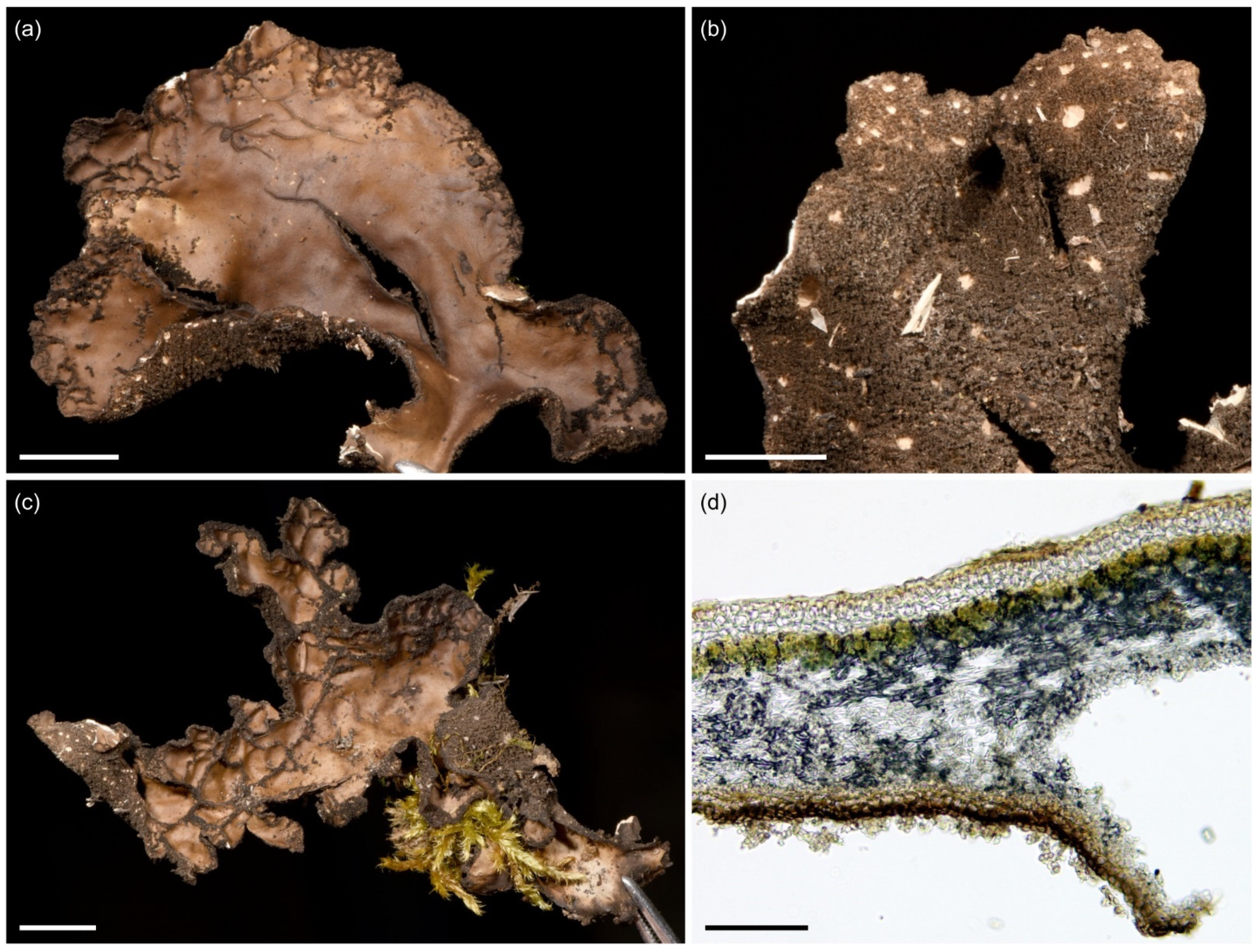
3.2.2. Sticta afromontana Kaasalainen and Rikkinen sp. nov. (Figure 7)

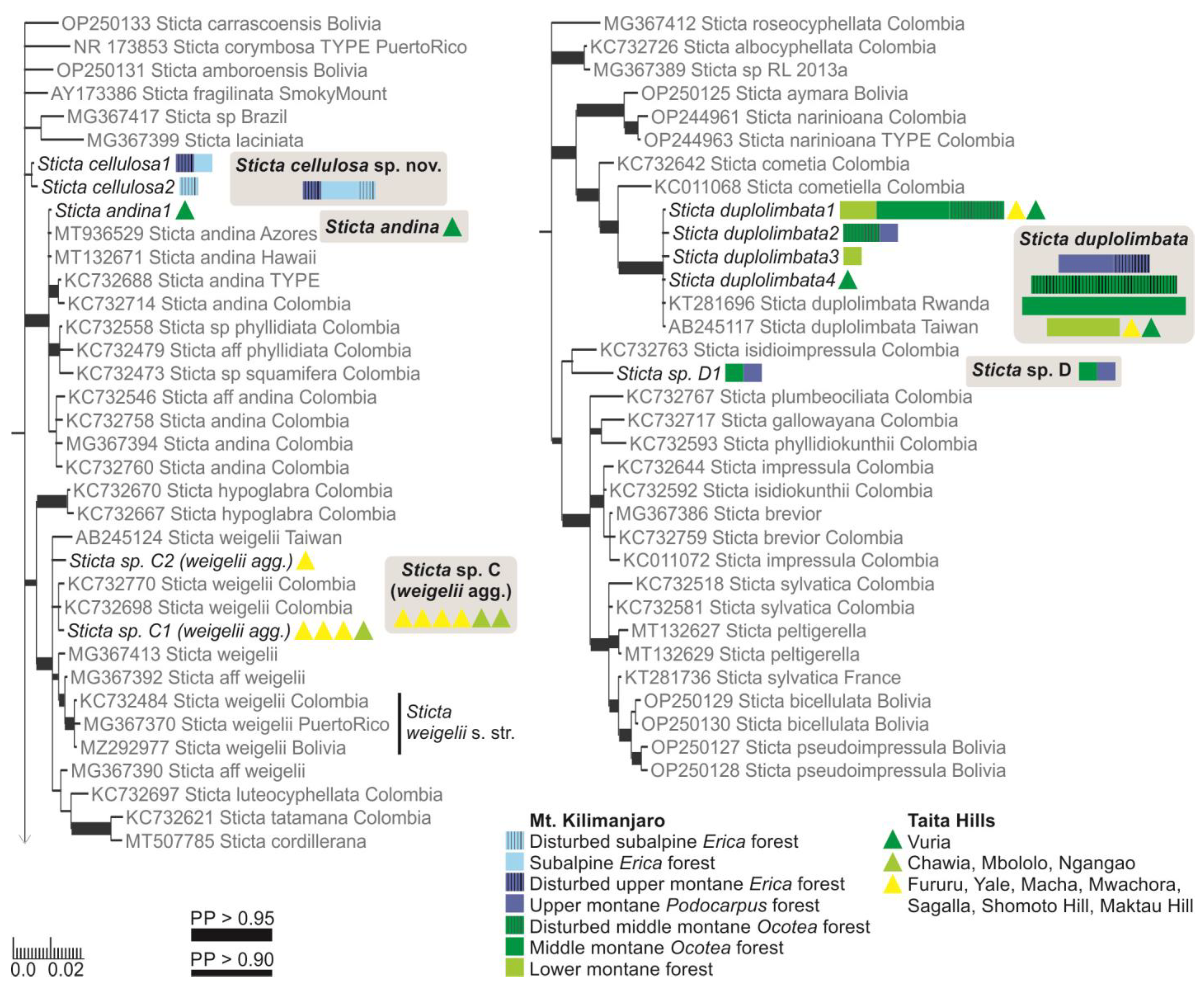
3.2.3. Sticta andina B. Moncada, Lücking and Sérus. (Figure 6d,e)
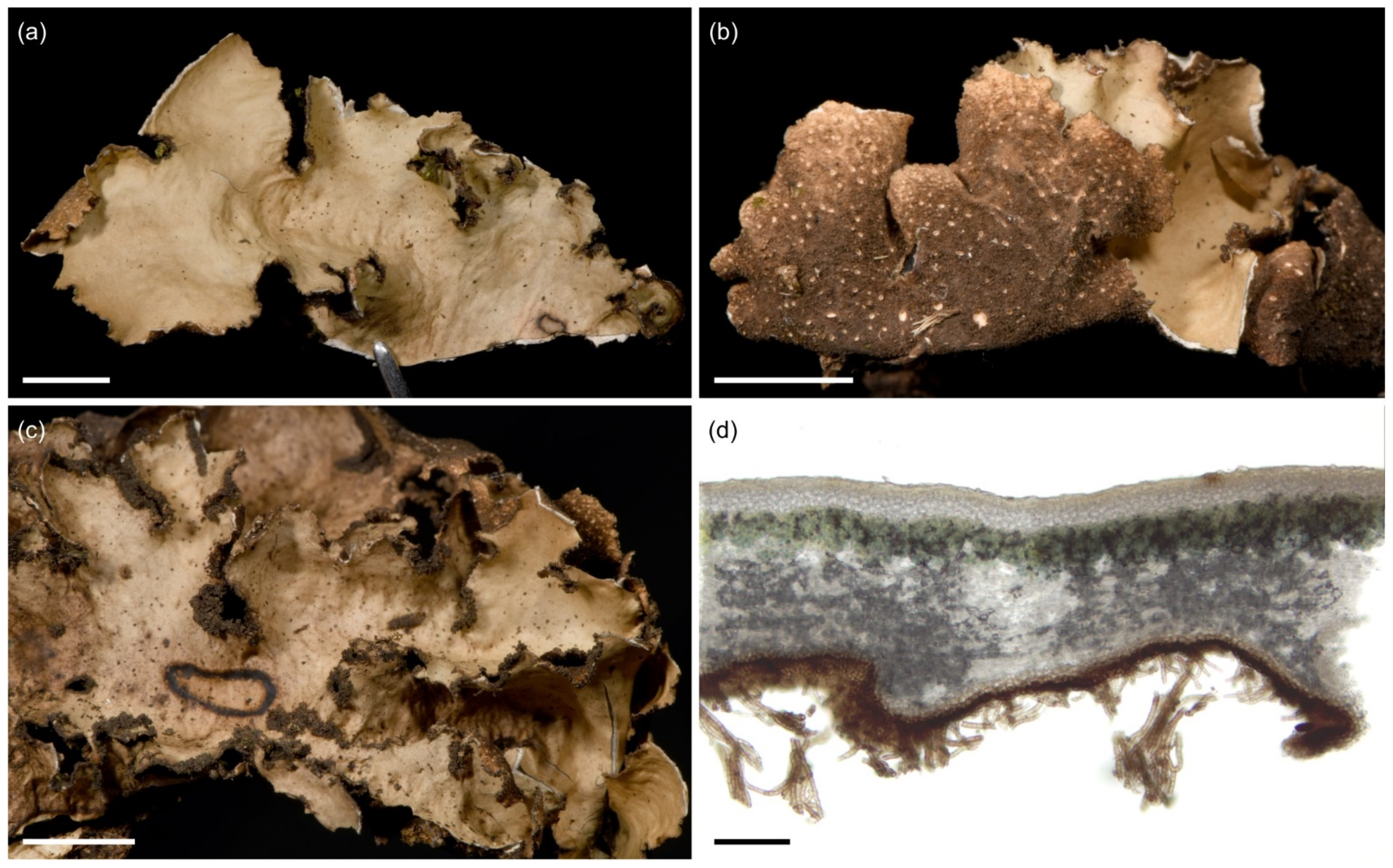
3.2.4. Sticta aspratilis Kaasalainen and Rikkinen sp. nov. (Figure 8)

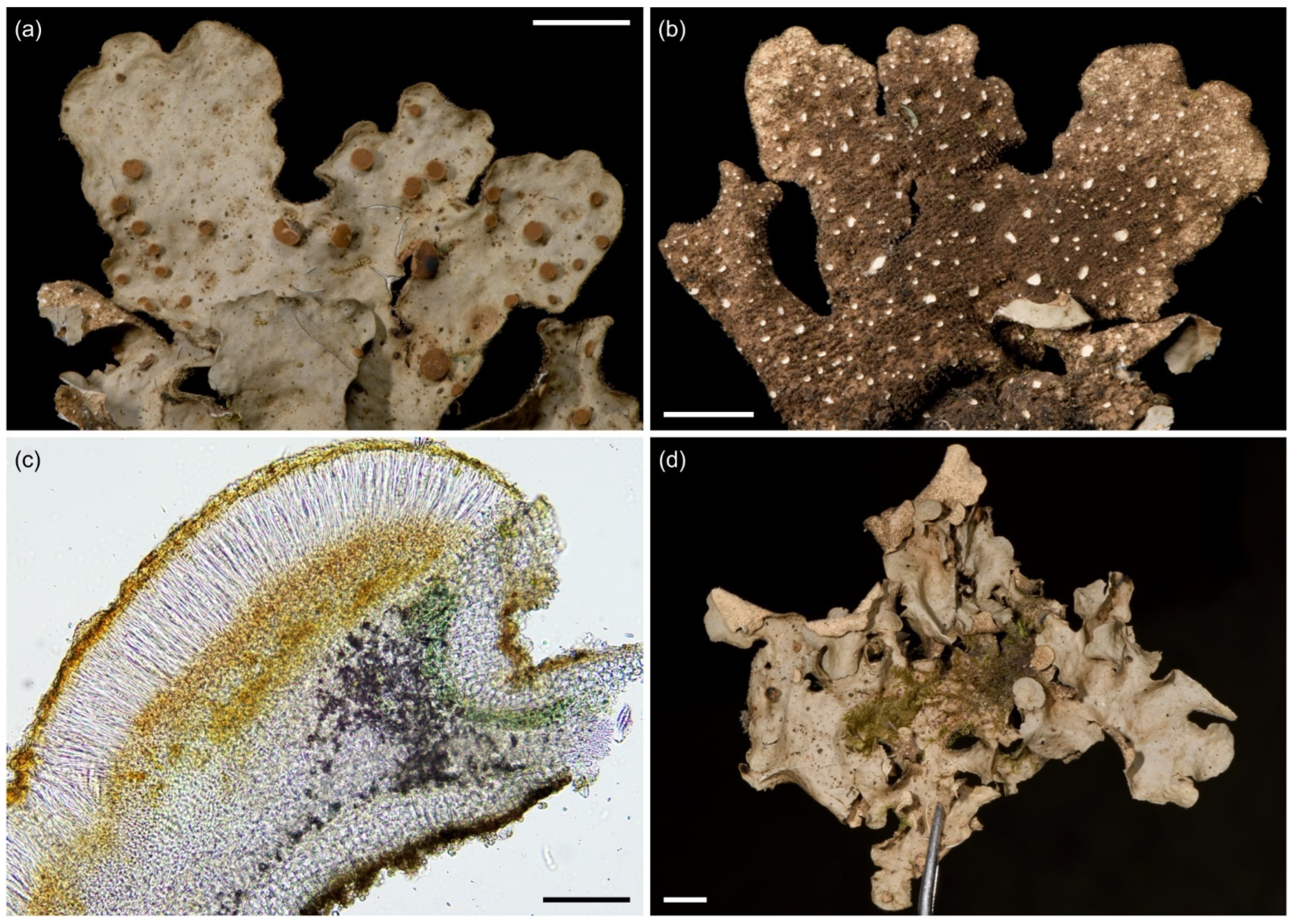
3.2.5. Sticta cellulosa Kaasalainen sp. nov. (Figure 9)

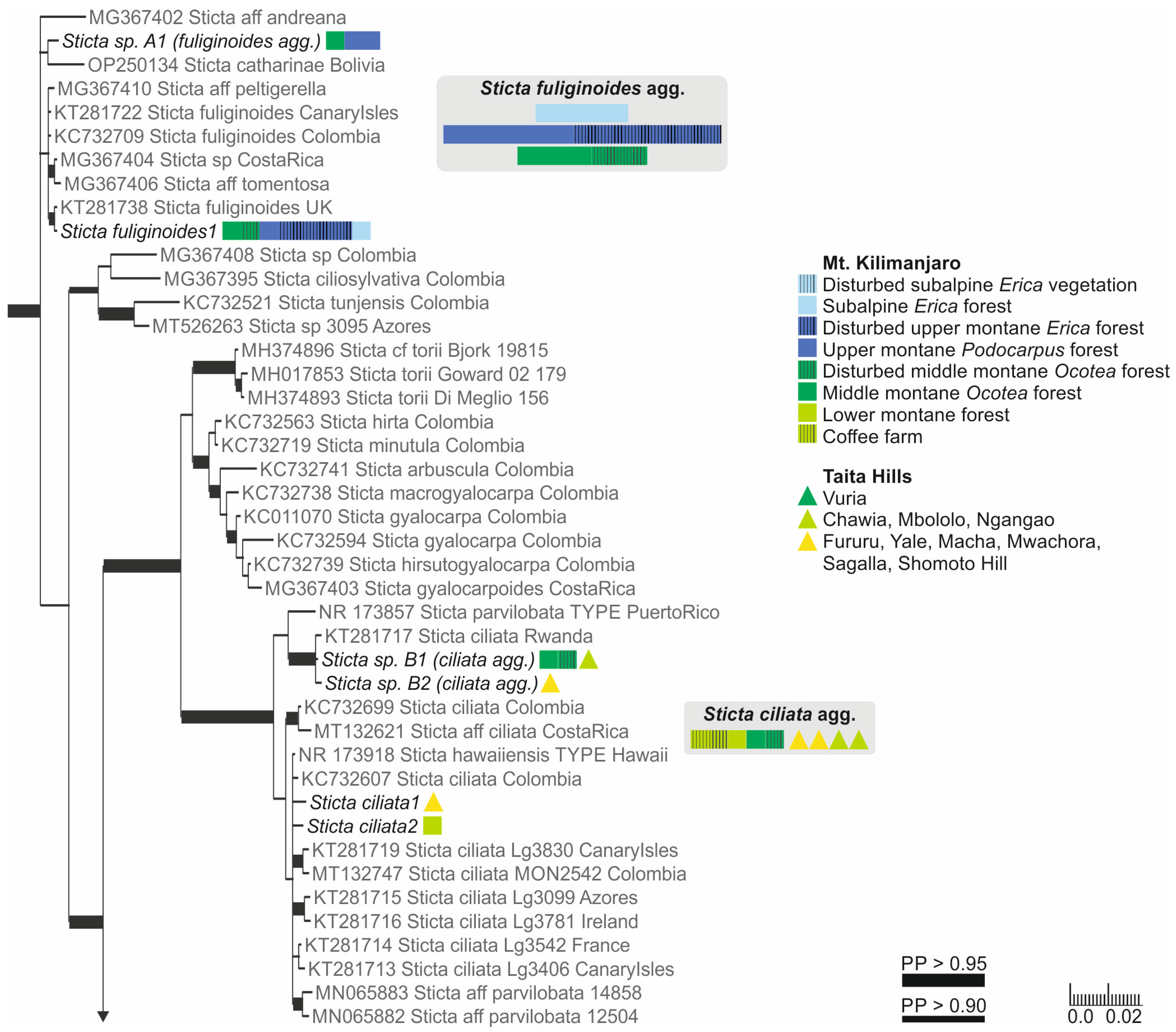
3.2.6. Sticta ciliata Tayl. (Figure 10a)

3.2.7. Sticta cyanocaperata Kaasalainen sp. nov. (Figure 11)

3.2.8. Sticta duplolimbata (Hue) Vain. (Figure 10b)
3.2.9. Sticta fuliginoides Magain and Sérus. (Figure 10c)
3.2.10. Sticta fuliginosa (Hoffm.) Ach. (Figure 10d,e)
3.2.11. Sticta marginalis Bory (Figure 10f–h)
3.2.12. Sticta munda Kaasalainen sp. nov. (Figure 12)

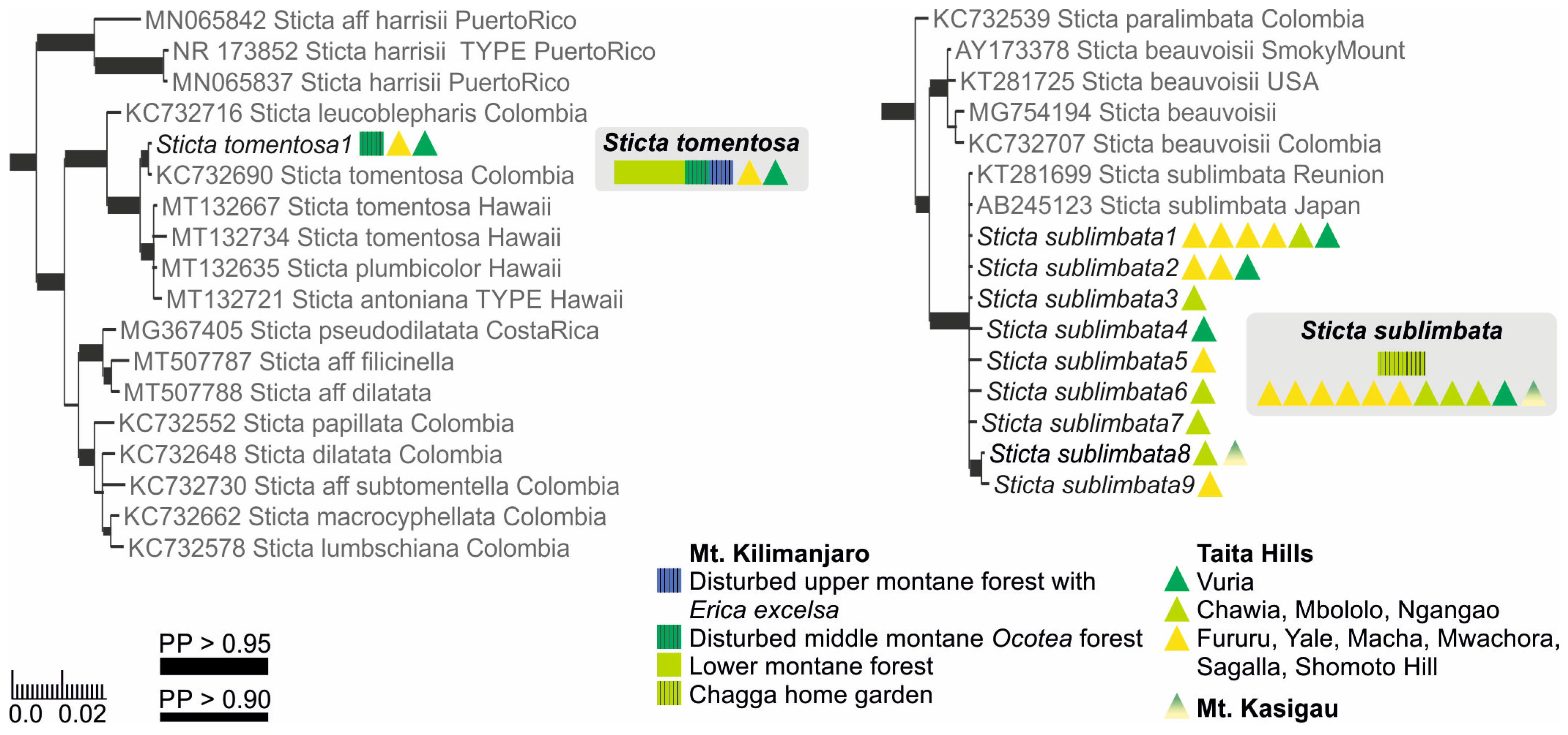
3.2.13. Sticta sublimbata (J. Steiner) Swinscow and Krog (Figure 13a,b)

3.2.14. Sticta tomentosa (Sw.) Ach. (Figure 13c,d)
3.2.15. Sticta umbilicariiformis Hochst. ex Flot. (Figure 13e–g)
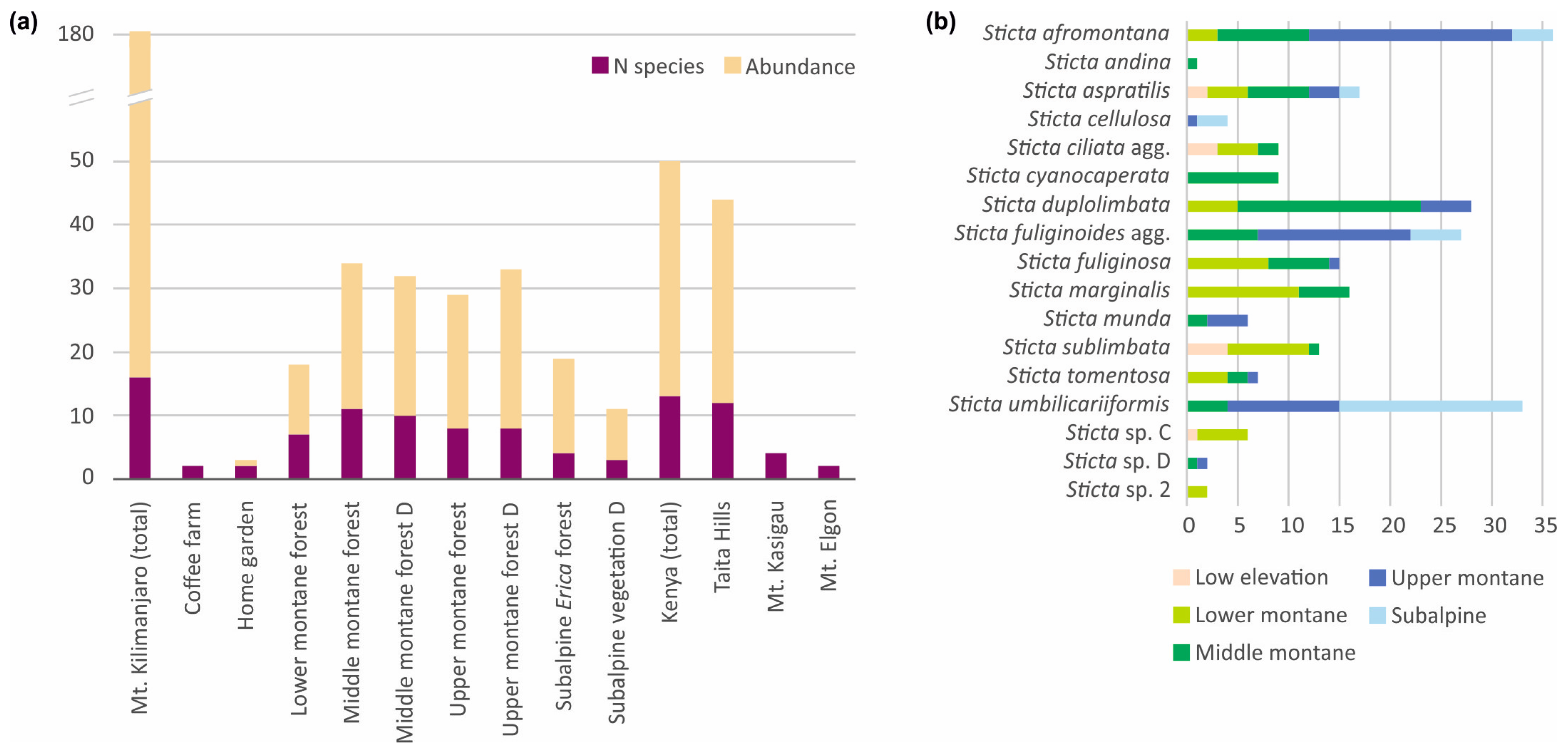
3.3. Sticta Diversity in the Montane Ecosystems of East Africa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Collection ID | ITS Variant | Collection Location | Accession | |
|---|---|---|---|---|---|
| Sticta afromontana | JR10112 | afromontana1 | Taita Hills | Vuria | OP999402 |
| Sticta afromontana | JR10121B | afromontana1 | Taita Hills | Vuria | OP999404 |
| Sticta afromontana | JR10189 | afromontana1 | Taita Hills | Yale | OP999412 |
| Sticta afromontana | JR16366 | afromontana1 | Taita Hills | Vuria | OP999453 |
| Sticta afromontana | JR25 | - | Taita Hills | Yale | - |
| Sticta afromontana | JR31 | - | Taita Hills | Vuria | - |
| Sticta afromontana | UK110523c | - | Taita Hills | Vuria | - |
| Sticta afromontana | UK170771f | afromontana1 | Kilimanjaro | FPo1 | OP999477 |
| Sticta afromontana | UK170775b | afromontana3 | Kilimanjaro | FPo1 | OP999480 |
| Sticta afromontana | UK170779c | afromontana1 | Kilimanjaro | FPo1 | OP999481 |
| Sticta afromontana | UK170781b | afromontana1 | Kilimanjaro | FPo1 | OP999482 |
| Sticta afromontana | UK170792d | afromontana1 | Kilimanjaro | FOc2 | OP999484 |
| Sticta afromontana | UK170794d | - | Kilimanjaro | FOc2 | - |
| Sticta afromontana | UK170800b | - | Kilimanjaro | FOc1 | - |
| Sticta afromontana | UK170806j | afromontana1 | Kilimanjaro | FPD4 | OP999491 |
| Sticta afromontana | UK170807a | - | Kilimanjaro | FPD4 | - |
| Sticta afromontana | UK170817h | - | Kilimanjaro | FPD4 | - |
| Sticta afromontana | UK170822k | - | Kilimanjaro | FPD4 | - |
| Sticta afromontana | UK170826e | afromontana1 | Kilimanjaro | FPD4 | OP999496 |
| Sticta afromontana | UK170830d | - | Kilimanjaro | FPD4 | - |
| Sticta afromontana | UK170832c | - | Kilimanjaro | FPD4 | - |
| Sticta afromontana | UK170834a | - | Kilimanjaro | FPD3 | - |
| Sticta afromontana | UK170836b | - | Kilimanjaro | FPD3 | - |
| Sticta afromontana | UK170841b | - | Kilimanjaro | FPD3 | - |
| Sticta afromontana | UK170842e | afromontana1 | Kilimanjaro | FPD3 | OP999498 |
| Sticta afromontana | UK170846e | afromontana1 | Kilimanjaro | FPD3 | OP999501 |
| Sticta afromontana | UK170856c | - | Kilimanjaro | FPo4 | - |
| Sticta afromontana | UK170857a | afromontana1 | Kilimanjaro | FPo4 | OP999503 |
| Sticta afromontana | UK170858g | afromontana1 | Kilimanjaro | FPo4 | OP999507 |
| Sticta afromontana | UK170880e | - | Kilimanjaro | FPo5 | - |
| Sticta afromontana | UK170886a | afromontana1 | Kilimanjaro | FPo5 | OP999509 |
| Sticta afromontana | UK170892b | - | Kilimanjaro | FPo5 | - |
| Sticta afromontana | UK170897ar | - | Kilimanjaro | FOD4 | - |
| Sticta afromontana | UK170911b | - | Kilimanjaro | FOD5 | - |
| Sticta afromontana | UK170915a | - | Kilimanjaro | FOD5 | - |
| Sticta afromontana | UK170930c | afromontana1 | Kilimanjaro | Flm1 | OP999523 |
| Sticta afromontana | UK170951g | - | Kilimanjaro | Flm1 | - |
| Sticta afromontana | UK171180a | afromontana4 | Kilimanjaro | FOD3 | OP999537 |
| Sticta afromontana | UK171185h | - | Kilimanjaro | FOD3 | - |
| Sticta afromontana | UK171405e | - | Kilimanjaro | FEr4 | - |
| Sticta afromontana | UK171413a | - | Kilimanjaro | FEr4 | - |
| Sticta afromontana | UK171438g | - | Kilimanjaro | FEr2 | - |
| Sticta afromontana | UK171439t | afromontana2 | Kilimanjaro | FEr2 | OP999554 |
| Sticta afromontana | UK171442c | - | Kilimanjaro | FEr2 | - |
| Sticta afromontana | UK171445j | - | Kilimanjaro | FEr2 | - |
| Sticta afromontana | UK171470v | - | Kilimanjaro | FPD1 | - |
| Sticta afromontana | UK171471c | - | Kilimanjaro | FPD1 | - |
| Sticta afromontana | UK171476e | afromontana1 | Kilimanjaro | FPD1 | OP999561 |
| Sticta afromontana | UK171490e | afromontana1 | Kilimanjaro | FOc3 | OP999566 |
| Sticta afromontana | UK171525a | - | Kilimanjaro | FPo2 | - |
| Sticta afromontana | UK171526a | afromontana1 | Kilimanjaro | FPo2 | OP999590 |
| Sticta afromontana | UK171528b | afromontana1_4 | Kilimanjaro | FPo2 | OP999591 |
| Sticta afromontana | UK171577f | afromontana2 | Kilimanjaro | FPD2 | OP999595 |
| Sticta afromontana | UK171578m | - | Kilimanjaro | FPD2 | - |
| Sticta afromontana | UK171581j | - | Kilimanjaro | FPD2 | - |
| Sticta afromontana | UK171582p | - | Kilimanjaro | FPD2 | - |
| Sticta afromontana | UK171584v | afromontana1 | Kilimanjaro | FPD2 | OP999601 |
| Sticta afromontana | UK171589b | - | Kilimanjaro | FPo3 | - |
| Sticta andina | JR10117 | andina1 | Taita Hills | Vuria | OP999403 |
| Sticta aspratilis | JR10044A | aspratilis4 | Taita Hills | Shomoto Hill | OP999394 |
| Sticta aspratilis | JR10057 | aspratilis1 | Taita Hills | Shomoto Hill | OP999395 |
