
Biodistribution and Adjuvant Effect of an Intranasal Vaccine Based on Chitosan Nanoparticles against Paracoccidioidomycosis

 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
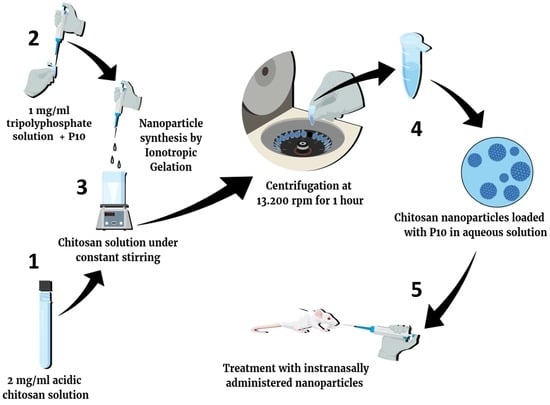
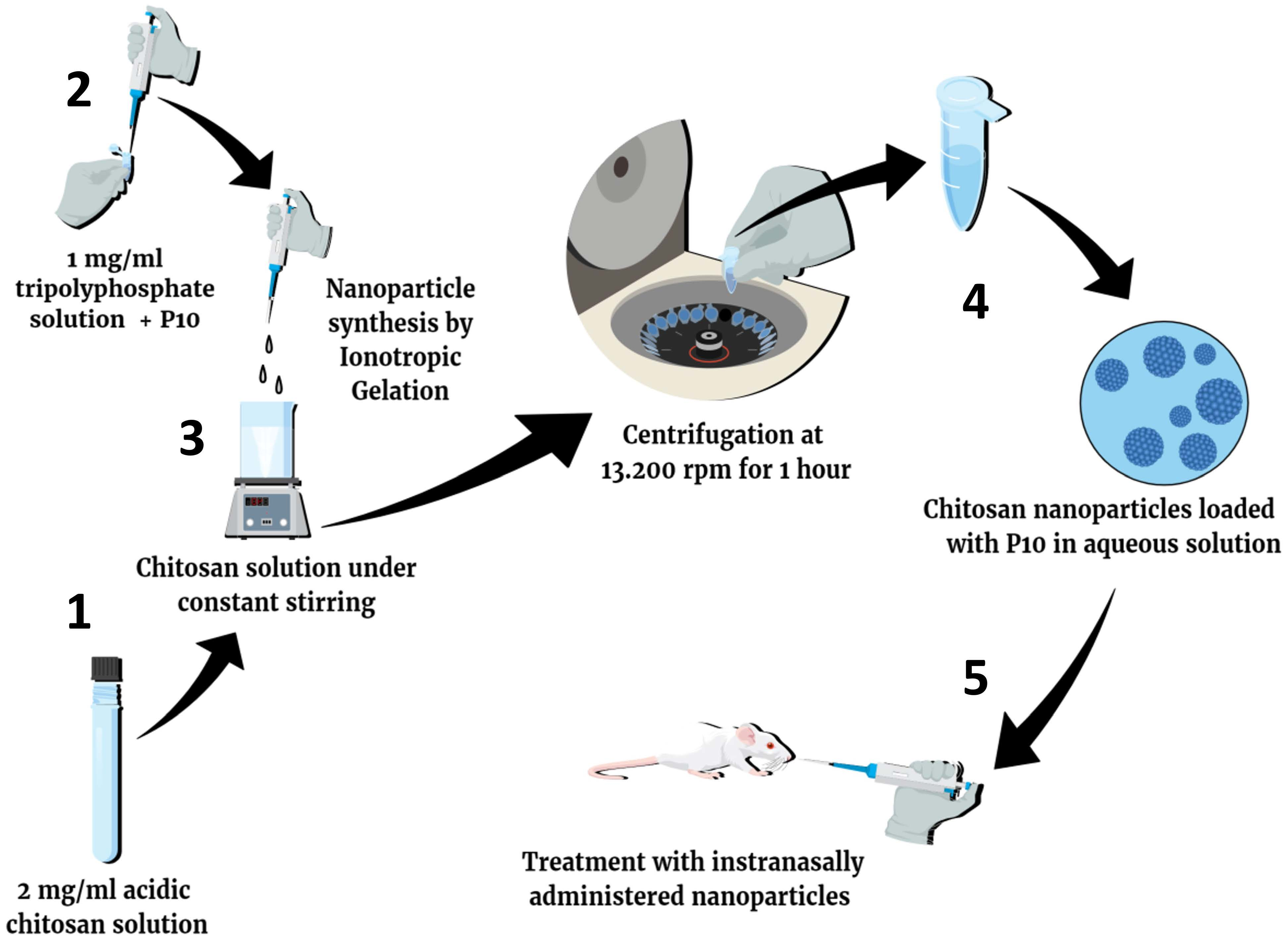
2.1. Non-Fluorescent Nanoparticles Preparation
2.2. Preparation of Fluorescent Nanoparticles
2.3. Physical-Chemical Characterization of the Nanoparticles
2.4. Animal Approvals
2.5. Experimental Design Used in the Biodistribution Protocol
2.6. Nanoparticles Phagocytosis Assay
2.7. Yeast Cells
2.8. Experimental Design Used in the Infection Protocol
2.9. Intratracheal Infection and Immunization
2.10. Antifungal and Cytokine Evaluation
2.11. Statistical Analysis
3. Results
3.1. Nanoparticles Preparation and Characterization
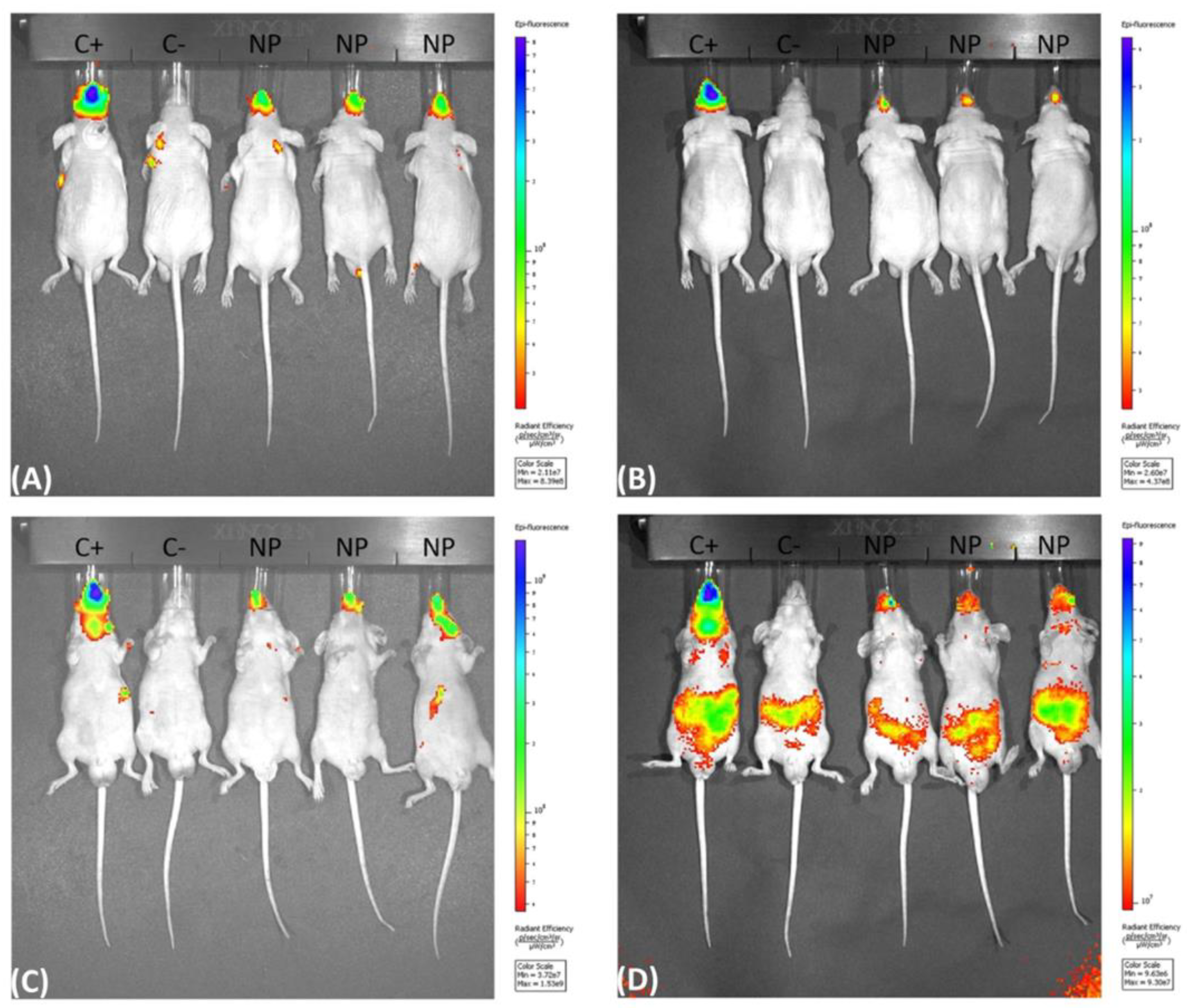
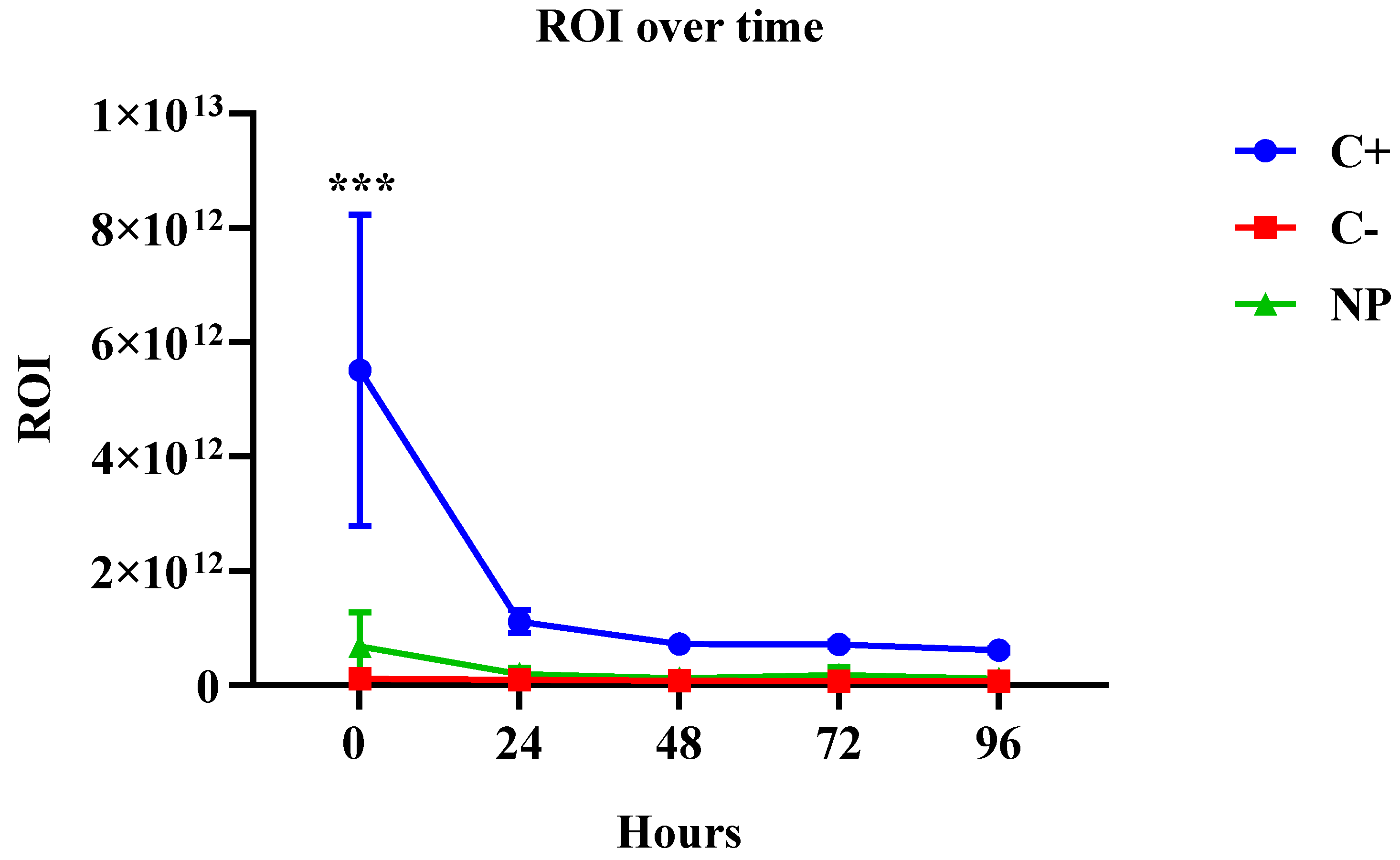
3.2. Biodistribution using the IVIS Spectrum
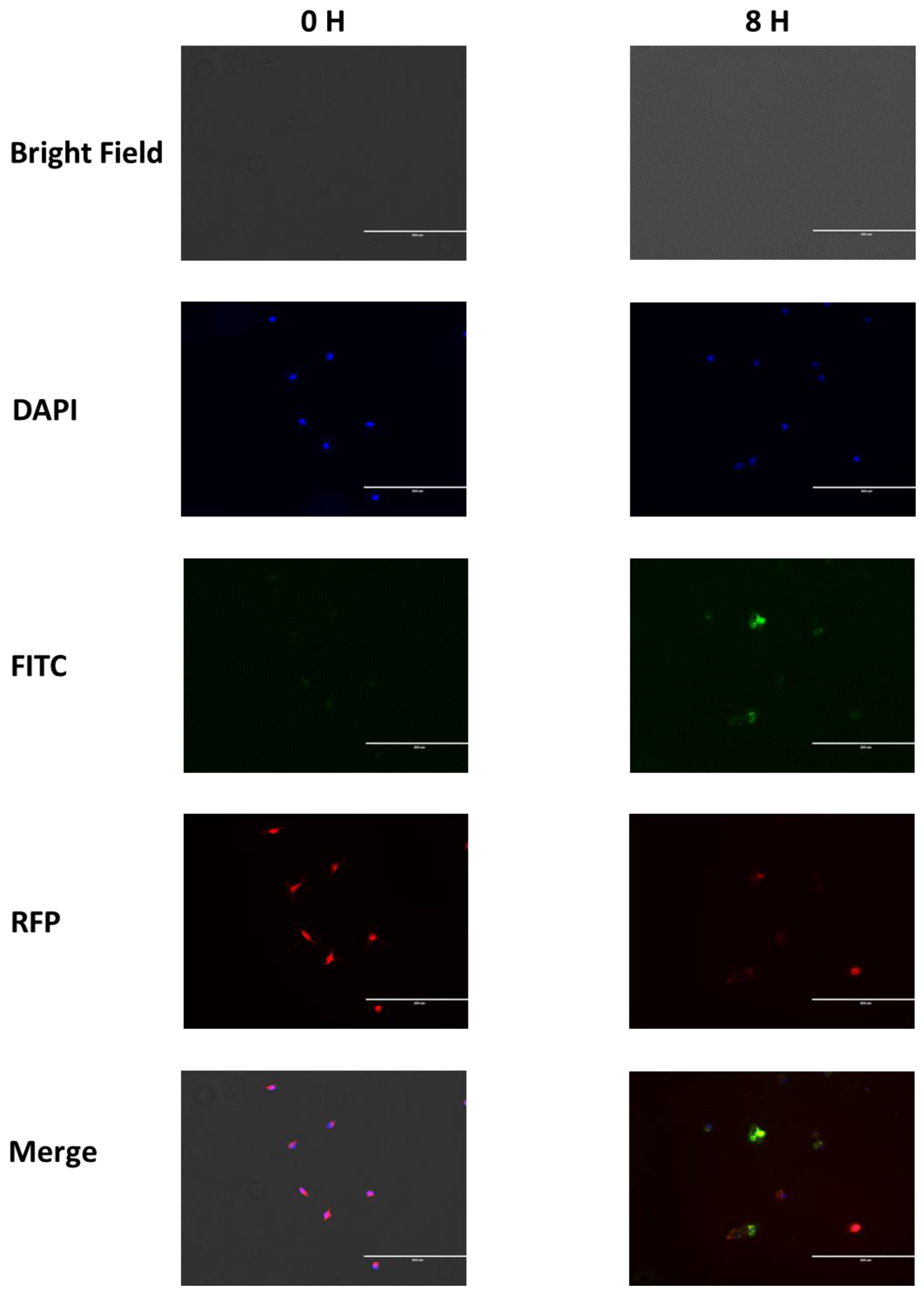
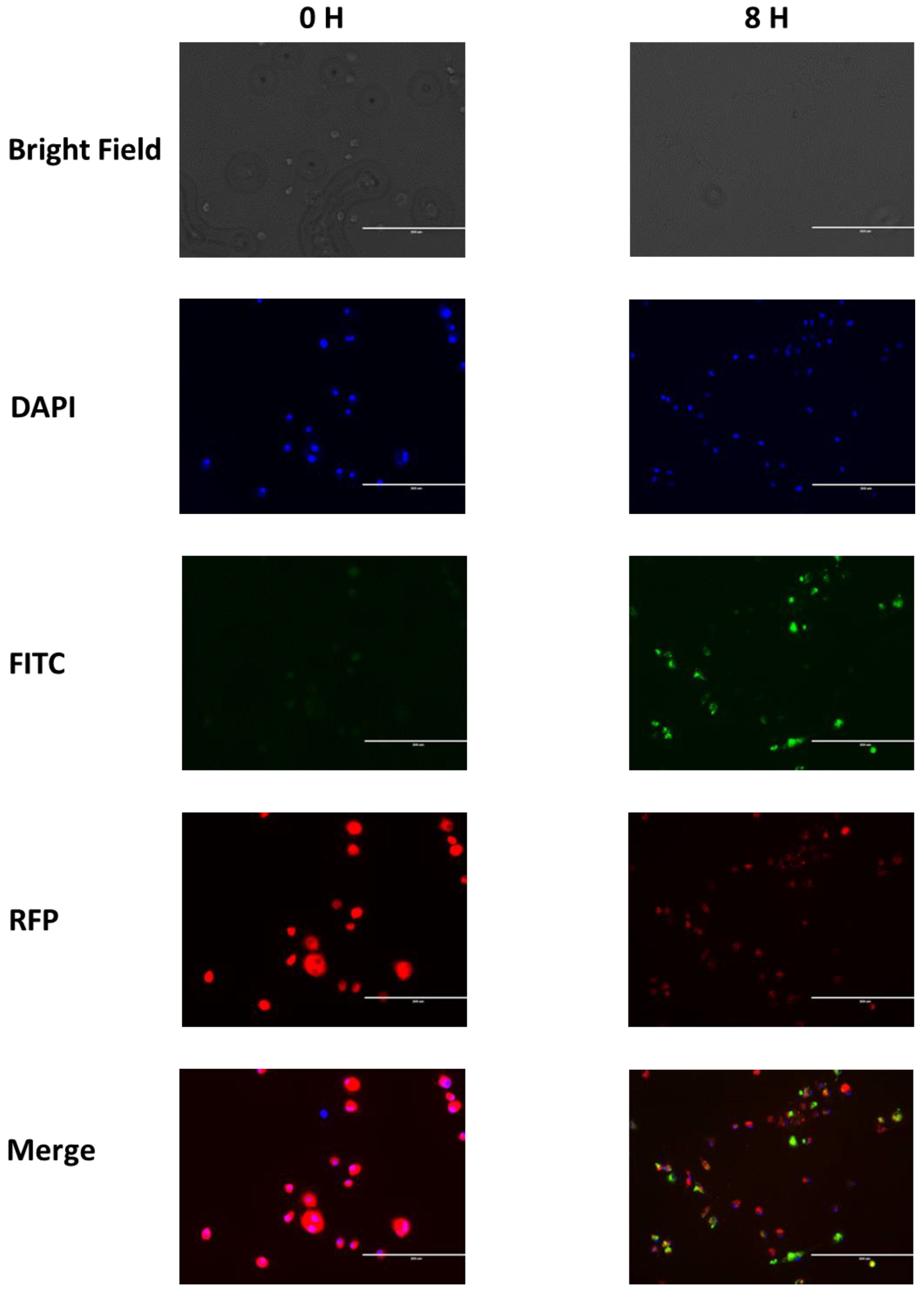
3.3. Nanoparticles Phagocytosis Assay
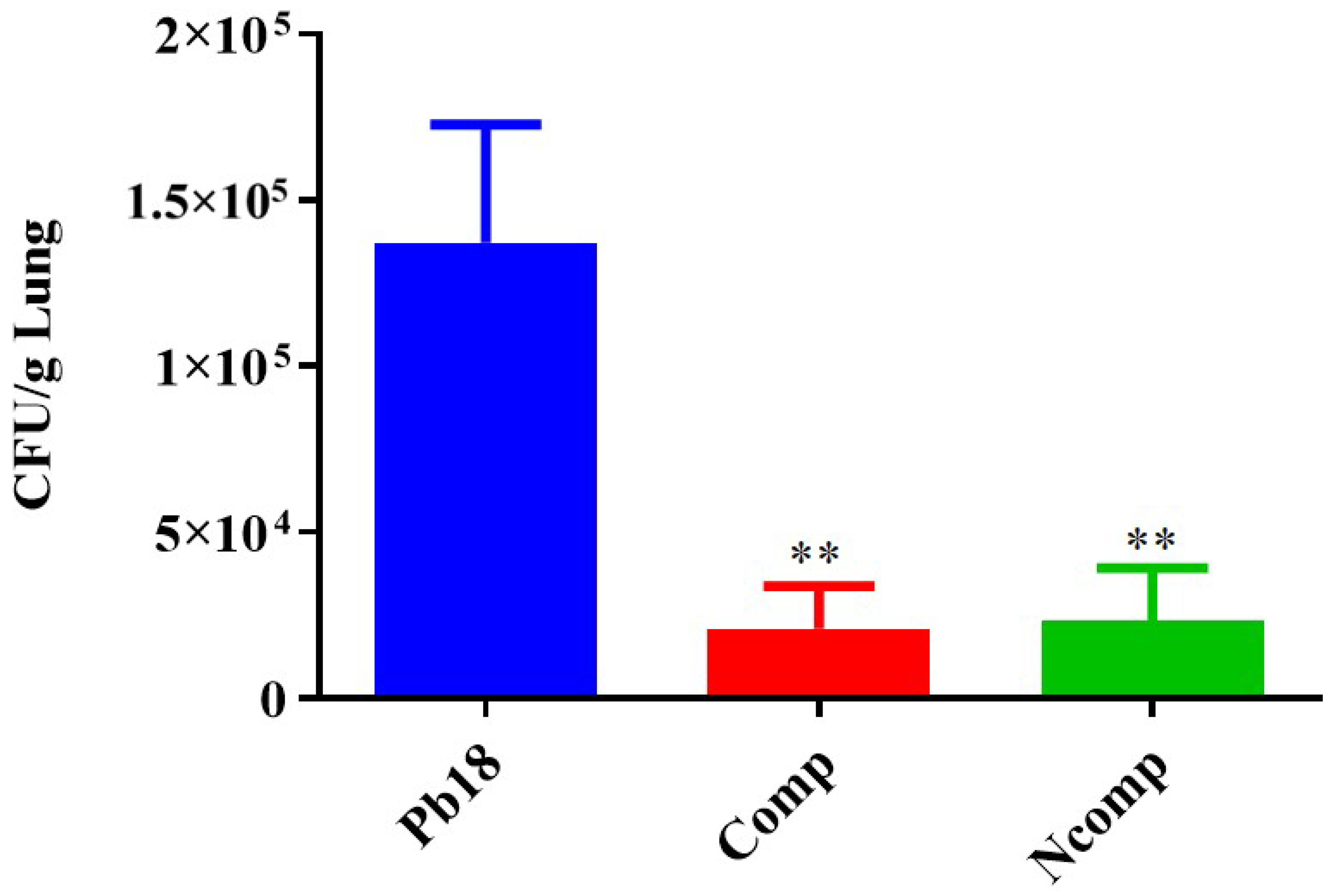
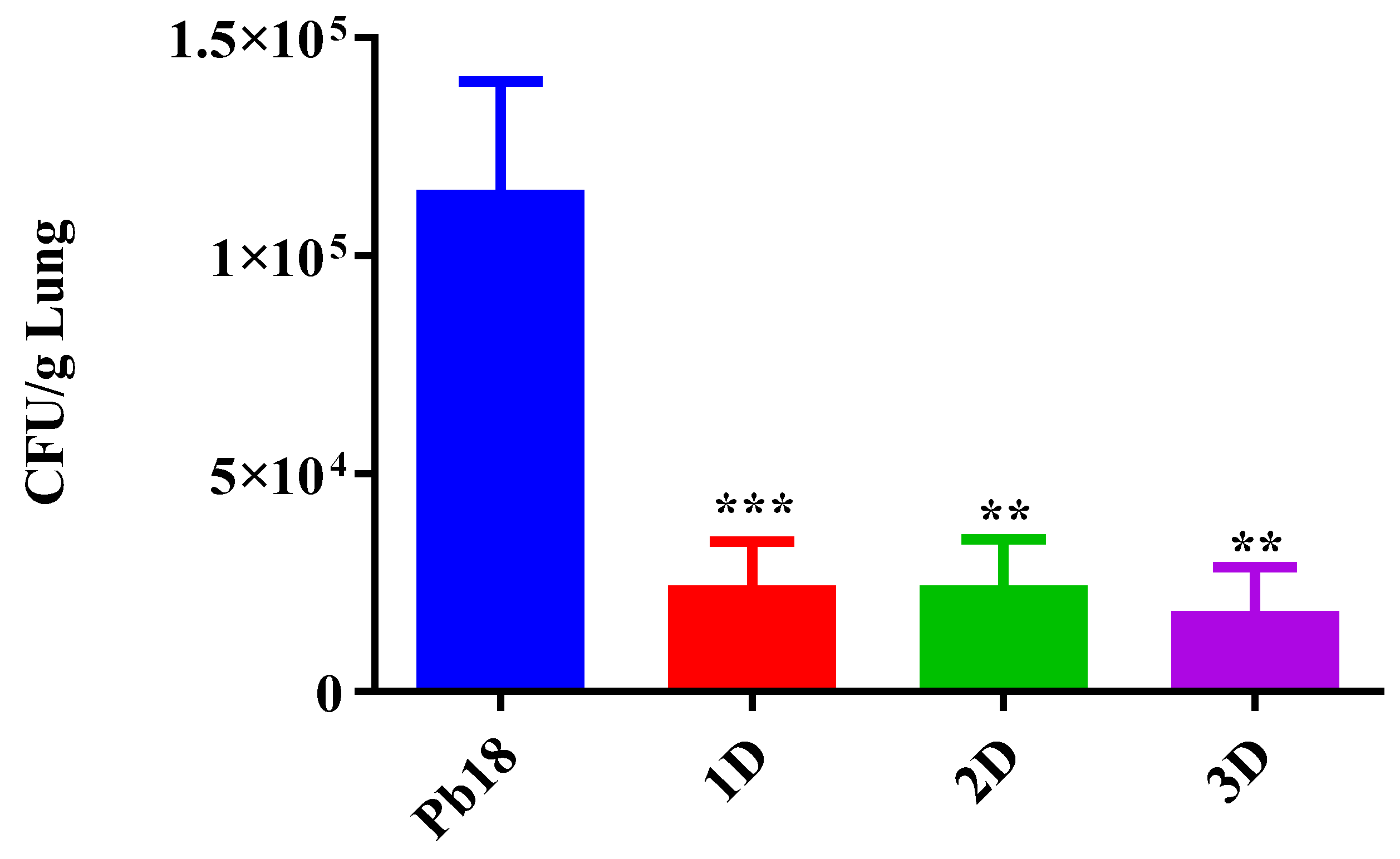
3.4. CFU from the Complexed or Co-Administered Nanoparticle Immunizations
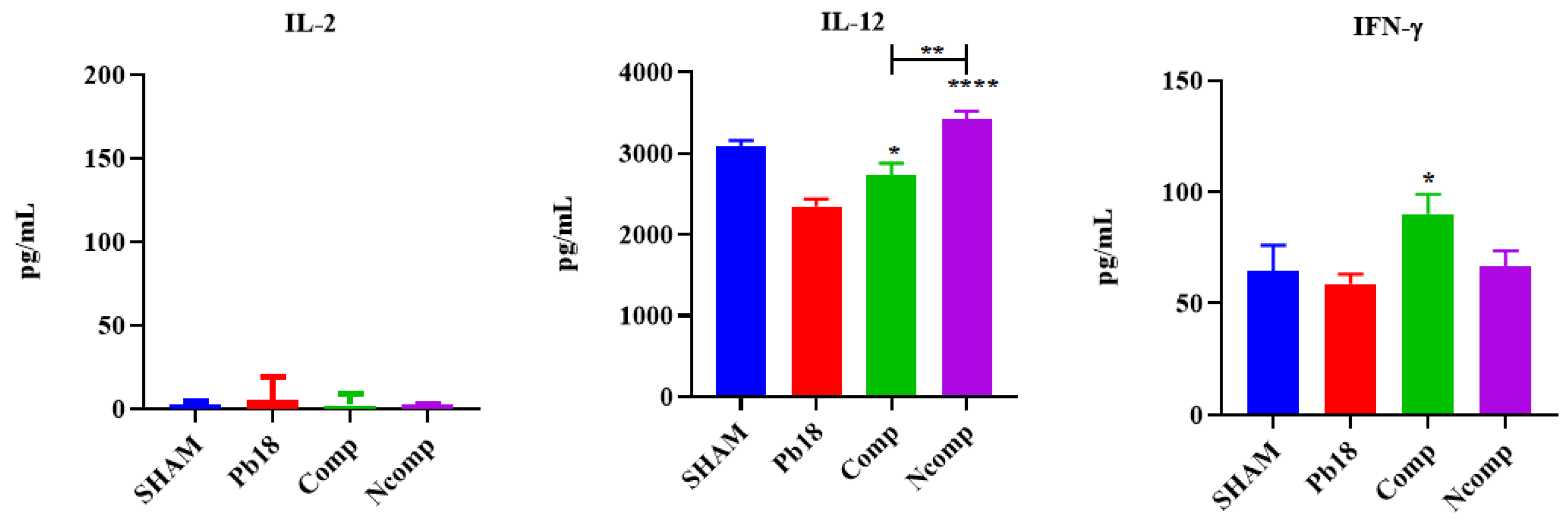
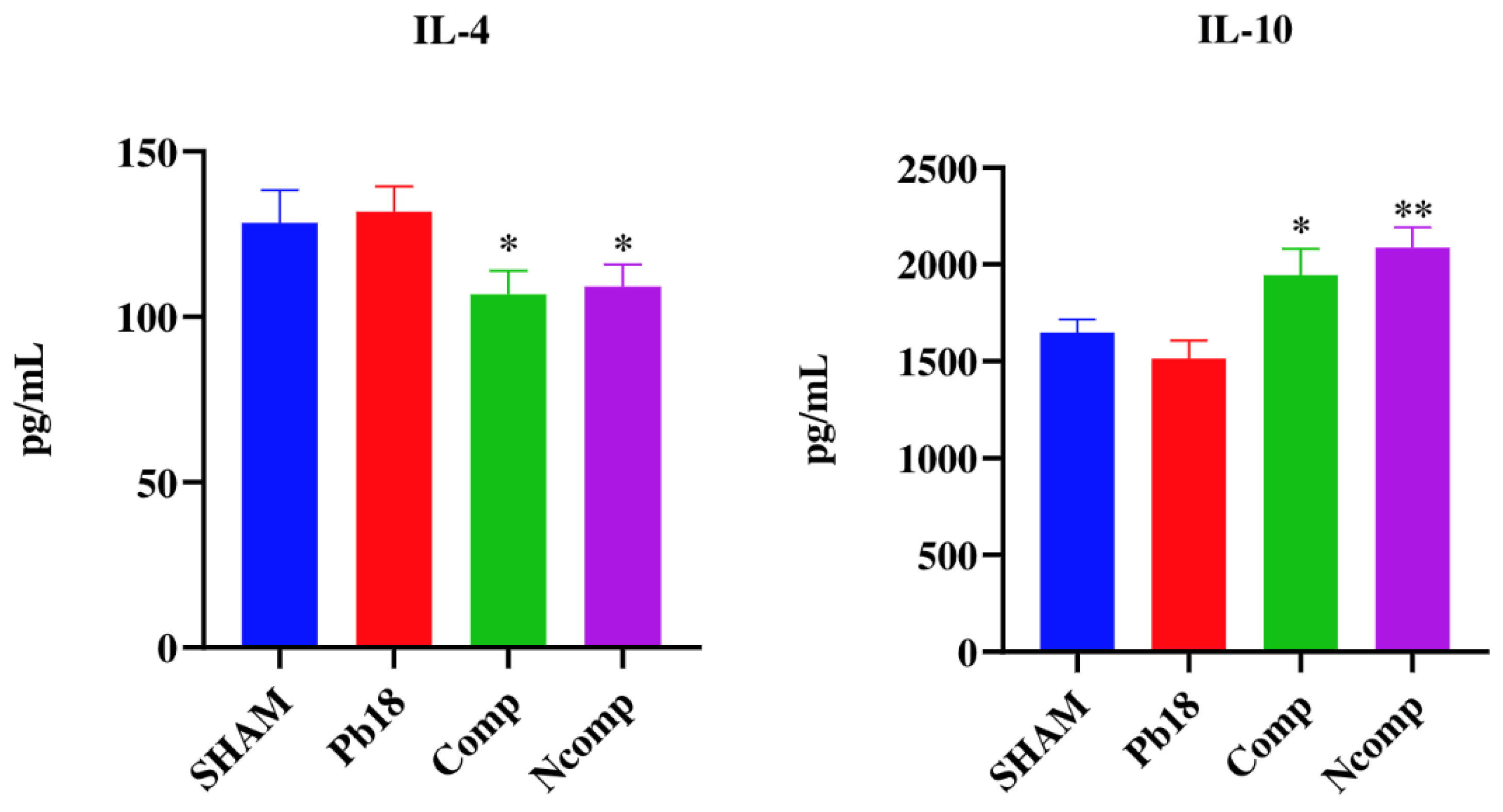
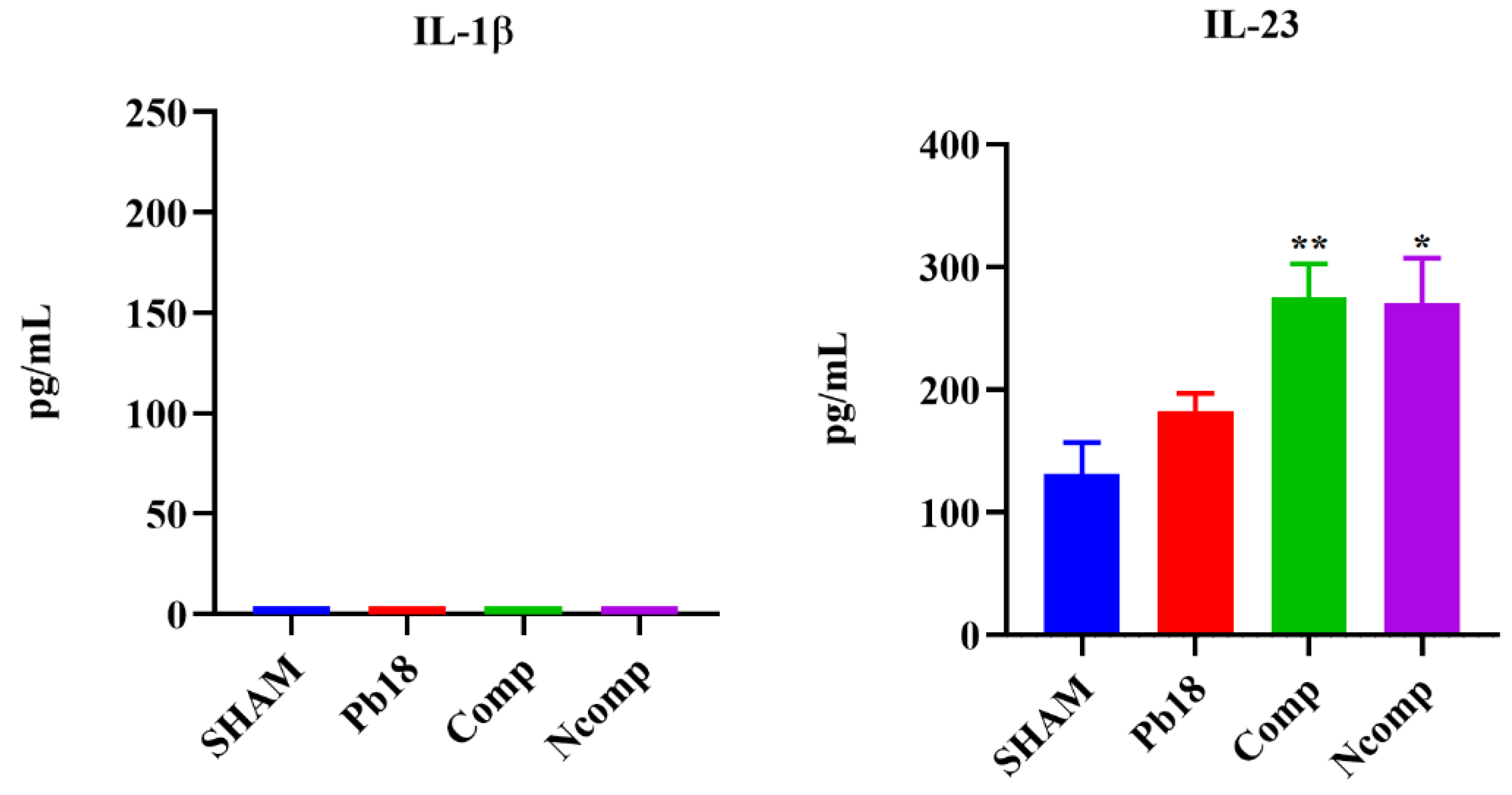
3.5. Cytokines from the Complexed or Co-Administered Nanoparticle Immunizations
3.6. CFU from the Immunization with Different Doses of Nanoparticles
3.7. Cytokines from the Immunization with Different Doses of Nanoparticles
4. Discussion
5. Conclusion
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Travassos, L.R.; Taborda, C.P. New advances in the development of a vaccine against paracoccidioidomycosis. Front. Microbiol. 2012, 3, 212. [Google Scholar] [CrossRef]
- Shikanai-yasuda, M.A.; De Queiroz, F.; Filho, T.; Mendes, R.P.; Colombo, A.L.; Moretti, M.L.; Consultores, D. Consenso em paracoccidioidomicose Guideliness in paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2006, 39, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; de Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; do Valle, A.C.F.; Bagagli, E.; Benard, G.; et al. Brazilian guidelines for the clinical management of paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2017, 50, 715–740. [Google Scholar] [CrossRef] [PubMed]
