
The Antidepressant Sertraline Affects Cell Signaling and Metabolism in Trichophyton rubrum

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Culture Conditions for RNA-Seq Analysis
2.2. RNA Isolation and cDNA Library Construction
2.3. Transcriptome Analysis
2.4. Data Analysis and Gene Selection
2.5. cDNA Synthesis and RT-qPCR Analysis
2.6. Quantitation of Ergosterol Content
3. Results
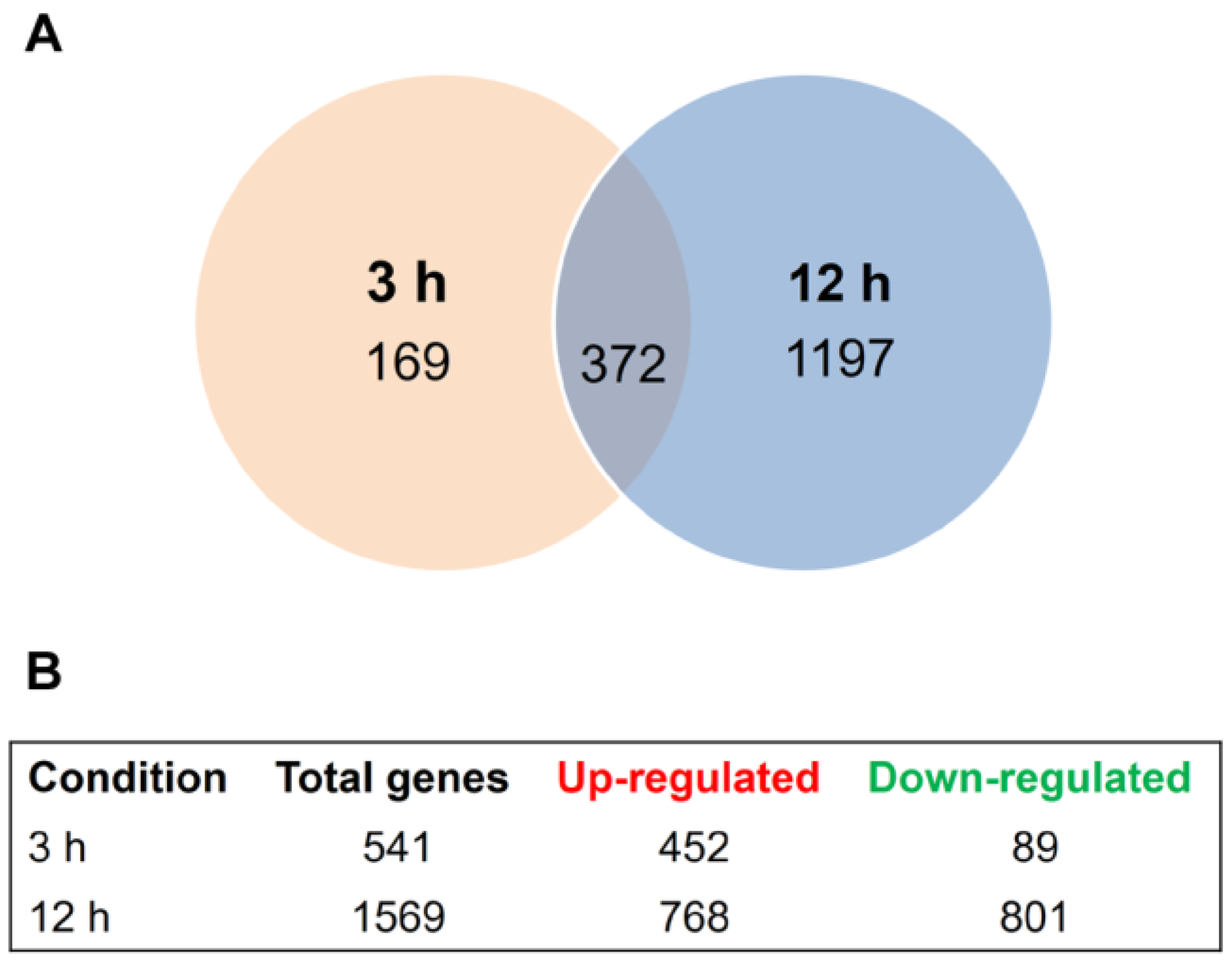
3.1. Transcriptional Profiling of Trichophyton rubrum Challenged by SRT
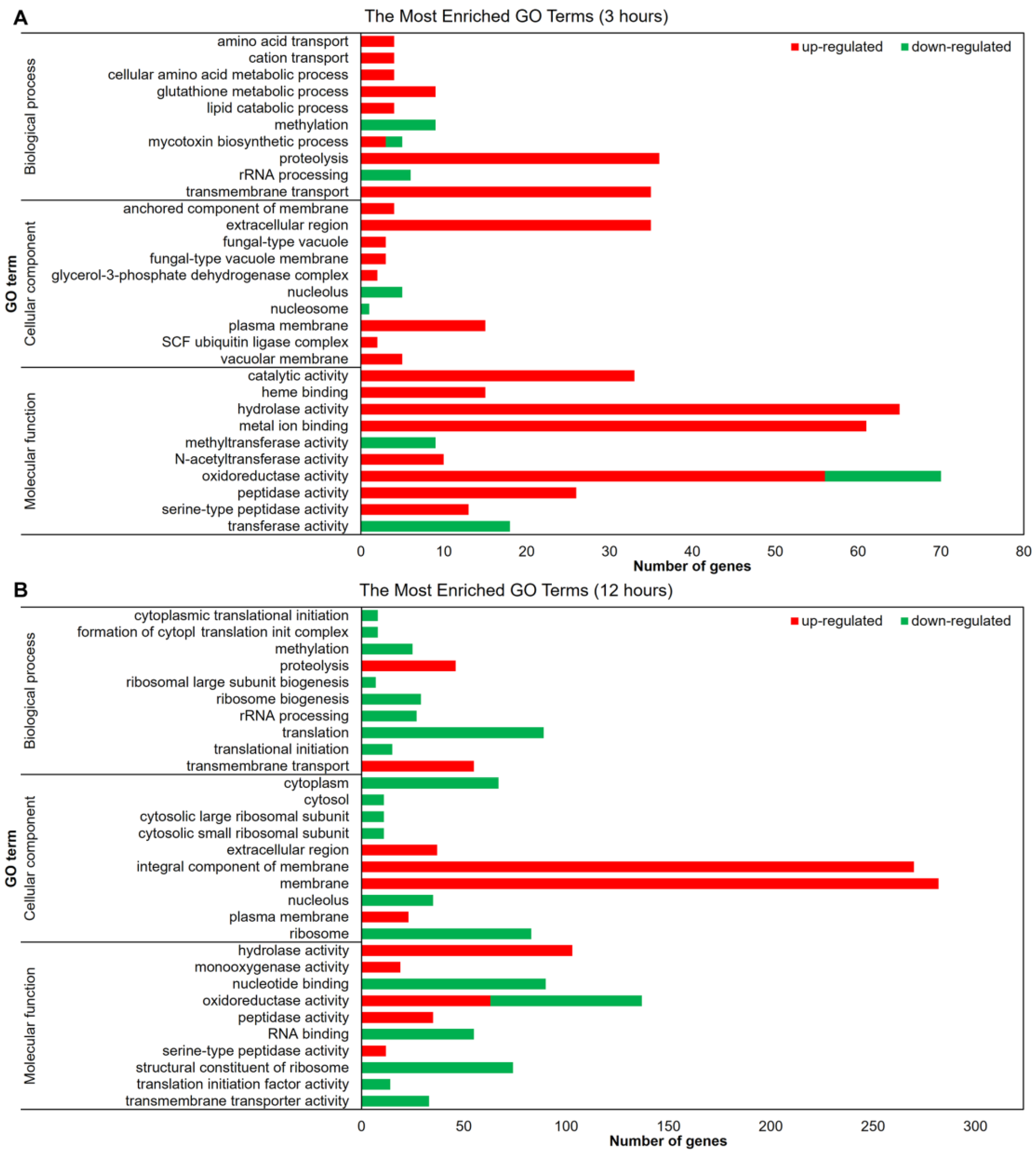
3.2. SRT Interferes with T. rubrum Signaling and Regulation
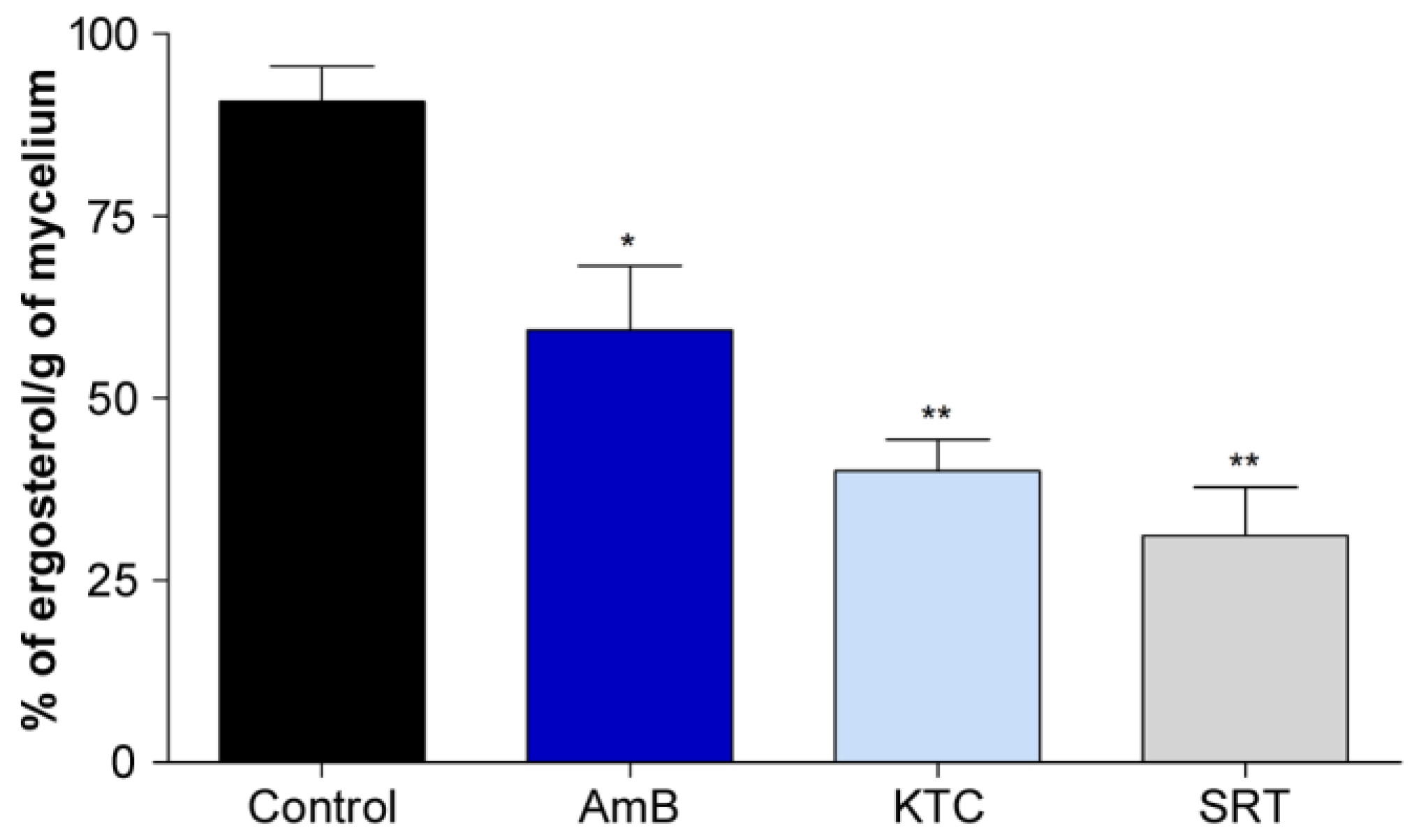
3.3. Cell Wall and Membrane Structure Are Affected by SRT
3.4. SRT Affects the Transcription of Oxidative Stress Genes in T. rubrum
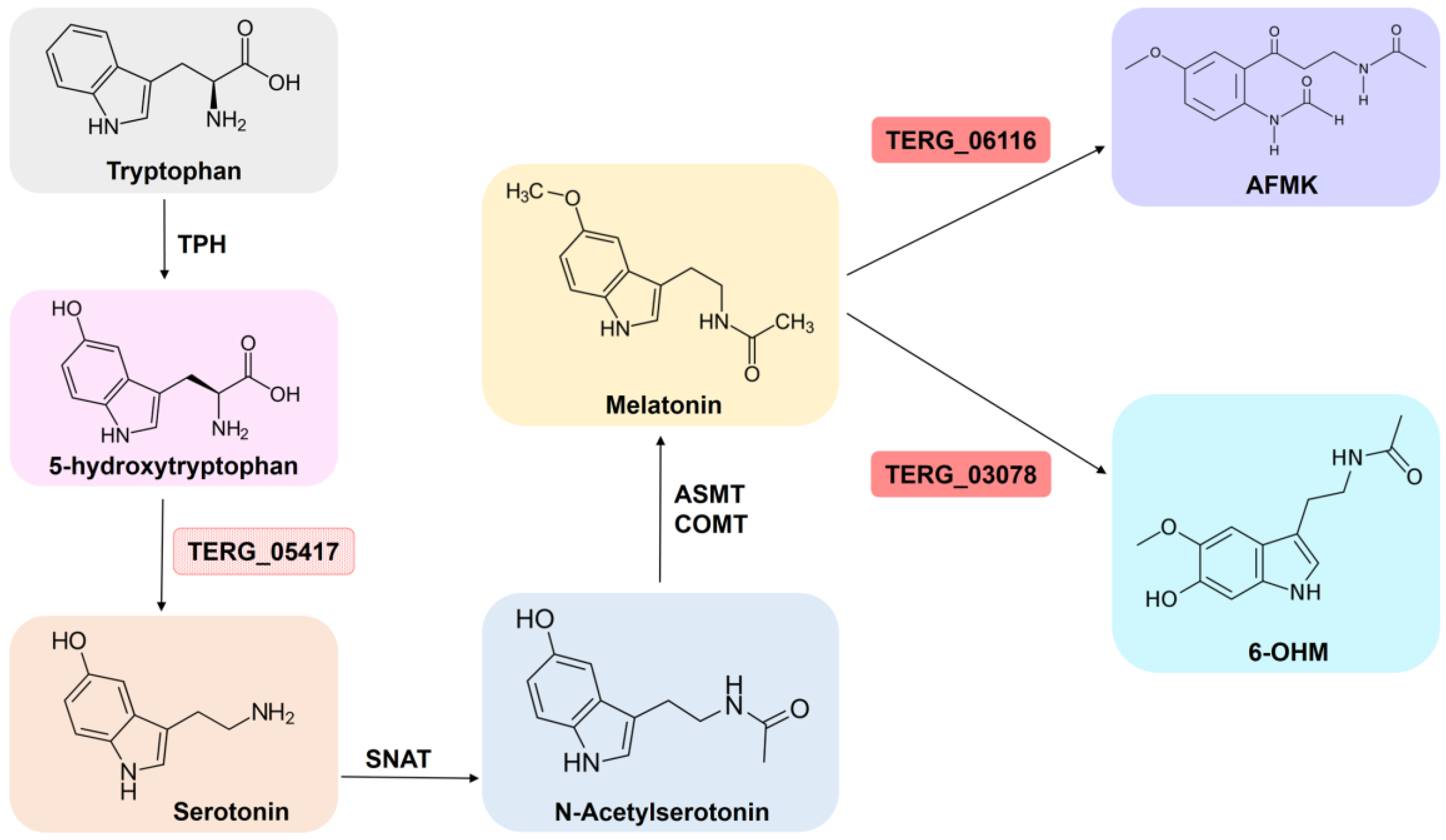
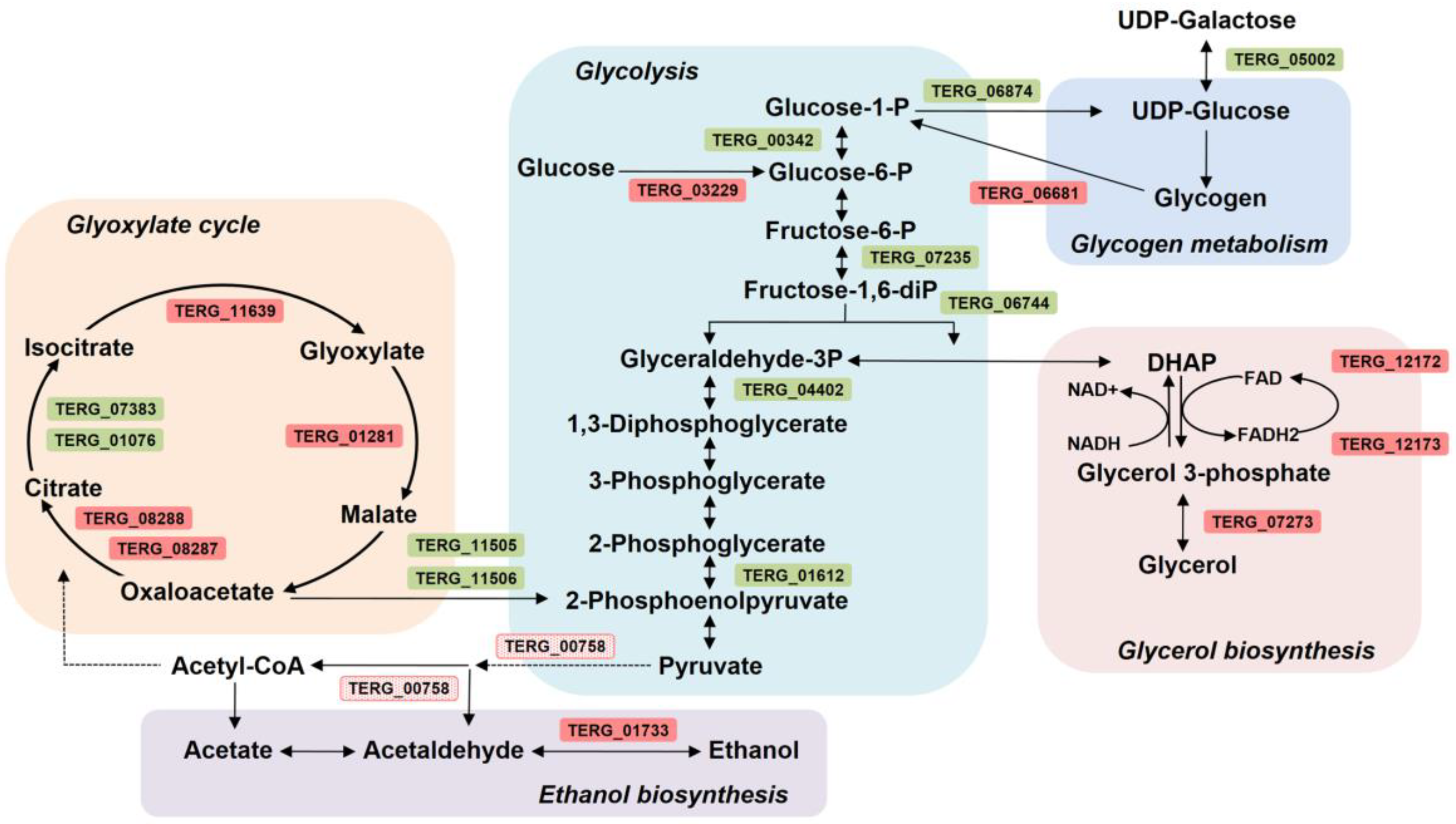
3.5. Effect of SRT on T. rubrum Metabolism
4. Discussion
4.1. SRT Affects Cell Signaling and Transcription in T. rubrum
4.2. SRT Disturbs T. rubrum Cell Wall and Membrane
4.3. SRT Overbalances Oxidative Stress Response
4.4. SRT Imposes Dynamic Metabolic Modulation
5. Concluding Remarks, Challenges, and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rouzaud, C.; Hay, R.; Chosidow, O.; Dupin, N.; Puel, A.; Lortholary, O.; Lanternier, F. Severe Dermatophytosis and Acquired or Innate Immunodeficiency: A Review. J. Fungi 2015, 2, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piraccini, B.M.; Alessandrini, A. Onychomycosis: A Review. J. Fungi 2015, 1, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Rossi, N.M.; Peres, N.T.; Bitencourt, T.A.; Martins, M.P.; Rossi, A. State-of-the-art Dermatophyte infections: Epidemiology aspects, pathophysiology, and resistance mechanisms. J. Fungi 2021, 7, 629. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Rossi, N.M.; Bitencourt, T.A.; Peres, N.T.A.; Lang, E.A.S.; Gomes, E.V.; Quaresemin, N.R.; Martins, M.P.; Lopes, L.; Rossi, A. Dermatophyte Resistance to Antifungal Drugs: Mechanisms and Prospectus. Front. Microbiol. 2018, 9, 1108. [Google Scholar] [CrossRef] [Green Version]
- Rossi, A.; Martins, M.r.P.; Bitencourt, T.A.; Peres, N.T.A.; Rocha, C.H.L.; Rocha, F.M.G.; Neves-da-Rocha, J.o.; Lopes, M.E.R.; Sanches, P.R.; Bortolossi, J.C. Reassessing the use of undecanoic acid as a therapeutic strategy for treating fungal infections. Mycopathologia 2021, 186, 327–340. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, F.; Zeng, M.; Mao, Y.; Song, Z. Drug repurposing strategies in the development of potential antifungal agents. Appl. Microbiol. Biotechnol. 2021, 105, 5259–5279. [Google Scholar] [CrossRef]