| Sticta aspratilis | JR10155A | aspratilis1 | Taita Hills | Yale | OP999406 |
| Sticta aspratilis | JR10155B | aspratilis1 | Taita Hills | Yale | OP999407 |
| Sticta aspratilis | JR10155C | aspratilis1 | Taita Hills | Yale | OP999408 |
| Sticta aspratilis | JR10171 | aspratilis1 | Taita Hills | Yale | OP999409 |
| Sticta aspratilis | JR16103 | aspratilis1 | Mt. Elgon | Erica zone | OP999436 |
| Sticta aspratilis | JR16107 | aspratilis1 | Mt. Elgon | Erica zone | OP999437 |
| Sticta aspratilis | UK110551f | aspratilis2 | Taita Hills | Fururu | OP999475 |
| Sticta aspratilis | UK110551g | aspratilis2 | Taita Hills | Fururu | OP999476 |
| Sticta aspratilis | UK110566c | - | Taita Hills | Chawia | - |
| Sticta aspratilis | UK170781c | - | Kilimanjaro | FPo1 | - |
| Sticta aspratilis | UK170892c | aspratilis1 | Kilimanjaro | FPo5 | OP999510 |
| Sticta aspratilis | UK170896b | aspratilis1and2 | Kilimanjaro | FPo5 | OP999512 |
| Sticta aspratilis | UK170910b | - | Kilimanjaro | FOD5 | - |
| Sticta aspratilis | UK170912b | - | Kilimanjaro | FOD5 | - |
| Sticta aspratilis | UK170916c | aspratilis1 | Kilimanjaro | Flm6 | OP999520 |
| Sticta aspratilis | UK170975f | aspratilis1 | Kilimanjaro | Cof5 | OP999530 |
| Sticta aspratilis | UK171478a | aspratilis1 | Kilimanjaro | FOc4 | OP999562 |
| Sticta aspratilis | UK171487c | aspratilis1 | Kilimanjaro | FOc4 | OP999565 |
| Sticta aspratilis | UK171508k | - | Kilimanjaro | FOD2 | - |
| Sticta aspratilis | UK171515b | aspratilis3 | Kilimanjaro | FOc5 | OP999584 |
| Sticta aspratilis | UK171562c | aspratilis1 | Kilimanjaro | FED2 | OP999592 |
| Sticta aspratilis | UK171587j | aspratilis1 | Kilimanjaro | FPo3 | OP999607 |
| Sticta cellulosa | UK171340t | cellulosa2 | Kilimanjaro | FED1 | OP999545 |
| Sticta cellulosa | UK171406e | cellulosa1 | Kilimanjaro | FEr4 | OP999548 |
| Sticta cellulosa | UK171407j | cellulosa1 | Kilimanjaro | FEr4 | OP999549 |
| Sticta cellulosa | UK171458k | - | Kilimanjaro | FEr4 | - |
| Sticta cellulosa | UK171584k | cellulosa1 | Kilimanjaro | FPD2 | OP999599 |
| Sticta ciliata | JR16294b | ciliata1 | Taita Hills | Sagalla | OP999443 |
| Sticta ciliata | UK170944j | ciliata2 | Kilimanjaro | Flm1 | OP999528 |
| Sticta ciliata (agg.) | UK110570a | - | Taita Hills | Chawia | - |
| Sticta ciliata (agg.) | UK160552r | - | Kilimanjaro | Home3 | - |
| Sticta ciliata (agg.) | UK170984 | - | Kilimanjaro | Cof5 | - |
| Sticta cyanocaperata | UK170897f | - | Kilimanjaro | FOD4 | - |
| Sticta cyanocaperata | UK170912a | cyanocaperata1 | Kilimanjaro | FOD5 | OP999516 |
| Sticta cyanocaperata | UK170912e | cyanocaperata1 | Kilimanjaro | FOD5 | OP999518 |
| Sticta cyanocaperata | UK171182c | cyanocaperata1 | Kilimanjaro | FOD3 | OP999539 |
| Sticta cyanocaperata | UK171480d | cyanocaperata1 | Kilimanjaro | FOc4 | OP999563 |
| Sticta cyanocaperata | UK171483v | - | Kilimanjaro | FOc4 | - |
| Sticta cyanocaperata | UK171486b | - | Kilimanjaro | FOc4 | - |
| Sticta cyanocaperata | UK171495f | cyanocaperata1 | Kilimanjaro | FOc3 | OP999570 |
| Sticta cyanocaperata | UK171506g | - | Kilimanjaro | FOD2 | - |
| Sticta cyanocaperata | UK171510m | - | Kilimanjaro | FOD2 | - |
| Sticta duplolimbata | JR10194A | duplolimbata1 | Taita Hills | Yale | OP999416 |
| Sticta duplolimbata | JR10194B | - | Taita Hills | Yale | - |
| Sticta duplolimbata | JR16374b | duplolimbata4 | Taita Hills | Vuria | OP999460 |
| Sticta duplolimbata | JR16374d | duplolimbata1 | Taita Hills | Vuria | OP999462 |
| Sticta duplolimbata | JR68a | duplolimbata1_2_4 | Taita Hills | Vuria | OP999470 |
| Sticta duplolimbata | JR69 | duplolimbata1 | Taita Hills | Vuria | OP999471 |
| Sticta duplolimbata | UK170790 | duplolimbata1 | Kilimanjaro | FOc2 | OP999483 |
| Sticta duplolimbata | UK170799d | duplolimbata1 | Kilimanjaro | FOc1 | OP999489 |
| Sticta duplolimbata | UK170845o | - | Kilimanjaro | FPD3 | - |
| Sticta duplolimbata | UK170893a | duplolimbata1 | Kilimanjaro | FPo5 | OP999511 |
| Sticta duplolimbata | UK170914b | duplolimbata2 | Kilimanjaro | FOD5 | OP999519 |
| Sticta duplolimbata | UK170929c | duplolimbata1 | Kilimanjaro | Flm6 | OP999521 |
| Sticta duplolimbata | UK170931h | duplolimbata3 | Kilimanjaro | Flm1 | OP999524 |
| Sticta duplolimbata | UK170933g | - | Kilimanjaro | Flm1 | - |
| Sticta duplolimbata | UK170936g | - | Kilimanjaro | Flm1 | - |
| Sticta duplolimbata | UK170950c | - | Kilimanjaro | Flm1 | - |
| Sticta duplolimbata | UK170952e | duplolimbata1 | Kilimanjaro | Flm1 | OP999529 |
| Sticta duplolimbata | UK171178f | duplolimbata1 | Kilimanjaro | FOD3 | OP999536 |
| Sticta duplolimbata | UK171181e | duplolimbata2 | Kilimanjaro | FOD3 | OP999538 |
| Sticta duplolimbata | UK171182b | - | Kilimanjaro | FOD3 | - |
| Sticta duplolimbata | UK171188b | - | Kilimanjaro | FOD3 | - |
| Sticta duplolimbata | UK171472d | - | Kilimanjaro | FPD1 | - |
| Sticta duplolimbata | UK171483g | - | Kilimanjaro | FOc4 | - |
| Sticta duplolimbata | UK171490f | duplolimbata1 | Kilimanjaro | FOc3 | OP999567 |
| Sticta duplolimbata | UK171493b | - | Kilimanjaro | FOc3 | - |
| Sticta duplolimbata | UK171495e | duplolimbata1 | Kilimanjaro | FOc3 | OP999569 |
| Sticta duplolimbata | UK171497e | duplolimbata1 | Kilimanjaro | FOD1 | OP999572 |
| Sticta duplolimbata | UK171497f | duplolimbata1 | Kilimanjaro | FOD1 | OP999573 |
| Sticta duplolimbata | UK171502b | duplolimbata1 | Kilimanjaro | FOD1 | OP999574 |
| Sticta duplolimbata | UK171502c | duplolimbata1 | Kilimanjaro | FOD1 | OP999575 |
| Sticta duplolimbata | UK171503c | - | Kilimanjaro | FOD1 | - |
| Sticta duplolimbata | UK171504b | duplolimbata1 | Kilimanjaro | FOD2 | OP999576 |
| Sticta duplolimbata | UK171513f | duplolimbata1 | Kilimanjaro | FOc5 | OP999582 |
| Sticta duplolimbata | UK171519g | duplolimbata1 | Kilimanjaro | FOc5 | OP999585 |