- Turissini, D.A.; Gomez, O.M.; Teixeira, M.M.; McEwen, J.G.; Matute, D.R. Species boundaries in the human pathogen Paracoccidioides. Fungal Genet. Biol. 2017, 106, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.C.O.; Taborda, C.P. Epidemiology of Dimorphic Fungi. In Encyclopedia of Mycology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 613–623. ISBN 9780323851800. [Google Scholar]
- Taborda, C.P.; Travassos, L.R.; Benard, G. Paracoccidioidomycosis. In Encyclopedia of Mycology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 654–675. [Google Scholar]
- Benard, G. An overview of the immunopathology of human paracoccidioidomycosis. Mycopathologia 2008, 165, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Bocca, A.L.; Amaral, A.C.; Teixeira, M.M.; Sato, P.K.; Sato, P.; Shikanai-Yasuda, M.A.; Soares Felipe, M.S. Paracoccidioidomycosis: Eco-epidemiology, taxonomy and clinical and therapeutic issues. Future Microbiol. 2013, 8, 1177–1191. [Google Scholar] [CrossRef]
- Amaral, A.C.; Bocca, A.L.; Ribeiro, A.M.; Nunes, J.; Peixoto, D.L.G.G.; Simioni, A.R.; Primo, F.L.; Lacava, Z.G.M.M.; Bentes, R.; Titze-de-Almeida, R.; et al. Amphotericin B in poly(lactic-co-glycolic acid) (PLGA) and dimercaptosuccinic acid (DMSA) nanoparticles against paracoccidioidomycosis. J. Antimicrob. Chemother. 2009, 63, 526–533. [Google Scholar] [CrossRef]
- Taborda, C.P.; Urán, M.E.; Nosanchuk, J.D.; Travassos, L.R. Paracoccidioidomycosis: Challenges in the development of a vaccine against an endemic mycosis in the americas. Rev. Inst. Med. Trop. Sao Paulo 2015, 57, 21–24. [Google Scholar] [CrossRef]
- Taborda, C.P.; Juliano, M.A.; Puccia, R.; Franco, M.; Travassos, L.R. Mapping of the T-cell epitope in the major 43-kilodalton glycoprotein of Paracoccidioides brasiliensis which induces a Th-1 response protective against fungal infection in BALB/c mice. Infect. Immun. 1998, 66, 786–793. [Google Scholar] [CrossRef]
- Amaral, A.C.; Marques, A.F.; Muñoz, J.E.; Bocca, A.L.; Simioni, A.R.; Tedesco, A.C.; Morais, P.C.; Travassos, L.R.; Taborda, C.P.; Felipe, M.S.S. Poly(lactic acid-glycolic acid) nanoparticles markedly improve immunological protection provided by peptide P10 against murine paracoccidioidomycosis. Br. J. Pharmacol. 2010, 159, 1126–1132. [Google Scholar] [CrossRef] [Green Version]
