
Selection of Autochthonous Yeasts Isolated from the Intestinal Tracts of Cobia Fish (Rachycentron canadum) with Probiotic Potential

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Intestinal Samples
2.2. Isolation of Yeast from Intestinal Samples by Culture Method
2.3. DNA Extraction
2.4. Sequencing for Yeast Identification
2.5. Specific PCR for Debaryomyces hansenii Identification
2.6. RAPD-PCR Profile
2.7. Yeast Biomass Production
2.8. Enzymatic Activity
2.9. Hemolytic Activity
2.10. Antagonistic Activity against Vibrio spp
2.11. Biofilm Production
2.12. Hydrophobicity Test
2.13. Autoaggregation Test
2.14. Polyamine Production
2.15. Effect of Yeast Strains on Larval Survival
2.16. Protective Effect of Cobia Larvae by Yeasts against a Salinity Stress
2.17. Yeast Growth Curve
2.18. Viability of Yeast Cells Stored at 4 °C
2.19. Statistical Analysis
3. Results
3.1. Criteria to Select Probiotic Yeasts
3.2. Isolation and Identification of Yeasts Isolates from Cobia
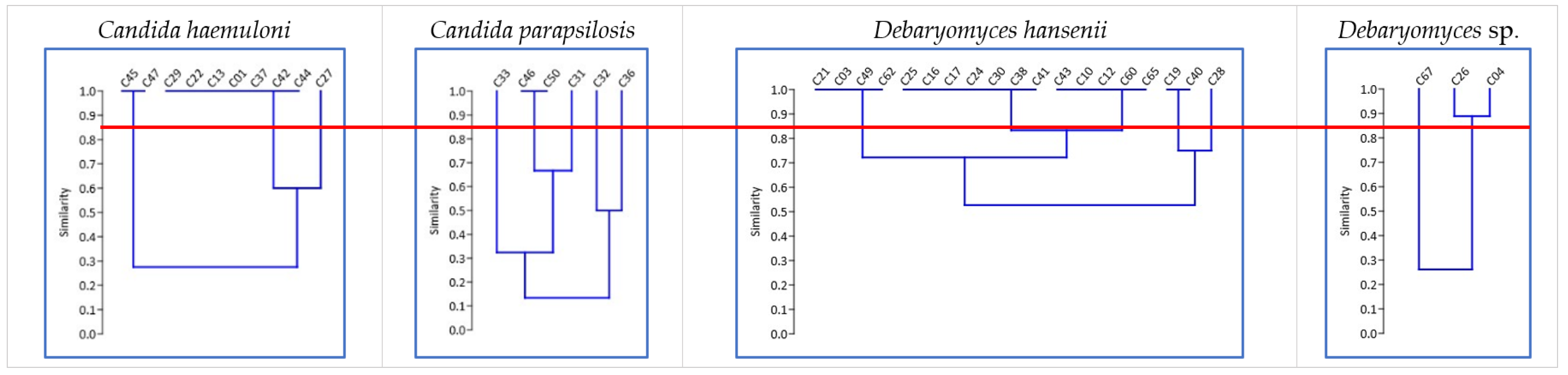
3.3. First Selection of Yeasts by RAPD-PCR Profile and Biomass Production
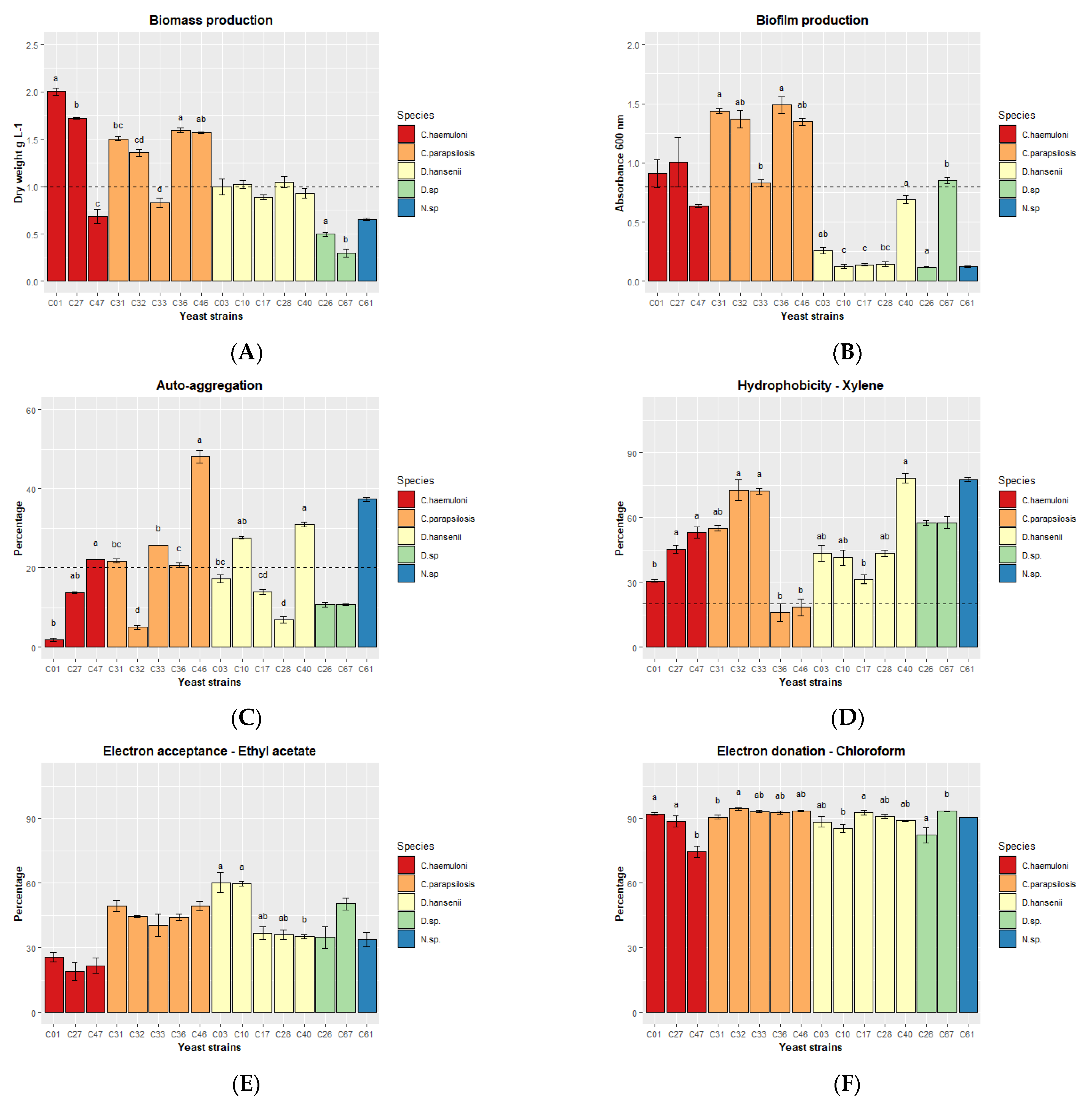
3.4. Second Selection of Yeast Strains Based on their Enzymatic, Hemolytic, and Antipathogen Activity and their Potential to Adhere to Intestinal Mucosa
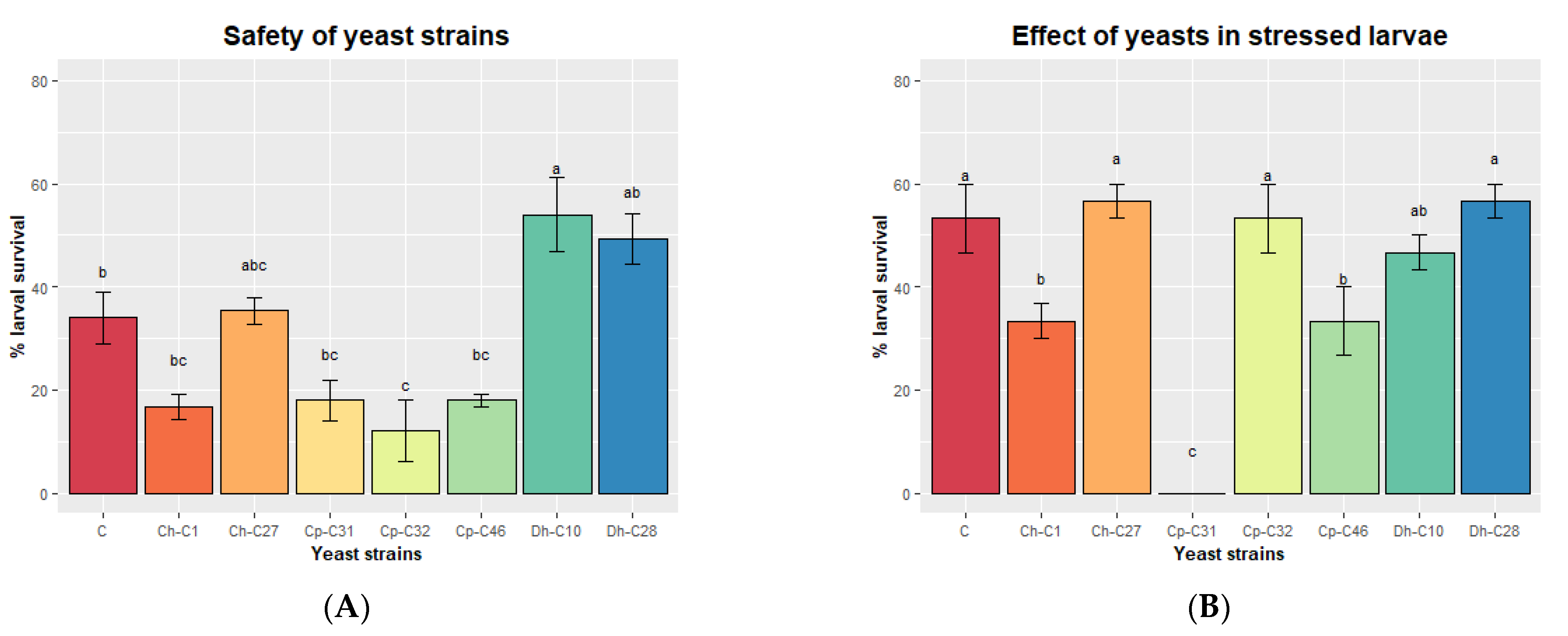
3.5. Third Selection Based on the Polyamine Production, Safety, and Protection of Larvae against a Saline Stress
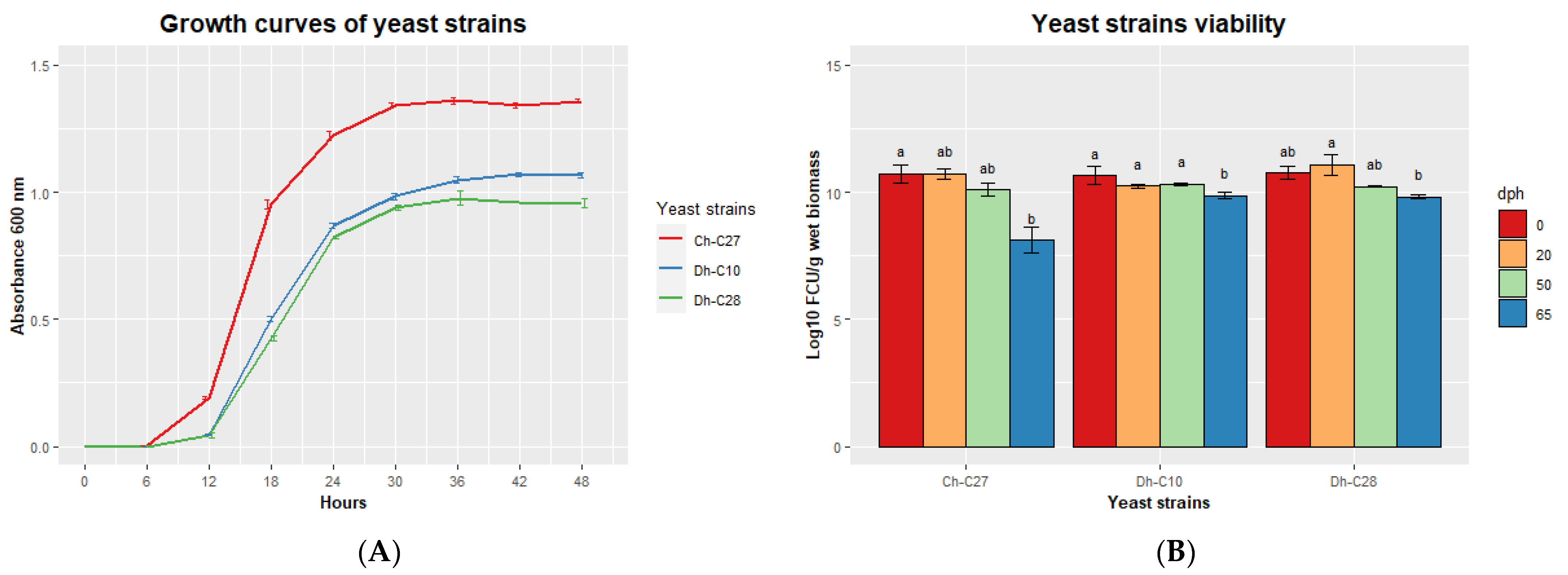
3.6. Characterization of Selected Potential Probiotic Yeast Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022; ISBN 978-92-5-136364-5. [Google Scholar]
- Qin, J.G. (Ed.) Larval Fish Aquaculture; Nova Science Publishers, Inc.: New York, NY, USA, 2013; ISBN 6312317269. [Google Scholar]
- Froese, R.; Pauly, D. Rachycentron Canadum (Linnaeus, 1766), Cobia. Available online: https://www.fishbase.se/summary/Rachycentron-canadum (accessed on 3 October 2022).
