
Global Transcriptomic Changes Elicited by sodB Deletion and Menadione Exposure in Aspergillus nidulans



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Culturing Conditions
2.2. Measuring Superoxide Formation and Superoxide Dismutase (SOD) Activities
2.3. Reverse-Transcriptional Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR) Assays
2.4. High-Throughput RNA Sequencing
- (1)
- Untreated cultures of A. nidulans THS30.3 reference strain;
- (2)
- MSB (0.16 mM) stress-treated cultures of A. nidulans THS30.3 reference strain;
- (3)
- Untreated cultures of A. nidulans ΔsodB mutant;
- (4)
- MSB (0.16 mM) stress-treated cultures of A. nidulans ΔsodB mutant
2.5. Evaluation of Transcriptome Data
3. Results
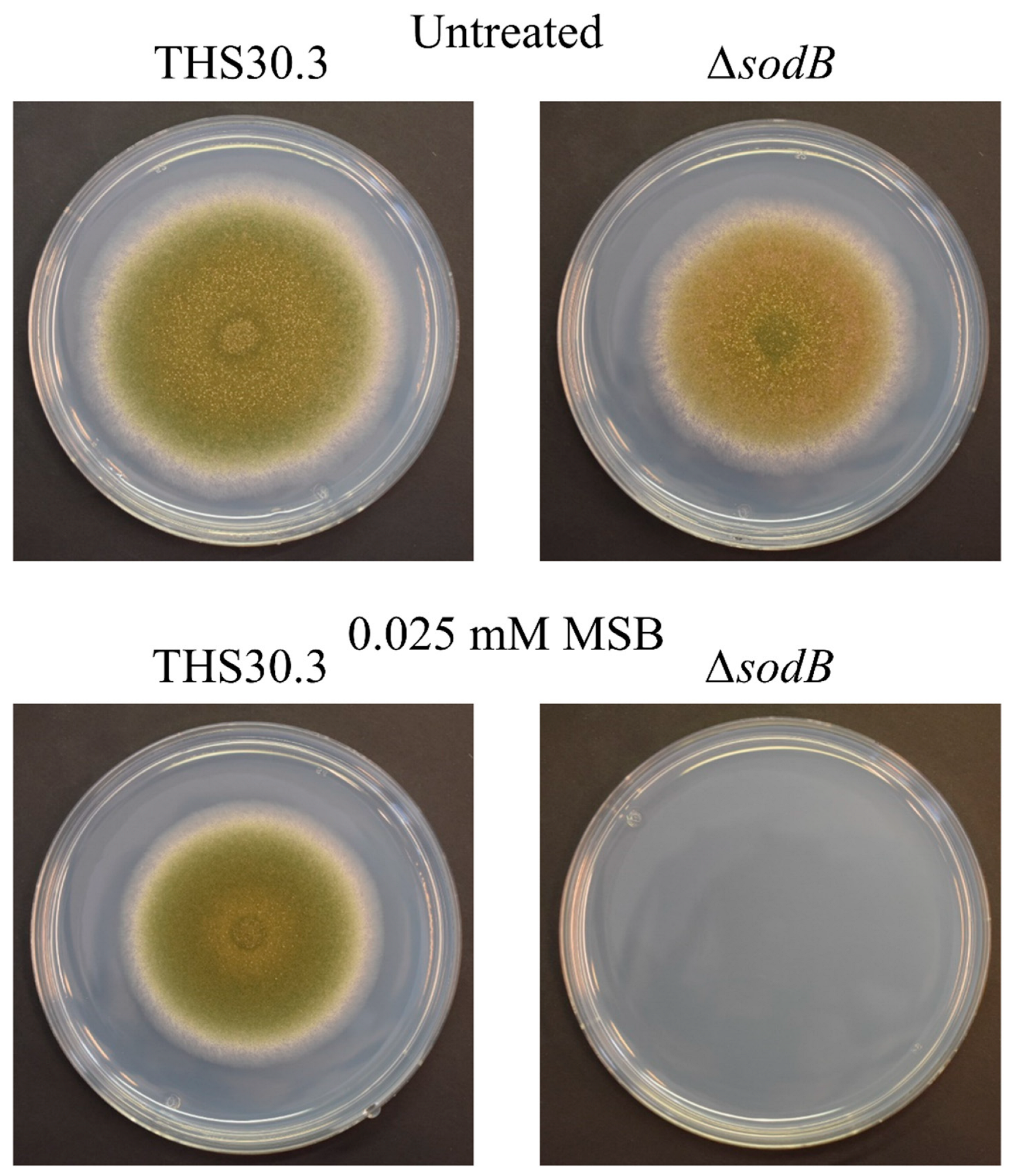
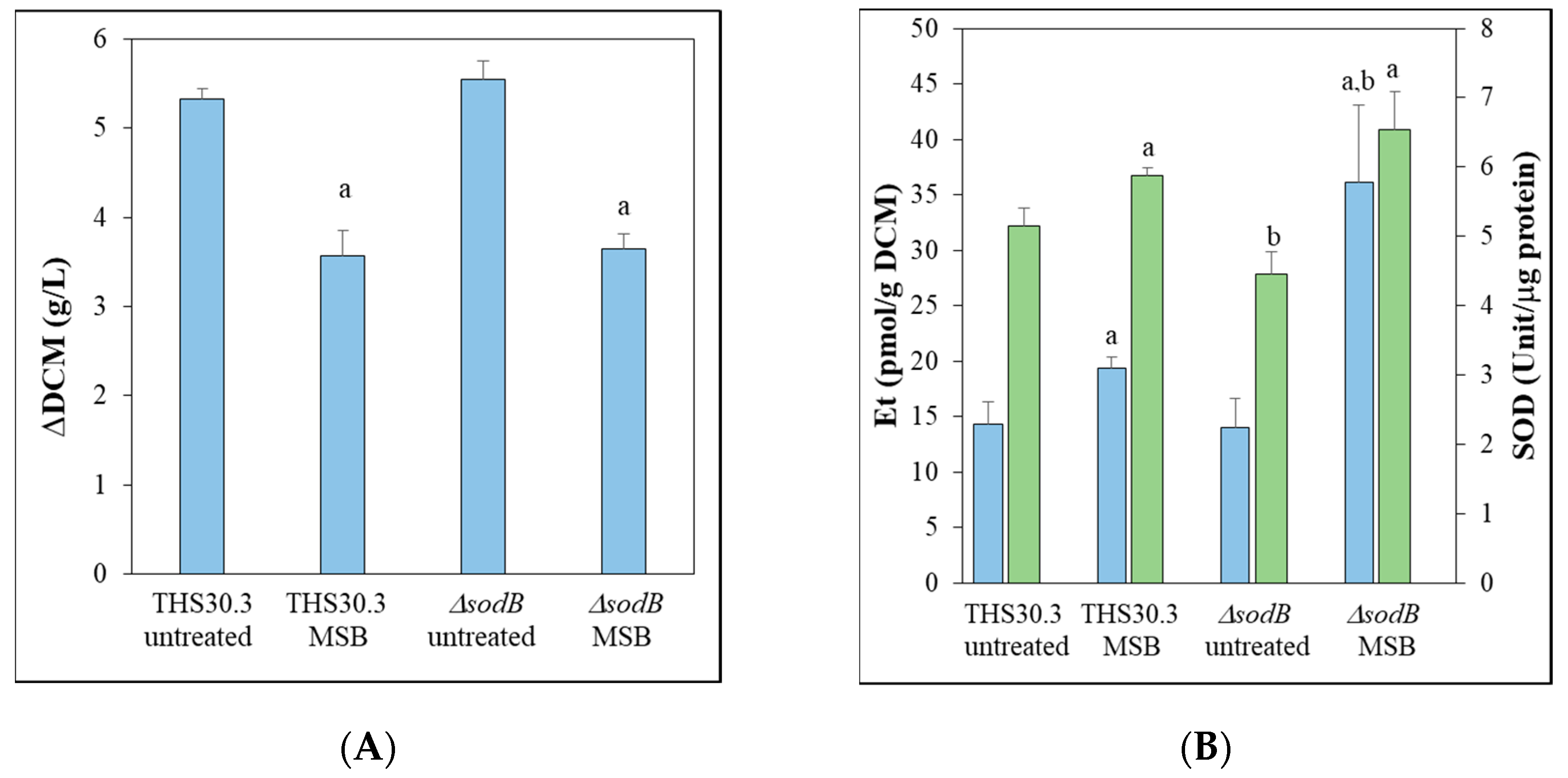
3.1. Intracellular Superoxide Formation and SOD Activity
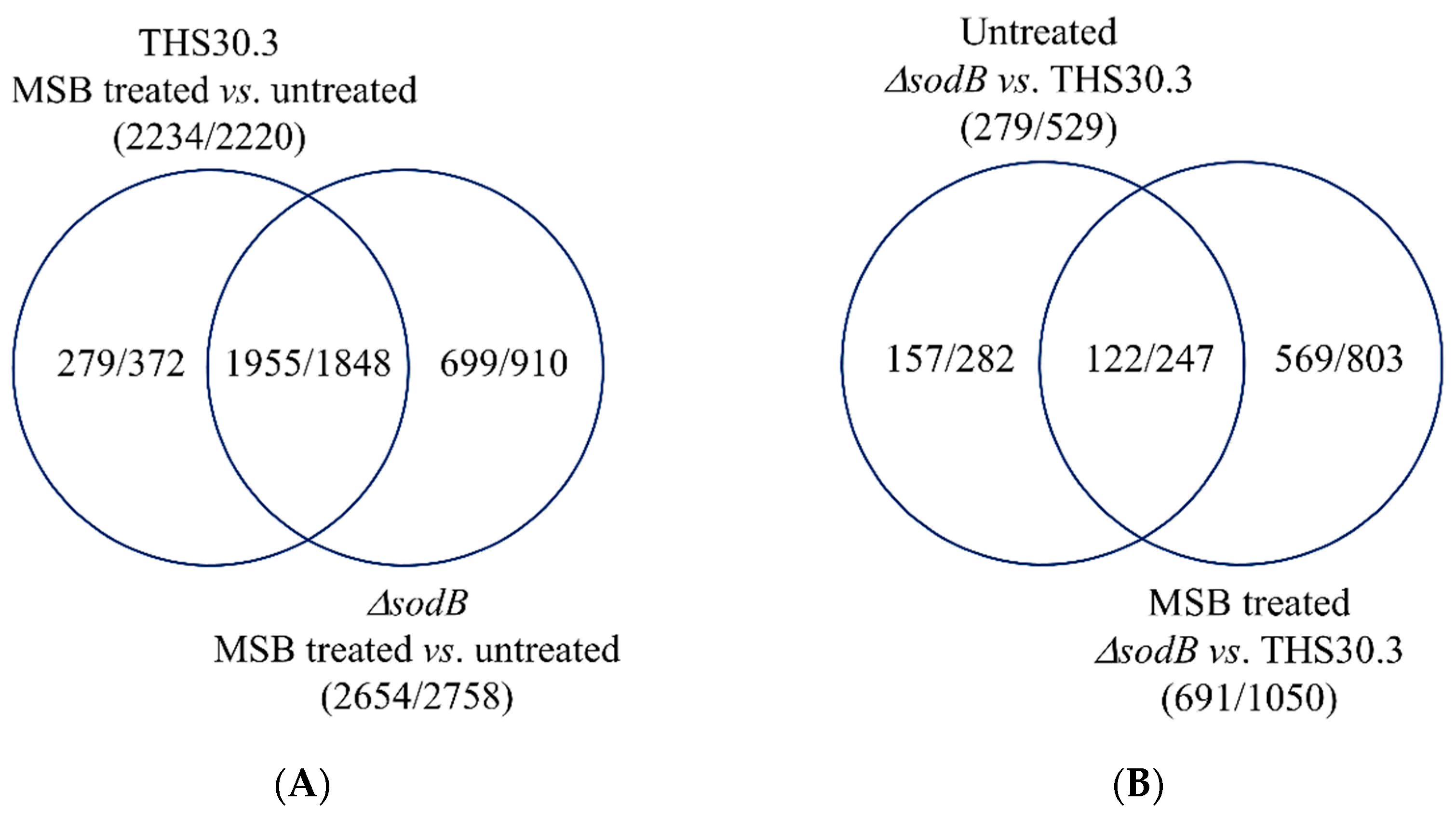
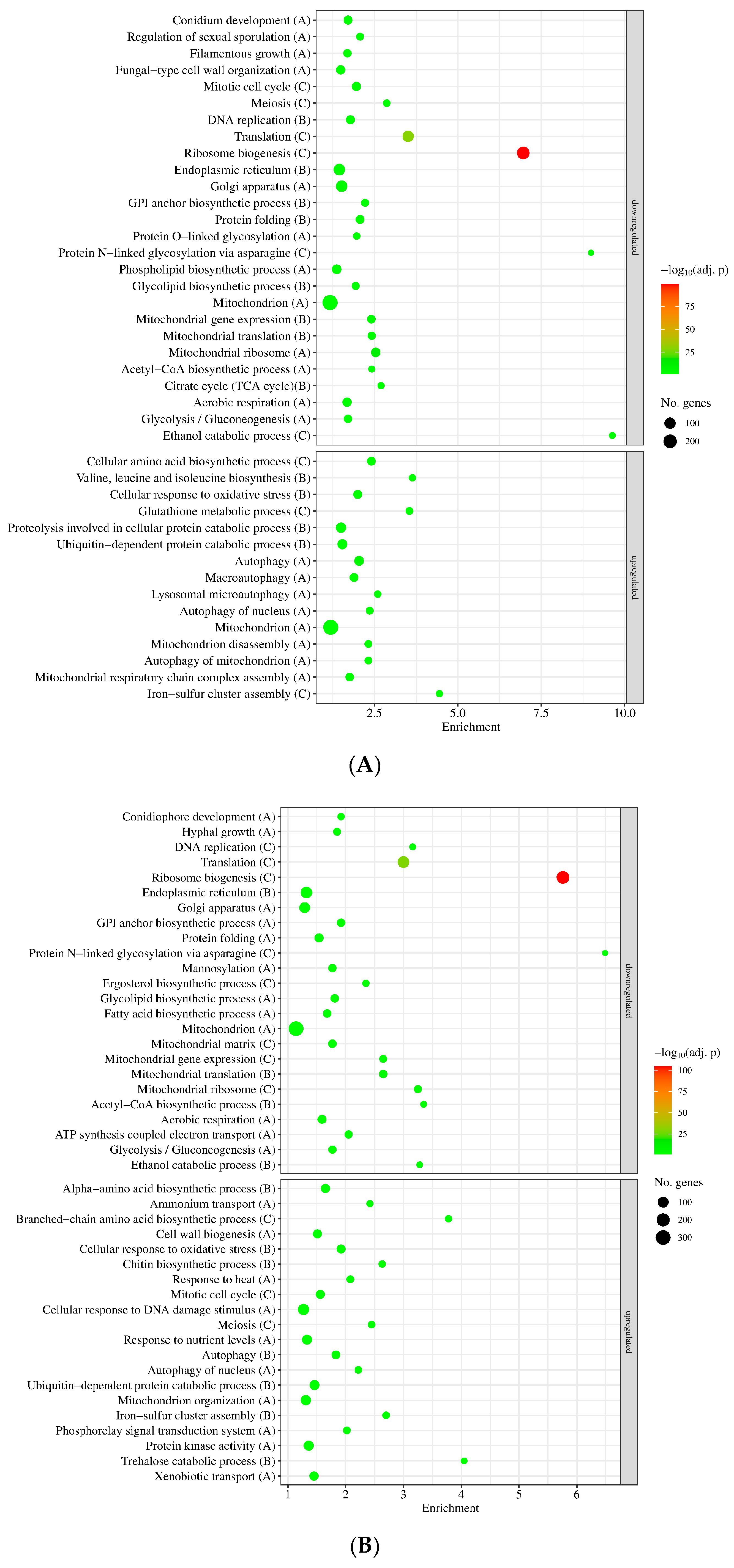
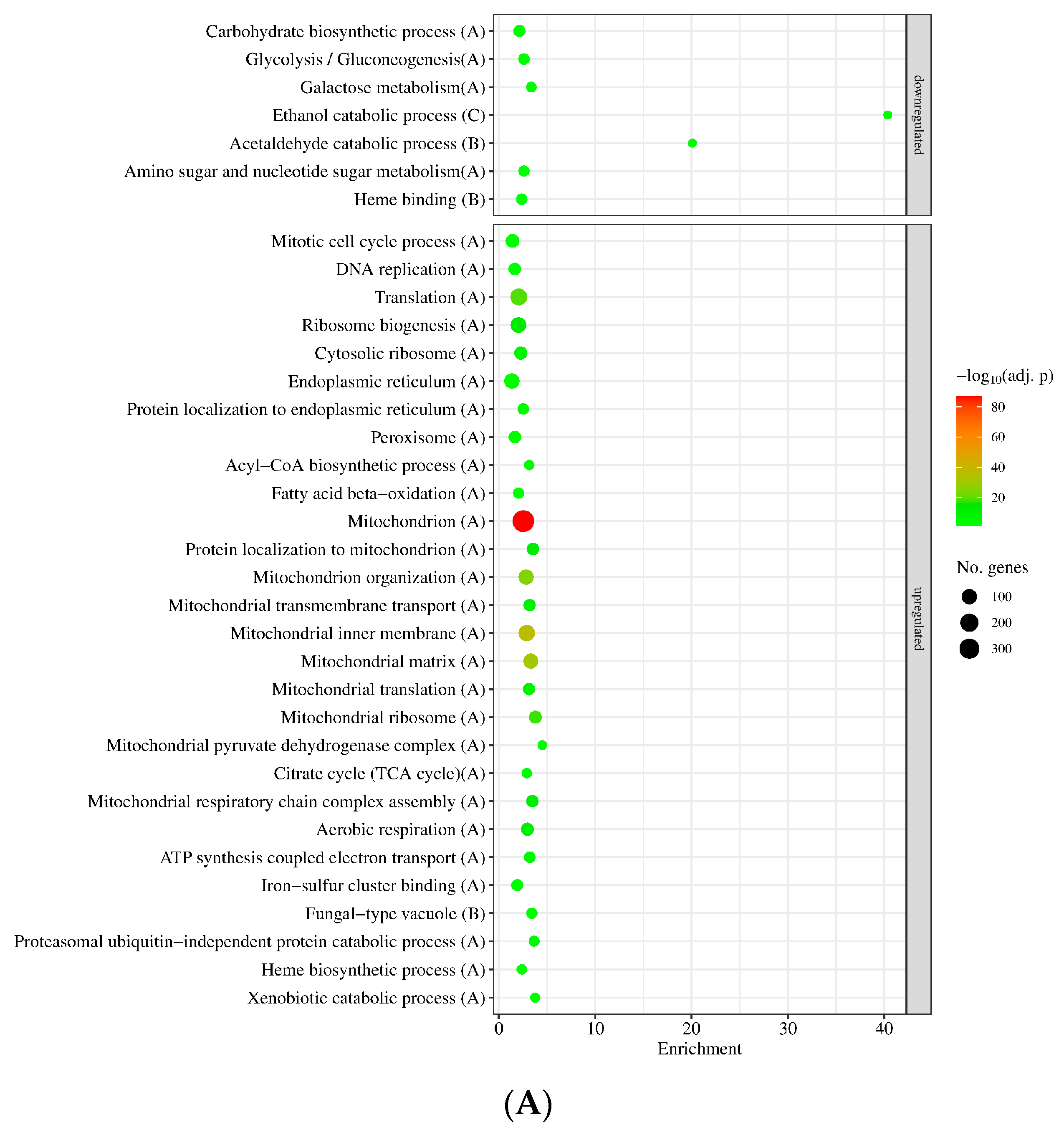
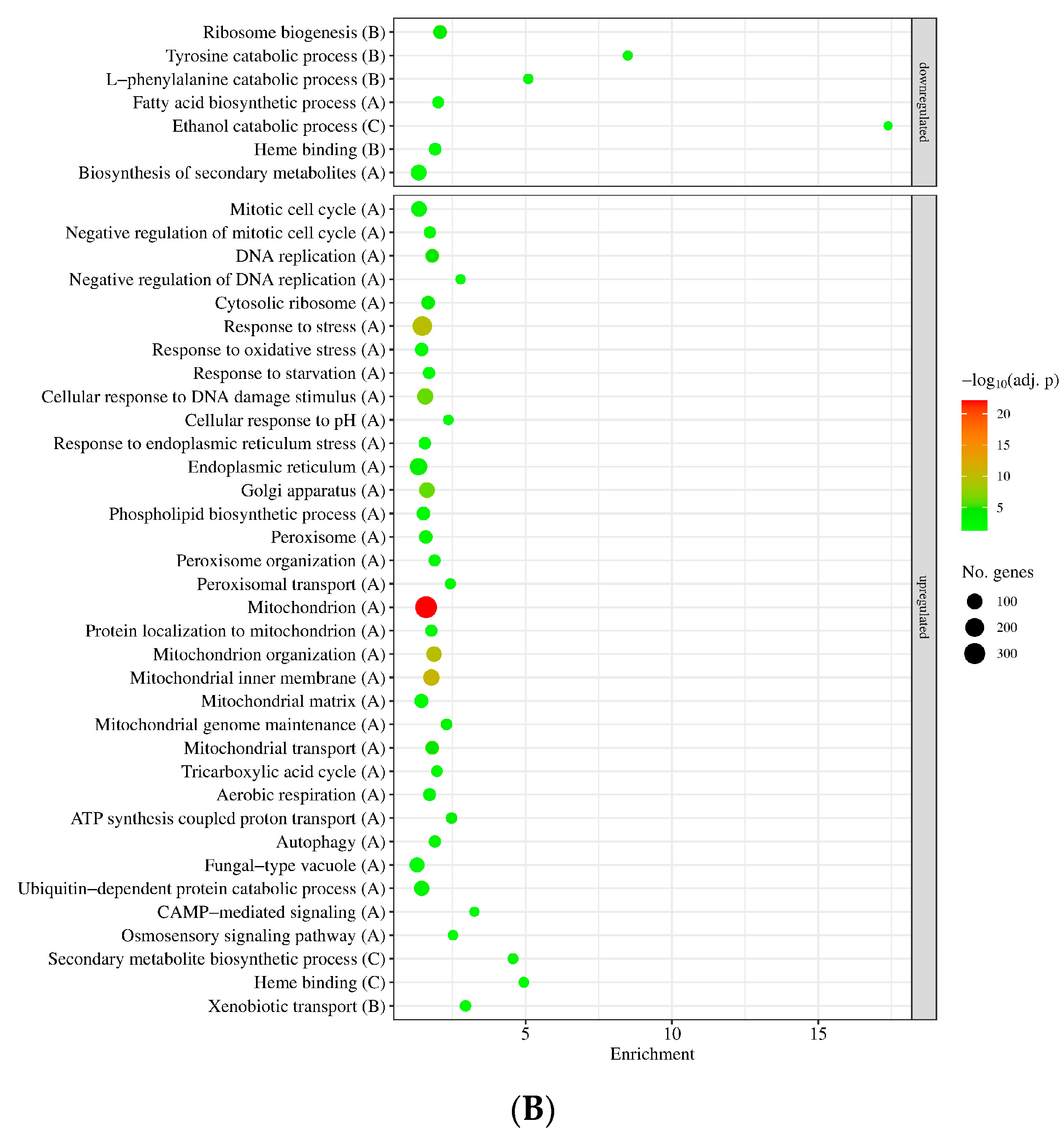
3.2. Genome-Wide Transcriptional Consequences of MSB Treatment and Deletion of the sodB Gene in A. nidulans
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miller, A.F. Superoxide dismutases: Ancient enzymes and new insights. FEBS Lett. 2012, 586, 585–595. [Google Scholar] [CrossRef] [PubMed]
- de Obeso Fernandez Del Valle, A.; Scheckhuber, C.Q. Superoxide dismutases in eukaryotic microorganisms: Four case studies. Antioxidants 2022, 11, 188. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.M.; Dirmeier, R.; Engle, M.; Poyton, R.O. Mitochondrial protein oxidation in yeast mutants lacking manganese-(MnSOD) or copper- and zinc-containing superoxide dismutase (CuZnSOD): Evidence that MnSOD and CuZnSOD have both unique and overlapping functions in protecting mitochondrial proteins from oxidative damage. J. Biol. Chem. 2004, 279, 51817–51827. [Google Scholar] [CrossRef]