- Jannuzzi, G.P.; Souza, N.d.A.; Françoso, K.S.; Pereira, R.H.; Santos, R.P.; Kaihami, G.H.; de Almeida, J.R.F.; Batista, W.L.; Amaral, A.C.; Maranhão, A.Q.; et al. Therapeutic treatment with scFv–PLGA nanoparticles decreases pulmonary fungal load in a murine model of paracoccidioidomycosis. Microbes Infect. 2018, 20, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues Dos Santos Junior, S.; Kelley Lopes da Silva, F.; Santos Dias, L.; Oliveira Souza, A.C.; Valdemir de Araujo, M.; Buffoni Roque da Silva, L.; Travassos, L.R.; Correa Amaral, A.; Taborda, C.P. Intranasal Vaccine Using P10 Peptide Complexed within Chitosan Polymeric Nanoparticles as Experimental Therapy for Paracoccidioidomycosis in Murine Model. J. Fungi 2020, 6, 160. [Google Scholar] [CrossRef]
- Kischkel, B.; Rossi, S.A.; Santos, S.R.; Nosanchuk, J.D.; Travassos, L.R.; Taborda, C.P. Therapies and Vaccines Based on Nanoparticles for the Treatment of Systemic Fungal Infections. Front. Cell. Infect. Microbiol. 2020, 10, 463. [Google Scholar] [CrossRef]
- dos Santos Junior, S.R.; Amaral, A.C.; Taborda, C.P. Application of Nanoparticles to Invasive Fungal Infections. In Nanotechnology for Infectious Diseases; Springer: Singapore, 2022; pp. 151–173. [Google Scholar]
- Van der Lubben, I.M.; Verhoef, J.C.; Borchard, G.; Junginger, H.E. Chitosan for mucosal vaccination. Adv. Drug Deliv. Rev. 2001, 52, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Zhang, Z.; Lv, J.; Zhou, P.; Hu, W.; Fang, Y.; Chen, H.; Liu, X.; Shao, J.; Zhao, F.; et al. Induction of mucosal immune responses and protection of cattle against direct-contact challenge by intranasal delivery with foot-and-mouth disease virus antigen mediated by nanoparticles. Int. J. Nanomed. 2014, 9, 5603–5618. [Google Scholar] [CrossRef] [PubMed]
- Kyd, J.M.; Foxwell, A.R.; Cripps, A.W. Mucosal immunity in the lung and upper airway. Vaccine 2001, 19, 2527–2533. [Google Scholar] [CrossRef]
- Xia, Y.; Fan, Q.; Hao, D.; Wu, J.; Ma, G.; Su, Z. Chitosan-based mucosal adjuvants: Sunrise on the ocean. Vaccine 2015, 33, 5997–6010. [Google Scholar] [CrossRef]
