
Benzimidazole Derivatives Suppress Fusarium Wilt Disease via Interaction with ERG6 of Fusarium equiseti and Activation of the Antioxidant Defense System of Pepper Plants
 , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
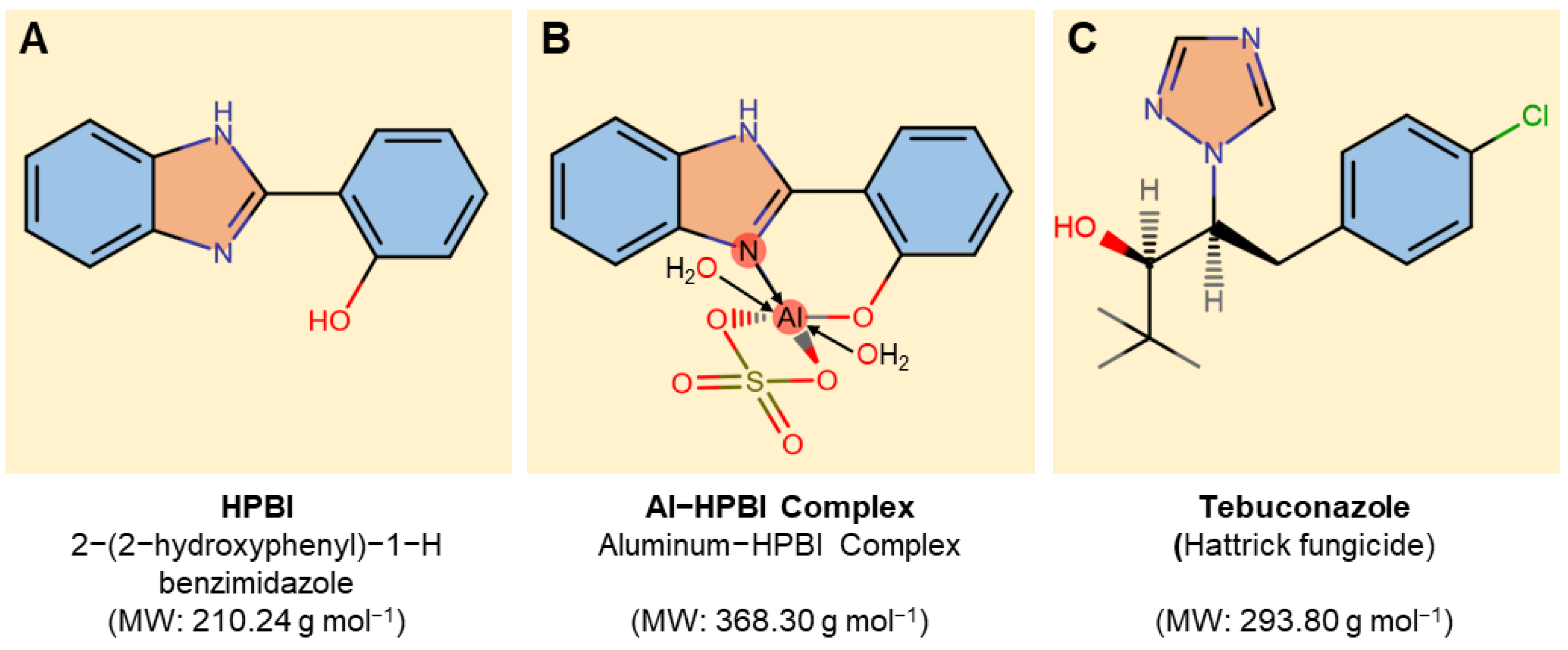
2.1. Tested Compounds
2.2. Fungal Isolate
2.3. In Vitro Antifungal Activity
2.4. Greenhouse Experiment, Disease Assessment, and Growth Parameters
2.5. In Situ Histochemical Localization of Hydrogen Peroxide (H2O2) and Superoxide Anion (O2•−)
2.6. Total Soluble Phenolic and Flavonoid Compounds
2.7. Enzymatic Activity
2.8. Gene Expression Analysis
2.9. In Silico Analysis of EGR6 from F. equiseti
2.9.1. Protein–Protein BLAST (BLASTp) Analysis
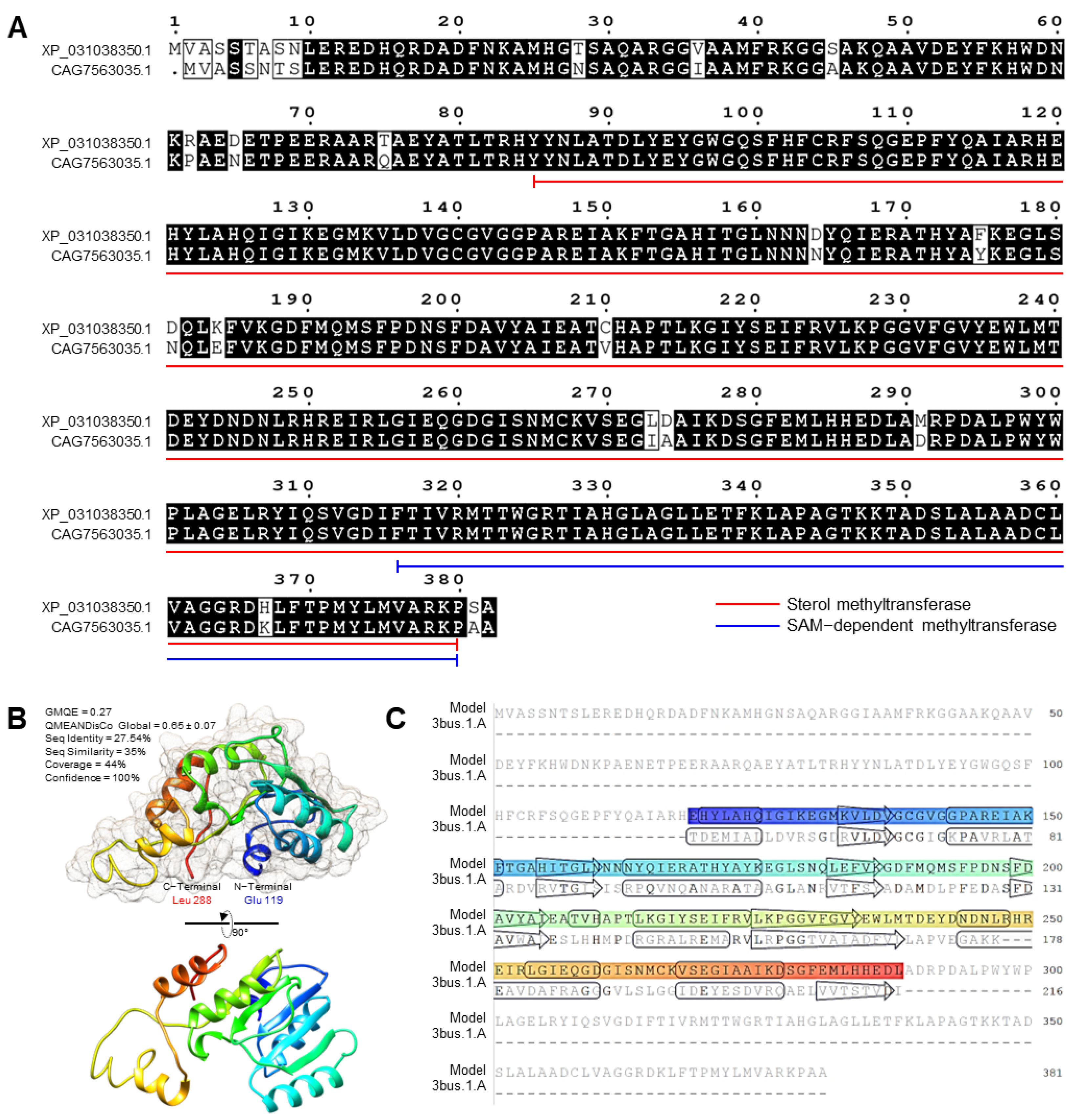
2.9.2. Multiple Sequence Alignment Analysis
2.9.3. Conserved Domains and Theoretical pI/Mw
2.9.4. Three-Dimensional (3D) Structure Modeling
2.10. Molecular Docking Analysis
2.11. Statistical Analyses
3. Results
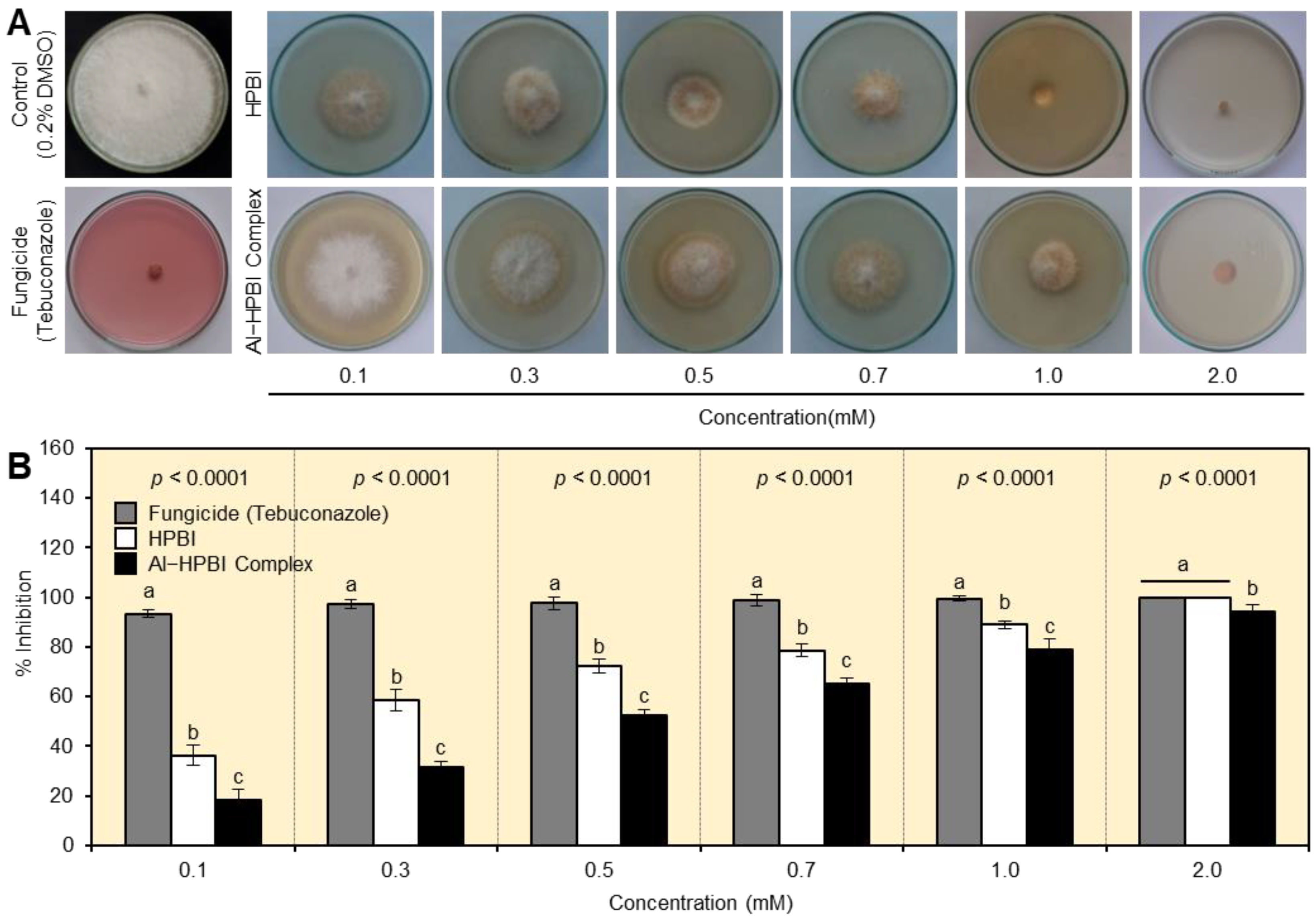
3.1. HPBI and Al−HPBI Complex Suppress the Mycelial Growth of F. equiseti
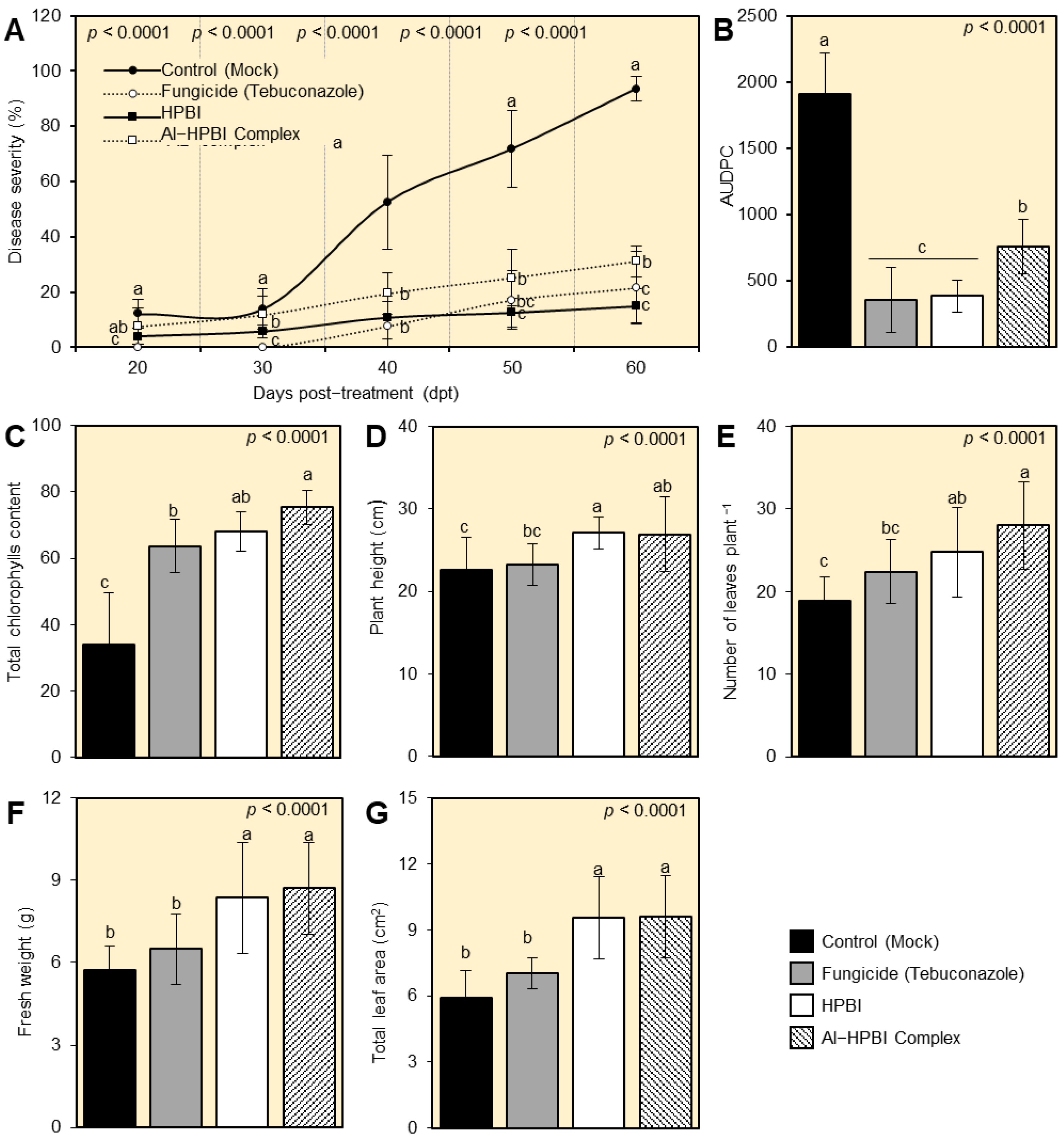
3.2. HPBI and Al−HPBI Complex Reduce the Development of Fusarium Wilt Disease
3.3. HPBI and Its Aluminum Complex Enhance the Growth Variables of Infected Pepper Plants
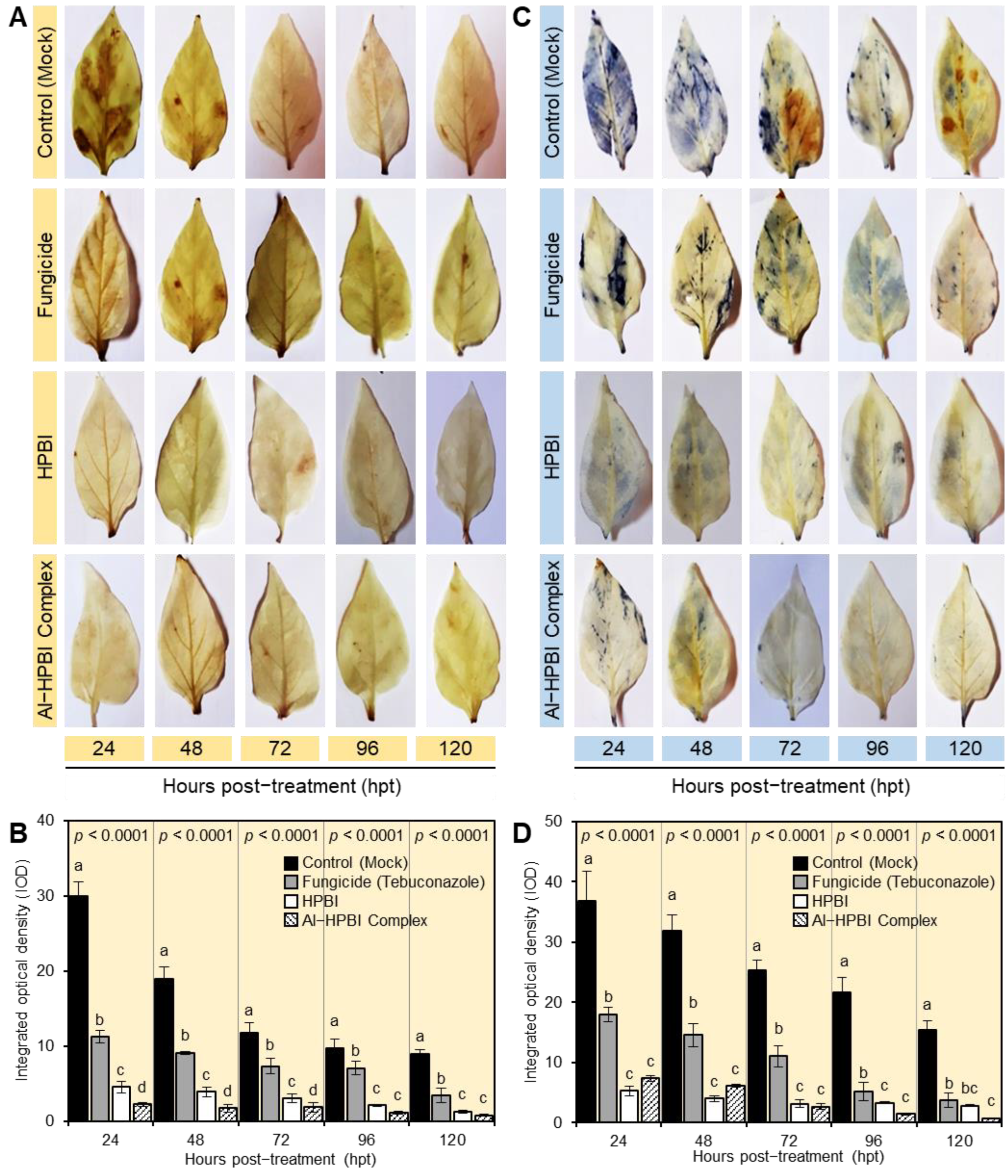
3.4. HPBI and Its Aluminum Complex Mitigate the Oxidative Stress of F. equiseti-Infected Plants
3.5. HPBI and Al−HPBI Complex Induce the Antioxidant Defense Machinery of F. equiseti-Infected Plants
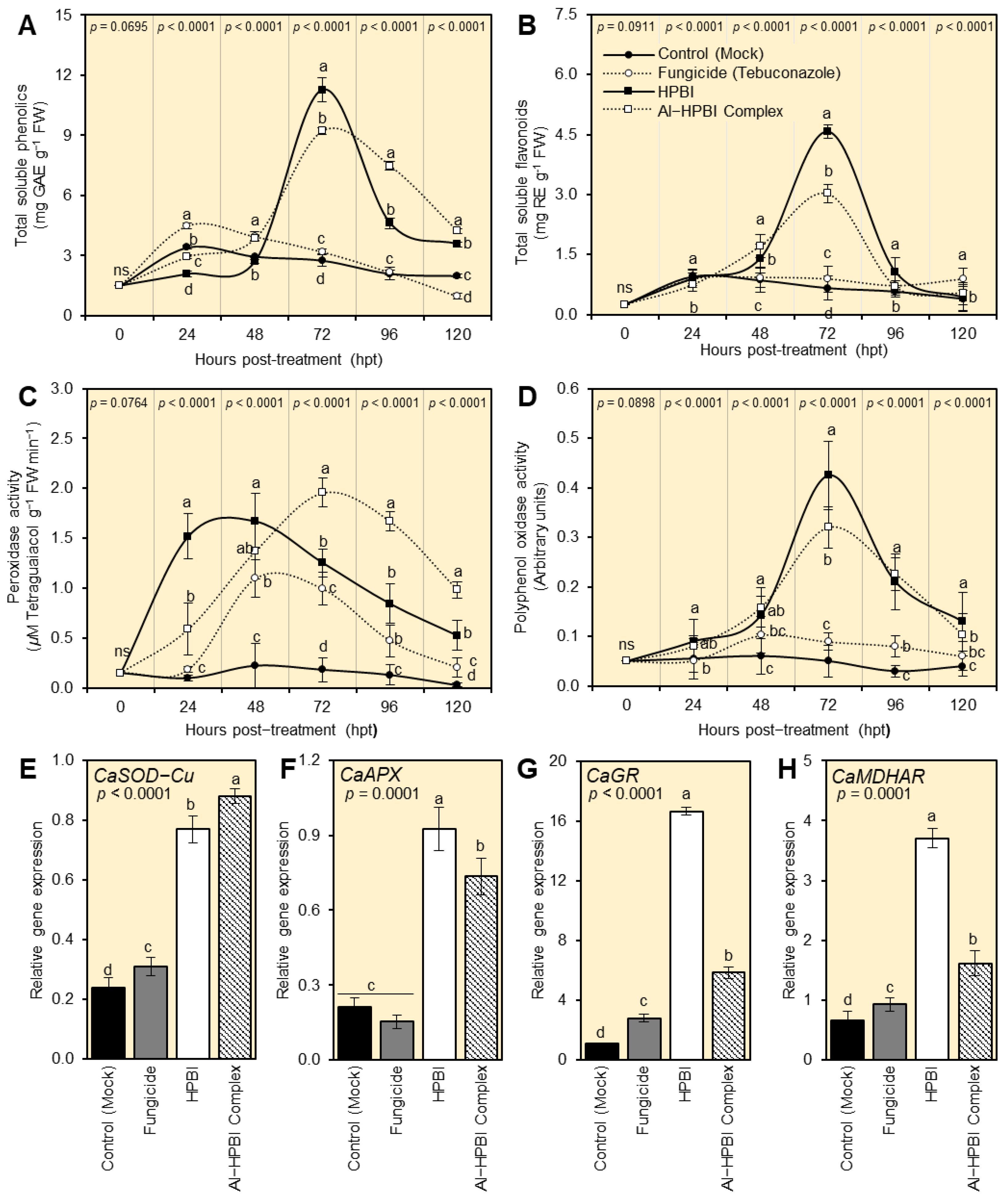
3.5.1. HPBI and Al−HPBI Complex Boost the Content of Total Soluble Phenolics and Flavonoids of F. equiseti-Infected Pepper Plants
3.5.2. HPBI and Al−HPBI Complex Induce Antioxidant-Related Enzymes of F. equiseti-Infected Pepper Plants
3.6. F. equiseti Genome Possesses a Putative Sterol 24-C-methyltransferase (EGR6)
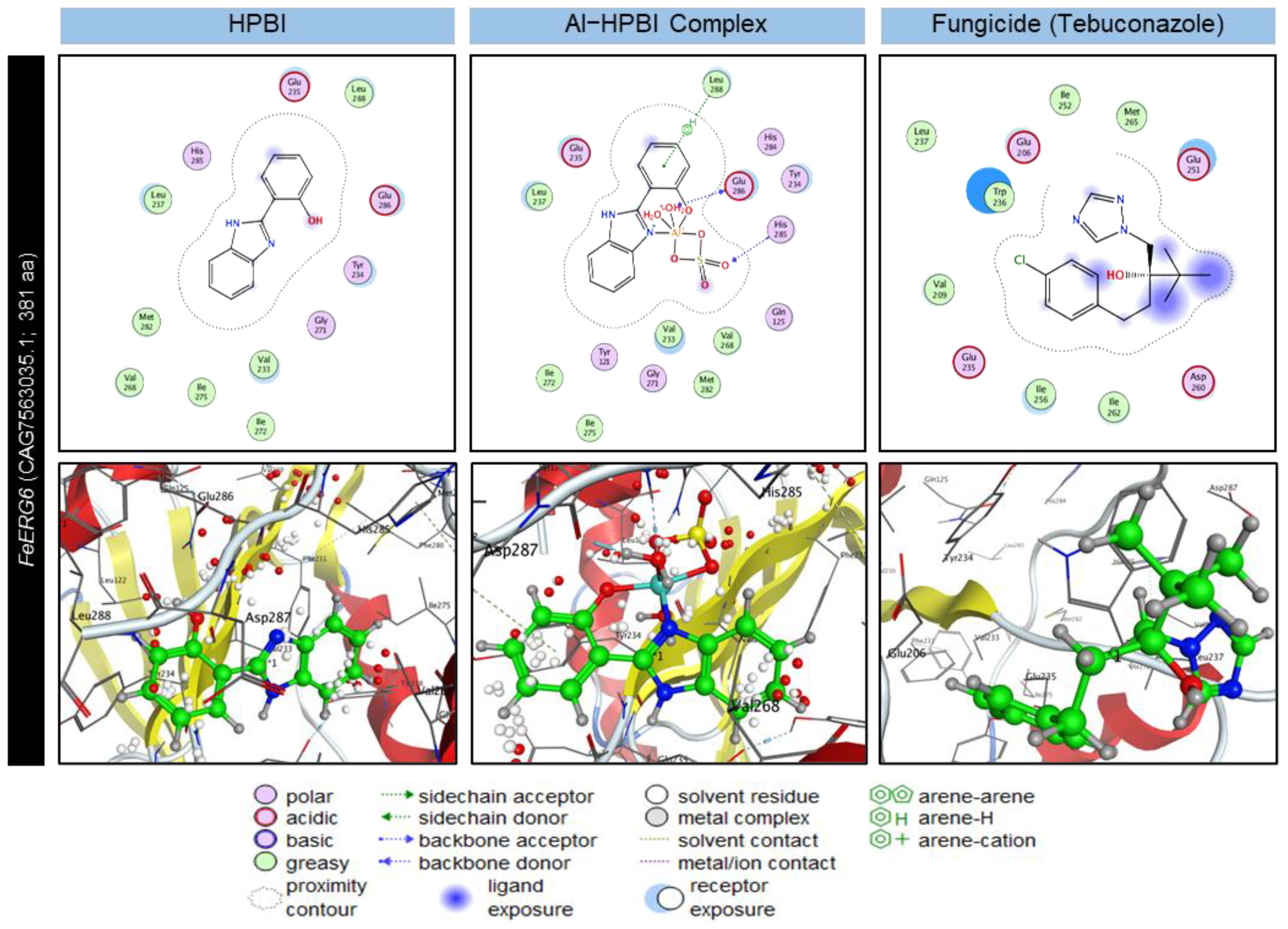
3.7. Molecular Docking Analysis Reveals Ligand–Protein Interactions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, L.-N.; Wang, D.-C.; Hu, Q.; Dai, X.-Q.; Xie, Y.-S.; Li, Q.; Liu, H.-M.; Guo, J.-H. Consortium of Plant Growth-Promoting Rhizobacteria Strains Suppresses Sweet Pepper Disease by Altering the Rhizosphere Microbiota. Front. Microbiol. 2019, 10, 1668. [Google Scholar] [CrossRef]
- Akram, W.; Mahboob, A.; Javed, A.A. Bacillus thuringiensis strain199 can induce systemic resistance in tomato againstFusariumwilt. Eur. J. Microbiol. Immunol. 2013, 3, 275–280. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Organization of the United Nations (FAO). Volume 25. Available online: http://www.fao.org/faostat/En/-Data/QC (accessed on 3 June 2020).