| Sticta duplolimbata | UK171521h | - | Kilimanjaro | FOc5 | - |
| Sticta duplolimbata | UK171586p | duplolimbata2 | Kilimanjaro | FPo3 | OP999605 |
| Sticta duplolimbata | UK171590q | duplolimbata2 | Kilimanjaro | FPo3 | OP999609 |
| Sticta duplolimbata | UK171596e | duplolimbata1 | Kilimanjaro | Flm2 | OP999610 |
| Sticta fuliginoides | UK170808e | fuliginoides1 | Kilimanjaro | FPD4 | OP999494 |
| Sticta fuliginoides | UK170844c | fuliginoides1 | Kilimanjaro | FPD3 | OP999499 |
| Sticta fuliginoides | UK170858e | fuliginoides1 | Kilimanjaro | FPo4 | OP999505 |
| Sticta fuliginoides | UK171438f | fuliginoides1 | Kilimanjaro | FEr2 | OP999552 |
| Sticta fuliginoides | UK171468d | fuliginoides1 | Kilimanjaro | FPD1 | OP999558 |
| Sticta fuliginoides | UK171485c | fuliginoides1 | Kilimanjaro | FOc4 | OP999564 |
| Sticta fuliginoides | UK171504c | fuliginoides1 | Kilimanjaro | FOD2 | OP999577 |
| Sticta fuliginoides | UK171577i | fuliginoides1 | Kilimanjaro | FPD2 | OP999597 |
| Sticta fuliginoides (agg.) | UK170773c | - | Kilimanjaro | FPo1 | - |
| Sticta fuliginoides (agg.) | UK170775c | - | Kilimanjaro | FPo1 | - |
| Sticta fuliginoides (agg.) | UK170826g | - | Kilimanjaro | FPD4 | - |
| Sticta fuliginoides (agg.) | UK170845m | - | Kilimanjaro | FPD3 | - |
| Sticta fuliginoides (agg.) | UK170888g | - | Kilimanjaro | FPo5 | - |
| Sticta fuliginoides (agg.) | UK170889e | - | Kilimanjaro | FPo5 | - |
| Sticta fuliginoides (agg.) | UK170897g | - | Kilimanjaro | FOD4 | - |
| Sticta fuliginoides (agg.) | UK171439h | - | Kilimanjaro | FEr2 | - |
| Sticta fuliginoides (agg.) | UK171445h | - | Kilimanjaro | FEr2 | - |
| Sticta fuliginoides (agg.) | UK171456b | - | Kilimanjaro | FEr2 | - |
| Sticta fuliginoides (agg.) | UK171458m | - | Kilimanjaro | FEr4 | - |
| Sticta fuliginoides (agg.) | UK171467j | - | Kilimanjaro | FEr4 | - |
| Sticta fuliginoides (agg.) | UK171471d | - | Kilimanjaro | FPD1 | - |
| Sticta fuliginoides (agg.) | UK171473m | - | Kilimanjaro | FPD1 | - |
| Sticta fuliginoides (agg.) | UK171497ab | - | Kilimanjaro | FOD1 | - |
| Sticta fuliginoides (agg.) | UK171505d | - | Kilimanjaro | FOD2 | - |
| Sticta fuliginoides (agg.) | UK171514d | - | Kilimanjaro | FOc5 | - |
| Sticta fuliginoides (agg.) | UK171523m | - | Kilimanjaro | FPo2 | - |
| Sticta fuliginoides (agg.) | UK171582j | - | Kilimanjaro | FPD2 | - |
| Sticta fuliginoides (agg.) | UK171584l | - | Kilimanjaro | FPD2 | - |
| Sticta fuliginoides (agg.) | UK171586m | - | Kilimanjaro | FPo3 | - |
| Sticta fuliginoides (agg.) | UK171590r | - | Kilimanjaro | FPo3 | - |
| Sticta fuliginosa | JR09A46 | fuliginosa1 | Taita Hills | Ngangao | OP999381 |
| Sticta fuliginosa | JR09D21B | fuliginosa1 | Taita Hills | Ngangao | OP999384 |
| Sticta fuliginosa | JR10076 | fuliginosa1 | Taita Hills | Vuria | OP999399 |
| Sticta fuliginosa | JR10151 | fuliginosa1 | Taita Hills | Yale | OP999405 |
| Sticta fuliginosa | JR10190B | fuliginosa1 | Taita Hills | Yale | OP999413 |
| Sticta fuliginosa | JR10245 | fuliginosa2 | Taita Hills | Mwachora | OP999425 |
| Sticta fuliginosa | JR105A | fuliginosa1 | Taita Hills | Vuria | OP999428 |
| Sticta fuliginosa | JR105B | fuliginosa1 | Taita Hills | Vuria | OP999429 |
| Sticta fuliginosa | JR10K564 | - | Mt. Kasigau | Plot W12 | - |
| Sticta fuliginosa | JR16335 | fuliginosa1 | Taita Hills | Sagalla | OP999448 |
| Sticta fuliginosa | JR16354 | fuliginosa1 | Taita Hills | Vuria | OP999450 |
| Sticta fuliginosa | JR16358 | fuliginosa1 | Taita Hills | Vuria | OP999452 |
| Sticta fuliginosa | JR16457 | - | Taita Hills | Mbololo | |
| Sticta fuliginosa | JR16463 | fuliginosa1 | Taita Hills | Mbololo | OP999468 |
| Sticta fuliginosa | JR19 | - | Taita Hills | Yale | - |
| Sticta fuliginosa | JR3 | fuliginosa1 | Taita Hills | Yale | OP999469 |
| Sticta fuliginosa | UK110504b | - | Taita Hills | Ngangao | - |
| Sticta fuliginosa | UK110512a | - | Taita Hills | Vuria | - |
| Sticta fuliginosa | UK170794g | fuliginosa1 | Kilimanjaro | FOc2 | OP999486 |
| Sticta fuliginosa | UK170796a | fuliginosa1 | Kilimanjaro | FOc2 | OP999487 |
| Sticta fuliginosa | UK170797j | fuliginosa1 | Kilimanjaro | FOc1 | OP999488 |
| Sticta fuliginosa | UK170846f | fuliginosa1 | Kilimanjaro | FPD3 | OP999502 |
| Sticta fuliginosa | UK170900h | fuliginosa1 | Kilimanjaro | FOD4 | OP999513 |
| Sticta fuliginosa | UK170906b | fuliginosa1 | Kilimanjaro | FOD4 | OP999515 |
| Sticta fuliginosa | UK170930b | fuliginosa1 | Kilimanjaro | Flm1 | OP999522 |
| Sticta fuliginosa | UK171494n | fuliginosa1 | Kilimanjaro | FOc3 | OP999568 |
| Sticta fuliginosa | UK171504m | fuliginosa1 | Kilimanjaro | FOD2 | OP999579 |
| Sticta fuliginosa | UK171594e | - | Kilimanjaro | Flm2 | - |
| Sticta marginalis | JR_58_7 | marginalis3 | Mt. Kasigau | Plot W14 | OP999379 |
| Sticta marginalis | JR090X8a | - | Taita Hills | Chawia | - |
| Sticta marginalis | JR09D12A | marginalis1 | Taita Hills | Ngangao | OP999383 |
| Sticta marginalis | JR10028B | marginalis1 | Taita Hills | Vuria | OP999391 |
| Sticta marginalis | JR10176A | marginalis1 | Taita Hills | Yale | OP999410 |
| Sticta marginalis | JR10193A | marginalis1 | Taita Hills | Yale | OP999415 |
| Sticta marginalis | JR10202B | marginalis1 | Taita Hills | Yale | OP999417 |
| Sticta marginalis | JR10K532 | - | Mt. Kasigau | Plot W9 | - |
| Sticta marginalis | JR10K552A | - | Mt. Kasigau | Plot W11 | - |
| Sticta marginalis | JR10K552B | - | Mt. Kasigau | Plot W11 | - |
| Sticta marginalis | JR10K563 | - | Mt. Kasigau | Plot W12 | - |
| Sticta marginalis | JR110030A | marginalis4 | Taita Hills | Vuria | OP999432 |
| Sticta marginalis | JR110055 | marginalis1 | Taita Hills | Vuria | OP999433 |
| Sticta marginalis | JR16248 | marginalis1 | Taita Hills | Mbololo | OP999441 |