- Chadwick, S.; Kriegel, C.; Amiji, M. Nanotechnology solutions for mucosal immunization. Adv. Drug Deliv. Rev. 2010, 62, 394–407. [Google Scholar] [CrossRef]
- Banerjee, T.; Mitra, S.; Kumar Singh, A.; Kumar Sharma, R.; Maitra, A. Preparation, characterization and biodistribution of ultrafine chitosan nanoparticles. Int. J. Pharm. 2002, 243, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Sonin, D.; Pochkaeva, E.; Zhuravskii, S.; Postnov, V.; Korolev, D.; Vasina, L.; Kostina, D.; Mukhametdinova, D.; Zelinskaya, I.; Skorik, Y.; et al. Biological Safety and Biodistribution of Chitosan Nanoparticles. Nanomaterials 2020, 10, 810. [Google Scholar] [CrossRef] [Green Version]
- Raj, P.M.; Raj, R.; Kaul, A.; Mishra, A.K.; Ram, A. Biodistribution and targeting potential assessment of mucoadhesive chitosan nanoparticles designed for ulcerative colitis via scintigraphy. RSC Adv. 2018, 8, 20809–20821. [Google Scholar] [CrossRef] [PubMed]
- Suri, S.; Ruan, G.; Winter, J.; Schmidt, C.E. Microparticles and Nanoparticles. In Biomaterials Science: An Introduction to Materials in Medicine, 3rd ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 360–388. [Google Scholar] [CrossRef]
- Buzea, C.; Pacheco, I. Toxicity of nanoparticles. In Nanotechnology in Eco-Efficient Construction: Materials, Processes and Applications; Woodhead Publishing: Soston, UK, 2018; pp. 705–754. [Google Scholar] [CrossRef]
- Kim, K.; Kim, J.H.; Park, H.; Kim, Y.S.; Park, K.; Nam, H.; Lee, S.; Park, J.H.; Park, R.W.; Kim, I.S.; et al. Tumor-homing multifunctional nanoparticles for cancer theragnosis: Simultaneous diagnosis, drug delivery, and therapeutic monitoring. J. Control. Release 2010, 146, 219–227. [Google Scholar] [CrossRef]
- Huang, M.; Ma, Z.; Khor, E.; Lim, L.Y. Uptake of FITC-chitosan nanoparticles by A549 cells. Pharm. Res. 2002, 19, 1488–1494. [Google Scholar] [CrossRef]
- Herb, M.; Farid, A.; Gluschko, A.; Krönke, M.; Schramm, M. Highly Efficient Transfection of Primary Macrophages with In Vitro Transcribed mRNA. J. Vis. Exp. 2019, 2019, e60143. [Google Scholar] [CrossRef]
- Zhang, X.; Goncalves, R.; Mosser, D.M. The Isolation and Characterization of Murine Macrophages. Curr. Protoc. Immunol. 2008, 83, 14. [Google Scholar] [CrossRef]