- Roberts, P.D.; Adkins, S.; Pernezny, K.; Jones, J.B. Diseases of Pepper and Their Management. In Diseases of Fruits and Vegetables: Volume II; Springer: Berlin/Heidelberg, Germany, 2004; pp. 333–387. [Google Scholar]
- El-Nagar, A.; Elzaawely, A.A.; Xuan, T.D.; Gaber, M.; El-Wakeil, N.; El-Sayed, Y.; Nehela, Y. Metal Complexation of Bis-Chalcone Derivatives Enhances Their Efficacy against Fusarium Wilt Disease, Caused by Fusarium equiseti, via Induction of Antioxidant Defense Machinery. Plants 2022, 11, 2418. [Google Scholar] [CrossRef] [PubMed]
- Poddar, R.K.; Singh, D.V.; Dubey, S.C. Management of Chickpea Wilt through Combination of Fungicides and Bioagents. Indian Phytopathol 2004, 57, 39–43. [Google Scholar]
- Cruz-Cerino, P.; Cristóbal-Alejo, J.; Ruiz-Carrera, V.; Carnevali, G.; Vera-Ku, M.; Martín, J.; Reyes, F.; Gamboa-Angulo, M. Extracts from Six Native Plants of the Yucatán Peninsula Hinder Mycelial Growth of Fusarium Equiseti and F. Oxysporum, Pathogens of Capsicum Chinense. Pathogens 2020, 9, 827. [Google Scholar] [CrossRef] [PubMed]
- Lomas-Cano, T.; Palmero-Llamas, D.; de Cara, M.; García-Rodríguez, C.; Boix-Ruiz, A.; Camacho-Ferre, F.; Tello-Marquina, J.C. First Report of Fusarium oxysporum on Sweet Pepper Seedlings in Almería, Spain. Plant Dis. 2014, 98, 1435. [Google Scholar] [CrossRef]
- Morid, B.; Hajmansoor, S.; Kakvan, N. Screening of Resistance Genes to Fusarium Root Rot and Fusarium Wilt Diseases in Tomato (Lycopersicon Esculentum) Cultivars Using RAPD and CAPs Markers. Eur. J. Exp. Biol 2012, 2, 931–939. [Google Scholar]
- Alegbejo, M.; Lawal, A.; Chindo, P.; Banwo, O. Outbreak of Basal Stem Rot and Wilt Disease of Pepper in Northern Nigeria. J. Plant Prot. Res. 2006, 46, 7–13. [Google Scholar]
- Metwally, N.H.; Radwan, I.T.; El-Serwy, W.S.; Mohamed, M.A. Design, synthesis, DNA assessment and molecular docking study of novel 2-(pyridin-2-ylimino)thiazolidin-4-one derivatives as potent antifungal agents. Bioorganic Chem. 2018, 84, 456–467. [Google Scholar] [CrossRef]
- Muto, M.; Takahashi, H.; Ishihara, K.; Yuasa, H.; Huang, J.-W. Antimicrobial Activity of Medicinal Plants Used by Indigenous People in Taiwan. J. Phytopathol. Soc. 2005, 14, 13–24. [Google Scholar]
- Abo-Elyousr, K.A.M.; Bagy, H.M.M.K.; Hashem, M.; Alamri, S.A.M.; Mostafa, Y.S. Biological control of the tomato wilt caused by Clavibacter michiganensis subsp. michiganensis using formulated plant growth-promoting bacteria. Egypt. J. Biol. Pest Control. 2019, 29, 54. [Google Scholar] [CrossRef]