- Fréalle, E.; Noël, C.; Nolard, N.; Symoens, F.; Felipe, M.S.; Dei-Cas, E.; Camus, D.; Viscogliosi, E.; Delhaes, L. Manganese superoxide dismutase based phylogeny of pathogenic fungi. Mol. Phylogenet. Evol. 2006, 41, 28–39. [Google Scholar] [CrossRef]
- Leiter, É.; Park, H.S.; Kwon, N.J.; Han, K.H.; Emri, T.; Oláh, V.; Mészáros, I.; Dienes, B.; Vincze, J.; Csernoch, L.; et al. Characterization of the aodA, dnmA, mnSOD and pimA genes in Aspergillus nidulans. Sci. Rep. 2016, 6, 20523. [Google Scholar] [CrossRef] [PubMed]
- Stuart, G.R.; Humble, M.H.; Strand, M.K.; William, C.; Copeland, W.C. Transcriptional response to mitochondrial NADH kinase deficiency in Saccharomyces cerevisiae. Mitochondrion 2009, 9, 211–221. [Google Scholar] [CrossRef]
- Stenberg, S.; Li, J.; Gjuvsland, A.B.; Persson, K.; Demitz-Helin, E.; Peña, C.G.; Yue, J.X.; Gilchrist, C.; Ärengård, T.; Ghiaci, P.; et al. Genetically controlled mtDNA deletions prevent ROS damage by arresting oxidative phosphorylation. eLife 2022, 11, e76095. [Google Scholar] [CrossRef]
- Knuppertz, L.; Warnsmann, V.; Hamann, A.; Grimm, C.; Osiewacz, H.D. Stress-dependent opposing roles for mitophagy in aging of the ascomycete Podospora anserina. Autophagy 2017, 13, 1037–1052. [Google Scholar] [CrossRef]
- Lambou, K.; Lamarre, C.; Beau, R.; Dufour, N.; Latgé, J.P. Functional analysis of the superoxide dismutase family in Aspergillus fumigatus. Mol. Microbiol. 2010, 75, 910–923. [Google Scholar] [CrossRef]