- Liao, I.C.; Huang, T.S.; Tsai, W.S.; Hsueh, C.M.; Chang, S.L.; Leaño, E.M. Cobia Culture in Taiwan: Current Status and Problems. Aquaculture 2004, 237, 155–165. [Google Scholar] [CrossRef]
- Gopakumar, G.; Abdul Nazar, A.K.; Tamilmani, G.; Sakthivel, M.; Kalidas, C.; Ramamoorthy, N.; Palanichamy, S.; Ashok Maharshi, V.; Srinivasa Rao, K.; Syda Rao, G. First Experience in the Larviculture of Cobia, Rachycentron Canadum (Linnaeus, 1752) in India. Indian J. Fish. 2012, 59, 59–63. [Google Scholar]
- Benetti, D.D.; Suarez, J.; Camperio, J.; Hoenig, R.H.; Tudela, C.E.; Daugherty, Z.; McGuigan, C.J.; Mathur, S.; Anchieta, L.; Buchalla, Y.; et al. A Review on Cobia, Rachycentron Canadum, Aquaculture. J. World Aquac. Soc. 2021, 52, 691–709. [Google Scholar] [CrossRef]
- Petersen, E.H.; Luan, T.D.; Chinh, D.T.M.; Tuan, V.A.; Binh, T.Q.; Van Truc, L.; Glencross, B.D. Bioeconomics of Cobia, Rachycentron Canadum, Culture in Vietnam. Aquac. Econ. Manag. 2014, 18, 28–44. [Google Scholar] [CrossRef]
- FAO. Global Aquaculture Production 1950–2020 (FishstatJ); Fishery and Aquaculture Statistics: Rome, Italy, 2022. [Google Scholar]
- Hamre, K.; Yúfera, M.; Rønnestad, I.; Boglione, C.; Conceição, L.; Izquierdo, M. Fish Larval Nutrition and Feed Formulation: Knowledge Gaps and Bottlenecks for Advances in Larval Rearing. In Success factors for fish larval production; Conceição, L., Tandler, A., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2018; pp. 200–280. [Google Scholar]
- Nguyen, H.Q.; Reinertsen, H.; Wold, P.A.; Tran, M.T.; Kjørsvik, E. Effects of Early Weaning Strategies on Growth, Survival and Digestive Enzyme Activities in Cobia (Rachycentron Canadum L.) Larvae. Aquac. Int. 2011, 19, 63–78. [Google Scholar] [CrossRef]
- Nhu, V.C.; Dierckens, K.; Nguyen, H.T.; Hoang, T.M.T.; Le, T.L.; Tran, M.T.; Nys, C.; Sorgeloos, P. Effect of Early Co-Feeding and Different Weaning Diets on the Performance of Cobia (Rachycentron Canadum) Larvae and Juveniles. Aquaculture 2010, 305, 52–58. [Google Scholar] [CrossRef]
- Benetti, D.; Sardenberg, B.; Welch, A.; Hoenig, R.; Orhun, M.R.; Zink, I. Intensive Larval Husbandry and Fingerling Production of Cobia Rachycentron Canadum. Aquaculture 2008, 281, 22–27. [Google Scholar] [CrossRef]
- Gisbert, E.; Morais, S.; Moyano, F.J. Feeding and Digestion. In Larval fish aquaculture; Qin, J.G., Ed.; Nova Publishers: New York, Ny, USA, 2013; pp. 73–124. ISBN 978-1-62618-152-6. [Google Scholar]
- Amenyogbe, E.; Huang, J.; Chen, G.; Wang, W. Probiotic Potential of Indigenous (Bacillus Sp. RCS1, Pantoea Agglomerans RCS2, and Bacillus Cereus Strain RCS3) Isolated From Cobia Fish (Rachycentron Canadum) and Their Antagonistic Effects on the Growth of Pathogenic Vibrio Alginolyticus, Vibrio Harveyi, Streptococcus Iniae, and Streptococcus Agalactiae. Front. Mar. Sci. 2021, 8, 560. [Google Scholar] [CrossRef]
- Garrido-Pereira, M.A.; Schwarz, M.; Delbos, B.; Rodrigues, R.V.; Romano, L.; Sampaio, L. Efectos Probióticos Sobre Las Larvas de Cobia Rachycentron Canadum Criadas En Un Sistema de Recirculación de Agua. Lat. Am. J. Aquat. Res. 2014, 42, 1169–1174. [Google Scholar] [CrossRef]
- Hitzfelder, G.M.; Holt, G.J.; Fox, J.M.; McKee, D.A. The Effect of Rearing Density on Growth and Survival of Cobia, Rachycentron Canadum, Larvae in Closed Recirculating Aquaculture System. J. World Aquac. Soc. 2006, 37, 204–209. [Google Scholar] [CrossRef]
- Yufera, M. Feeding Behavior in Larval Fish. In Larval Fish Nutrition; Holt, G.J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 285–307. ISBN 9780813817927. [Google Scholar]
- Faulk, C.K.; Holt, G.J. Advances in Rearing Cobia Rachycentron Canadum Larvae in Recirculating Aquaculture Systems: Live Prey Enrichment and Greenwater Culture. Aquaculture 2005, 249, 231–243. [Google Scholar] [CrossRef]
- Salze, G.; McLean, E.; Craig, S.R. Dietary Taurine Enhances Growth and Digestive Enzyme Activities in Larval Cobia. Aquaculture 2012, 362–363, 44–49. [Google Scholar] [CrossRef]
- Salze, G.; Craig, S.R.; Smith, B.H.; Smith, E.P.; McLean, E. Morphological Development of Larval Cobia Rachycentron Canadum and the Influence of Dietary Taurine Supplementation. J. Fish Biol. 2011, 78, 1470–1491. [Google Scholar] [CrossRef] [PubMed]
- Salze, G.; McLean, E.; Schwarz, M.; Craig, S.R. Dietary Mannan Oligosaccharide Enhances Salinity Tolerance and Gut Development of Larval Cobia. Aquaculture 2008, 274, 148–152. [Google Scholar] [CrossRef]
- Nayak, S.K. Role of Gastrointestinal Microbiota in Fish. Aquac. Res. 2010, 41, 1553–1573. [Google Scholar] [CrossRef]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic Bacteria as Biological Control Agents in Aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef] [Green Version]
- Thirumurugan, R.; Vignesh, V. Probiotics: Live Boon to Aquaculture. In Advances in Marine and Brackishwater Aquaculture; Perumal, S., Thirunavukkarasu, A., Pachiappan, Eds.; Springer India: New Delhi, India, 2015; pp. 51–61. ISBN 9788132222712. [Google Scholar]
- Navarrete, P.; Tovar-Ramírez, D. Use of Yeasts as Probiotics in Fish Aquaculture. In Sustainable Aquaculture Techniques; Hernandez-Vergara, M., Perez-Rostro, C., Eds.; IntechOpen: Rijeka, Croatia, 2014; pp. 135–172. ISBN 978-953-51-1224-2. [Google Scholar]
- Raggi, P.; Lopez, P.; Diaz, A.; Carrasco, D.; Silva, A.; Velez, A.; Opazo, R.; Magne, F.; Navarrete, P. Debaryomyces Hansenii and Rhodotorula Mucilaginosa Comprised the Yeast Core Gut Microbiota of Wild and Reared Carnivorous Salmonids, Croaker and Yellowtail. Environ. Microbiol. 2014, 16, 2791–2803. [Google Scholar] [CrossRef]
- Gatesoupe, F.J. Live Yeasts in the Gut: Natural Occurrence, Dietary Introduction, and Their Effects on Fish Health and Development. Aquaculture 2007, 267, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Gotcheva, V.; Hristozova, E.; Hristozova, T.; Guo, M.; Roshkova, Z.; Angelov, A. Assessment of Potential Probiotic Properties of Lactic Acid Bacteria and Yeast Strains. Food Biotechnol. 2002, 16, 211–225. [Google Scholar] [CrossRef]
- Neut, C.; Mahieux, S.; Dubreuil, L.J. Antibiotic Susceptibility of Probiotic Strains: Is It Reasonable to Combine Probiotics with Antibiotics? Med. Mal. Infect. 2017, 47, 477–483. [Google Scholar] [CrossRef]