- Lass-Flörl, C.; Dierich, M.; Fuchs, D.; Semenitz, E.; Jenewein, I.; Ledochowski, M. Antifungal properties of selective serotonin reuptake inhibitors against Aspergillus species in vitro. J. Antimicrob. Chemother. 2001, 48, 775–779. [Google Scholar] [CrossRef] [Green Version]
- Ayaz, M.; Subhan, F.; Ahmed, J.; Khan, A.U.; Ullah, F.; Ullah, I.; Ali, G.; Syed, N.I.; Hussain, S. Sertraline enhances the activity of antimicrobial agents against pathogens of clinical relevance. J. Biol. Res. 2015, 22, 4. [Google Scholar] [CrossRef] [Green Version]
- Villanueva-Lozano, H.; Treviño-Rangel, R.J.; Téllez-Marroquín, R.; Bonifaz, A.; Rojas, O.C.; Hernández-Rodríguez, P.A.; González, G.M. In vitro inhibitory activity of sertraline against clinical isolates of Sporothrix schenckii. Rev. Iberoam. Micol. 2019, 36, 139–141. [Google Scholar] [CrossRef]
- Breuer, M.R.; Dasgupta, A.; Vasselli, J.G.; Lin, X.; Shaw, B.D.; Sachs, M.S. The Antidepressant Sertraline Induces the Formation of Supersized Lipid Droplets in the Human Pathogen Cryptococcus neoformans. J. Fungi 2022, 8, 642. [Google Scholar] [CrossRef]
- Zhai, B.; Wu, C.; Wang, L.; Sachs, M.S.; Lin, X. The antidepressant sertraline provides a promising therapeutic option for neurotropic cryptococcal infections. Antimicrob. Agents Chemother. 2012, 56, 3758–3766. [Google Scholar] [CrossRef] [Green Version]
- Cong, L.; Liao, Y.; Yang, S.; Yang, R. In vitro antifungal activity of sertraline and synergistic effects in combination with antifungal drugs against planktonic forms and biofilms of clinical Trichosporon asahii isolates. PLoS ONE 2016, 11, e0167903. [Google Scholar] [CrossRef] [Green Version]
- Rossato, L.; Loreto, É.S.; Zanette, R.A.; Chassot, F.; Santurio, J.M.; Alves, S.H. In vitro synergistic effects of chlorpromazine and sertraline in combination with amphotericin B against Cryptococcus neoformans var. grubii. Folia Microbiol. 2016, 61, 399–403. [Google Scholar] [CrossRef]
- Villanueva-Lozano, H.; González, G.M.; Espinosa-Mora, J.E.; Bodden-Mendoza, B.A.; Andrade, A.; Martínez-Reséndez, M.F.; Treviño-Rangel, R.J. Evaluation of the expanding spectrum of sertraline against uncommon fungal pathogens. J. Infect. Chemother. 2020, 26, 309–311. [Google Scholar] [CrossRef]
- Rocha, C.H.L.; Rocha, F.M.G.; Bitencourt, T.A.; Martins, M.P.; Sanches, P.R.; Rossi, A.; Martinez-Rossi, N.M. Synergism between the Antidepressant Sertraline and Caspofungin as an Approach to Minimise the Virulence and Resistance in the Dermatophyte Trichophyton rubrum. J. Fungi 2022, 8, 815. [Google Scholar] [CrossRef]
