
A New Species of Neoscytalidium hylocereum sp. nov. Causing Canker on Red-Fleshed Dragon Fruit (Hylocereus polyrhizus) in Southern Thailand



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Fungal Isolation
2.2. Pathogenicity Test
2.3. Morphological Observation
2.4. Molecular Identification
3. Results
3.1. Disease Observation and Symptom Recognition
3.2. Pathogenicity Test
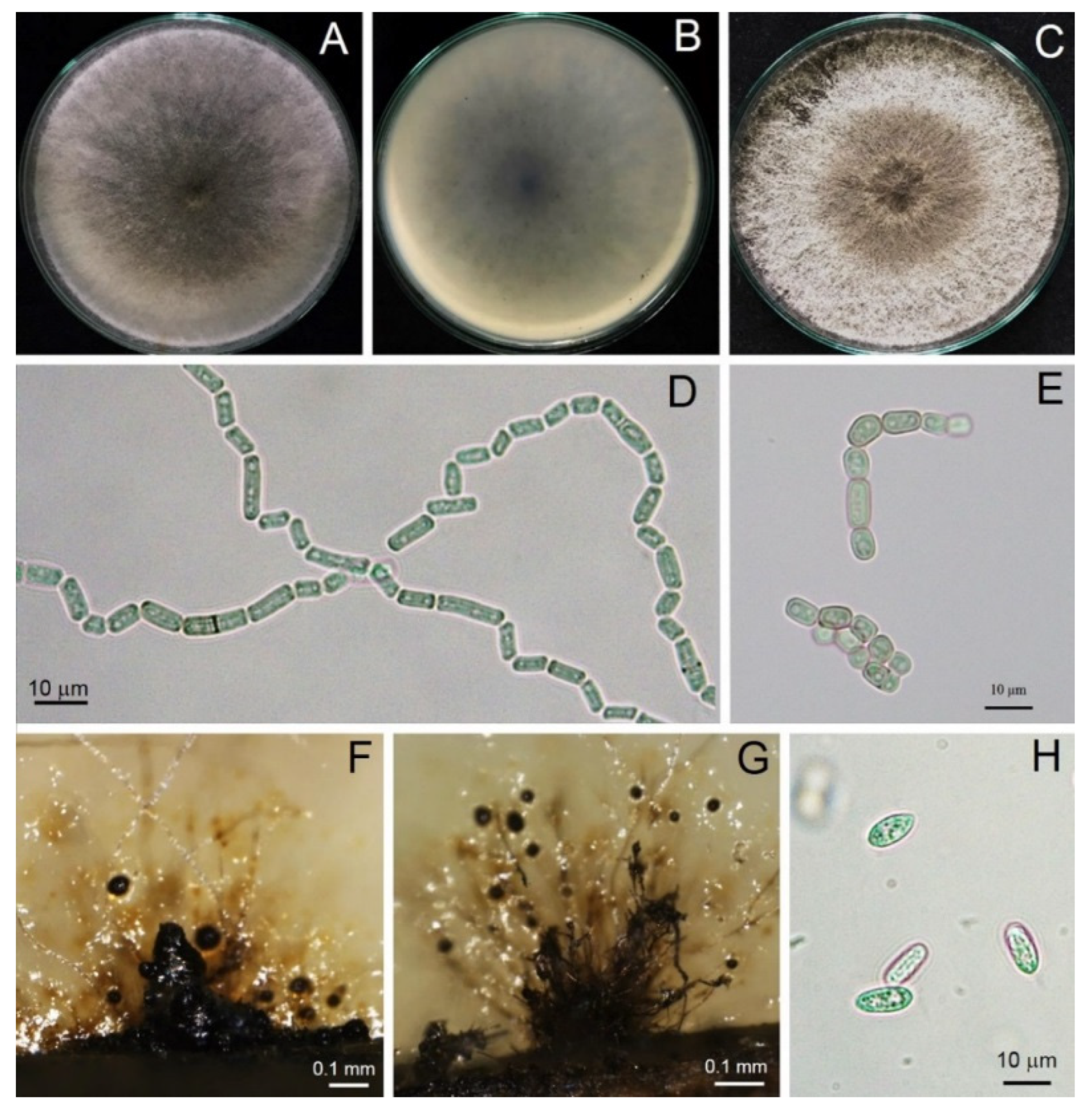
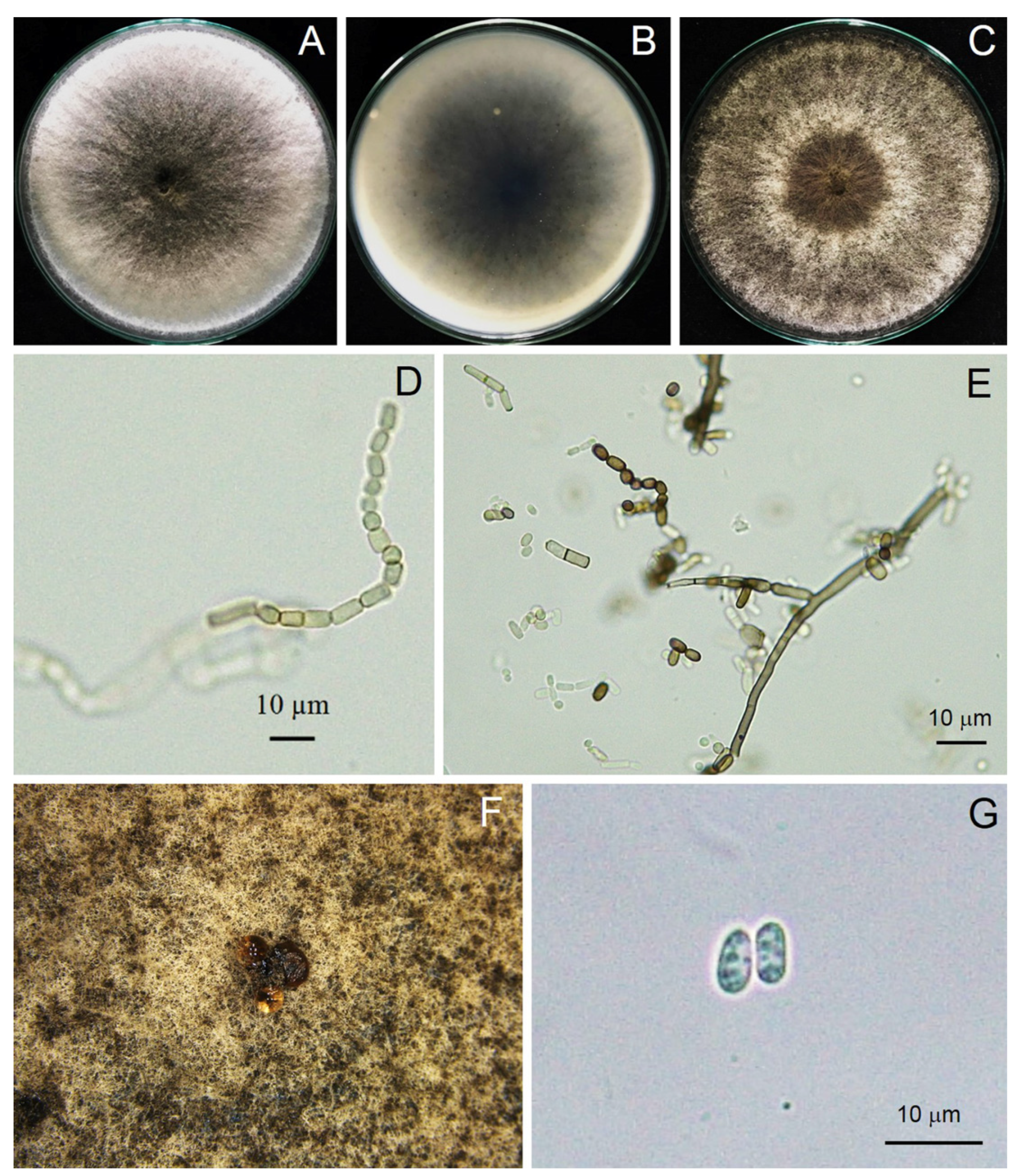
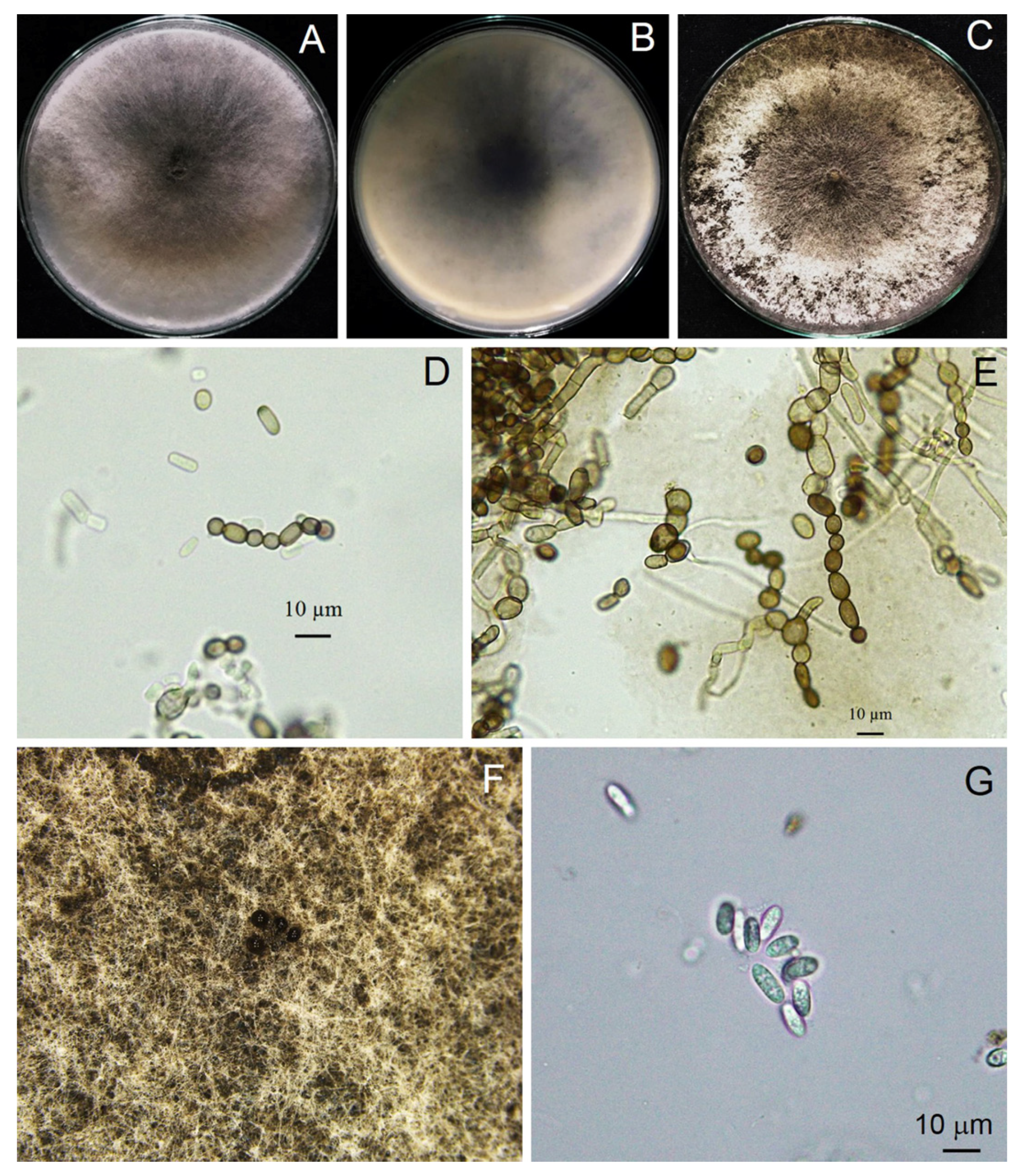
3.3. Morphological Study

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Colony | Arthroconidia | Pycnidia | Pycnidial Conidia | Reference |
|---|---|---|---|---|---|
| Neoscytalidium dimidiatum | White or hyaline colony, turned dark gray to black after 12 days on PDA | Cylindrical to round, hyaline to brown, singly or in arthric chains, 9.4 ± 1.2 × 5.1 ± 0.8 μm (length/width ratio = 1.85) | Produced after 3–4 weeks, black, irregularly shaped to ovoid, singly or in aggregates | Hyaline, ellipsoidal to ovoid, 11.1 ± 0.7 × 5.5 ± 0.5 μm (length/width ratio = 2.03) | [32] |
| N. novaehollandiae | – | Cylindrical, oblong to obtuse to doliiform, 0- to 1-septate, thick-walled, 5.2–11.2 × 2.9–4.9 μm, in arthric chains | Pycnidia (produced on pine needles on 1.5% water agar) semi-immersed or superficial with a mean diameter of 310 μm | Hyaline, ellipsoidal, becoming sepia, 0- to 1-septate, 3.1–4.1 × 12.2–13.8 μm | [33] |
| N. hyalinum | White with dense and hairy aerial mycelium and gradually turned gray to olive green | Truncated-cylindrical, oblong–obtuse to doliiform, hyaline to dark brown, 0- to 2-septate, in arthric chains, 4.57–9.85 × 3.125–27 μm | – | – | [34] |