- Romero, J.; Feijoó, C.G.; Navarrete, P. Antibiotics in Aquaculture-Use, Abuse and Alternatives. In Health and Environment in Aquaculture; Carvalho, E., Silva, G., Silva, R., Eds.; IntechOpen: Rijeka, Croatia, 2012; pp. 159–198. ISBN 978-953-51-0497-1. [Google Scholar]
- Andlid, T.; Vasquez-Juárez, R.; Gustaffson, L. Yeast Colonizing the Intestine of Rainbow Trout (Salmo Gairdneri) and Turbot (Scophtalmus Maximus). Microb. Ecol. 1995, 30, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Andlid, T.; Vázquez-Juárez, R.; Gustafsson, L. Yeasts Isolated from the Intestine of Rainbow Trout Adhere to and Grow in Intestinal Mucus. Mol. Mar. Biol. Biotechnol. 1998, 7, 115–126. [Google Scholar] [PubMed]
- Tovar, D. Potencial Probiótico de Levaduras Productoras de Poliaminas En El Desarrollo Del Sistema Digestivo de La Lubina Europea y La Cabrilla Arenera; CIBNOR: La Paz, Mexico, 2002. [Google Scholar]
- Caruffo, M.; Navarrete, N.; Salgado, O.; Díaz, A.; López, P.; García, K.; Feijóo, C.G.; Navarrete, P. Potential Probiotic Yeasts Isolated from the Fish Gut Protect Zebrafish (Danio Rerio) from a Vibrio Anguillarum Challenge. Front. Microbiol. 2015, 6, 1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutty, S.; Philip, R. Marine Yeasts—A Review. Yeast 2008, 25, 465–483. [Google Scholar] [CrossRef] [PubMed]
- Valderrama, B.; Ruiz, J.J.; Gutiérrez, M.S.; Alveal, K.; Caruffo, M.; Oliva, M.; Flores, H.; Silva, A.; Toro, M.; Reyes-Jara, A.; et al. Cultivable Yeast Microbiota from the Marine Fish Species Genypterus Chilensis and Seriolella Violacea. J. Fungi 2021, 7, 515. [Google Scholar] [CrossRef]
- Reyes-Becerril, M.; Esteban, M.Á.; Tovar-Ramírez, D.; Ascencio-Valle, F. Polyamine Determination in Different Strains of the Yeast Debaryomyces Hansenii by High Pressure Liquid Chromatography. Food Chem. 2011, 127, 1862–1865. [Google Scholar] [CrossRef]
- Tovar-Ramírez, D.; Zambonino-Infante, J.L.; Cahu, C.L.; Gatesoupe, F.J.; Vázquez-Juárez, R. Influence of Dietary Live Yeast on European Sea Bass (Dicentrarchus Labrax) Larval Development. Aquaculture 2004, 234, 415–427. [Google Scholar] [CrossRef] [Green Version]
- Tovar-Ramírez, D.; Mazurais, D.; Gatesoupe, F.J.; Quazuguel, P.; Cahu, C.L.; Zambonino-Infante, J.L. Dietary Probiotic Live Yeast Modulates Antioxidant Enzyme Activities and Gene Expression of Sea Bass (Dicentrarchus Labrax) Larvae. Aquaculture 2010, 300, 142–147. [Google Scholar] [CrossRef] [Green Version]
- Caruffo, M.; Navarrete, N.C.; Salgado, O.A.; Faúndez, N.B.; Gajardo, M.C.; Feijóo, C.G.; Reyes-Jara, A.; García, K.; Navarrete, P. Protective Yeasts Control V. Anguillarum Pathogenicity and Modulate the Innate Immune Response of Challenged Zebrafish (Danio Rerio) Larvae. Front. Cell. Infect. Microbiol. 2016, 6, 127. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.H.; Cheng, C.H.; Gua, W.R.; Guu, Y.K.; Cheng, W. Dietary Administration of the Probiotic, Saccharomyces Cerevisiae P13, Enhanced the Growth, Innate Immune Responses, and Disease Resistance of the Grouper, Epinephelus Coioides. Fish Shellfish Immunol. 2010, 29, 1053–1059. [Google Scholar] [CrossRef]
- Reyes-Becerril, M.; Tovar-Ramírez, D.; Ascencio-Valle, F.; Civera-Cerecedo, R.; Gracia-López, V.; Barbosa-Solomieu, V.; Esteban, M.Á. Effects of Dietary Supplementation with Probiotic Live Yeast Debaryomyces Hansenii on the Immune and Antioxidant Systems of Leopard Grouper Mycteroperca Rosacea Infected with Aeromonas Hydrophila. Aquac. Res. 2011, 42, 1676–1686. [Google Scholar] [CrossRef]
- Ross, S.S.; Morris, E.O. An Investigation of the Yeast Flora of Marine Fish from Scottish Coastal Waters and a Fishing Ground off Iceland. J. Appl. Bacteriol. 1965, 28, 224–234. [Google Scholar] [CrossRef]
- Tovar, D.; Zambonino-Infante, J.L.; Cahu, C.L.; Gatesoupe, F.J.; Vázquez-Juárez, R.; Lésel, R. Effect of Live Yeast Incorporation in Compound Diet on Digestive Enzyme Activity in Sea Bass (Dicentrarchus Labrax) Larvae. Aquaculture 2002, 204, 113–123. [Google Scholar] [CrossRef]
- Apolinario Castillo, D.A. Composición Química Proximal de Tres Especies de Peces Pelágicos Pequeños de Importancia Comercial En El Puerto Pesquero de Anconcito, La Libertad; Universidad Estatal Península de Santa Elena: La Libertad, Ecuador, 2017. [Google Scholar]
- Green, M.R.; Sambrook, J. Molecular Cloning, A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012; ISBN 978-1-936113-41-5. [Google Scholar]
- Wrent, P.; Rivas, E.M.; Gil de Prado, E.; Peinado, J.M.; de Silóniz, M.I. Development of Species-Specific Primers for Rapid Identification of Debaryomyces Hansenii. Int. J. Food Microbiol. 2015, 193, 109–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Bhathena, Z.P.; Kumar, S.; Srivastava, P.P.; Jadhao, S.B. Quantification and Characterization of Mannan Oligosaccharide Producing Yeasts Isolated from Various Food Products. Proc. Natl. Acad. Sci. India Sect. B-Biol. Sci. 2018, 88, 1237–1247. [Google Scholar] [CrossRef]
- Padilla, B.; Manzanares, P.; Belloch, C. Yeast Species and Genetic Heterogeneity within Debaryomyces Hansenii along the Ripening Process of Traditional Ewes’ and Goats’ Cheeses. Food Microbiol. 2014, 38, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Souza, F.A.; da Silva, V.G.; Bitencourt, T.B. Use of McFarland Standards and Spectrophotometry for Yarrowia Lipolytica QU69 Cell Counting. Int. J. Environ. Agric. Biotechnol. 2020, 5, 1089–1091. [Google Scholar] [CrossRef]
- Guinea, J.; Recio, S.; Escribano, P.; Torres-Narbona, M.; Peláez, T.; Sánchez-Carrillo, C.; Rodríguez-Créixems, M.; Bouza, E. Rapid Antifungal Susceptibility Determination for Yeast Isolates by Use of Etest Performed Directly on Blood Samples from Patients with Fungemia. J. Clin. Microbiol. 2010, 48, 2205–2212. [Google Scholar] [CrossRef] [Green Version]
- Amoah, K.; Dong, X.H.; Tan, B.P.; Zhang, S.; Kuebutornye, F.K.A.; Chi, S.Y.; Yang, Q.H.; Liu, H.Y.; Zhang, H.T.; Yang, Y.Z. In Vitro Assessment of the Safety and Potential Probiotic Characteristics of Three Bacillus Strains Isolated From the Intestine of Hybrid Grouper (Epinephelus Fuscoguttatus♀ × Epinephelus Lanceolatus♂). Front. Vet. Sci. 2021, 8, 426. [Google Scholar] [CrossRef]