| Sticta marginalis | JR16294a | - | Taita Hills | Sagalla | - |
| Sticta marginalis | JR16312 | marginalis1 | Taita Hills | Sagalla | OP999446 |
| Sticta marginalis | JR16322a | marginalis1 | Taita Hills | Sagalla | OP999447 |
| Sticta marginalis | JR16420 | - | Taita Hills | Mbololo | - |
| Sticta marginalis | JR16440 | - | Taita Hills | Mbololo | - |
| Sticta marginalis | UK110501a | - | Taita Hills | Ngangao | - |
| Sticta marginalis | UK110535b | - | Taita Hills | Vuria | - |
| Sticta marginalis | UK170800m | marginalis2 | Kilimanjaro | FOc1 | OP999490 |
| Sticta marginalis | UK170927a | - | Kilimanjaro | Flm6 | - |
| Sticta marginalis | UK170932b | marginalis1 | Kilimanjaro | Flm1 | OP999525 |
| Sticta marginalis | UK170933h | marginalis1 | Kilimanjaro | Flm1 | OP999526 |
| Sticta marginalis | UK170934b | - | Kilimanjaro | Flm1 | - |
| Sticta marginalis | UK170937j | - | Kilimanjaro | Flm1 | - |
| Sticta marginalis | UK170942c | marginalis1 | Kilimanjaro | Flm1 | OP999527 |
| Sticta marginalis | UK170943d | - | Kilimanjaro | Flm1 | - |
| Sticta marginalis | UK170950d | - | Kilimanjaro | Flm1 | - |
| Sticta marginalis | UK171183a | marginalis1 | Kilimanjaro | FOD3 | OP999540 |
| Sticta marginalis | UK171183b | - | Kilimanjaro | FOD3 | - |
| Sticta marginalis | UK171186a | - | Kilimanjaro | FOD3 | - |
| Sticta marginalis | UK171504k | marginalis1 | Kilimanjaro | FOD2 | OP999578 |
| Sticta marginalis | UK171596k | marginalis1 | Kilimanjaro | Flm2 | OP999611 |
| Sticta munda | UK170888c | - | Kilimanjaro | FPo5 | - |
| Sticta munda | UK171497d | munda3 | Kilimanjaro | FOD1 | OP999571 |
| Sticta munda | UK171508j | munda1 | Kilimanjaro | FOD2 | OP999580 |
| Sticta munda | UK171510k | munda1 | Kilimanjaro | FOD2 | OP999581 |
| Sticta munda | UK171582i | munda2 | Kilimanjaro | FPD2 | OP999598 |
| Sticta munda | UK171584u | munda1 | Kilimanjaro | FPD2 | OP999600 |
| Sticta munda | UK171586k | munda1 | Kilimanjaro | FPo3 | OP999604 |
| Sticta munda | UK171586l | - | Kilimanjaro | FPo3 | - |
| Sticta sublimbata | JR09A60 | sublimbata1 | Taita Hills | Ngangao | OP999382 |
| Sticta sublimbata | JR09D6A | sublimbata1 | Taita Hills | Ngangao | OP999385 |
| Sticta sublimbata | JR09W12a | sublimbata8 | Mt. Kasigau | - | OP999387 |
| Sticta sublimbata | JR09Y12a | sublimbata6 | Taita Hills | Mbololo | OP999388 |
| Sticta sublimbata | JR09Y15b | sublimbata6 | Taita Hills | Mbololo | OP999389 |
| Sticta sublimbata | JR09Z2 | sublimbata1 | Taita Hills | Ngangao | OP999390 |
| Sticta sublimbata | JR10032A | sublimbata2 | Taita Hills | Shomoto Hill | OP999392 |
| Sticta sublimbata | JR10039B | sublimbata2 | Taita Hills | Shomoto Hill | OP999393 |
| Sticta sublimbata | JR10066A | sublimbata2 | Taita Hills | Shomoto Hill | OP999397 |
| Sticta sublimbata | JR10066B | sublimbata2 | Taita Hills | Shomoto Hill | OP999398 |
| Sticta sublimbata | JR10191 | sublimbata1 | Taita Hills | Yale | OP999414 |
| Sticta sublimbata | JR10208A | sublimbata5 | Taita Hills | Yale | OP999418 |
| Sticta sublimbata | JR10214B | sublimbata1 | Taita Hills | Yale | OP999420 |
| Sticta sublimbata | JR10220A | sublimbata2 | Taita Hills | Mwachora | OP999421 |
| Sticta sublimbata | JR10228D | sublimbata1 | Taita Hills | Mwachora | OP999422 |
| Sticta sublimbata | JR10230A | sublimbata1 | Taita Hills | Mwachora | OP999423 |
| Sticta sublimbata | JR10241 | sublimbata1 | Taita Hills | Mwachora | OP999424 |
| Sticta sublimbata | JR10282B | sublimbata1 | Taita Hills | Macha | OP999427 |
| Sticta sublimbata | JR10K494 | - | Mt. Kasigau | Plot W5 | - |
| Sticta sublimbata | JR10K495 | - | Mt. Kasigau | Plot W5 | - |
| Sticta sublimbata | JR16275 | sublimbata9 | Taita Hills | Sagalla | OP999442 |
| Sticta sublimbata | JR16355 | sublimbata2 | Taita Hills | Vuria | OP999451 |
| Sticta sublimbata | JR16367a | sublimbata2 | Taita Hills | Vuria | OP999454 |
| Sticta sublimbata | JR16367b | sublimbata2 | Taita Hills | Vuria | OP999455 |
| Sticta sublimbata | JR16373 | sublimbata4 | Taita Hills | Vuria | OP999458 |
| Sticta sublimbata | JR16374a | sublimbata1 | Taita Hills | Vuria | OP999459 |
| Sticta sublimbata | JR16374c | sublimbata2 | Taita Hills | Vuria | OP999461 |
| Sticta sublimbata | JR16417 | sublimbata3 | Taita Hills | Mbololo | OP999463 |
| Sticta sublimbata | JR16419 | sublimbata7 | Taita Hills | Mbololo | OP999464 |
| Sticta sublimbata | JR16443 | sublimbata8 | Taita Hills | Mbololo | OP999466 |
| Sticta sublimbata | UK110502 | - | Taita Hills | Ngangao | - |
| Sticta sublimbata | UK110510a | - | Taita Hills | Vuria | - |
| Sticta sublimbata | UK110520a | - | Taita Hills | Vuria | - |
| Sticta sublimbata | UK110539d | - | Taita Hills | Vuria | - |
| Sticta sublimbata | UK110547a | sublimbata1 | Taita Hills | Fururu | OP999473 |
| Sticta sublimbata | UK110547b | sublimbata1 | Taita Hills | Fururu | OP999474 |
| Sticta sublimbata | UK110554a | - | Taita Hills | Chawia | - |
| Sticta sublimbata | UK110556a | - | Taita Hills | Chawia | - |
| Sticta sublimbata | UK160482e | - | Kilimanjaro | Home1 | - |
| Sticta sublimbata | UK160552m | - | Kilimanjaro | Home3 | - |
| Sticta sublimbata | UK160557f | - | Kilimanjaro | Home3 | - |
| Sticta sublimbata | UK160559b | - | Kilimanjaro | Home3 | - |
| Sticta tomentosa | JR10086 | tomentosa1 | Taita Hills | Vuria | OP999400 |
| Sticta tomentosa | JR10095A | tomentosa1 | Taita Hills | Vuria | OP999401 |
| Sticta tomentosa | JR10260 | tomentosa1 | Taita Hills | Mwachora | OP999426 |
| Sticta tomentosa | JR16352 | tomentosa1 | Taita Hills | Vuria | OP999449 |
| Sticta tomentosa | JR16367c | tomentosa1 | Taita Hills | Vuria | OP999456 |
| Sticta tomentosa | JR16368 | tomentosa1 | Taita Hills | Vuria | OP999457 |