- Reverberi, M.; Punelli, M.; Scala, V.; Scarpari, M.; Uva, P.; Mentzen, W.I.; Dolezal, A.L.; Woloshuk, C.; Pinzari, F.; Fabbri, A.A.; et al. Genotypic and phenotypic versatility of Aspergillus flavus during maize exploitation. PLoS ONE 2013, 8, e68735. [Google Scholar] [CrossRef]
- Barratt, R.W.; Johnson, G.B.; Ogata, W.N. Wild type and mutant stocks of Aspergillus nidulans. Genetics 1965, 52, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Emri, T.; Pócsi, I.; Szentirmai, A. Analysis of the oxidative stress response of Penicillium chrysogenum to menadione. Free Radic. Res. 1999, 30, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Oberley, L.W.; Spitz, D.R. Assay of superoxide dismutase activity in tumour tissue. Meth. Enzymol. 1984, 105, 457–464. [Google Scholar]
- Chomczynski, P.A. Reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. BioTechniques 1993, 15, 532–534, 536–537. [Google Scholar]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and 169 HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Inglis, D.O.; Binkley, J.; Skrzypek, M.S.; Arnaud, M.B.; Cerqueira, G.C.; Shah, P.; Wymore, F.; Wortman, J.R.; Sherlock, G. Comprehensive annotation of secondary metabolite biosynthetic genes and gene clusters of Aspergillus nidulans, A. fumigatus, A. niger and A. oryzae. BMC Microbiol. 2013, 13, 91. [Google Scholar] [CrossRef]
- Karányi, Z.; Holb, I.; Hornok, L.; Pócsi, I.; Miskei, M. FSRD: Fungal stress response database. Database 2013, 2013, bat037. [Google Scholar] [CrossRef]