- Khabnadideh, S.; Rezaei, Z.; Pakshir, K.; Zomorodian, K.; Ghafari, N. Synthesis and antifungal activity of benzimidazole, benzotriazole and aminothiazole derivatives. Res. Pharm. Sci. 2012, 7, 65–72. [Google Scholar]
- Reddy, V.M.; Reddy, K.R. Synthesis and Biological Evaluation of Some Novel-3-(5-Substituted Benzimidaz-ol-2-Yl)-5-Arylisoxazolines. Chin. Chem. Lett. 2010, 21, 1145–1148. [Google Scholar] [CrossRef]
- Hosamani, K.M.; Shingalapur, R.V. Synthesis of 2-Mercaptobenzimidazole Derivatives as Potential Anti-microbial and Cytotoxic Agents. Arch. der Pharm. 2011, 344, 311–319. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, J.; Zhu, Y.; Duan, Y.; Zhou, M. Mechanism of Action of the Benzimidazole Fungicide on Fusarium graminearum: Interfering with Polymerization of Monomeric Tubulin But Not Polymerized Microtubule. Phytopathology 2016, 106, 807–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tunçbilek, M.; Kiper, T.; Altanlar, N. Synthesis and in vitro antimicrobial activity of some novel substituted benzimidazole derivatives having potent activity against MRSA. Eur. J. Med. Chem. 2009, 44, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, G.G.; Attia, H.A.E.; Ibrahim, N.A. Synthesis, Characterization and Fungicidal Potentialities of Some Transition Metal Complexes of Benzimidazole Dithiocarbamate Based Ligand. J. Agric. Res. 2018, 96, 527–543. [Google Scholar]
- Raducka, A.; Świątkowski, M.; Korona-Głowniak, I.; Kapro, B.; Plech, T.; Szczesio, M.; Gobis, K.; Szynkowska-Jó´zwik, M.I.; Czylkowska, A. Zinc Coordination Compounds with Benzimidazole Derivatives: Synthesis, Structure, Antimicrobial Activity and Potential Anticancer Application. Int. J. Mol. Sci. 2022, 23, 6595. [Google Scholar] [CrossRef] [PubMed]
- Sahyon, H.A.; El-Bindary, A.A.; Shoair, A.F.; Abdellatif, A.A. Synthesis and Characterization of Ruthenium (III) Complex Containing 2-Aminomethyl Benzimidazole, and Its Anticancer Activity of in Vitro and in Vivo Models. J. Mol. Liq. 2018, 255, 122–134. [Google Scholar] [CrossRef]
- Apohan, E.; Yilmaz, U.; Yilmaz, O.; Serindag, A.; Küçükbay, H.; Yesilada, O.; Baran, Y. Synthesis, cytotoxic and antimicrobial activities of novel cobalt and zinc complexes of benzimidazole derivatives. J. Organomet. Chem. 2017, 828, 52–58. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, S.; Qian, P.; Li, Y.; Ren, W.; Deng, H.; Jiang, L. Synthesis and fungicidal activity of novel benzimidazole derivatives bearing pyrimidine-thioether moiety against Botrytis cinerea. Pest Manag. Sci. 2021, 77, 5529–5536. [Google Scholar] [CrossRef]