- Sangkuhl, K.; Klein, T.E.; Altman, R.B. Selective serotonin reuptake inhibitors pathway. Pharm. Genom. 2009, 19, 907–909. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Park, S.Y.; Hwang, O. Up-regulation of tryptophan hydroxylase expression and serotonin synthesis by sertraline. Mol. Pharmacol. 2002, 61, 778–785. [Google Scholar] [CrossRef] [Green Version]
- Rainey, M.M.; Korostyshevsky, D.; Lee, S.; Perlstein, E.O. The antidepressant sertraline targets intracellular vesiculogenic membranes in yeast. Genetics 2010, 185, 1221–1233. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Persinoti, G.F.; de Aguiar Peres, N.T.; Jacob, T.R.; Rossi, A.; Vêncio, R.Z.; Martinez-Rossi, N.M. RNA-sequencing analysis of Trichophyton rubrum transcriptome in response to sublethal doses of acriflavine. BMC Genom. 2014, 15 (Suppl 7), S1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vencio, R.Z.; Koide, T.; Gomes, S.L.; Pereira, C.A. BayGO: Bayesian analysis of ontology term enrichment in microarray data. BMC Bioinform. 2006, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [Green Version]
- Jacob, T.R.; Peres, N.T.A.; Persinoti, G.F.; Silva, L.G.; Mazucato, M.; Rossi, A.; Martinez-Rossi, N.M. rpb2 is a reliable reference gene for quantitative gene expression analysis in the dermatophyte Trichophyton rubrum. Med. Mycol. 2012, 50, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Arthington-Skaggs, B.A.; Jradi, H.; Desai, T.; Morrison, C.J. Quantitation of ergosterol content: Novel method for determination of fluconazole susceptibility of Candida albicans. J. Clin. Microbiol. 1999, 37, 3332–3337. [Google Scholar] [CrossRef] [Green Version]
- Mendes, N.S.; Bitencourt, T.A.; Sanches, P.R.; Silva-Rocha, R.; Martinez-Rossi, N.M.; Rossi, A. Transcriptome-wide survey of gene expression changes and alternative splicing in Trichophyton rubrum in response to undecanoic acid. Sci. Rep. 2018, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Martinez, D.A.; Oliver, B.G.; Graser, Y.; Goldberg, J.M.; Li, W.; Martinez-Rossi, N.M.; Monod, M.; Shelest, E.; Barton, R.C.; Birch, E.; et al. Comparative Genome Analysis of Trichophyton rubrum and Related Dermatophytes Reveals Candidate Genes Involved in Infection. MBio 2012, 3, e00259-12. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Fu, X.D. Regulation of splicing by SR proteins and SR protein-specific kinases. Chromosoma 2013, 122, 191–207. [Google Scholar] [CrossRef] [Green Version]
- Bullock, A.N.; Das, S.; Debreczeni, J.E.; Rellos, P.; Fedorov, O.; Niesen, F.H.; Guo, K.; Papagrigoriou, E.; Amos, A.L.; Cho, S.; et al. Kinase domain insertions define distinct roles of CLK kinases in SR protein phosphorylation. Structure 2009, 17, 352–362. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.D.; Strange, R.C. Glutathione S-transferase polymorphisms and their biological consequences. Pharmacology 2000, 61, 154–166. [Google Scholar] [CrossRef]