| Sticta tomentosa | UK110523b | tomentosa1 | Taita Hills | Vuria | OP999472 |
| Sticta tomentosa | UK170912c | tomentosa1 | Kilimanjaro | FOD5 | OP999517 |
| Sticta tomentosa | UK170934c | - | Kilimanjaro | Flm1 | - |
| Sticta tomentosa | UK170936f | - | Kilimanjaro | Flm1 | - |
| Sticta tomentosa | UK170943c | - | Kilimanjaro | Flm1 | - |
| Sticta tomentosa | UK170950b | - | Kilimanjaro | Flm1 | - |
| Sticta tomentosa | UK171473j | - | Kilimanjaro | FPD1 | - |
| Sticta umbilicariiformis | JR16098 | umbilicariiformis1 | Mt. Elgon | Erica zone | OP999434 |
| Sticta umbilicariiformis | JR16102 | umbilicariiformis3 | Mt. Elgon | Erica zone | OP999435 |
| Sticta umbilicariiformis | JR16108 | umbilicariiformis1 | Mt. Elgon | Erica zone | OP999438 |
| Sticta umbilicariiformis | UK170774k | umbilicariiformis3 | Kilimanjaro | FPo1 | OP999478 |
| Sticta umbilicariiformis | UK170775a | umbilicariiformis3 | Kilimanjaro | FPo1 | OP999479 |
| Sticta umbilicariiformis | UK170794c | - | Kilimanjaro | FOc2 | - |
| Sticta umbilicariiformis | UK170806m | umbilicariiformis3 | Kilimanjaro | FPD4 | OP999492 |
| Sticta umbilicariiformis | UK170808c | umbilicariiformis3 | Kilimanjaro | FPD4 | OP999493 |
| Sticta umbilicariiformis | UK170821d | - | Kilimanjaro | FPD4 | - |
| Sticta umbilicariiformis | UK170821h | umbilicariiformis1 | Kilimanjaro | FPD4 | OP999495 |
| Sticta umbilicariiformis | UK170826f | - | Kilimanjaro | FPD4 | - |
| Sticta umbilicariiformis | UK170842d | umbilicariiformis3 | Kilimanjaro | FPD3 | OP999497 |
| Sticta umbilicariiformis | UK170846d | umbilicariiformis3 | Kilimanjaro | FPD3 | OP999500 |
| Sticta umbilicariiformis | UK170858d | umbilicariiformis3 | Kilimanjaro | FPo4 | OP999504 |
| Sticta umbilicariiformis | UK171102b | umbilicariiformis1 | Kilimanjaro | FED5 | OP999531 |
| Sticta umbilicariiformis | UK171106b | umbilicariiformis2 | Kilimanjaro | FED5 | OP999532 |
| Sticta umbilicariiformis | UK171119b | umbilicariiformis5 | Kilimanjaro | FED5 | OP999533 |
| Sticta umbilicariiformis | UK171131a | umbilicariiformis5 | Kilimanjaro | FED4 | OP999534 |
| Sticta umbilicariiformis | UK171131b | umbilicariiformis1 | Kilimanjaro | FED4 | OP999535 |
| Sticta umbilicariiformis | UK171320e | umbilicariiformis4 | Kilimanjaro | FED1 | OP999541 |
| Sticta umbilicariiformis | UK171320f | umbilicariiformis4 | Kilimanjaro | FED1 | OP999542 |
| Sticta umbilicariiformis | UK171320g | umbilicariiformis4 | Kilimanjaro | FED1 | OP999543 |
| Sticta umbilicariiformis | UK171337 | umbilicariiformis1 | Kilimanjaro | FED1 | OP999544 |
| Sticta umbilicariiformis | UK171340s | - | Kilimanjaro | FED1 | - |
| Sticta umbilicariiformis | UK171404e | umbilicariiformis1 | Kilimanjaro | FEr4 | OP999546 |
| Sticta umbilicariiformis | UK171405f | umbilicariiformis1 | Kilimanjaro | FEr4 | OP999547 |
| Sticta umbilicariiformis | UK171411c | umbilicariiformis4 | Kilimanjaro | FEr4 | OP999550 |
| Sticta umbilicariiformis | UK171429 | umbilicariiformis4 | Kilimanjaro | FEr3 | OP999551 |
| Sticta umbilicariiformis | UK171433a | - | Kilimanjaro | FEr3 | - |
| Sticta umbilicariiformis | UK171435f | - | Kilimanjaro | FEr3 | - |
| Sticta umbilicariiformis | UK171438h | umbilicariiformis4 | Kilimanjaro | FEr2 | OP999553 |
| Sticta umbilicariiformis | UK171439u | umbilicariiformis1 | Kilimanjaro | FEr2 | OP999555 |
| Sticta umbilicariiformis | UK171442b | - | Kilimanjaro | FEr2 | - |
| Sticta umbilicariiformis | UK171449i | umbilicariiformis4 | Kilimanjaro | FEr2 | OP999556 |
| Sticta umbilicariiformis | UK171455h | umbilicariiformis1 | Kilimanjaro | FEr2 | OP999557 |
| Sticta umbilicariiformis | UK171458h | - | Kilimanjaro | FEr4 | - |
| Sticta umbilicariiformis | UK171467k | - | Kilimanjaro | FEr4 | - |
| Sticta umbilicariiformis | UK171468e | umbilicariiformis3 | Kilimanjaro | FPD1 | OP999559 |
| Sticta umbilicariiformis | UK171468f | umbilicariiformis3 | Kilimanjaro | FPD1 | OP999560 |
| Sticta umbilicariiformis | UK171494k | - | Kilimanjaro | FOc3 | - |
| Sticta umbilicariiformis | UK171514c | umbilicariiformis3 | Kilimanjaro | FOc5 | OP999583 |
| Sticta umbilicariiformis | UK171521f | umbilicariiformis3 | Kilimanjaro | FOc5 | OP999588 |
| Sticta umbilicariiformis | UK171524b | umbilicariiformis3 | Kilimanjaro | FPo2 | OP999589 |
| Sticta umbilicariiformis | UK171563a | umbilicariiformis2 | Kilimanjaro | FED2 | OP999593 |
| Sticta umbilicariiformis | UK171569d | umbilicariiformis4 | Kilimanjaro | FED2 | OP999594 |
| Sticta umbilicariiformis | UK171577h | umbilicariiformis1 | Kilimanjaro | FPD2 | OP999596 |
| Sticta umbilicariiformis | UK171578t | - | Kilimanjaro | FPD2 | - |
| Sticta umbilicariiformis | UK171584x | umbilicariiformis2 | Kilimanjaro | FPD2 | OP999602 |
| Sticta umbilicariiformis | UK171584y | umbilicariiformis1 | Kilimanjaro | FPD2 | OP999603 |
| Sticta umbilicariiformis | UK171590j | umbilicariiformis3 | Kilimanjaro | FPo3 | OP999608 |
| Sticta sp. A (fuliginoides agg.) | UK170858f | A1 | Kilimanjaro | FPo4 | OP999506 |
| Sticta sp. A (fuliginoides agg.) | UK170884b | A1 | Kilimanjaro | FPo5 | OP999508 |
| Sticta sp. A (fuliginoides agg.) | UK171519h | A1 | Kilimanjaro | FOc5 | OP999586 |
| Sticta sp. A (fuliginoides agg.) | UK171521g | - | Kilimanjaro | FOc5 | - |
| Sticta sp. B (ciliata agg.) | JR09D8A | B1 | Taita Hills | Ngangao | OP999386 |
| Sticta sp. B (ciliata agg.) | JR10060C | B2 | Taita Hills | Shomoto Hill | OP999396 |
| Sticta sp. B (ciliata agg.) | UK170792q | B1 | Kilimanjaro | FOc2 | OP999485 |
| Sticta sp. B (ciliata agg.) | UK170901b | B1 | Kilimanjaro | FOD4 | OP999514 |
| Sticta sp. C (weigelii agg.) | JR090X3 | C1 | Taita Hills | Chawia | OP999380 |
| Sticta sp. C (weigelii agg.) | JR10180B | C1 | Taita Hills | Yale | OP999411 |