| N. orchidacearum | Cream or white from above and reverse, with filamentous form or margin, flat | – | Stromatic, immersed, eventually erumpent, unilocular to multilocular (2~4-loculate), glabrous, brown to black, globose to subglobose, papillate, 200~500 μm diameter | Ellipsoidal to ovoid, hyaline, smooth, guttulate, aseptate becoming 2~3-septate, (10~) 12~13 (~15) × 3~5 (~6) μm (n = 50) | [31] |
| TSU-HP01 | White, moderate growth of 3 cm/day, olive green to gray color and turned dark gray to black after 12 days | Ellipsoid to ovoid, globose, oblong–cylindrical, singly or in arthric chains, 0- to 2-septate, 3.70–9.65 (6.24 ± 1.77) × 2.46–6.15 (3.73 ± 1.04) μm, length/width ratio = 1.67 | Rarely produced on PDA after 4 weeks of incubation, brown to dark brown, singly or in aggregates | Ellipsoid, oblong or cylindrical, 9.81–13.52 (11.48 ± 0.93) × 3.73–5.04 (4.27 ± 0.36) μm, length/width ratio = 2.68 | This study |
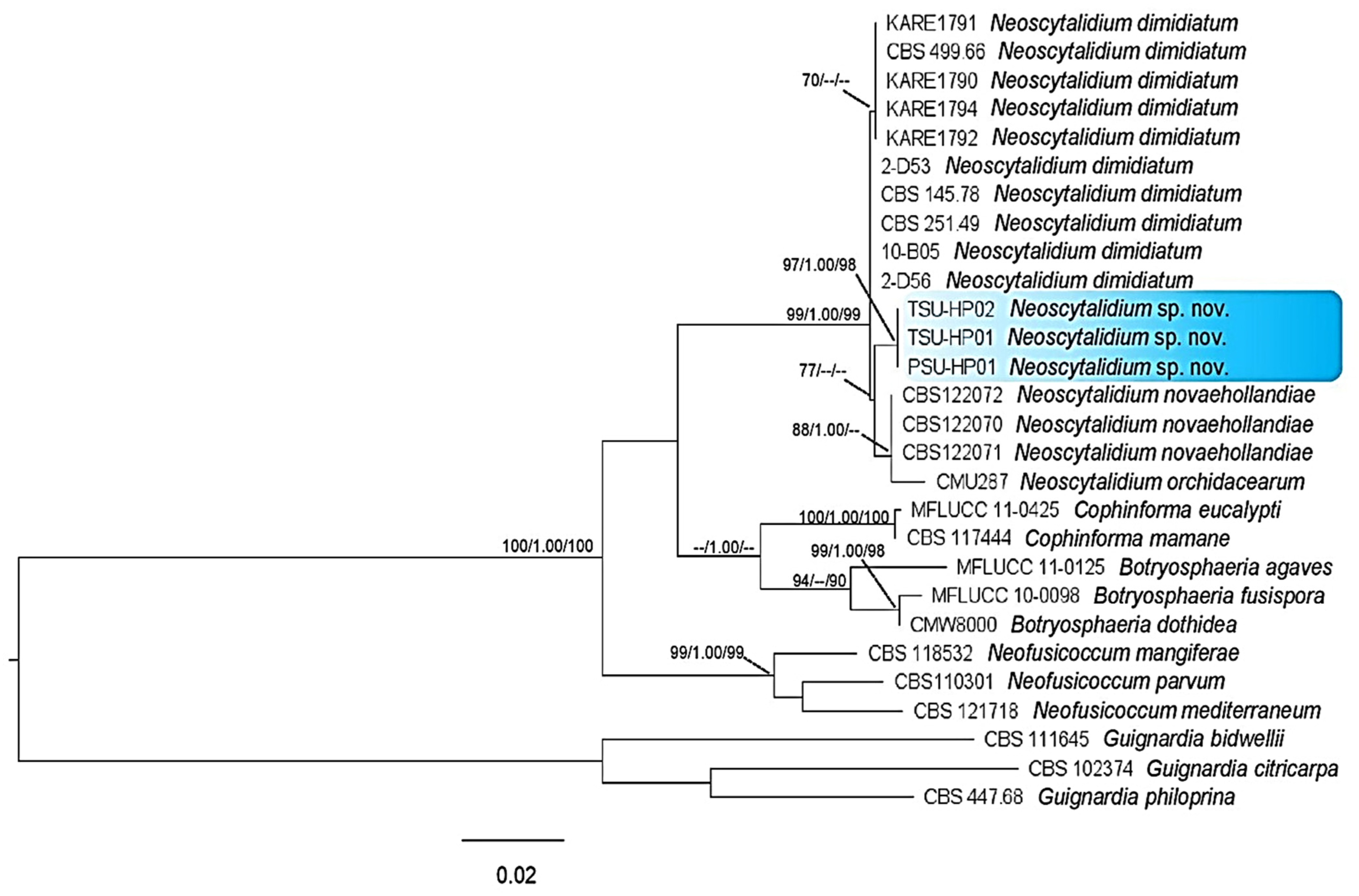
3.4. Phylogenetic Tree Analysis
3.5. Taxonomy of a New Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jaafar, R.A.; Abdul Rahman, A.R.B.; Mahnood, N.Z.C.; Vasudevan, R. Proximate analysis of dragon fruit (Hylocereus polyrhizus). Am. J. Applied Sci. 2009, 6, 1341–1346. [Google Scholar]
- Sonawane, M.S. Nutritive and medicinal value of dragon fruit. Asian J. Hort. 2017, 12, 267–271. [Google Scholar] [CrossRef]
- Mizrahi, Y.; Nerd, A.; Nobel, P.S. Cacti as crops. Hort. Rev. 1997, 18, 291–320. [Google Scholar]
- Masyahit, M.; Sijam, K.; Awang, Y.; Satar, M.G.M. The first report of the occurrence of anthracnose disease caused by Colletotrichum gloeospo rioides (Penz.) Penz. & Sacc. on dragon fruit (Hylocereus spp.) in Peninsular Malaysia. Am. J. Applied Sci. 2009, 6, 902–912. [Google Scholar]
- Barbeau, G.C. La pitahaya rouge, un nouveau fruit exotique (The red pitahaya, a new exotic fruit). Fruits 1990, 45, 141–147. [Google Scholar]