- Muñiz-Calvo, S.; Bisquert, R.; Fernández-Cruz, E.; García-Parrilla, M.C.; Guillamón, J.M. Deciphering the melatonin metabolism in Saccharomyces cerevisiae by the bioconversion of related metabolites. J. Pineal Res. 2019, 66, e12554. [Google Scholar] [CrossRef] [Green Version]
- Gallardo-Fernández, M.; Valls-Fonayet, J.; Valero, E.; Hornedo-Ortega, R.; Richard, T.; Troncoso, A.M.; Garcia-Parrilla, M.C. Isotopic labelling-based analysis elucidates biosynthesis pathways in Saccharomyces cerevisiae for Melatonin, Serotonin and Hydroxytyrosol formation. Food Chem. 2022, 374, 131742. [Google Scholar] [CrossRef]
- Muthu, D.; Gowri, M.; Suresh Kumar, G.; Kattimani, V.S.; Girija, E.K. Repurposing of antidepression drug sertraline for antimicrobial activity against Staphylococcus aureus: A potential approach for the treatment of osteomyelitis. New J. Chem. 2019, 43, 5315–5324. [Google Scholar] [CrossRef]
- Baú-Carneiro, J.L.; Akemi Guirao Sumida, I.; Gallon, M.; Zaleski, T.; Boia-Ferreira, M.; Bridi Cavassin, F. Sertraline repositioning: An overview of its potential use as a chemotherapeutic agent after four decades of tumor reversal studies. Transl. Oncol. 2022, 16, 101303. [Google Scholar] [CrossRef]
- Ferreira, D.D.; Mesquita, J.T.; da Costa Silva, T.A.; Romanelli, M.M.; da Gama Jaen Batista, D.; da Silva, C.F.; da Gama, A.N.S.; Neves, B.J.; Melo-Filho, C.C.; Correia Soeiro, M.N.; et al. Efficacy of sertraline against Trypanosoma cruzi: An in vitro and in silico study. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 30. [Google Scholar] [CrossRef]
- Krzyżek, P.; Franiczek, R.; Krzyżanowska, B.; Laczmański, L.; Migdal, P.; Gościniak, G. In Vitro Activity of Sertraline, an Antidepressant, Against Antibiotic-Susceptible and Antibiotic-Resistant Helicobacter pylori Strains. Pathogens 2019, 8, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, N.A.; Cartwright, T.N.; Lawless, C.; Cámara-Donoso, M.; Sen, O.; Sako, K.; Hirota, T.; Kimura, H.; Higgins, J.M.G. Kinase inhibition profiles as a tool to identify kinases for specific phosphorylation sites. Nat. Commun. 2020, 11, 1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piazzi, M.; Bavelloni, A.; Gallo, A.; Faenza, I.; Blalock, W.L. Signal Transduction in Ribosome Biogenesis: A Recipe to Avoid Disaster. Int. J. Mol. Sci. 2019, 20, 2718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelest, E. Transcription factors in fungi. FEMS Microbiol. Lett. 2008, 286, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Beauvais, A.; Latgé, J.-P. Special Issue: Fungal Cell Wall. J. Fungi 2018, 4, 91. [Google Scholar] [CrossRef] [Green Version]