- Domínguez-Borbor, C.; Ardiles, V.; Bermeo, M.; Bolívar-Alvarado, C.; Tomalá, C.; Sonnenholzner, S.; Rodríguez, J.A. The Marine Symbiont Pseudovibrio Denitrificans, Is Effective to Control Pathogenic Vibrio Spp. in Shrimp Aquaculture. Aquaculture 2019, 508, 127–136. [Google Scholar] [CrossRef]
- Restrepo, L.; Domínguez-Borbor, C.; Bajaña, L.; Betancourt, I.; Rodríguez, J.; Bayot, B.; Reyes, A. Microbial Community Characterization of Shrimp Survivors to AHPND Challenge Test Treated with an Effective Shrimp Probiotic (Vibrio Diabolicus). Microbiome 2021, 9, 88. [Google Scholar] [CrossRef]
- Vargas, O.; Gutiérrez, M.S.; Caruffo, M.; Valderrama, B.; Medina, D.A.; García, K.; Reyes-Jara, A.; Toro, M.; Feijóo, C.G.; Navarrete, P. Probiotic Yeasts and Vibrio Anguillarum Infection Modify the Microbiome of Zebrafish Larvae. Front. Microbiol. 2021, 12, 1639. [Google Scholar] [CrossRef]
- Naghmouchi, K.; Drider, D.; Kheadr, E.; Lacroix, C.C.; Prévost, H.; Fliss, I. Multiple Characterizations of Listeria Monocytogenes Sensitive and Insensitive Variants to Divergicin M35, a New Pediocin-like Bacteriocin. J. Appl. Microbiol. 2006, 100, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.M.; Kothari, D.; Lee, W.D.; Lim, J.M.; Khosravi, S.; Lee, S.M.; Lee, B.J.; Kim, K.W.; Han, H.S.; Kim, S.K. Autochthonous Bacillus Licheniformis: Probiotic Potential and Survival Ability in Low-Fishmeal Extruded Pellet Aquafeed. Microbiologyopen 2019, 8, e00767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, M. Bacterial Adherence to Hydrocarbons: A Useful Technique for Studying Cell Surface Hydrophobicity. FEMS Microbiol. Lett. 1984, 22, 289–295. [Google Scholar] [CrossRef]
- Bellon-Fontaine, M.N.; Rault, J.; Van Oss, C.J. Microbial Adhesion to Solvents: A Novel Method to Determine the Electron-Donor/Electron-Acceptor or Lewis Acid-Base Properties of Microbial Cells. Colloids Surf. B Biointerfaces 1996, 7, 47–53. [Google Scholar] [CrossRef]
- Hazen, K.C.; Plotkin, B.J.; Klimas, D.M. Influence of Growth Conditions on Cell Surface Hydrophobicity of Candida Albicans and Candida Glabrata. Infect. Immun. 1986, 54, 269–271. [Google Scholar] [CrossRef] [Green Version]
- Galarce, O.; Henríquez-Aedo, K.; Peterssen, D.; Peña-Farfal, C.; Aranda, M. A Selective Chromatographic Method to Determine the Dynamic of Biogenic Amines During Brewing Process. Food Anal. Methods 2016, 9, 3385–3395. [Google Scholar] [CrossRef]
- Faulk, C.K.; Benninghoff, A.D.; Holt, G.J. Ontogeny of the Gastrointestinal Tract and Selected Digestive Enzymes in Cobia Rachycentron Canadum (L.). J. Fish Biol. 2007, 70, 567–583. [Google Scholar] [CrossRef]
- Faulk, C.K.; Holt, G.J. Responses of Cobia Rachycentron Canadum Larvae to Abrupt or Gradual Changes in Salinity. Aquaculture 2006, 254, 275–283. [Google Scholar] [CrossRef]
- Romero, J.; Ringø, E.; Merrifield, D.L. The Gut Microbiota of Fish. In Aquaculture Nutrition: Gut Health, Probiotics and Prebiotics; Merrifield, D.L., Ringø, E., Eds.; Aquaculture Nutrition: West Sussex, UK, 2014; pp. 75–100. ISBN 9780470672716. [Google Scholar]
- Navarrete, P.; Tovar-Ramírez, D. Use of Yeasts as Probiotics in Fish Aquaculture. Sustain. Aquac. Tech. 2014, 1, 135–172. [Google Scholar] [CrossRef] [Green Version]
- Legrand, T.P.R.A.; Wynne, J.W.; Weyrich, L.S.; Oxley, A.P.A. A Microbial Sea of Possibilities: Current Knowledge and Prospects for an Improved Understanding of the Fish Microbiome. Rev. Aquac. 2020, 12, 1101–1134. [Google Scholar] [CrossRef]
- Siriyappagouder, P.; Kiron, V.; Lokesh, J.; Rajeish, M.; Kopp, M.; Fernandes, J. The Intestinal Mycobiota in Wild Zebrafish Comprises Mainly Dothideomycetes While Saccharomycetes Predominate in Their Laboratory-Reared Counterparts. Front. Microbiol. 2018, 9, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghori, I.; Tabassum, M.; Ahmad, T.; Zuberi, A.; Imra, M. Geotrichum Candidum Enhanced the Enterococcus Faecium Impact in Improving Physiology, and Health of Labeo Rohita (Hamilton, 1822) by Modulating Gut Microbiome Under Mimic Aquaculture Conditions. Turkish J. Fish. Aquat. Sci. 2018, 18, 1255–1267. [Google Scholar] [CrossRef]
- van Uden, N.; Kolipinski, M.C. Torulopsis Haemulonii Nov. Spec. a Yeast from the Atlantic Ocean. Antonie Van Leeuwenhoek 1962, 28, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Merrifield, D.L.; Dimitroglou, A.; Foey, A.; Davies, S.J.; Baker, R.T.M.; Bøgwald, J.; Castex, M.; Ringø, E. The Current Status and Future Focus of Probiotic and Prebiotic Applications for Salmonids. Aquaculture 2010, 302, 1–18. [Google Scholar] [CrossRef]
- Gómez-Pastor, R.; Pérez-Torrado, R.; Garre, E.; Matallana, E. Recent Advances in Yeast Biomass Production. In Biomass: Detection, Production and Usage; Matovic, D., Ed.; IntechOpen: Rijeka, Croatia, 2011; pp. 202–222. ISBN 9789533074924. [Google Scholar]
- Lapeña, D.; Kosa, G.; Hansen, L.D.; Mydland, L.T.; Passoth, V.; Horn, S.J.; Eijsink, V.G.H. Production and Characterization of Yeasts Grown on Media Composed of Spruce-Derived Sugars and Protein Hydrolysates from Chicken by-Products. Microb. Cell Fact. 2020, 19, 19. [Google Scholar] [CrossRef]
- Vandermies, M.; Fickers, P. Bioreactor-Scale Strategies for the Production of Recombinant Protein in the Yeast Yarrowia Lipolytica. Microorganisms 2019, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Balcázar, J.L.; Blas, I.d.; Ruiz-Zarzuela, I.; Cunningham, D.; Vendrell, D.; Múzquiz, J.L. The Role of Probiotics in Aquaculture. Vet. Microbiol. 2006, 114, 173–186. [Google Scholar] [CrossRef]
- Garcia-Martos, P.; Mira-Gutierrez, J. Contribution to the Knowledge of the Enzymatic Activity of Yeasts of Clinical Interest. Mycopathologia 1995, 132, 9–13. [Google Scholar] [CrossRef]
- Chan, M.Y.; Tay, S.T. Enzymatic Characterisation of Clinical Isolates of Cryptococcus neoformans, Cryptococcus gattii and Other Environmental Cryptococcus Spp. Mycoses 2010, 53, 26–31. [Google Scholar] [CrossRef]
- Mudryk, Z.J.; Podgórska, B. Enzymatic Activity of Bacterial Strains Isolated from Marine Beach Sediments. Polish J. Environ. Stud. 2006, 15, 441–448. [Google Scholar]
- Kim, S.; Baek, S.H.; Lee, K.; Hahn, J.S. Cellulosic Ethanol Production Using a Yeast Consortium Displaying a Minicellulosome and β-Glucosidase. Microb. Cell Fact. 2013, 12, 14. [Google Scholar] [CrossRef] [Green Version]