| Sticta sp. C (weigelii agg.) | JR10180C | - | Taita Hills | Yale | - |
| Sticta sp. C (weigelii agg.) | JR10212B | C1 | Taita Hills | Yale | OP999419 |
| Sticta sp. C (weigelii agg.) | JR14634/082 | - | Taita Hills | Maktau Hill | - |
| Sticta sp. C (weigelii agg.) | JR16201a | C2 | Taita Hills | Fururu | OP999439 |
| Sticta sp. C (weigelii agg.) | JR16201b | C2 | Taita Hills | Fururu | OP999440 |
| Sticta sp. C (weigelii agg.) | JR16295 | C1 | Taita Hills | Sagalla | OP999444 |
| Sticta sp. C (weigelii agg.) | JR16310 | C1 | Taita Hills | Sagalla | OP999445 |
| Sticta sp. C (weigelii agg.) | JR16439 | C1 | Taita Hills | Mbololo | OP999465 |
| Sticta sp. C (weigelii agg.) | JR16462 | C1 | Taita Hills | Mbololo | OP999467 |
| Sticta sp. C (weigelii agg.) | UK110555e | - | Taita Hills | Chawia | - |
| Sticta sp. D | UK171519k | D1 | Kilimanjaro | FOc5 | OP999587 |
| Sticta sp. D | UK171586q | D1 | Kilimanjaro | FPo3 | OP999606 |
| Sticta sp. 2 | JR09Y12b | - | Taita Hills | Mbololo | - |
| Sticta sp. 2 | JR09Y13 | - | Taita Hills | Mbololo | - |
| Sticta sp. 2 | JR10K302 | sp. 2 | Mt. Kasigau | Plot E15 | OP999430 |
| Sticta sp. 2 | JR10K303 | sp. 2 | Mt. Kasigau | Plot E15 | OP999431 |
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Merckx, V.S.F.T.; Hendriks, K.P.; Beentjes, K.K.; Mennes, C.B.; Becking, L.E.; Peijnenburg, K.T.C.A.; Afendy, A.; Arumugam, N.; de Boer, H.; Biun, A.; et al. Evolution of endemism on a young tropical mountain. Nature 2015, 524, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Hemp, C.; Kehl, S.; Schultz, O.; Wägele, J.W.; Hemp, A. Climatic fluctuations and orogenesis as motors for speciation in East Africa: Case study on Parepistaurus Karsch, 1896 (Orthoptera). Syst. Entomol. 2015, 40, 17–34. [Google Scholar] [CrossRef]
- Schwery, O.; Onstein, R.E.; Bouchenak-Khelladi, Y.; Xing, Y.; Carter, R.J.; Linder, H.P. As old as the mountains: The radiations of the Ericaceae. New Phytol. 2015, 207, 355–367. [Google Scholar] [CrossRef]
- Griffiths, C.J. The ecological evolution of East Africa. In Biogeography and Ecology of the Rainforests of Eastern Africa; Lovett, J.C., Wasser, S.K., Eds.; Cambridge University Press: Cambridge, UK, 1993; pp. 9–21. ISBN 0-521-43083-6. [Google Scholar]
- Wasser, S.K.; Lovett, J.C. (Eds.) Introduction to the biography and ecology of the rain forests of eastern Africa. In Biogeography and Ecology of the Rainforests of Eastern Africa; Cambridge University Press: Cambridge, UK, 1993; pp. 3–8. ISBN 0-521-43083-6. [Google Scholar]
- Lovett, J.C. Eastern Arc moist forest flora. In Biogeography and Ecology of the Rainforests of Eastern Africa; Lovett, J.C., Wasser, S.K., Eds.; Cambridge University Press: Cambridge, UK, 1993; pp. 33–55. ISBN 0-521-43083-6. [Google Scholar]
- Hemp, A.; Hemp, C. Broken bridges: The isolation of Kilimanjaro’s ecosystem. Glob. Chang. Biol. 2018, 24, 3499–3507. [Google Scholar] [CrossRef]
- Hemp, C.; Grzywacz, B.; Warchałowska-Śliwa, E.; Hemp, A. Topography and climatic fluctuations boosting speciation: Biogeography and a molecular phylogeny of the East African genera Afroanthracites Hemp & Ingrisch and Afroagraecia Ingrisch & Hemp (Orthoptera, Tettigoniidae, Conocephalinae, Agraeciini). Org. Divers. Evol. 2016, 16, 211–223. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Robles, G.P.; Hoffmann, M.; Pilgrim, J.; Brooks, T.; Mittermeier, C.G.; Lamoreux, J.; Da Fonseca, G.A.B. Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Ecoregions; CEMEX: Mexico City, Mexico, 2004; ISBN 968-6397-77-9. [Google Scholar]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global Biodiversity Conservation: The Critical Role of Hotspots. In Biodiversity Hotspots; Zachos, F.E., Habel, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–22. [Google Scholar] [CrossRef]
- Swinscow, T.D.V.; Krog, H. Macrolichens of East Africa; Natural History Museum Publications: London, UK, 1988; ISBN 0565010395. [Google Scholar]
- Kaasalainen, U.; Tuovinen, V.; Kirika, P.; Mollel, N.; Hemp, A.; Rikkinen, J. Diversity of Leptogium (Collemataceae, Ascomycota) in East African Montane Ecosystems. Microorganisms 2021, 9, 314. [Google Scholar] [CrossRef]
- Kirika, P.M.; Divakar, P.K.; Buaruang, K.; Leavitt, S.D.; Crespo, A.; Gatheri, G.W.; Mugambi, G.; Benatti, M.N.; Lumbsch, H.T. Molecular phylogenetic studies unmask overlooked diversity in the tropical lichenized fungal genus Bulbothrix s.l. (Parmeliaceae, Ascomycota). Bot. J. Linn. Soc. 2017, 184, 387–399. [Google Scholar] [CrossRef]
- Kirika, P.M.; Divakar, P.K.; Crespo, A.; Leavitt, S.D.; Mugambi, G.; Gatheri, G.W.; Lumbsch, H.T. Polyphyly of the genus Canoparmelia—Uncovering incongruences between phenotype-based classification and molecular phylogeny within lichenized Ascomycota (Parmeliaceae). Phytotaxa 2016, 289, 36–48. [Google Scholar] [CrossRef]
- Kirika, P.M.; Divakar, P.K.; Crespo, A.; Lumbsch, H.T. Molecular and phenotypical studies on species diversity of Hypotrachyna (Parmeliaceae, Ascomycota) in Kenya, East Africa. Bryologist 2019, 122, 140–150. [Google Scholar] [CrossRef]
- Kaasalainen, U.; Biermann, L.; Mollel, N.P.; Schmidt, A.R.; Hemp, A. Peltigera (Lecanoromycetes) on Mt Kilimanjaro, East Africa. Lichenologist 2022, 54, 231–243. [Google Scholar] [CrossRef]
- Suija, A.; Kaasalainen, U.; Kirika, P.M.; Rikkinen, J. Taitaia, a novel lichenicolous fungus in tropical montane forests in Kenya (East Africa). Lichenologist 2018, 50, 173–184. [Google Scholar] [CrossRef]
- Moncada, B.; Lücking, R.; Suárez, A. Molecular phylogeny of the genus Sticta (lichenized Ascomycota: Lobariaceae) in Colombia. Fungal Divers. 2014, 64, 205–231. [Google Scholar] [CrossRef]