- Martins, M.P.; Silva, L.G.; Rossi, A.; Sanches, P.R.; Souza, L.D.R.; Martinez-Rossi, N.M. Global Analysis of Cell Wall Genes Revealed Putative Virulence Factors in the Dermatophyte Trichophyton rubrum. Front. Microbiol. 2019, 10, 2168. [Google Scholar] [CrossRef] [Green Version]
- Ibe, C.; Munro, C.A. Fungal cell wall: An underexploited target for antifungal therapies. PLoS Pathog. 2021, 17, e1009470. [Google Scholar] [CrossRef]
- Roncero, C.; Vázquez de Aldana, C.R. Glucanases and Chitinases. Curr. Top. Microbiol. Immunol. 2020, 425, 131–166. [Google Scholar]
- Yamamoto, T.; Umegawa, Y.; Tsuchikawa, H.; Hanashima, S.; Matsumori, N.; Funahashi, K.; Seo, S.; Shinoda, W.; Murata, M. The Amphotericin B-Ergosterol Complex Spans a Lipid Bilayer as a Single-Length Assembly. Biochemistry 2019, 58, 5188–5196. [Google Scholar] [CrossRef]
- Abe, F.; Usui, K.; Hiraki, T. Fluconazole modulates membrane rigidity, heterogeneity, and water penetration into the plasma membrane in Saccharomyces cerevisiae. Biochemistry 2009, 48, 8494–8504. [Google Scholar] [CrossRef]
- Hinkovska-Galcheva, V.; Treadwell, T.; Shillingford, J.M.; Lee, A.; Abe, A.; Tesmer, J.J.G.; Shayman, J.A. Inhibition of lysosomal phospholipase A2 predicts drug-induced phospholipidosis. J. Lipid Res. 2021, 62, 100089. [Google Scholar] [CrossRef]
- Rhein, C.; Reichel, M.; Kramer, M.; Rotter, A.; Lenz, B.; Mühle, C.; Gulbins, E.; Kornhuber, J. Alternative splicing of SMPD1 coding for acid sphingomyelinase in major depression. J. Affect. Disord. 2017, 209, 10–15. [Google Scholar] [CrossRef]
- Ramrakhiani, L.; Chand, S. Recent progress on phospholipases: Different sources, assay methods, industrial potential and pathogenicity. Appl. Biochem. Biotechnol. 2011, 164, 991–1022. [Google Scholar] [CrossRef]
- O'Luanaigh, N.; Pardo, R.; Fensome, A.; Allen-Baume, V.; Jones, D.; Holt, M.R.; Cockcroft, S. Continual production of phosphatidic acid by phospholipase D is essential for antigen-stimulated membrane ruffling in cultured mast cells. Mol. Biol. Cell 2002, 13, 3730–3746. [Google Scholar] [CrossRef] [Green Version]
- Martins, M.P.; Rossi, A.; Sanches, P.R.; Bortolossi, J.C.; Martinez-Rossi, N.M. Comprehensive analysis of the dermatophyte Trichophyton rubrum transcriptional profile reveals dynamic metabolic modulation. Biochem. J. 2020, 477, 873–885. [Google Scholar] [CrossRef]
- Bitencourt, T.A.; Rezende, C.P.; Quaresemin, N.R.; Moreno, P.; Hatanaka, O.; Rossi, A.; Martinez-Rossi, N.M.; Almeida, F. Extracellular Vesicles From the Dermatophyte Trichophyton interdigitale Modulate Macrophage and Keratinocyte Functions. Front. Immunol. 2018, 9, 2343. [Google Scholar] [CrossRef] [Green Version]