- Janeczko, M.; Kazimierczuk, Z.; Orzeszko, A.; Niewiadomy, A.; Król, E.; Szyszka, R.; Masłyk, M. In Search of the Antimicrobial Potential of Benzimidazole Derivatives. Pol. J. Microbiol. 2016, 65, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Casanova, I.; Durán, M.L.; Viqueira, J.; Sousa-Pedrares, A.; Zani, F.; Real, J.A.; García-Vázquez, J.A. Metal complexes of a novel heterocyclic benzimidazole ligand formed by rearrangement-cyclization of the corresponding Schiff base. Electrosynthesis, structural characterization and antimicrobial activity. Dalton Trans. 2018, 47, 4325–4340. [Google Scholar] [CrossRef]
- El-Safty, S.M.; El-Wakiel, N.; El-Oleimy, G.; Gaber, M.; El-Sayed, Y.S. Microhardness and Fluoride Release of Glass Ionomer Cement Modified with a Novel Al+3 Complex to Enhance Its Antimicrobial Activity. Int. J. Biomater. 2021, 2021, 1–12. [Google Scholar] [CrossRef]
- Grover, R.; Moore, J.D. Toximetric Studies of Fungicides against the Brown Root Organisms, Sclerotinia Fructicola and S. Laxa. Phytopathology 1962, 52, 876–880. [Google Scholar]
- Song, W.; Zhou, L.; Yang, C.; Cao, X.; Zhang, L.; Liu, X. Tomato Fusarium wilt and its chemical control strategies in a hydroponic system. Crop. Prot. 2004, 23, 243–247. [Google Scholar] [CrossRef]
- Jeger, M.J.; Viljanen-Rollinson, S.L.H. The use of the area under the disease-progress curve (AUDPC) to assess quantitative disease resistance in crop cultivars. Theor. Appl. Genet. 2001, 102, 32–40. [Google Scholar] [CrossRef]
- Romero-Puertas, M.C.; Rodriguez-Serrano, M.; Corpas, F.J.; del Gomez, M.; DEL Rio, L.A.; Sandalio, L.M. Cadmium-induced subcellular accumulation of O2.- and H2O2 in pea leaves. Plant Cell Environ. 2004, 27, 1122–1134. [Google Scholar] [CrossRef]
- Shi, J.; Fu, X.-Z.; Peng, T.; Huang, X.-S.; Fan, Q.-J.; Liu, J.-H. Spermine pretreatment confers dehydration tolerance of citrus in vitro plants via modulation of antioxidative capacity and stomatal response. Tree Physiol. 2010, 30, 914–922. [Google Scholar] [CrossRef]
- Killiny, N.; Nehela, Y.; Hijaz, F.; Gonzalez-Blanco, P.; Hajeri, S.; Gowda, S. Knock-down of $δ$-Aminolevulinic Acid Dehy-dratase via Virus-Induced Gene Silencing Alters the MicroRNA Biogenesis and Causes Stress-Related Reactions in Citrus Plants. Plant Sci. 2020, 299, 110622. [Google Scholar] [CrossRef]