- Bates, J.M.; Akerlund, J.; Mittge, E.; Guillemin, K. Intestinal Alkaline Phosphatase Detoxifies Lipopolysaccharide and Prevents Inflammation in Zebrafish in Response to the Gut Microbiota. Cell Host Microbe 2007, 2, 371–382. [Google Scholar] [CrossRef] [Green Version]
- Kanpiengjai, A.; Khanongnuch, C.; Lumyong, S.; Kummasook, A.; Kittibunchakul, S. Characterization of Sporidiobolus Ruineniae A45.2 Cultivated in Tannin Substrate for Use as a Potential Multifunctional Probiotic Yeast in Aquaculture. J. Fungi 2020, 6, 378. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Shen, J.; Tian, D.; Yu, J.; He, T.; Yi, J.; Li, Y. A Potential Alternative to Traditional Antibiotics in Aquaculture: Yeast Glycoprotein Exhibits Antimicrobial Effect in Vivo and in Vitro on Aeromonas Caviae Isolated from Carassius Auratus Gibelio. Vet. Med. Sci. 2020, 6, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Zara, G.; Budroni, M.; Mannazzu, I.; Fancello, F.; Zara, S. Yeast Biofilm in Food Realms: Occurrence and Control. World J. Microbiol. Biotechnol. 2020, 36, 134. [Google Scholar] [CrossRef]
- Hatoum, R.; Labrie, S.; Fliss, I. Antimicrobial and Probiotic Properties of Yeasts: From Fundamental to Novel Applications. Front. Microbiol. 2012, 3, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Jeong, H.S.; Lee, H.Y.; Ahn, J. Assessment of Cell Surface Properties and Adhesion Potential of Selected Probiotic Strains. Lett. Appl. Microbiol. 2009, 49, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Del Re, B.; Sgorbati, B.; Miglioli, M.; Palenzona, D. Adhesion, Autoaggregation and Hydrophobicity of 13 Strains of Bifidobacterium Longum. Lett. Appl. Microbiol. 2000, 31, 438–442. [Google Scholar] [CrossRef]
- Masuoka, J.; Hazen, K.C. Cell Wall Protein Mannosylation Determines Candida Albicans Cell Surface Hydrophobicity. Microbiology 1997, 143 Pt 9, 3015–3021. [Google Scholar] [CrossRef] [Green Version]
- Lichtenberger, L.M. The Hydrophobic Barrier Properties of Gastrointestinal Mucus. Annu. Rev. Physiol. 1995, 57, 565–583. [Google Scholar] [CrossRef] [PubMed]
- Simões, L.A.; Cristina de Souza, A.; Ferreira, I.; Melo, D.S.; Lopes, L.A.A.; Magnani, M.; Schwan, R.F.; Dias, D.R. Probiotic Properties of Yeasts Isolated from Brazilian Fermented Table Olives. J. Appl. Microbiol. 2021, 131, 1983–1997. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, T.B.; Fink, G.R. Bakers’ Yeast, a Model for Fungal Biofilm Formation. Science 2001, 291, 878–881. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, K.J.; Klis, F.M. Flocculation, Adhesion and Biofilm Formation in Yeasts. Mol. Microbiol. 2006, 60, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Goktas, H.; Dikmen, H.; Demirbas, F.; Sagdic, O.; Dertli, E. Characterisation of Probiotic Properties of Yeast Strains Isolated from Kefir Samples. Int. J. Dairy Technol. 2021, 74, 715–722. [Google Scholar] [CrossRef]
- Tomičić, R.; Tomičić, Z.; Raspor, P. Influence of Culture Conditions on Co-Aggregation of Probiotic Yeast Saccharomyces Boulardii with Candida Spp. and Their Auto-Aggregation. Folia Microbiol. 2022, 67, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Schlomann, B.H.; Wiles, T.J.; Wall, E.S.; Guillemin, K.; Parthasarathy, R. Bacterial Cohesion Predicts Spatial Distribution in the Larval Zebrafish Intestine. Biophys. J. 2018, 115, 2271–2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardócz, S.; Duguid, T.J.; Brown, D.S.; Grant, G.; Pusztai, A.; White, A.; Ralph, A. The Importance of Dietary Polyamines in Cell Regeneration and Growth. Br. J. Nutr. 1995, 73, 819–828. [Google Scholar] [CrossRef] [Green Version]
- Bardocz, S. The Role of Dietary Polyamines. Eur. J. Clin. Nutr. 1993, 47, 683–690. [Google Scholar]
- Peulen, O.; Deloyer, P.; Grandfils, C.; Loret, S.; Dandrifosse, G. Intestinal Maturation Induced by Spermine in Young Animals. Livest. Prod. Sci. 2000, 66, 109–120. [Google Scholar] [CrossRef]
- Pères, A.; Cahu, C.L.; Zambonino-Infante, J.L. Dietary Spermine Supplementation Induces Intestinal Maturation in Sea Bass (Dicentrarchus Labrax) Larvae. Fish Physiol. Biochem. 1997, 16, 479–485. [Google Scholar] [CrossRef]
- Teles, A.; Alvarez-González, C.A.; Llera-Herrera, R.; Gisbert, E.; Salas-Leiva, J.; del Carmen Rodríguez-Jaramillo, M.; Tovar-Ramírez, D. Debaryomyces Hansenii CBS 8339 Promotes Larval Development in Seriola Rivoliana. Aquaculture 2022, 560, 738587. [Google Scholar] [CrossRef]
- Tovar-Ramírez, D.; Zambonino-Infante, J.L.; Cahu, C.L.; Gatesoupe, F.J.; Vázquez-Juárez, R. Efecto de La Administración de Levaduras En El Processo de Maduración Del Tracto Digestivo de Peces. In Avances en Nutrición Acuícola V, Memorias del V Simposium Internacional de Nutrición Acuícola; Cruz-Suárez, L.E., Ricque-Marie, D., Tapia-Salazar, M., Olvera Novoa, M.A., Civera-Cerecedo, R., Eds.; Universidad Autónoma de Nuevo León: Monterrey, México, 2000; pp. 33–46. [Google Scholar]
- Reyes-Becerril, M.; Tovar-Ramírez, D.; Ascencio-Valle, F.; Civera-Cerecedo, R.; Gracia-López, V.; Barbosa-Solomieu, V. Effects of Dietary Live Yeast Debaryomyces Hansenii on the Immune and Antioxidant System in Juvenile Leopard Grouper Mycteroperca Rosacea Exposed to Stress. Aquaculture 2008, 280, 39–44. [Google Scholar] [CrossRef]
- Jeong, J.W.; Cha, H.J.; Han, M.H.; Hwang, S.J.; Lee, D.S.; Yoo, J.S.; Choi, I.W.; Kim, S.; Kim, H.S.; Kim, G.Y.; et al. Spermidine Protects against Oxidative Stress in Inflammation Models Using Macrophages and Zebrafish. Biomol. Ther. 2018, 26, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swapna, A.; Philip, R.; Sajeevan, T.P.; Simi, J.; Bright, S. Differential Expression of Antimicrobial Peptides in Penaeus Monodon in Response to the Administration of Marine Yeasts/Β Glucans and White Spot Virus Challenge. Blue Biotechnol. J. 2012, 1, 69–90. [Google Scholar]
- Abadias, M.; Benabarre, A.; Teixidó, N.; Usall, J.; Vias, I. Effect of Freeze Drying and Protectants on Viability of the Biocontrol Yeast Candida Sake. Int. J. Food Microbiol. 2001, 65, 173–182. [Google Scholar] [CrossRef]