- Sato, I.; Shimizu, M.; Hoshino, T.; Takaya, N. The glutathione system of Aspergillus nidulans involves a fungus-specific glutathione S-transferase. J. Biol. Chem. 2009, 284, 8042–8053. [Google Scholar] [CrossRef] [Green Version]
- Pócsi, I.; Prade, R.A.; Penninckx, M.J. Glutathione, altruistic metabolite in fungi. Adv. Microb. Physiol. 2004, 49, 1–76. [Google Scholar]
- He, Y.; Zhou, C.; Huang, M.; Tang, C.; Liu, X.; Yue, Y.; Diao, Q.; Zheng, Z.; Liu, D. Glyoxalase system: A systematic review of its biological activity, related-diseases, screening methods and small molecule regulators. Biomed. Pharmacother. 2020, 131, 110663. [Google Scholar] [CrossRef]
- Missall, T.A.; Lodge, J.K. Function of the thioredoxin proteins in Cryptococcus neoformans during stress or virulence and regulation by putative transcriptional modulators. Mol. Microbiol. 2005, 57, 847–858. [Google Scholar] [CrossRef]
- Pannala, V.R.; Dash, R.K. Mechanistic characterization of the thioredoxin system in the removal of hydrogen peroxide. Free Radic. Biol. Med. 2015, 78, 42–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, J.; Shadkchan, Y.; Osherov, N.; Krappmann, S. The Essential Thioredoxin Reductase of the Human Pathogenic Mold Aspergillus fumigatus Is a Promising Antifungal Target. Front. Microbiol. 2020, 11, 1383. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.; González, B.; Sempere, V.; Mas, A.; Torija, M.J.; Beltran, G. Melatonin Reduces Oxidative Stress Damage Induced by Hydrogen Peroxide in Saccharomyces cerevisiae. Front. Microbiol. 2017, 8, 1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkhofer, S.; Niederegger, H.; Blum, G.; Burgstaller, W.; Ledochowski, M.; Dierich, M.P.; Lass-Flörl, C. Interaction of 5-hydroxytryptamine (serotonin) against Aspergillus spp. in vitro. Int. J. Antimicrob. Agents 2007, 29, 424–429. [Google Scholar] [CrossRef] [Green Version]
- New, A.M.; Cerulus, B.; Govers, S.K.; Perez-Samper, G.; Zhu, B.; Boogmans, S.; Xavier, J.B.; Verstrepen, K.J. Different levels of catabolite repression optimize growth in stable and variable environments. PLoS Biol. 2014, 12, e1001764. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Zheng, W.; Islam, W.; Arif, M.; Abubakar, Y.S.; Wang, Z.; Lu, G. Carbon Catabolite Repression in Filamentous Fungi. Int. J. Mol. Sci. 2017, 19, 48. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, M.C.; Fink, G.R. The glyoxylate cycle is required for fungal virulence. Nature 2001, 412, 83–86. [Google Scholar] [CrossRef]