- Hawa, M.M.; Salleh, B.; Latiffah, Z. Characterization and intraspecific variation of Fusarium semitectum (Berkeley and Ravenel) associated with red-fleshed dragon fruit (Hylocereus polyrhizus [Weber] Britton and Rose) in Malaysia. Afr. J. Biotechnol. 2010, 9, 273–284. [Google Scholar]
- Ghani, M.A.A.; Awang, Y.; Sijam, K. Disease occurrence and fruit quality of pre-harvest calcium treated red flesh dragon fruit (Hylocereus polyrhizus). Afr. J. Biotechnol. 2013, 10, 1550–1558. [Google Scholar]
- Hawa, M.M.; Salleh, B.; Latiffah, Z. Characterization and pathogenicity of Fusarium proliferatum causing stem rot of Hylocereus polyrhizus in Malaysia. Ann. Appl. Biol. 2013, 163, 269–280. [Google Scholar] [CrossRef]
- Guo, L.W.; Wu, Y.X.; Ho, H.H.; Su, Y.Y.; Mao, Z.C.; He, P.F.; He, Y.Q. First report of dragon fruit (Hylocereus undatus) anthracnose caused by Colletotrichum truncatum in China. J. Phytopath. 2014, 162, 272–275. [Google Scholar] [CrossRef]
- Mohd, M.H.; Salleh, B.; Zakaria, L. Identification and molecular characterizations of Neoscytalidium dimidiatum causing stem canker of red-fleshed dragon fruit (Hylocereus polyrhizus) in Malaysia. J. Phytopath. 2013, 161, 841–849. [Google Scholar] [CrossRef]
- Wonglom, P.; Thithuan, N.; Bunjongsiri, P.; Sunpapao, A. Plant-Parasitic algae (Cephaleuros spp.) in Thailand, including four new records. Pacific Sci. 2018, 72, 363–371. [Google Scholar] [CrossRef]
- Thithuan, N.; Bunjonsiri, P.; Sunpapao, A. Morphology and behavior of gametes and zoospores from the plant-parasitic green algae, Cephaleuros (Chlorophyta, Ulvophyceae). Pac. Sci. 2019, 73, 403–410. [Google Scholar] [CrossRef]
- Ruangwong, O.-U.; Kunasakdakul, K.; Wonglom, P.; Dy, K.S.; Sunpapao, A. Morphological and molecular studies of a rare mucoralean species causing flower rot in Hylocereus polyrhizus . J. Phytopathol. 2022, 170, 217–220. [Google Scholar] [CrossRef]