- Carter, W.O.; Narayanan, P.K.; Robinson, J.P. Intracellular hydrogen peroxide and superoxide anion detection in endothelial cells. J. Leukoc. Biol. 1994, 55, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Emri, T.; Szarvas, V.; Orosz, E.; Antal, K.; Park, H.; Han, K.; Yu, J.; Pócsi, I. Core oxidative stress response in Aspergillus nidulans. BMC Genom. 2015, 16, 478. [Google Scholar] [CrossRef] [PubMed]
- Orosz, E.; Antal, K.; Gazdag, Z.; Szabó, Z.; Han, K.H.; Yu, J.H.; Pócsi, I.; Emri, T. Transcriptome-based modeling reveals that oxidative stress induces modulation of the AtfA-dependent signaling networks in Aspergillus nidulans. Int. J. Genom. 2017, 2017, 6923849. [Google Scholar] [CrossRef]
- Antal, K.; Gila, B.C.; Pócsi, I.; Emri, T. General stress response or adaptation to rapid growth in Aspergillus nidulans? Fungal Biol. 2020, 124, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.Y.; Wu, J.; Miller, B.L. StuA is required for cell pattern formation in Aspergillus. Genes Dev. 1992, 6, 1770–1782. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.D.; Gralla, E.B.; Valentine, J.S. Superoxide dismutase activity is essential for stationary phase survival in Saccharomyces cerevisiae. Mitochondrial production of toxic oxygen species in vivo. J. Biol. Chem. 1996, 271, 12275–12280. [Google Scholar] [CrossRef]
- Fabrizio, P.; Liou, L.L.; Moy, V.N.; Diaspro, A.; Valentine, J.S.; Gralla, E.B.; Longo, V.D. SOD2 functions downstream of Sch9 to extend longevity in yeast. Genetics 2003, 163, 35–46. [Google Scholar] [CrossRef]
- Fabrizio, P.; Pletcher, S.D.; Minois, N.; Vaupel, J.W.; Longo, V.D. Chronological aging-independent replicative life span regulation by Msn2/Msn4 and Sod2 in Saccharomyces cerevisiae. FEBS Lett. 2004, 557, 136–142. [Google Scholar] [CrossRef]