- Busch, C.; Favret, J.; Geirsdóttir, L.; Molawi, K.; Sieweke, M. Isolation and Long-term Cultivation of Mouse Alveolar Macrophages. Bio-Protocol 2019, 9, e3302. [Google Scholar] [CrossRef] [PubMed]
- Netto, C.F.; Vegas, V.S.; Sciannaméa, I.M.; Guarnieri, D.B. The polysaccharidic antigen from Paracoccidioides brasiliensis. Study of the time of cultivation necessary for the preparation of the antigen. Rev. Inst. Med. Trop. Sao Paulo 1969, 11, 177–181. [Google Scholar] [PubMed]
- Del Rosal, B.; Villa, I.; Jaque, D.; Sanz-Rodríguez, F. In vivo autofluorescence in the biological windows: The role of pigmentation. J. Biophotonics 2016, 9, 1059–1067. [Google Scholar] [CrossRef]
- Kwon, S.; Davies-Venn, C.; Sevick-Muraca, E.M. In vivo dynamic imaging of intestinal motions using diet-related autofluorescence. Neurogastroenterol. Motil. 2012, 24, 494–497. [Google Scholar] [CrossRef] [PubMed]
- del Rosal, B.; Benayas, A. Strategies to Overcome Autofluorescence in Nanoprobe-Driven In Vivo Fluorescence Imaging. Small Methods 2018, 2, 1800075. [Google Scholar] [CrossRef]
- Bhaumik, S.; Depuy, J.; Klimash, J. Strategies to minimize background autofluorescence in live mice during noninvasive fluorescence optical imaging. Lab Anim. 2007, 36, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Izawa, K.; Kiryu, S.; Tojo, A.; Ohtomo, K. Diet and Abdominal Autofluorescence Detected by In Vivo Fluorescence Imaging of Living Mice. Mol. Imaging 2008, 7, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The Still Underestimated Problem of Fungal Diseases Worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef]
- Mayorga, O.; Munoz, J.E.; Travassos, L.R.; Carlos, P. The role of adjuvants in therapeutic protection against paracoccidioidomycosis after immunization with the P10 peptide. Front. Microbiol. 2012, 3, 154. [Google Scholar] [CrossRef]
- Shim, S.; Soh, S.H.; Bin Im, Y.; Park, H.E.; Cho, C.S.; Kim, S.; Yoo, H.S. Elicitation of Th1/Th2 related responses in mice by chitosan nanoparticles loaded with Brucella abortus malate dehydrogenase, outer membrane proteins 10 and 19. Int. J. Med. Microbiol. 2020, 310, 151362. [Google Scholar] [CrossRef]