- Dy, K.S.; Wonglom, P.; Pornsuriya, C.; Sunpapao, A. Morphological, molecular identification and pathogenicity of Neoscytalidium dimidiatum causing stem canker of Hylocereus polyrhizus in southern Thailand. Plants 2022, 11, 504. [Google Scholar] [CrossRef]
- Daengsuwan, W.; Wonglom, P.; Arikit, S.; Sunpapao, A. Morphological and molecular identification of Neopestalotiopsis clavispora causing flower blight on Anthurium andraeanum in Thailand. Hort. Plant J. 2021, 7, 573–578. [Google Scholar] [CrossRef]
- Sunpapao, A.; Suwannarach, N.; Kumla, J.; Dumhai, R.; Riangwong, K.; Sanguansub, S.; Wanchana, S.; Arikit, S. Morphological and molecular identification of plant pathogenic fungi associated with dirty panicle disease in coconuts (Cocos nucifera) in Thailand. J. Fungi 2022, 8, 335. [Google Scholar] [CrossRef]
- Thaochan, N.; Pornsuriya, C.; Chairin, T.; Chomnunti, P.; Sunpapao, A. Morphological and molecular characterization of Calonectria foliicola associated with leaf blight on rubber tree (Hevea brasiliensis) in Thailand. J. Fungi 2022, 8, 986. [Google Scholar] [CrossRef]
- Pornsuriya, C.; Chairin, T.; Thaochan, N.; Sunpapao, A. Identification and characterization of Neopestalotiopsis fungi associated with a novel leaf fall disease of rubber trees (Hevea brasiliensis) in Thailand. J. Phytopathol. 2020, 168, 416–427. [Google Scholar] [CrossRef]
- Saqib, S.; Zaman, W.; Ayaz, A.; Habib, S.; Bahadur, S.; Hussain, S.; Muhammad, S.; Ullah, F. Postharvest disease inhibition in fruit by synthesis and characterization of chitosan iron oxide nanoparticles. Biocatal. Agric. Biotechnol. 2020, 28, 101729. [Google Scholar] [CrossRef]