- Magain, N.; Sérusiaux, E. Dismantling the treasured flagship lichen Sticta fuliginosa (Peltigerales) into four species in Western Europe. Mycol. Prog. 2015, 14, 125. [Google Scholar] [CrossRef]
- Moncada, B.; Mercado-Díaz, J.A.; Smith, C.W.; Bungartz, F.; Sérusiaux, E.; Lumbsch, H.T.; Lücking, R. Two new common, previously unrecognized species in the Sticta weigelii morphodeme (Ascomycota: Peltigeraceae). Willdenowia 2021, 51, 35–45. [Google Scholar] [CrossRef]
- Ossowska, E.A.; Moncada, B.; Kukwa, M.; Flakus, A.; Rodriguez-Flakus, P.; Olszewska, S.; Lücking, R. New species of Sticta (lichenised Ascomycota, lobarioid Peltigeraceae) from Bolivia suggest a high level of endemism in the Central Andes. Mycokeys 2022, 92, 131–160. [Google Scholar] [CrossRef]
- Mercado-Díaz, J.A.; Lücking, R.; Moncada, B.; Widhelm, T.J.; Lumbsch, H.T. Elucidating species richness in lichen fungi: The genus Sticta (Ascomycota: Peltigeraceae) in Puerto Rico. Taxon 2020, 69, 851–891. [Google Scholar] [CrossRef]
- Widhelm, T.J.; Bertoletti, F.R.; Asztalos, M.J.; Mercado-Díaz, J.A.; Huang, J.-P.; Moncada, B.; Lücking, R.; Magain, N.; Sérusiaux, E.; Goffinet, B.; et al. Oligocene origin and drivers of diversification in the genus Sticta (Lobariaceae, Ascomycota). Mol. Phylogenetics Evol. 2018, 126, 58–73. [Google Scholar] [CrossRef]
- Simon, A.; Goffinet, B.; Magain, N.; Sérusiaux, E. High diversity, high insular endemism and recent origin in the lichen genus Sticta (lichenized Ascomycota, Peltigerales) in Madagascar and the Mascarenes. Mol. Phylogenetics Evol. 2018, 122, 15–28. [Google Scholar] [CrossRef]
- Kirika, P.; Mugambi, G.; Lücking, R.; Lumbsch, H.T. New Records of Lichen-Forming Fungi from Kenya. J. East Afr. Nat. Hist. 2012, 101, 73–98. [Google Scholar] [CrossRef]
- Frisch, A.; Hertel, H. Flora of macrolichens in the alpine and subalpine zones of Mount Kenya (Kenya). Sauteria 1998, 9, 363–370. [Google Scholar]
- Alstrup, V.; Christensen, S.N. New records of lichens with cyanobacteria from Tanzania and Kenya. Cryptogam. Mycol. 2006, 27, 57–68. [Google Scholar]
- Kirika, P.M.; Ndiritu, G.G.; Mugambi, G.K.; Newton, L.E.; Lumbsch, H.T. Diversity and Altitudinal Distribution of Understorey Corticolous lichens in a tropical montane forest in Kenya (East Africa). Crypt. Biodivers. Assess. 2018, 2018, 47–70. [Google Scholar] [CrossRef]
- Krog, H. Corticolous macrolichens of low montane rainforests and moist woodlands of eastern Tanzania. Sommerfeltia 2000, 28, 1. [Google Scholar] [CrossRef]
- Kaasalainen, U.; Tuovinen, V.; Mwachala, G.; Pellikka, P.; Rikkinen, J. Complex Interaction Networks Among Cyanolichens of a Tropical Biodiversity Hotspot. Front. Microbiol. 2021, 12, 672333. [Google Scholar] [CrossRef]
- Galloway, D.J.; Thomas, M.A. Sticta. In Lichen Flora of the Greater Sonoran Desert Region; Nash, T.H., III, Ryan, B.D., Diederich, P., Gries, C., Bungartz, F., Eds.; Lichens Unlimited, Arizona State University: Tempe, AZ, USA, 2004; Volume 2, pp. 513–524. ISBN 0971675910. [Google Scholar]
- Galloway, D.J. Studies on lichen genus Sticta (Schreber) Ach.: III*. Notes on species described by Bory de St-Vincent, William Hooker, and Delise, between 1804 and 1825. Nova Hedwig. 1995, 61, 147–188. [Google Scholar]
- Rutten, G.; Ensslin, A.; Hemp, A.; Fischer, M. Vertical and Horizontal Vegetation Structure across Natural and Modified Habitat Types at Mount Kilimanjaro. PLoS ONE 2015, 10, e0138822. [Google Scholar] [CrossRef]
- Bytebier, B. Taita Hills Biodiversity Project Report; National Museums of Kenya: Nairobi, Kenya, 2001. [Google Scholar]
- Pellikka, P.K.E.; Lötjönen, M.; Siljander, M.; Lens, L. Airborne remote sensing of spatiotemporal change (1955–2004) in indigenous and exotic forest cover in the Taita Hills, Kenya. Int. J. Appl. Earth Obs. Geoinf. 2009, 11, 221–232. [Google Scholar] [CrossRef]
- Beentje, H.J. An ecological and floristical study of the forests of the Taita Hills, Kenya. Utafiti 1988, 1, 23–66. [Google Scholar]
- Aerts, R.; Matthijsen, E.; Lehouck, V.; Beentje, H.; Bytebier, B.; Matthysen, E.; Gulinck, H.; Lens, L.; Muys, B. Woody plant communities of isolated Afromontane cloud forests in Taita Hills, Kenya. Plant Ecol. 2011, 212, 639–649. [Google Scholar] [CrossRef]
- Thijs, K.W.; Roelen, I.; Musila, W.M. Field Guide to the Woody Plants of Taita Hills, Kenya. J. East Afr. Nat. Hist. 2014, 102, 1–272. [Google Scholar] [CrossRef]
- Medley, K.E.; Maingi, J.K. Biogeographic Patterns of Forest Diversity at Mount Kasigau, Kenya. J. East Afr. Nat. Hist. 2015, 103, 1–24. [Google Scholar] [CrossRef]
- Henkin, M.A.; Medley, K.E.; Maingi, J.K. Biophysical analysis of afromontane forest community types at Mount Kasigau, Kenya. Afr. J. Ecol. 2015, 53, 454–464. [Google Scholar] [CrossRef]
- Enroth, J.; Nyqvist, P.; Malombe, I.; Pellikka, P.; Rikkinen, J. Additions to the moss flora of the Taita Hills and Mount Kasigau, Kenya. Pol. Bot. J. 2013, 58, 495–510. [Google Scholar] [CrossRef]
- Moncada, B.; Lücking, R. Ten new species of Sticta and counting: Colombia as a hot spot for unrecognized diversification in a conspicuous macrolichen genus. Phytotaxa 2012, 74, 1–29. [Google Scholar] [CrossRef]
- Fedrowitz, K.; Kaasalainen, U.; Rikkinen, J. Genotype variability of Nostoc symbionts associated with three epiphytic Nephroma species in a boreal forest landscape. Bryologist 2011, 114, 220–230. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PRC Protcols: A guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: London, UK, 1990; pp. 315–322. ISBN 0-12-372181-4. [Google Scholar]
- CodonCode Corporation. CodonCode Aligner v8; CodonCode Corporation: Centerville, UT, USA, 2018. [Google Scholar]
- National Center for Biotechnology Information. NCBI GenBank. Available online: https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 9 October 2022).