- Rençber, S.; Karavana, S.Y.; Yılmaz, F.F.; Eraç, B.; Nenni, M.; Ozbal, S.; Pekçetin, Ç.; Gurer-Orhan, H.; Hoşgör Limoncu, M.; Güneri, P.; et al. Development, characterization, and in vivo assessment of mucoadhesive nanoparticles containing fluconazole for the local treatment of oral candidiasis. Int. J. Nanomed. 2016, 11, 2641. [Google Scholar] [CrossRef]
- Southam, D.S.; Dolovich, M.; O’Byrne, P.M.; Inman, M.D. Distribution of intranasal instillations in mice: Effects of volume, time, body position, and anesthesia. Am. J. Physiol. Cell. Mol. Physiol. 2002, 282, L833–L839. [Google Scholar] [CrossRef]
- Miller, M.A.; Stabenow, J.M.; Parvathareddy, J.; Wodowski, A.J.; Fabrizio, T.P.; Bina, X.R.; Zalduondo, L.; Bina, J.E. Visualization of Murine Intranasal Dosing Efficiency Using Luminescent Francisella tularensis: Effect of Instillation Volume and Form of Anesthesia. PLoS ONE 2012, 7, e31359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visweswaraiah, A. Tracking the tissue distribution of marker dye following intranasal delivery in mice and chinchillas: A multifactorial analysis of parameters affecting nasal retention. Vaccine 2002, 20, 3209–3220. [Google Scholar] [CrossRef]
- Swanson, K.V.; Deng, M.; Ting, J.P.Y. The NLRP3 inflammasome: Molecular activation and regulation to therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef]
- Carroll, E.C.; Jin, L.; Mori, A.; Muñoz-Wolf, N.; Oleszycka, E.; Moran, H.B.T.; Mansouri, S.; McEntee, C.P.; Lambe, E.; Agger, E.M.; et al. The Vaccine Adjuvant Chitosan Promotes Cellular Immunity via DNA Sensor cGAS-STING-Dependent Induction of Type I Interferons. Immunity 2016, 44, 597–608. [Google Scholar] [CrossRef]
- Dubensky, T.W.; Kanne, D.B.; Leong, M.L. Rationale, progress and development of vaccines utilizing STING-activating cyclic dinucleotide adjuvants. Ther. Adv. Vaccines 2013, 1, 131–143. [Google Scholar] [CrossRef]
- Villiers, C.; Chevallet, M.; Diemer, H.; Couderc, R.; Freitas, H.; Van Dorsselaer, A.; Marche, P.N.; Rabilloud, T. From secretome analysis to immunology: Chitosan induces major alterations in the activation of dendritic cells via a TLR4-dependent mechanism. Mol. Cell. Proteom. 2009, 8, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Bueter, C.L.; Lee, C.K.; Wang, J.P.; Ostroff, G.R.; Specht, C.A.; Levitz, S.M. Spectrum and Mechanisms of Inflammasome Activation by Chitosan. J. Immunol. 2014, 192, 5943–5951. [Google Scholar] [CrossRef] [PubMed]