- Saqib, S.; Zaman, W.; Ullah, F.; Majeed, I.; Ayaz, A.; Munis, M.F.H. Organometallic assembling of chitosan-Iron oxide nanoparticles with their antifungal evaluation against Rhizopus oryzae . Appl. Organomet. Chem. 2019, 33, e5190. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetic. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Saitoh, K.; Togashi, K.; Arie, T.; Teraoka, T. A simple method for a mini-preparation of fungal DNA. J. Gen. Plant Pathol. 2006, 72, 348–350. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nuclic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamaru, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MRBAYES 3.2: Efficient Bayesian phylogenetic inference and model selection across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.O.; Philips, A.J.L.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef]
- Nouri, M.T.; Lawrence, D.P.; Yaghmour, M.A.; Michailides, T.J.; Trouillas, F.P. Neoscytalidium dimidiatum causing canker, shoot blight and fruit rot of almond in California. Plant Dis. 2018, 102, 1638–1647. [Google Scholar] [CrossRef] [PubMed]
- Derviş, S.; Özer, G.; Türkölmez, S. First report of Neoscytalidium novaehollandiae causing stem blight on tomato in Turkey. J. Plant Pathol. 2020, 102, 1339–1340. [Google Scholar] [CrossRef]
- Mirtalebi, M.; Sabahi, F.; Banihashemi, Z. Fruit rot caused by Neoscytalidium hyalinum on melon in Iran. Australasian Plant Dis. Notes 2019, 14, 8. [Google Scholar] [CrossRef]
- Wonglom, P.; Sunpapao, A. Fusarium incarnatum is associated with postharvest fruit rot of muskmelon (Cucumis melo). J. Phytopathol. 2020, 168, 204–210. [Google Scholar] [CrossRef]
- Daengsuwan, W.; Wonglom, P.; Sunpapao, A. First report of Lasiodiplodia theobromae causing spadix rot in Anthurium andraeanum . J. Phytopathol. 2020, 168, 129–133. [Google Scholar] [CrossRef]
- Slippers, B.; Crous, P.W.; Denman, S.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Combined multiple gene genealogies and phenotypic characters differentiate several species previously identified as Botryosphaeria dothidea . Mycologia 2004, 96, 83–101. [Google Scholar] [CrossRef]