- Nehela, Y.; Taha, N.A.; Elzaawely, A.A.; Xuan, T.D.; Amin, M.A.; Ahmed, M.E.; Nagar, A. El Benzoic Acid and Its Hydrox-ylated Derivatives Suppress Early Blight of Tomato (Alternaria Solani) via the Induction of Salicylic Acid Biosynthesis and Enzymatic and Nonenzymatic Antioxidant Defense Machinery. J. Fungi 2021, 7, 663. [Google Scholar] [CrossRef] [PubMed]
- Hückelhoven, R.; Fodor, J.; Preis, C.; Kogel, K.-H. Hypersensitive Cell Death and Papilla Formation in Barley Attacked by the Powdery Mildew Fungus Are Associated with Hydrogen Peroxide but Not with Salicylic Acid Accumulation. Plant Physiol. 1999, 119, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.-P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant Activity of Plant Extracts Containing Phenolic Compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant activity of some algerian medicinal plants extracts containing phenolic compounds. Food Chem. 2006, 97, 654–660. [Google Scholar] [CrossRef]
- Harrach, B.D.; Fodor, J.; Pogány, M.; Preuss, J.; Barna, B. Antioxidant, ethylene and membrane leakage responses to powdery mildew infection of near-isogenic barley lines with various types of resistance. Eur. J. Plant Pathol. 2007, 121, 21–33. [Google Scholar] [CrossRef]
- Malik, C.P.; Singh, M.B. Plant Enzymology and Histo-Enzymology; Kalyani Publishers: Delhi, India, 1980. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Killiny, N.; Nehela, Y. Metabolomic Response to Huanglongbing: Role of Carboxylic Compounds in Citrus sinensis Response to ‘Candidatus Liberibacter asiaticus’ and Its Vector, Diaphorina citri. Mol. Plant-Microbe Interact. 2017, 30, 666–678. [Google Scholar] [CrossRef] [Green Version]
- Nehela, Y.; Killiny, N. Melatonin Is Involved in Citrus Response to the Pathogen Huanglongbing via Modulation of Phytohormonal Biosynthesis. Plant Physiol. 2020, 184, 2216–2239. [Google Scholar] [CrossRef]
- Nehela, Y.; Killiny, N. ‘Candidatus Liberibacter asiaticus’ and Its Vector, Diaphorina citri, Augment the Tricarboxylic Acid Cycle of Their Host via the γ-Aminobutyric Acid Shunt and Polyamines Pathway. Mol. Plant-Microbe Interact. 2019, 32, 413–427. [Google Scholar] [CrossRef]
- Nehela, Y.; Killiny, N. Not Just a Cycle: Three gab Genes Enable the Non-Cyclic Flux Toward Succinate Via GABA Shunt in ‘Candidatus Liberibacter asiaticus’–Infected Citrus. Mol. Plant-Microbe Interact. 2022, 35, 200–214. [Google Scholar] [CrossRef]