- Giles, S.S.; Batinic-Haberle, I.; Perfect, J.R.; Cox, G.M. Cryptococcus neoformans mitochondrial superoxide dismutase: An essential link between antioxidant function and high-temperature growth. Eukaryot. Cell 2005, 4, 46–54. [Google Scholar] [CrossRef]
- Narasipura, S.D.; Chaturvedi, V.; Chaturvedi, S. Characterization of Cryptococcus neoformans variety gattii SOD2 reveals distinct roles of the two superoxide dismutases in fungal biology and virulence. Mol. Microbiol. 2005, 55, 1782–1800. [Google Scholar] [CrossRef]
- Xie, X.Q.; Li, F.; Ying, S.H.; Feng, M.G. Additive contributions of two manganese-cored superoxide dismutases (MnSODs) to antioxidation, UV tolerance and virulence of Beauveria bassiana. PLoS ONE 2012, 7, e30298. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.G.; Tong, S.M.; Ying, S.H.; Ming-Guang Feng, M.G. Antioxidant activities of four superoxide dismutases in Metarhizium robertsii and their contributions to pest control potential. Appl. Microbiol. Biotechnol. 2018, 102, 9221–9230. [Google Scholar] [CrossRef] [PubMed]
- Montibus, M.; Ducos, C.; Bonnin-Verdal, M.N.; Bormann, J.; Ponts, N.; Richard-Forget, F.; Christian Barreau, C. The bZIP transcription factor Fgap1 mediates oxidative stress response and trichothecene biosynthesis but not virulence in Fusarium graminearum. PLoS ONE 2013, 8, e83377. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, T.; Yoshinari, T.; Sakuda, S. Intracellular superoxide level controlled by manganese superoxide dismutases affects trichothecene production in Fusarium graminearum. FEMS Microbiol. Lett. 2017, 364, fnx213. [Google Scholar] [CrossRef] [PubMed]