- Rolshausen, P.E.; Akgül, D.S.; Perez, R.; Eskalen, A.; Gispert, C. First report of wood canker caused by Neoscytalidium dimidiatum on grapevine in California. Plant Dis. 2013, 97, 1511. [Google Scholar] [CrossRef]
- Fernández-Herrera, E.; Moreno-Salazar, S.F.; Rentería-Martínez, M.E.; Arratia-Castro, A.A.; Villar-Luna, E. Neoscytalidium dimidiatum: Causal agent of dieback in Ficus benjamina L. in Mexico. Rev. Chapingo Ser. Horticu. 2017, 23, 203–210. [Google Scholar] [CrossRef]
- Türkölmez, S.; Derviş, S.; Çiftçi, O.; Serçe, C.U.; Türkölmez, C.G.; Dikilitas, M. First report of Neoscytalidium dimidiatum causing dieback, shoot blight, and branch canker of willow trees in Turkey. Plant Dis. 2019, 103, 2139. [Google Scholar] [CrossRef]
- Oksal, E.; Yiğit, T.; Özer, G. First report of Neoscytalidium dimidiatum causing shoot blight, dieback and canker of apricot in Turkey. J. Plant Pathol. 2020, 102, 579–580. [Google Scholar] [CrossRef]
- Chuang, M.F.; Ni, H.F.; Yang, H.R.; Shu, S.L.; Lai, S.Y.; Jiang, Y.L. First report of stem canker disease of pitaya (Hylocereus undatus and H. polyrhizus) caused by Neoscytalidium dimidiatum in Taiwan. Plant Dis. 2012, 96, 906. [Google Scholar] [CrossRef] [PubMed]
- Sanahuja, G.; Lopez, P.; Palmateer, A.J. First report of Neoscytalidium dimidiatum causing stem and fruit canker of Hylocereus undatus in Florida. Plant Dis. 2016, 100, 1499. [Google Scholar] [CrossRef]
- Xu, M.; Peng, Y.; Qi, Z.; Yan, Z.; Yang, L.; He, M.-D.; Li, Q.-X.; Liu, C.-L.; Ruan, Y.-Z.; Wei, S.-S.; et al. Identification of Neoscytalidium dimidiatum causing canker disease of pitaya in Hainan, China. Australas. Plant Pathol. 2018, 47, 547–553. [Google Scholar] [CrossRef]