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Müller, K.; Müller, J.; Quandt, D. PhyDE. Available online: http://www.phyde.de/ (accessed on 23 November 2022).
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F.; Nielsen, R.; Bollback, J.P. Bayesian Inference of Phylogeny and Its Impact on Evolutionary Biology. Science 2001, 294, 2310–2314. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Larget, B.; Miller, R.E.; Ronquist, F. Potential Applications and Pitfalls of Bayesian Inference of Phylogeny. Syst. Biol. 2002, 51, 673–688. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Ossowska, E.A. First records of Sticta weigelii s.str. from Bolivia confirmed by molecular data. Folia Cryptogam. Est. 2021, 58, 65–72. [Google Scholar] [CrossRef]
- Galloway, D.J. Studies on the lichen genus Sticta (Schreber) Ach.: V*. Australian species. Bryophyt. Divers. Evol. 1998, 15, 117–160. [Google Scholar] [CrossRef]
- Moncada, B.; Smith, C.W.; Lücking, R. A taxonomic reassessment of the genus Sticta (lichenized Ascomycota: Peltigeraceae) in the Hawaiian archipelago. Lichenologist 2021, 53, 117–133. [Google Scholar] [CrossRef]
- Nylander, W. Lichenes in Regionibus Exoticis Quibusnam Vigentes. Ann. Sci. Nat. Bot. 1859, 11, 205–264. [Google Scholar]
- Lindgren, H.; Moncada, B.; Lücking, R.; Magain, N.; Simon, A.; Goffinet, B.; Sérusiaux, E.; Nelsen, M.P.; Mercado-Díaz, J.A.; Widhelm, T.J.; et al. Cophylogenetic patterns in algal symbionts correlate with repeated symbiont switches during diversification and geographic expansion of lichen-forming fungi in the genus Sticta (Ascomycota, Peltigeraceae). Mol. Phylogenetics Evol. 2020, 150, 106860. [Google Scholar] [CrossRef]
- Magain, N.; Goffinet, B.; Sérusiaux, E. Further photomorphs in the lichen family Lobariaceae from Reunion (Mascarene archipelago) with notes on the phylogeny of Dendriscocaulon cyanomorphs. Bryol. 2012, 115, 243–254. [Google Scholar] [CrossRef]
- Hue, A.M. Lichens récoltés á Java en 1894-1895 par M. Jean Massart. I. Ann. Du Jard. Bot. De Buitenzorg 1901, 11, 171–194. [Google Scholar]
- Nylander, W. Synopsis Methodica Lichenum; Martinet: Paris, France, 1858. [Google Scholar]
- Galloway, D.J. Studies on the lichen genus Sticta (Schreber) Ach.: II. Typification of taxa from Swartz’s Prodromus of 1788. Bull. Nat. Hist. Mus. Lond. 1994, 24, 35–48. [Google Scholar]
- Flotow, J.V. Lichenen des Reisevereins und des Hochstetterschen Herbarii. Linnaea: Ein Journal für die Botanik in ihrem ganzen Umfange 1843, 17, 15–30. [Google Scholar]
- Moncada, B.; Luecking, R.K.; Lumbsch, H.T. Rewriting the evolutionary history of the lichen genus Sticta (Ascomycota: Peltigeraceae subfam. Lobarioideae) in the Hawaiian islands. Plant Fungal Syst. 2020, 65, 95–119. [Google Scholar] [CrossRef]
- Galloway, D.J. Notes on the holotype of Sticta damaecornis β weigelii Ach. (=Sticta weigelii). Lichenologist 2006, 38, 89–92. [Google Scholar] [CrossRef]
- Aptroot, A.; Stech, M. An updated checklist of the lichens of St. Eustatius, Netherlands Antilles. Mycokeys 2018, 33, 69–84. [Google Scholar] [CrossRef]
- Delise, D.F. Histoire des Lichens, Genre Sticta; Harvard University: Cambridge, MA, USA, 1825; Volume 1—2, 167. [Google Scholar]
- Benítez, Á.; Prieto, M.; Gonzáles, Y.; Aragón, G. Effects of tropical montane forest disturbance on epiphytic macrolichens. Sci. Total Env. 2012, 441, 169–175. [Google Scholar] [CrossRef]
- Tripp, E.A.; Lendemer, J.C.; McCain, C.M. Habitat quality and disturbance drive lichen species richness in a temperate biodiversity hotspot. Oecologia 2019, 190, 445–457. [Google Scholar] [CrossRef]
- Rikkinen, J. Cyanolichens. Biodivers. Conserv. 2015, 24, 973–993. [Google Scholar] [CrossRef]






| Species | Distribution | Ecology | Comments | References |
|---|---|---|---|---|
| Sticta afromontana sp. nov. | K*, T* | Common especially in middle and upper montane forests. | ||
| Sticta ambavillaria | - | Not found in this study. | Previous reports may represent S. munda. | [12,27,28,29,30] |
| Sticta andina | K* | Middle montane forest (single observation). | ||
| Sticta aspratilis sp. nov. | K*, T* | Lower montane to subalpine zones. | ||
| Sticta cellulosa sp. nov. | T* | Rare in upper montane and subalpine zones. | ||
| Sticta ciliata | K*, T* | Rare in lower montane forest zone. | Previous report from Rwanda [20] represents Sticta sp. B. | |
| Sticta cyanocaperata sp. nov. | T* | Uncommon in middle montane forest. | ||
| Sticta cyphellulata | - | Not found in this study. | Previous reports may represent S. duplolimbata and/or S. marginalis. | [12,28,30] |
| Sticta dichotoma | T | Not found in this study. | [12,28,30] | |
| Sticta duplolimbata | K*, T*, R | Common in lower and middle montane forests. | [20] | |
| Sticta fuliginoides | T* | Common especially in upper montane forests. | ||
| Sticta fuliginosa | K, T, R | Common in lower and middle montane forests. | Some previous reports may represent other species of S. fuliginosa morphodeme. | [12,20,28,29,30,31] |
| Sticta kunthii | - | Not found in this study. | Previous report may represent S. umbilicariiformis and/or S. aspratilis. | [26] |
| Sticta limbata | - | Not found in this study. | Previous reports represent S. umbilicariiformis [20]. | [12,27] |
| Sticta marginalis | K*, T*, U* | Common in lower and middle montane forests. | Presence in Uganda based on previous reports of S. orbicularis [12,28,30]. | |
| Sticta munda sp. nov. | T* | Rare in middle and upper montane forests. | ||
| Sticta orbicularis | - | Not found in this study. | Previous reports may represent S. marginalis. | [12,28,30] |
| Sticta papyracea/variabilis | - | Not found in this study. | Sticta papyracea and S. variabilis are synonymous [33]. The previous reports may represent Sticta sp. 2. | [12,30] |
| Sticta sublimbata | E, K, T, U | Common in low elevation and lower montane habitats. | [12,30,31] | |
| Sticta tomentosa | K, T, U | Lower and middle montane forests. | [12,28,30,31] | |
| Sticta umbilicariiformis | E, K, T, R, U | Abundant in the upper montane and subalpine zones. | Presence in Uganda based on previous reports of S. limbata [12]. | [20] |
| Sticta weigelii | - | Not found in this study. | Previous reports may represent Sticta sp. C. | [12,27,28,29,30,31] |
| Sticta xanthotropa | - | Not found in this study. | Previously reported as S. weigelii var. xanthotropa. The reports may represent other species with marginal flattened isidia. | [12] |
| Sticta sp. A | T* | Rare in middle and upper montane forests. | Sticta fuliginoides agg. | |
| Sticta sp. B | K*, T*, R | Rare in lower and middle montane forests. | Sticta ciliata agg. Previously reported from Rwanda as S. ciliata [20]. | |
| Sticta sp. C | K* | Low elevation and lower montane habitats. | Sticta weigelii agg. May include two distinct taxa. | |
| Sticta sp. D | T* | Rare in middle and upper montane forests. | ||
| Sticta sp. 2 | K*, T | Rare in lower montane forests. | Presence in Tanzania based on previous reports of S. papyracea [12,30]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaasalainen, U.; Kirika, P.M.; Mollel, N.P.; Hemp, A.; Rikkinen, J. The Lichen Genus Sticta (Lobariaceae, Peltigerales) in East African Montane Ecosystems. J. Fungi 2023, 9, 246. https://doi.org/10.3390/jof9020246
Kaasalainen U, Kirika PM, Mollel NP, Hemp A, Rikkinen J. The Lichen Genus Sticta (Lobariaceae, Peltigerales) in East African Montane Ecosystems. Journal of Fungi. 2023; 9(2):246. https://doi.org/10.3390/jof9020246
Chicago/Turabian StyleKaasalainen, Ulla, Paul M. Kirika, Neduvoto P. Mollel, Andreas Hemp, and Jouko Rikkinen. 2023. "The Lichen Genus Sticta (Lobariaceae, Peltigerales) in East African Montane Ecosystems" Journal of Fungi 9, no. 2: 246. https://doi.org/10.3390/jof9020246
APA StyleKaasalainen, U., Kirika, P. M., Mollel, N. P., Hemp, A., & Rikkinen, J. (2023). The Lichen Genus Sticta (Lobariaceae, Peltigerales) in East African Montane Ecosystems. Journal of Fungi, 9(2), 246. https://doi.org/10.3390/jof9020246