- Novodvorska, M.; Hayer, K.; Pullan, S.T.; Wilson, R.; Blythe, M.J.; Stam, H.; Stratford, M.; Archer, D.B. Transcriptional landscape of Aspergillus niger at breaking of conidial dormancy revealed by RNA-sequencing. BMC Genom. 2013, 14, 246. [Google Scholar] [CrossRef]
- Van Leeuwen, M.R.; Krijgsheld, P.; Bleichrodt, R.; Menke, H.; Stam, H.; Stark, J.; Wösten, H.A.B.; Dijksterhuis, J. Germination of conidia of Aspergillus niger is accompanied by major changes in RNA profiles. Stud. Mycol. 2013, 74, 59–70. [Google Scholar] [CrossRef]
- Imlay, J.A. Iron-sulphur clusters and the problem with oxygen. Mol. Microbiol. 2006, 59, 1073–1082. [Google Scholar] [CrossRef]
- Balázs, A.; Pócsi, I.; Hamari, Z.; Leiter, É.; Emri, T.; Miskei, M.; Oláh, J.; Tóth, V.; Hegedűs, N.; Prade, R.A.; et al. AtfA BZIP-type transcription factor regulates oxidative and osmotic stress responses in Aspergillus nidulans. Mol. Genet. Genom. 2010, 283, 289–303. [Google Scholar] [CrossRef]
- Boczonádi, I.; Török, Z.; Jakab, Á.; Kónya, G.; Gyurcsó, K.; Baranyai, E.; Szoboszlai, Z.; Döncző, B.; Fábián, I.; Leiter, É.; et al. Increased Cd2+ biosorption capability of Aspergillus nidulans elicited by crpA deletion. J. Basic Microbiol. 2020, 60, 574–584. [Google Scholar] [CrossRef]
- Lalève, A.; Vallières, C.; Golinelli-Cohen, M.P.; Bouton, C.; Song, Z.; Pawlik, G.; Tindall, S.M.; Avery, S.V.; Clain, J.; Meunier, B. The antimalarial drug primaquine targets Fe-S cluster proteins and yeast respiratory growth. Redox Biol. 2016, 7, 21–29. [Google Scholar] [CrossRef]