| Species | Voucher/Culture | Location | Host | GenBank Accession Number | ||
|---|---|---|---|---|---|---|
| ITS | tef1-α | tub | ||||
| Botryosphaeria agaves | MFLUCC 11-0125 T | Thailand | Agaves sp. | JX646791 | JX646856 | JX646841 |
| B. fusispora | MFLUCC 10-0098 T | Thailand | Caryota sp. | JX646789 | JX646854 | JX646839 |
| B. dothidea | CMW8000 | Switzerland | Prunus sp. | AY236949 | AY236898 | AY236927 |
| Cophinforma eucalypti | MFLUCC 11-0425 | Thailand | Eucalyptus sp. | JX646800 | JX646865 | JX646848 |
| C. mamane | CBS 117444 | Venezuela | Eucalyptus sp. | KF531822 | KF531801 | KF531802 |
| Guignardia bidwellii | CBS 111645 | USA: Missouri | Parthenocissus quinquefolia | FJ824766 | FJ824772 | FJ824777 |
| G. citricarpa | CBS 102374 T | Brazil | Citrus aurantium | FJ824767 | FJ538371 | FJ824778 |
| G. philoprina | CBS 447.68 | USA | Taxus baccata | FJ824768 | FJ824773 | FJ824779 |
| Neofusicoccum mangiferae | CBS 118532 | Australia | M. indica | AY615186 | DQ093220 | AY615173 |
| N. mediterraneum | CBS 121718 T | Greece | Eucalyptus sp. | GU251176 | GU251308 | GU251836 |
| N. parvum | CBS 110301 | Portugal | Vitis vinifera | AY259098 | AY573221 | EU673095 |
| Neoscytalidium dimidiatum | 10-B05 | USA: Kern | P. dulcis | MG021589 | MG021541 | MG021486 |
| KARE1790 | USA: Madera | P. dulcis | MG021578 | MG021567 | MF991145 | |
| KARE1791 | USA: Madera | P. dulcis | MG021579 | MG021531 | MG021476 | |
| KARE1792 | USA: Madera | P. dulcis | MG021580 | MG021532 | MG021477 | |
| KARE1794 | USA: Madera | P. dulcis | MG021582 | MG021534 | MG021479 | |
| 2-D53 | USA: Madera | Ficus carica | MG021568 | MG021521 | MG021511 | |
| 2-D56 | USA: Fresno | F. carica | MG021570 | MG021523 | MG021513 | |
| CBS 145.78 I,* | United Kingdom | Homo sapiens | KF531816 | KF531795 | KF531796 | |
| CBS 499.66 * | Mali | Mangifera indica | KF531820 | KF531798 | KF531800 | |