- Altschul, S.F.; Wootton, J.C.; Gertz, E.M.; Agarwala, R.; Morgulis, A.; Schaffer, A.A.; Yu, Y.-K. Protein database searches using compositionally adjusted substitution matrices. FEBS J. 2005, 272, 5101–5109. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, J.S.; Agarwala, R. COBALT: Constraint-based alignment tool for multiple protein sequences. Bioinformatics 2007, 23, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Jeanmougin, F.; Thompson, J.D.; Gouy, M.; Higgins, D.G.; Gibson, T.J. Multiple sequence alignment with Clustal X. Trends Biochem. Sci. 1998, 23, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef]
- Bjellqvist, B.; Basse, B.; Olsen, E.; Celis, J.E. Reference points for comparisons of two-dimensional maps of proteins from different human cell types defined in a pH scale where isoelectric points correlate with polypeptide compositions. Electrophoresis 1994, 15, 529–539. [Google Scholar] [CrossRef]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera?A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Kiwaan, H.A.; El-Mowafy, A.S.; El-Bindary, A.A. Synthesis, spectral characterization, DNA binding, catalytic and in vitro cytotoxicity of some metal complexes. J. Mol. Liq. 2021, 326, 115381. [Google Scholar] [CrossRef]
- Sharfalddin, A.A.; Emwas, A.-H.; Jaremko, M.; Hussien, M.A. Synthesis and Theoretical Calculations of Metal-Antibiotic Chelation with Thiamphenicol: In Vitro DNA and HSA Binding, Molecular Docking, and Cytotoxicity Studies. New J. Chem. 2021, 45, 9598–9613. [Google Scholar] [CrossRef]
- Frisch, M.; Trucks, G.; Schlegel, H.; Scuseria, G.; Robb, M.; Cheeseman, J.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.; et al. Gaussian 09, Revision D.01; Gaussian Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Yanai, T.; Tew, D.P.; Handy, N.C. A new hybrid exchange–correlation functional using the Coulomb-attenuating method (CAM-B3LYP). Chem. Phys. Lett. 2004, 393, 51–57. [Google Scholar] [CrossRef]
- Prentice, R.L. A Generalization of the Probit and Logit Methods for Dose Response Curves. Biometrics 1976, 32, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Zarrinmayeh, H.; Nunes, A.M.; Ornstein, P.L.; Zimmerman, D.M.; Arnold, M.B.; Schober, D.A.; Gackenheimer, S.L.; Bruns, R.F.; Hipskind, P.A.; Britton, T.C.; et al. Synthesis and Evaluation of a Series of Novel 2-[(4-Chlorophenoxy) Me-thyl]-Benzimidazoles as Selective Neuropeptide Y Y1 Receptor Antagonists. J. Med. Chem. 1998, 41, 2709–2719. [Google Scholar] [CrossRef]
- Diener, A.C.; Li, H.; Zhou, W.-X.; Whoriskey, W.J.; Nes, W.D.; Fink, G.R. STEROL METHYLTRANSFERASE 1 Controls the Level of Cholesterol in Plants. Plant Cell 2000, 12, 853–870. [Google Scholar] [CrossRef]
- Bouvier-Navé, P.; Husselstein, T.; Benveniste, P. Two families of sterol methyltransferases are involved in the first and the second methylation steps of plant sterol biosynthesis. Eur. J. Biol. 1998, 256, 88–96. [Google Scholar] [CrossRef]