- Zyrina, A.N.; Smirnova, E.A.; Markova, O.V.; Severin, F.F.; Dmitry, A.; Knorre, D.A. Mitochondrial superoxide dismutase and Yap1p act as a signaling module contributing to ethanol tolerance of the yeast Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2017, 83, e02759-16. [Google Scholar] [CrossRef] [PubMed]
- Grant, C.M. Role of the glutathione/glutaredoxin and thioredoxin systems in yeast growth and response to stress conditions. Mol. Microbiol. 2001, 39, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Pócsi, I.; Prade, R.A.; Penninckx, M.J. Glutathione, altruistic metabolite in fungi. Adv. Microb. Physiol. 2004, 49, 1–76. [Google Scholar] [CrossRef] [PubMed]
- Muid, K.A.; Karakaya, H.C.; Ahmet Koc, A. Absence of superoxide dismutase activity causes nuclear DNA fragmentation during the aging process. Biochem. Biophys. Res. Commun. 2004, 444, 260–263. [Google Scholar] [CrossRef]
- Ogata, T.; Senoo, T.; Kawano, S.; Shogo Ikeda, S. Mitochondrial superoxide dismutase deficiency accelerates chronological aging in the fission yeast Schizosaccharomyces pombe. Cell Biol. Int. 2016, 40, 100–106. [Google Scholar] [CrossRef]
- Bradshaw, R.E.; Bird, D.M.; Brown, S.; Gardiner, R.E.; Hirst, P. Cytochrome c is not essential for viability of the fungus Aspergillus nidulans. Mol. Genet. Genom. 2001, 266, 48–55. [Google Scholar] [CrossRef]
- Molnár, Á.P.; Németh, Z.; Fekete, E.; Flipphi, M.; Keller, N.P.; Karaffa, L. Analysis of the relationship between alternative respiration and sterigmatocystin formation in Aspergillus nidulans. Toxins 2018, 10, 168. [Google Scholar] [CrossRef]
- Adams, T.H.; Wieser, J.K.; Yu, J.H. Asexual sporulation in Aspergillus nidulans. Microbiol. Mol. Biol. Rev. 1998, 62, 35–54. [Google Scholar] [CrossRef]