| CBS 251.49 * | USA | Juglans regia | KF531819 | KF531797 | KF531799 | |
| N. novaehollandiae | CBS122071 T | Australia | Crotalaria medcaginea | EF585540 | EF585580 | MT592760 |
| CBS122070 | Australia | Grevillea agrifolia | EF585539 | EF585579 | MT592759 | |
| CBS122072 | Australia | Adansonia gibbosa | EF585535 | EF585581 | MT592761 | |
| N. orchidacearum | CMU287 T | Thailand | Cattleya sp. | KY933091 | N/A | N/A |
| Neoscytalidium sp. | PSU-HP01 | Thailand | Hylocereus polyrhizus | LC590859 | LC590862 | LC647832 |
| Neoscytalidium sp. | TSU-HP01 | Thailand | H. polyrhizus | LC590860 | LC590863 | LC647833 |
| Neoscytalidium sp. | TSU-HP02 | Thailand | H. polyrhizus | LC590861 | LC590864 | LC647834 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wonglom, P.; Pornsuriya, C.; Sunpapao, A. A New Species of Neoscytalidium hylocereum sp. nov. Causing Canker on Red-Fleshed Dragon Fruit (Hylocereus polyrhizus) in Southern Thailand. J. Fungi 2023, 9, 197. https://doi.org/10.3390/jof9020197
Wonglom P, Pornsuriya C, Sunpapao A. A New Species of Neoscytalidium hylocereum sp. nov. Causing Canker on Red-Fleshed Dragon Fruit (Hylocereus polyrhizus) in Southern Thailand. Journal of Fungi. 2023; 9(2):197. https://doi.org/10.3390/jof9020197
Chicago/Turabian StyleWonglom, Prisana, Chaninun Pornsuriya, and Anurag Sunpapao. 2023. "A New Species of Neoscytalidium hylocereum sp. nov. Causing Canker on Red-Fleshed Dragon Fruit (Hylocereus polyrhizus) in Southern Thailand" Journal of Fungi 9, no. 2: 197. https://doi.org/10.3390/jof9020197
APA StyleWonglom, P., Pornsuriya, C., & Sunpapao, A. (2023). A New Species of Neoscytalidium hylocereum sp. nov. Causing Canker on Red-Fleshed Dragon Fruit (Hylocereus polyrhizus) in Southern Thailand. Journal of Fungi, 9(2), 197. https://doi.org/10.3390/jof9020197