- Bueter, C.L.; Lee, C.K.; Rathinam, V.A.K.; Healy, G.J.; Taron, C.H.; Specht, C.A.; Levitz, S.M. Chitosan but not chitin activates the inflammasome by a mechanism dependent upon phagocytosis. J. Biol. Chem. 2011, 286, 35447–35455. [Google Scholar] [CrossRef]
- Mori, A.; Oleszycka, E.; Sharp, F.A.; Coleman, M.; Ozasa, Y.; Singh, M.; O’Hagan, D.T.; Tajber, L.; Corrigan, O.I.; McNeela, E.A.; et al. The vaccine adjuvant alum inhibits IL-12 by promoting PI3 kinase signaling while chitosan does not inhibit IL-12 and enhances Th1 and Th17 responses. Eur. J. Immunol. 2012, 42, 2709–2719. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, E.C.; Ward, R.W. Mucosal vaccines—Fortifying the frontiers. Nat. Rev. Immunol. 2022, 22, 236–250. [Google Scholar] [CrossRef]
- Holmgren, J.; Czerkinsky, C. Mucosal immunity and vaccines. Nat. Med. 2005, 11, S45–S53. [Google Scholar] [CrossRef]
- Marttin, E.; Schipper, N.G.M.; Coos Verhoef, J.; Merkus, F.W.H.M. Nasal mucociliary clearance as a factor in nasal drug delivery. Adv. Drug Deliv. Rev. 1998, 29, 13–38. [Google Scholar] [CrossRef]
- Ogra, P.L.; Faden, H.; Welliver, R.C. Vaccination Strategies for Mucosal Immune Responses. Clin. Microbiol. Rev. 2001, 14, 430–445. [Google Scholar] [CrossRef]
- Kolls, J.K.; Khader, S.A. The role of Th17 cytokines in primary mucosal immunity. Cytokine Growth Factor Rev. 2010, 21, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Khader, S.A.; Gaffen, S.L.; Kolls, J.K. Th17 cells at the crossroads of innate and adaptive immunity against infectious diseases at the mucosa. Mucosal Immunol. 2009, 2, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Burger, E. Paracoccidioidomycosis Protective Immunity. J. Fungi 2021, 7, 137. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.M.; Bocca, A.L.; Amaral, A.C.; Faccioli, L.H.; Galetti, F.C.S.; Zárate-Bladés, C.R.; Figueiredo, F.; Silva, C.L.; Felipe, M.S.S. DNAhsp65 vaccination induces protection in mice against Paracoccidioides brasiliensis infection. Vaccine 2009, 27, 606–613. [Google Scholar] [CrossRef]