- Cruz, A.H.S.; Santos, R.S.; Martins, M.P.; Peres, N.T.A.; Trevisan, G.L.; Mendes, N.S.; Martinez-Rossi, N.M.; Rossi, A. Relevance of Nutrient-Sensing in the Pathogenesis of Trichophyton rubrum and Trichophyton interdigitale. Front. Fungal Biol. 2022, 3. [Google Scholar] [CrossRef]
- Chew, S.Y.; Ho, K.L.; Cheah, Y.K.; Ng, T.S.; Sandai, D.; Brown, A.J.P.; Than, L.T.L. Glyoxylate cycle gene ICL1 is essential for the metabolic flexibility and virulence of Candida glabrata. Sci. Rep. 2019, 9, 2843. [Google Scholar] [CrossRef] [Green Version]
- Ishchuk, O.P.; Voronovsky, A.Y.; Stasyk, O.V.; Gayda, G.Z.; Gonchar, M.V.; Abbas, C.A.; Sibirny, A.A. Overexpression of pyruvate decarboxylase in the yeast Hansenula polymorpha results in increased ethanol yield in high-temperature fermentation of xylose. FEMS Yeast Res. 2008, 8, 1164–1174. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upregulated | log2 (Fold Change) | ||
|---|---|---|---|
| ID | Gene Product Name | 3 h | 12 h |
| TERG_06548 | hypothetical protein (oxidoreductase, M. canis) | 10.86 | 11.80 |
| TERG_00785 | endoplasmic reticulum vesicle protein 25 | 8.60 | |
| TERG_01782 | hypothetical protein | 8.34 | 9.02 |
| TERG_00010 | amidase family protein (T. verrucosum) | 8.72 | |
| TERG_03829 | FAD binding domain-containing protein (T. equinum) | 6.92 | |
| TERG_04952 | multidrug resistance protein (T. equinum) | 6.87 | 7.69 |
| TERG_08954 | hypothetical protein | 6.61 | |
| TERG_06106 | sulfate permease 2 (T. tonsurans) | 6.45 | |
| TERG_08751 | ABC multidrug transporter, putative (A. benhamiae) | 5.82 | 6.33 |
| TERG_01543 | S-adenosylmethionine (SAM)-dependent methyltransferase (M. gypseum) | 5.42 | 7.48 |
| TERG_08041 | aminotransferase, putative (A. benhamiae) | 5.42 | |
| TERG_04937 | alpha/beta hydrolase (T. equinum) | 5.41 | 6.40 |
| TERG_07830 | hypothetical protein | 5.41 | 8.22 |
| Downregulated | log2 (Fold Change) | ||
| ID | Gene Product Name | 3 h | 12 h |
| TERG_02653 | hypothetical protein | −6.21 | |
| TERG_02652 | O-methyltransferase, putative (T. verrucosum) | −4.91 | |
| TERG_04066 | filamentation protein (Rhf1), putative (T. verrucosum) | −4.67 | |
| TERG_02959 | hypothetical protein | −4.16 | |
| TERG_05816 | hypothetical protein | −3.74 | |
| TERG_02650 | NmrA family protein (T. equinum) | −3.56 | |
| TERG_12339 | hypothetical protein | −3.31 | |
| TERG_01619 | toxin biosynthesis protein (Tri7), putative (T. verrucosum) | −3.20 | |
| TERG_00490 | erythromycin esterase (T. tonsurans) | −3.13 | |
| TERG_03826 | hypothetical protein | −2.77 | |
| TERG_02959 | hypothetical protein | −5.90 | |
| TERG_11536 | hypothetical protein | −5.69 | |
| TERG_11771 | hypothetical protein | −5.51 | |
| TERG_04742 | hypothetical protein | −5.29 | |
| TERG_01148 | hypothetical protein | −5.26 | |