- Kocsis, B.; Lee, M.K.; Antal, K.; Yu, J.H.; Pócsi, I.; Leiter, É.; Emri, T. Genome-wide gene expression analyses of the AtfA/AtfB-mediated menadione stress response in Aspergillus nidulans. Cells 2023, 12, 463. [Google Scholar] [CrossRef]
- Taubitz, A.; Bauer, B.; Heesemann, J.; Ebel, F. Role of respiration in the germination process of the pathogenic mold Aspergillus fumigatus. Curr. Microbiol. 2007, 54, 354–360. [Google Scholar] [CrossRef]
- Grahl, N.; Dinamarco, T.M.; Willger, S.D.; Goldman, G.H.; Cramer, R.A. Aspergillus fumigatus mitochondrial electron trans-port chain mediates oxidative stress homeostasis, hypoxia responses and fungal pathogenesis. Mol. Microbiol. 2012, 84, 383–399. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pákozdi, K.; Emri, T.; Antal, K.; Pócsi, I. Global Transcriptomic Changes Elicited by sodB Deletion and Menadione Exposure in Aspergillus nidulans. J. Fungi 2023, 9, 1060. https://doi.org/10.3390/jof9111060
Pákozdi K, Emri T, Antal K, Pócsi I. Global Transcriptomic Changes Elicited by sodB Deletion and Menadione Exposure in Aspergillus nidulans. Journal of Fungi. 2023; 9(11):1060. https://doi.org/10.3390/jof9111060
Chicago/Turabian StylePákozdi, Klaudia, Tamás Emri, Károly Antal, and István Pócsi. 2023. "Global Transcriptomic Changes Elicited by sodB Deletion and Menadione Exposure in Aspergillus nidulans" Journal of Fungi 9, no. 11: 1060. https://doi.org/10.3390/jof9111060
APA StylePákozdi, K., Emri, T., Antal, K., & Pócsi, I. (2023). Global Transcriptomic Changes Elicited by sodB Deletion and Menadione Exposure in Aspergillus nidulans. Journal of Fungi, 9(11), 1060. https://doi.org/10.3390/jof9111060