- Sojka, D.K.; Bruniquel, D.; Schwartz, R.H.; Singh, N.J. IL-2 Secretion by CD4 + T Cells In Vivo Is Rapid, Transient, and Influenced by TCR-Specific Competition. J. Immunol. 2004, 172, 6136–6143. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fluorescent Intensity of the Region of Interest (ROI) | |||
|---|---|---|---|
| Hours | C+ | C− | NP |
| 0 | 5.5 × 1012 | 1.0 × 1011 | 6.7 × 1011 |
| 24 | 1.1 × 1012 | 9.3 × 1010 | 1.9 × 1011 |
| 48 | 7.2 × 1011 | 7.6 × 1010 | 1.1 × 1011 |
| 72 | 7.1 × 1011 | 7.3 × 1010 | 1.7 × 1011 |
| 96 | 6.0 × 1011 | 7.2 × 1010 | 1.1 × 1011 |
| Cytokines | |||||||
|---|---|---|---|---|---|---|---|
| Groups | Th1 | Th2 | Th17 | ||||
| IL-2 | IL-12 | IFN-γ | IL-4 | IL-10 | IL-1β | IL-23 | |
| Sham | 0 | 2605 ± 480.1 | 80.96 ± 22 | 148.4 ± 57.84 | 1965 ± 297.8 | 0 | 339.4 ± 208.9 |
| Pb 18 | 0 | 2491 ± 502.8 | 66.80 ± 9.9 | 140.4 ± 37.61 | 2000 ± 387.7 | 0 | 117.0 ± 107.2 |
| 1D | 0 | 3168 ± 438.5 *** | 92.84 ± 29.38 * | 193.6 ± 47.72 ** | 2418 ± 299.8 ** | 0 | 232.4 ± 87.20 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos Júnior, S.R.D.; Barbalho, F.V.; Nosanchuk, J.D.; Amaral, A.C.; Taborda, C.P. Biodistribution and Adjuvant Effect of an Intranasal Vaccine Based on Chitosan Nanoparticles against Paracoccidioidomycosis. J. Fungi 2023, 9, 245. https://doi.org/10.3390/jof9020245
Santos Júnior SRD, Barbalho FV, Nosanchuk JD, Amaral AC, Taborda CP. Biodistribution and Adjuvant Effect of an Intranasal Vaccine Based on Chitosan Nanoparticles against Paracoccidioidomycosis. Journal of Fungi. 2023; 9(2):245. https://doi.org/10.3390/jof9020245
Chicago/Turabian StyleSantos Júnior, Samuel Rodrigues Dos, Filipe Vieira Barbalho, Joshua D. Nosanchuk, Andre Correa Amaral, and Carlos Pelleschi Taborda. 2023. "Biodistribution and Adjuvant Effect of an Intranasal Vaccine Based on Chitosan Nanoparticles against Paracoccidioidomycosis" Journal of Fungi 9, no. 2: 245. https://doi.org/10.3390/jof9020245
APA StyleSantos Júnior, S. R. D., Barbalho, F. V., Nosanchuk, J. D., Amaral, A. C., & Taborda, C. P. (2023). Biodistribution and Adjuvant Effect of an Intranasal Vaccine Based on Chitosan Nanoparticles against Paracoccidioidomycosis. Journal of Fungi, 9(2), 245. https://doi.org/10.3390/jof9020245