- Iwaki, T.; Iefuji, H.; Hiraga, Y.; Hosomi, A.; Morita, T.; Giga-Hama, Y.; Takegawa, K. Multiple functions of ergosterol in the fission yeast Schizosaccharomyces pombe. Microbiology 2008, 154, 830–841. [Google Scholar] [CrossRef]
- Pagadala, N.S.; Syed, K.; Tuszynski, J. Software for molecular docking: A review. Biophys. Rev. 2017, 9, 91–102. [Google Scholar] [CrossRef]
- Pantsar, T.; Poso, A. Binding Affinity via Docking: Fact and Fiction. Molecules 2018, 23, 1899. [Google Scholar] [CrossRef] [PubMed]
- Adeniyi, A.A.; Soliman, M.E.S. Implementing QM in docking calculations: Is it a waste of computational time? Drug Discov. Today 2017, 22, 1216–1223. [Google Scholar] [CrossRef]
- Gioia, D.; Bertazzo, M.; Recanatini, M.; Masetti, M.; Cavalli, A. Dynamic Docking: A Paradigm Shift in Computational Drug Discovery. Molecules 2017, 22, 2029. [Google Scholar] [CrossRef] [PubMed]
- Jyrkkärinne, J.; Küblbeck, J.; Pulkkinen, J.; Honkakoski, P.; Laatikainen, R.; Poso, A.; Laitinen, T. Molecular Dynamics Simulations for Human CAR Inverse Agonists. J. Chem. Inf. Model. 2012, 52, 457–464. [Google Scholar] [CrossRef]
- Bai, Y.-B.; Zhang, A.-L.; Tang, J.-J.; Gao, J.-M. Synthesis and Antifungal Activity of 2-Chloromethyl-1H-benzimidazole Derivatives against Phytopathogenic Fungi in Vitro. J. Agric. Food Chem. 2013, 61, 2789–2795. [Google Scholar] [CrossRef]
- Chandrika, N.T.; Shrestha, S.K.; Ngo, H.X.; Garneau-Tsodikova, S. Synthesis and Investigation of Novel Benzimidazole De-rivatives as Antifungal Agents. Bioorg. Med. Chem. 2016, 24, 3680–3686. [Google Scholar] [CrossRef]
- Karmaker, N.; Lira, D.N.; Das, B.K.; Kumar, U.; Shara, A.; Rouf, S. Synthesis and Antioxidant Activity of Some Novel Ben-zimidazole Derivatives. Dhaka Univ. J. Pharm. Sci. 2017, 16, 245–249. [Google Scholar] [CrossRef]
- Archie, S.R.; Das, B.K.; Hossain, S.; Kumar, U.; Rouf, A.S.S. Synthesis and antioxidant activity of 2-substituted-5-nitro benzimidazole derivatives. Int. J. Pharm. Pharm. Sci. 2016, 9, 308–310. [Google Scholar] [CrossRef]
- Dvornikova, I.A.; Buravlev, E.V.; Fedorova, I.V.; Shevchenko, O.G.; Chukicheva, I.Y.; Kutchin, A. V Synthesis and Antiox-idant Properties of Benzimidazole Derivatives with Isobornylphenol Fragments. Russ. Chem. Bull. 2019, 68, 1000–1005. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Bestwick, C.S.; Brown, I.R.; Mansfield, J.W. Localized Changes in Peroxidase Activity Accompany Hydrogen Peroxide Gen-eration during the Development of a Nonhost Hypersensitive Reaction in Lettuce. Plant Physiol. 1998, 118, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Eaton-rye, J.; Foyer, C.H.; Wu, Q.; Kuc, K.; Jac, V.; Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Racchi, M.L. Antioxidant Defenses in Plants with Attention to Prunus and Citrus spp. Antioxidants 2013, 2, 340–369. [Google Scholar] [CrossRef] [PubMed]