| TERG_00490 | erythromycin esterase (T. tonsurans) | −5.24 | |
| TERG_03919 | phytoene dehydrogenase (T. equinum) | −5.20 | |
| TERG_01599 | hypothetical protein | −4.95 | |
| TERG_12035 | NB-ARC and TPR domain protein (A. benhamiae) | −4.87 | |
| TERG_11963 | hypothetical protein | −4.79 | |
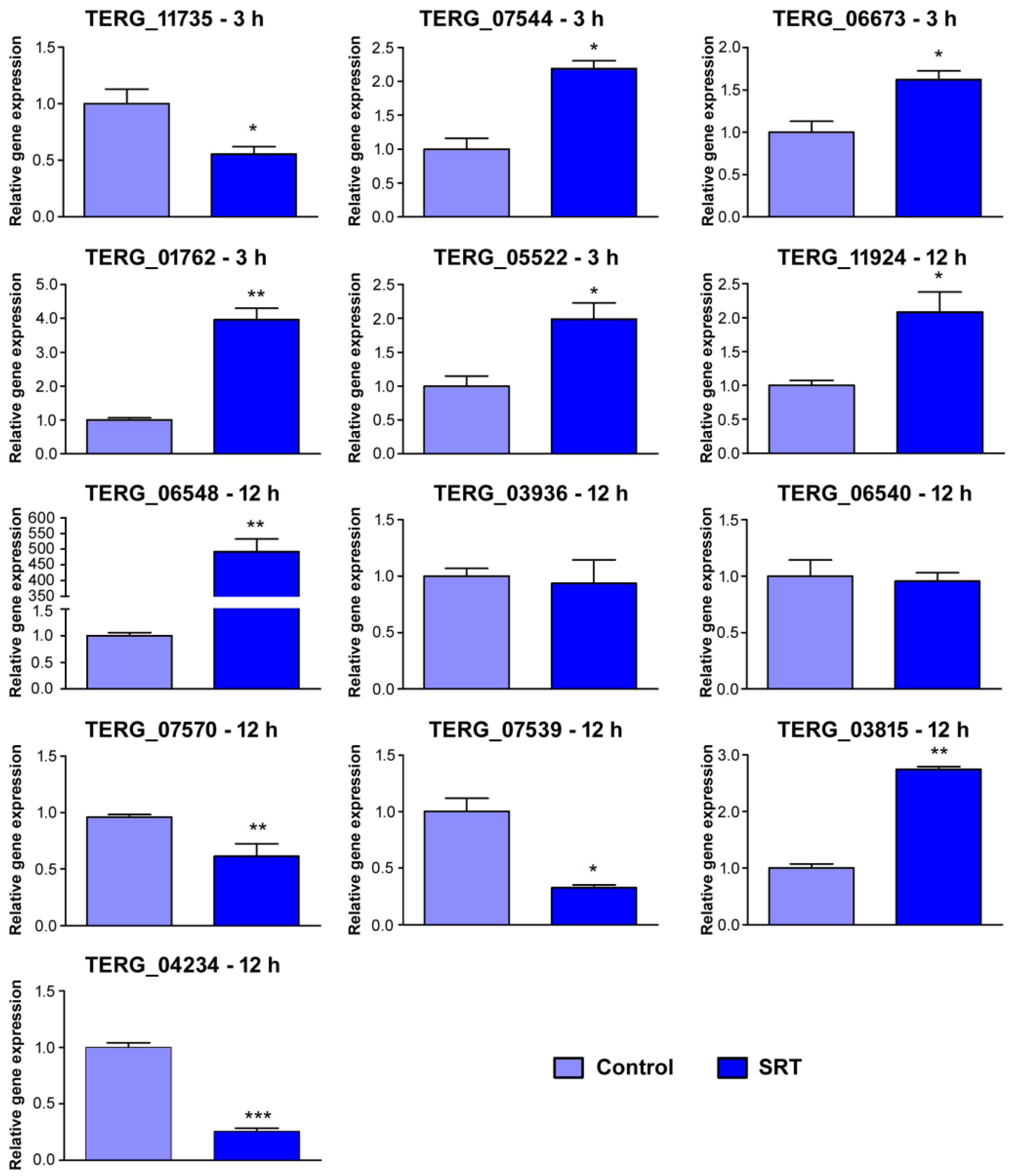
| ID | Condition | Gene Product Name | RNA-seq | RT-qPCR |
|---|---|---|---|---|
| TERG_11735 | 3 h | microtubule-associated protein (T. tonsurans) | −2.4 | −1.41 |
| TERG_07544 | 3 h | lipase (T. tonsurans) | 2.02 | 1.13 |
| TERG_06673 | 3 h | pachytene checkpoint component Pch2 (T. tonsurans) | 2.0 | 0.70 |
| TERG_01762 | 3 h | sulfite reductase (NADPH) hemoprotein beta-component | 2.14 | 1.99 |
| TERG_05522 | 3 h | lysophospholipase (T. equinum) | 1.55 | 0.97 |
| TERG_11924 | 12 h | ankyrin repeat protein (T. tonsurans) | 3.85 | 1.06 |
| TERG_06548 | 12 h | hypothetical protein (oxidoreductase, M. canis) | 11.8 | 8.63 |
| TERG_03936 | 12 h | CAMK protein kinase | −4.19 | −0.09 |
| TERG_06540 | 12 h | glutathione S-transferase (T. tonsurans) | −3.44 | −0.07 |
| TERG_07570 | 12 h | G-protein signaling regulator putative (T. verrucosum) | −3.47 | −0.70 |
| TERG_04234 | 12 h | hydrophobin putative (T. verrucosum) | −3.41 | −1.98 |
| TERG_07539 | 12 h | multidrug resistance protein (T. tonsurans) | −1.72 | −1.61 |
| TERG_03815 | 12 h | subtilisin-like protease 3 | 1.78 | 1.46 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galvão-Rocha, F.M.; Rocha, C.H.L.; Martins, M.P.; Sanches, P.R.; Bitencourt, T.A.; Sachs, M.S.; Martinez-Rossi, N.M.; Rossi, A. The Antidepressant Sertraline Affects Cell Signaling and Metabolism in Trichophyton rubrum. J. Fungi 2023, 9, 275. https://doi.org/10.3390/jof9020275
Galvão-Rocha FM, Rocha CHL, Martins MP, Sanches PR, Bitencourt TA, Sachs MS, Martinez-Rossi NM, Rossi A. The Antidepressant Sertraline Affects Cell Signaling and Metabolism in Trichophyton rubrum. Journal of Fungi. 2023; 9(2):275. https://doi.org/10.3390/jof9020275
Chicago/Turabian StyleGalvão-Rocha, Flaviane M., Carlos H. L. Rocha, Maíra P. Martins, Pablo R. Sanches, Tamires A. Bitencourt, Matthew S. Sachs, Nilce M. Martinez-Rossi, and Antonio Rossi. 2023. "The Antidepressant Sertraline Affects Cell Signaling and Metabolism in Trichophyton rubrum" Journal of Fungi 9, no. 2: 275. https://doi.org/10.3390/jof9020275
APA StyleGalvão-Rocha, F. M., Rocha, C. H. L., Martins, M. P., Sanches, P. R., Bitencourt, T. A., Sachs, M. S., Martinez-Rossi, N. M., & Rossi, A. (2023). The Antidepressant Sertraline Affects Cell Signaling and Metabolism in Trichophyton rubrum. Journal of Fungi, 9(2), 275. https://doi.org/10.3390/jof9020275