- Lodato, P.; Segovia de Huergo, M.; Buera, M.P. Viability and Thermal Stability of a Strain of Saccharomyces Cerevisiae Freeze-Dried in Different Sugar and Polymer Matrices. Appl. Microbiol. Biotechnol. 1999, 52, 215–220. [Google Scholar] [CrossRef]
- Cerrutti, P.; Segovia De Huergo, M.; Galvagno, M.; Schebor, C.; Del Pilar Buera, M. Commercial Baker’s Yeast Stability as Affected by Intracellular Content of Trehalose, Dehydration Procedure and the Physical Properties of External Matrices. Appl. Microbiol. Biotechnol. 2000, 54, 575–580. [Google Scholar] [CrossRef]
- Bond, C. Freeze-Drying of Yeast Cultures. Methods Mol. Biol. 2007, 368, 99–107. [Google Scholar] [CrossRef]
- Miyamoto-Shinohara, Y.; Imaizumi, T.; Sukenobe, J.; Murakami, Y.; Kawamura, S.; Komatsu, Y. Survival Rate of Microbes after Freeze-Drying and Long-Term Storage. Cryobiology 2000, 41, 251–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fish Characteristics | Characteristics of Yeast Isolates | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Origin | Weight (g) | Feed | Fish Code | N° | Yeast Isolate Code | Sequence Length (bp) | Closest Relatives | Identity (%) | Phylum | Species Assigned by Specific Primers | NCBI Accession Codes |
| E | 1447.8 | FFP | FC2 | 1 | C29 | 860 | Candida haemuloni JX459773.1 | 99.19 | A | Candida haemuloni OQ184038 | |
| 2 | C31 | 1059 | Candida parapsilosis MH545914.1 and MK394125.1 | 99.62 | A | Candida parapsilosis OQ184039 | |||||
| E | 1410.2 | FFP | FC4 | 3 | C47 | 874 | Candida haemuloni JX459773.1 | 99.31 | A | Candida haemuloni OQ184040 | |
| E | 1271.2 | FFP | FC6 | 4 | C32 | 1044 | Candida parapsilosis MH545914.1 and MK394125.1 | 99.52 | A | Candida parapsilosis OQ184041 | |
| 5 | C36 | 513 | Candida parapsilosis KX652404.1 | 99.22 | A | Candida parapsilosis OQ184042 | |||||
| 6 | C46 | 1058 | Candida parapsilosis MH545914.1 | 99.06 | A | Candida parapsilosis OQ184043 | |||||
| 7 | C50 | 1045 | Candida parapsilosis MH545914.1 and MK394125.1 | 97.63 | A | Candida parapsilosis OQ184044 | |||||
| E | 1087.6 | FFP | FC3 | 8 | C62 | 1073 | Debaryomyces fabryi MK394103.1, and Debaryomyces hansenii KX981201.1 | 99.24 | A | D. hansenii | Debaryomyces sp. OQ184045 |
| E | 871.8 | FFP | FC5 | 9 | C27 | 861 | Candida haemuloni JX459773.1 | 99.42 | A | Candida haemuloni OQ184046 | |
| 10 | C24 | 1175 | Debaryomyces hansenii FR686594.1 | 98.98 | A | D. hansenii | Debaryomyces sp. OQ184047 | ||||
| 11 | C28 | 1130 | Debaryomyces hansenii FR686594.1 and GQ458025.1 | 98.41 | A | D. hansenii | Debaryomyces sp. OQ184048 | ||||
| E | 801.8 | FFP | FC1 | 12 | C37 | 878 | Candida haemuloni JX459773.1 | 99.43 | A | Candida haemuloni OQ184049 | |
| 13 | C33 | 993 | Candida parapsilosis MZ375521.1 and MZ375508.1 | 97.79 | A | Candida parapsilosis OQ184050 | |||||
| 14 | C25 | 1154 | Debaryomyces hansenii FR686594.1 and GQ458025.1 | 98.70 | A | D. hansenii | Debaryomyces sp. OQ184051 | ||||
| 15 | C30 | 1149 | Debaryomyces hansenii FR686594.1 and GQ458025.1 | 97.05 | A | D. hansenii | Debaryomyces sp. OQ184052 | ||||
| 16 | C38 | 1091 | Debaryomyces fabryi MK394103.1, and Debaryomyces hansenii KC111444.1 | 99.91 | A | D. hansenii | Debaryomyces sp. OQ184053 | ||||
| 17 | C26 | 1175 | Debaryomyces hansenii FR686594.1 | 98.56 | A | Debaryomyces sp. OQ184054 | |||||
| 18 | C67 | 1156 | Debaryomyces hansenii FR686594.1 and GQ458025.1 | 98.45 | A | Debaryomyces sp. OQ184055 | |||||
| 19 | C61 | 978 | Naganishia adeliensis KY218697.1, and Naganishia sp. LC529181.1 | 97.75 | B | Naganishia sp. OQ184056 | |||||
| C | 5200.0 | FFP | FC32 | 20 | C13 | 842 | Candida haemuloni JX459773.1 | 99.17 | A | Candida haemuloni OQ184057 | |
| 21 | C16 | 1157 | Debaryomyces hansenii FR686594.1 and GQ458025.1 | 97.84 | A | D. hansenii | Debaryomyces sp. OQ184058 | ||||
| 22 | C17 | 858 | Debaryomyces fabryi MK394103.1, and Debaryomyces hansenii KC111444.1 | 98.26 | A | D. hansenii | Debaryomyces sp. OQ184059 | ||||
| C | 4780.0 | FFP | FC34 | 23 | C01 | 674 | Candida haemuloni OW987803.1 | 94.51 | A | Candida haemuloni OQ184060 | |
| C | 511.2 | FFP | FC20 | 24 | C03 | 1154 | Debaryomyces hansenii FR686594.1 and GQ458025.1 | 99.39 | A | D. hansenii | Debaryomyces sp. OQ184061 |
| 25 | C40 | 880 | Debaryomyces fabryi MK394103.1, and Debaryomyces hansenii KX981201.1 | 98.86 | A | D. hansenii | Debaryomyces sp. OQ184062 | ||||
| 26 | C41 | 1026 | Debaryomyces fabryi MK394103.1, Debaryomyces hansenii KX981201.1, and Debaryomyces sp. MW959732.1 | 99.81 | A | D. hansenii | Debaryomyces sp. OQ184063 | ||||
| 27 | C04 | 1094 | Debaryomyces fabryi MK394103.1, and Debaryomyces hansenii KX981201.1 | 99.91 | A | Debaryomyces sp. OQ184064 | |||||
| C | 498.6 | FFP | FC23 | 28 | C44 | 847 | Candida haemuloni JX459773.1 | 99.41 | A | Candida haemuloni OQ184065 | |
| 29 | C45 | 863 | Candida haemuloni JX459773.1 | 99.42 | A | Candida haemuloni OQ184066 | |||||
| 30 | C10 | 1159 | Debaryomyces hansenii FR686594.1 and GQ458025.1 | 99.13 | A | D. hansenii | Debaryomyces sp. OQ184067 | ||||
| C | 489.2 | FFP | FC24 | 31 | C21 | 1022 | Debaryomyces fabryi MK394103.1, and Debaryomyces hansenii KX981201.1 | 97.36 | A | D. hansenii | Debaryomyces sp. OQ184068 |
| C | 410.4 | FFP | FC25 | 32 | C43 | 1151 | Debaryomyces hansenii FR686594.1 and GQ458025.1 | 97.93 | A | D. hansenii | Debaryomyces sp. OQ184069 |
| 33 | C65 | 1141 | Debaryomyces hansenii FR686594.1 and GQ458025.1 | 98.87 | A | D. hansenii | Debaryomyces sp. OQ184070 | ||||
| C | 370.6 | FF | FC28 | 34 | C19 | 1152 | Debaryomyces hansenii FR686594.1 and GQ458025.1 | 98.35 | A | D. hansenii | Debaryomyces sp. OQ184071 |
| C | 366.8 | FF | FC27 | 35 | C22 | 853 | Candida haemuloni JX459773.1 | 98.59 | A | Candida haemuloni OQ184072 | |
| 36 | C42 | 365 | Candida haemuloni MH667577.1 and MH636864.1 | 100.00 | A | Candida haemuloni OQ184073 | |||||
| 37 | C12 | 1160 | Debaryomyces hansenii HE799666.1 and HE799660.1 | 97.76 | A | D. hansenii | Debaryomyces sp. OQ184074 | ||||
| 38 | C60 | 1190 | Debaryomyces hansenii FR686594.1 | 96.83 | A | D. hansenii | Debaryomyces sp. OQ184075 | ||||
| C | 327.6 | FF | FC30 | 39 | C49 | 1154 | Debaryomyces hansenii FR686594.1 and GQ458025.1; and Debaryomyces sp. MW959725.1 | 98.43 | A | D. hansenii | Debaryomyces sp. OQ184076 |