- Obianom, C.; Romanazzi, G.; Sivakumar, D. Effects of chitosan treatment on avocado postharvest diseases and expression of phenylalanine ammonia-lyase, chitinase and lipoxygenase genes. Postharvest Biol. Technol. 2018, 147, 214–221. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Docking Scores (S) (Kcal·mol−1) | RMSD-Refine (Å) | Strain Energy (Kcal·mol−1) | |||
|---|---|---|---|---|---|---|
| FoEGR6 | FeEGR6 | FoEGR6 | FeEGR6 | FoEGR6 | FeEGR6 | |
| HPBI | −5.38 | −5.36 | 2.50 | 1.94 | 39.06 | 39.49 |
| Al−HPBI Complex | −5.56 | −6.51 | 1.60 | 1.65 | 40.23 | 40.50 |
| Hattrick Fungicide | −6.64 | −6.05 | 1.32 | 3.15 | 39.56 | 39.60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Nagar, A.; Elzaawely, A.A.; El-Zahaby, H.M.; Xuan, T.D.; Khanh, T.D.; Gaber, M.; El-Wakeil, N.; El-Sayed, Y.; Nehela, Y. Benzimidazole Derivatives Suppress Fusarium Wilt Disease via Interaction with ERG6 of Fusarium equiseti and Activation of the Antioxidant Defense System of Pepper Plants. J. Fungi 2023, 9, 244. https://doi.org/10.3390/jof9020244
El-Nagar A, Elzaawely AA, El-Zahaby HM, Xuan TD, Khanh TD, Gaber M, El-Wakeil N, El-Sayed Y, Nehela Y. Benzimidazole Derivatives Suppress Fusarium Wilt Disease via Interaction with ERG6 of Fusarium equiseti and Activation of the Antioxidant Defense System of Pepper Plants. Journal of Fungi. 2023; 9(2):244. https://doi.org/10.3390/jof9020244
Chicago/Turabian StyleEl-Nagar, Asmaa, Abdelnaser A. Elzaawely, Hassan M. El-Zahaby, Tran Dang Xuan, Tran Dang Khanh, Mohamed Gaber, Nadia El-Wakeil, Yusif El-Sayed, and Yasser Nehela. 2023. "Benzimidazole Derivatives Suppress Fusarium Wilt Disease via Interaction with ERG6 of Fusarium equiseti and Activation of the Antioxidant Defense System of Pepper Plants" Journal of Fungi 9, no. 2: 244. https://doi.org/10.3390/jof9020244
APA StyleEl-Nagar, A., Elzaawely, A. A., El-Zahaby, H. M., Xuan, T. D., Khanh, T. D., Gaber, M., El-Wakeil, N., El-Sayed, Y., & Nehela, Y. (2023). Benzimidazole Derivatives Suppress Fusarium Wilt Disease via Interaction with ERG6 of Fusarium equiseti and Activation of the Antioxidant Defense System of Pepper Plants. Journal of Fungi, 9(2), 244. https://doi.org/10.3390/jof9020244