| N° | Yeast Isolate Code | Species | Dry Weight (g L−1) | RAPD-PCR Pattern | Selected Yeast Strains |
|---|---|---|---|---|---|
| 1 | C47 | Candida haemuloni | 0.69 ± 0.13 | A | ✓ |
| 2 | C45 | Candida haemuloni | 0.57 ± 0.07 | A | |
| 3 | C44 | Candida haemuloni | 2.14 ± 0.17 | B | |
| 4 | C01 | Candida haemuloni | 2.00 ± 0.07 | B | ✓ |
| 5 | C13 | Candida haemuloni | 1.94 ± 0.05 | B | |
| 6 | C29 | Candida haemuloni | 1.89 ± 0.06 | B | |
| 7 | C37 | Candida haemuloni | 1.86 ± 0.08 | B | |
| 8 | C22 | Candida haemuloni | 1.84 ± 0.10 | B | |
| 9 | C42 | Candida haemuloni | 1.65 ± 0.03 | B | |
| 10 | C27 | Candida haemuloni | 1.72 ± 0.01 | C | ✓ |
| 11 | C33 | Candida parapsilosis | 0.83 ± 0.09 | D | ✓ |
| 12 | C46 | Candida parapsilosis | 1.57 ± 0.02 | E | ✓ |
| 13 | C50 | Candida parapsilosis | 1.53 ± 0.04 | E | |
| 14 | C31 | Candida parapsilosis | 1.50 ± 0.04 | F | ✓ |
| 15 | C32 | Candida parapsilosis | 1.36 ± 0.07 | G | ✓ |
| 16 | C36 | Candida parapsilosis | 1.59 ± 0.04 | H | ✓ |
| 17 | C28 | Debaryomyces hansenii | 1.05 ± 0.06 | I | ✓ |
| 18 | C40 | Debaryomyces hansenii | 0.93 ± 0.09 | J | ✓ |
| 19 | C19 | Debaryomyces hansenii | 0.88 ± 0.06 | J | |
| 20 | C03 | Debaryomyces hansenii | 1.00 ± 0.15 | K | ✓ |
| 21 | C21 | Debaryomyces hansenii | 0.86 ± 0.08 | K | |
| 22 | C49 | Debaryomyces hansenii | 0.78 ± 0.07 | K | |
| 23 | C62 | Debaryomyces hansenii | 0.07 ± 0.01 | K | |
| 24 | C38 | Debaryomyces hansenii | 0.98 ± 0.13 | L | |
| 25 | C17 | Debaryomyces hansenii | 0.89 ± 0.04 | L | ✓ |
| 26 | C24 | Debaryomyces hansenii | 0.88 ± 0.05 | L | |
| 27 | C30 | Debaryomyces hansenii | 0.87 ± 0.08 | L | |
| 28 | C25 | Debaryomyces hansenii | 0.86 ± 0.02 | L | |
| 29 | C16 | Debaryomyces hansenii | 0.83 ± 0.08 | L | |
| 30 | C41 | Debaryomyces hansenii | 0.70 ± 0.12 | L | |
| 31 | C10 | Debaryomyces hansenii | 1.02 ± 0.08 | M | ✓ |
| 32 | C65 | Debaryomyces hansenii | 0.95 ± 0.06 | M | |
| 33 | C12 | Debaryomyces hansenii | 0.94 ± 0.09 | M | |
| 34 | C43 | Debaryomyces hansenii | 0.93 ± 0.04 | M | |
| 35 | C60 | Debaryomyces hansenii | 0.91 ± 0.07 | M | |
| 36 | C26 | Debaryomyces sp. | 0.87 ± 0.06 | N | ✓ |
| 37 | C04 | Debaryomyces sp. | 0.50 ± 0.04 | N | |
| 38 | C67 | Debaryomyces sp. | 0.30 ± 0.07 | O | ✓ |
| 39 | C61 | Naganishia sp. | 0.66 ± 0.02 | P | ✓ |
| N° | Enzymes | Candida haemuloni | Candida parapsilosis | Debaryomyces hansenii | Debaryomyces sp. | Naganishia sp. | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C01 | C27 | C47 | C31 | C32 | C33 | C36 | C46 | C03 | C10 | C17 | C28 | C40 | C26 | C67 | C61 | ||
| 1 | Alkaline phosphatase | - | - | - | ± | ± | - | ± | ± | ± | ± | ± | ± | - | - | - | - |
| 2 | Esterase (C 4) | + | ± | ± | ± | ± | ± | ± | + | ± | ± | ± | ± | ± | ± | ± | ± |
| 3 | Esterase Lipase (C 8) | + | ± | ± | ± | ± | ± | ± | ± | ± | ± | ± | ± | ± | ± | ± | ± |
| 4 | Lipase (C 14) | - | ± | - | - | - | - | - | - | - | ± | - | ± | - | - | - | - |
| 5 | Leucine arylamidase | + | + | + | + | + | + | + | + | ± | ± | + | + | + | + | + | + |
| 6 | Valine arylamidase | + | + | ± | + | + | ± | + | + | ± | ± | ± | ± | ± | ± | ± | ± |
| 7 | Cystine arylamidase | ± | ± | - | ± | ± | - | - | ± | - | - | - | - | - | - | - | - |
| 8 | Phosphatase acid | - | ± | + | + | + | ± | + | + | - | ± | ± | ± | ± | ± | ± | + |
| 9 | Naphthol-AS-BI-phosphohydrolase | - | ± | ± | ± | + | ± | + | + | - | ± | ± | ± | ± | ± | ± | ± |
| 10 | α glucosidase | - | - | - | ± | ± | ± | ± | + | - | ± | - | - | - | - | - | - |
| 11 | β glucosidase | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Total enzymes | 5 | 8 | 7 | 9 | 9 | 7 | 8 | 9 | 5 | 9 | 7 | 8 | 6 | 6 | 6 | 7 | |
| Yeast strains | Safety | Action against Pathogens | Potential to Adhere to Intestinal Mucosa | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Biomass Production | Enzymatic Activity | Hemolysis of Cobia Erythrocytes | Antagonism against Vibrio spp. | Biofilm | Autoaggregation | Hydrophobicity | Total | Selected Yeast Strains | |
| Ch−C01 | + | - | - | - | + | - | + | 3 | ✓ |
| Ch−C27 | + | + | - | - | + | - | + | 4 | ✓ |
| Ch−C47 | - | - | - | - | - | + | + | 2 | |
| Cp−C31 | + | + | - | - | + | + | + | 5 | ✓ |
| Cp−C32 | + | + | - | - | + | - | + | 4 | ✓ |
| Cp−C33 | - | - | - | - | + | + | + | 3 | |
| Cp−C36 | + | + | - | - | + | + | - | 4 | |
| Cp−C46 | + | + | - | - | + | + | - | 4 | ✓ |
| Dh−C03 | + | - | - | - | - | - | + | 2 | |
| Dh−C10 | + | + | - | - | - | + | + | 4 | ✓ |
| Dh−C17 | - | - | - | - | - | - | + | 1 | |
| Dh−C28 | + | + | - | - | - | - | + | 3 | ✓ |
| Dh−C40 | - | - | - | - | - | + | + | 2 | |
| Dsp−C26 | - | - | - | - | - | - | + | 1 | |
| Dsp−C67 | - | - | - | - | + | - | + | 2 | |
| Nsp−C61 | - | - | - | - | - | + | + | 2 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reinoso, S.; Gutiérrez, M.S.; Domínguez-Borbor, C.; Argüello-Guevara, W.; Bohórquez-Cruz, M.; Sonnenholzner, S.; Nova-Baza, D.; Mardones, C.; Navarrete, P. Selection of Autochthonous Yeasts Isolated from the Intestinal Tracts of Cobia Fish (Rachycentron canadum) with Probiotic Potential. J. Fungi 2023, 9, 274. https://doi.org/10.3390/jof9020274
Reinoso S, Gutiérrez MS, Domínguez-Borbor C, Argüello-Guevara W, Bohórquez-Cruz M, Sonnenholzner S, Nova-Baza D, Mardones C, Navarrete P. Selection of Autochthonous Yeasts Isolated from the Intestinal Tracts of Cobia Fish (Rachycentron canadum) with Probiotic Potential. Journal of Fungi. 2023; 9(2):274. https://doi.org/10.3390/jof9020274
Chicago/Turabian StyleReinoso, Samira, María Soledad Gutiérrez, Cristóbal Domínguez-Borbor, Wilfrido Argüello-Guevara, Milton Bohórquez-Cruz, Stanislaus Sonnenholzner, Daniela Nova-Baza, Claudia Mardones, and Paola Navarrete. 2023. "Selection of Autochthonous Yeasts Isolated from the Intestinal Tracts of Cobia Fish (Rachycentron canadum) with Probiotic Potential" Journal of Fungi 9, no. 2: 274. https://doi.org/10.3390/jof9020274
APA StyleReinoso, S., Gutiérrez, M. S., Domínguez-Borbor, C., Argüello-Guevara, W., Bohórquez-Cruz, M., Sonnenholzner, S., Nova-Baza, D., Mardones, C., & Navarrete, P. (2023). Selection of Autochthonous Yeasts Isolated from the Intestinal Tracts of Cobia Fish (Rachycentron canadum) with Probiotic Potential. Journal of Fungi, 9(2), 274. https://doi.org/10.3390/jof9020274