
Fusarium Photoreceptors


Abstract
:1. Introduction
2. Fusarium Photobiology
2.1. Effects of Light on Development
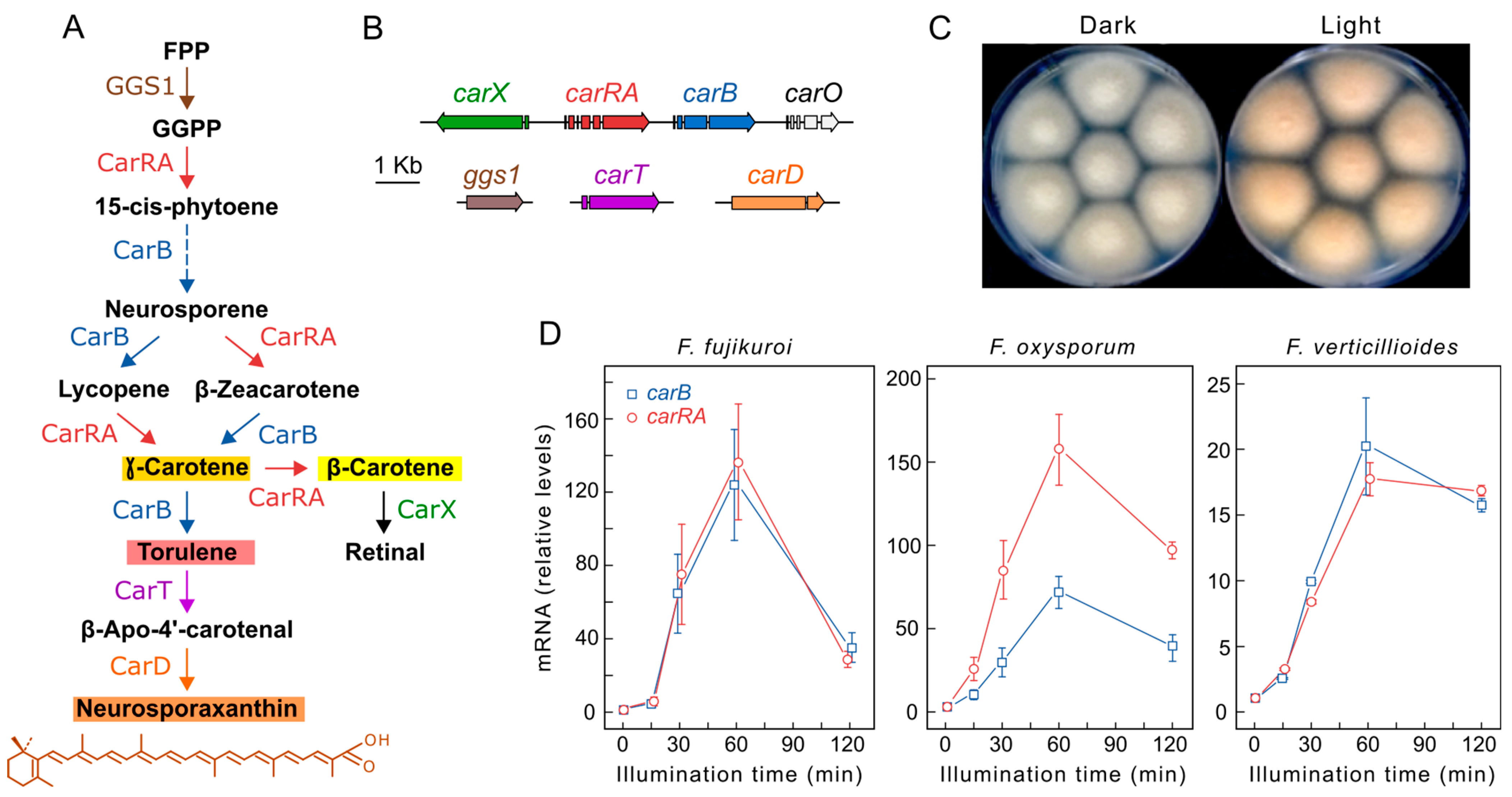
2.2. Effect of Light on Carotenogenesis
2.3. Effect of Light on the Production of Other Secondary Metabolites
3. Fungal Photoreceptors
4. Flavoproteins
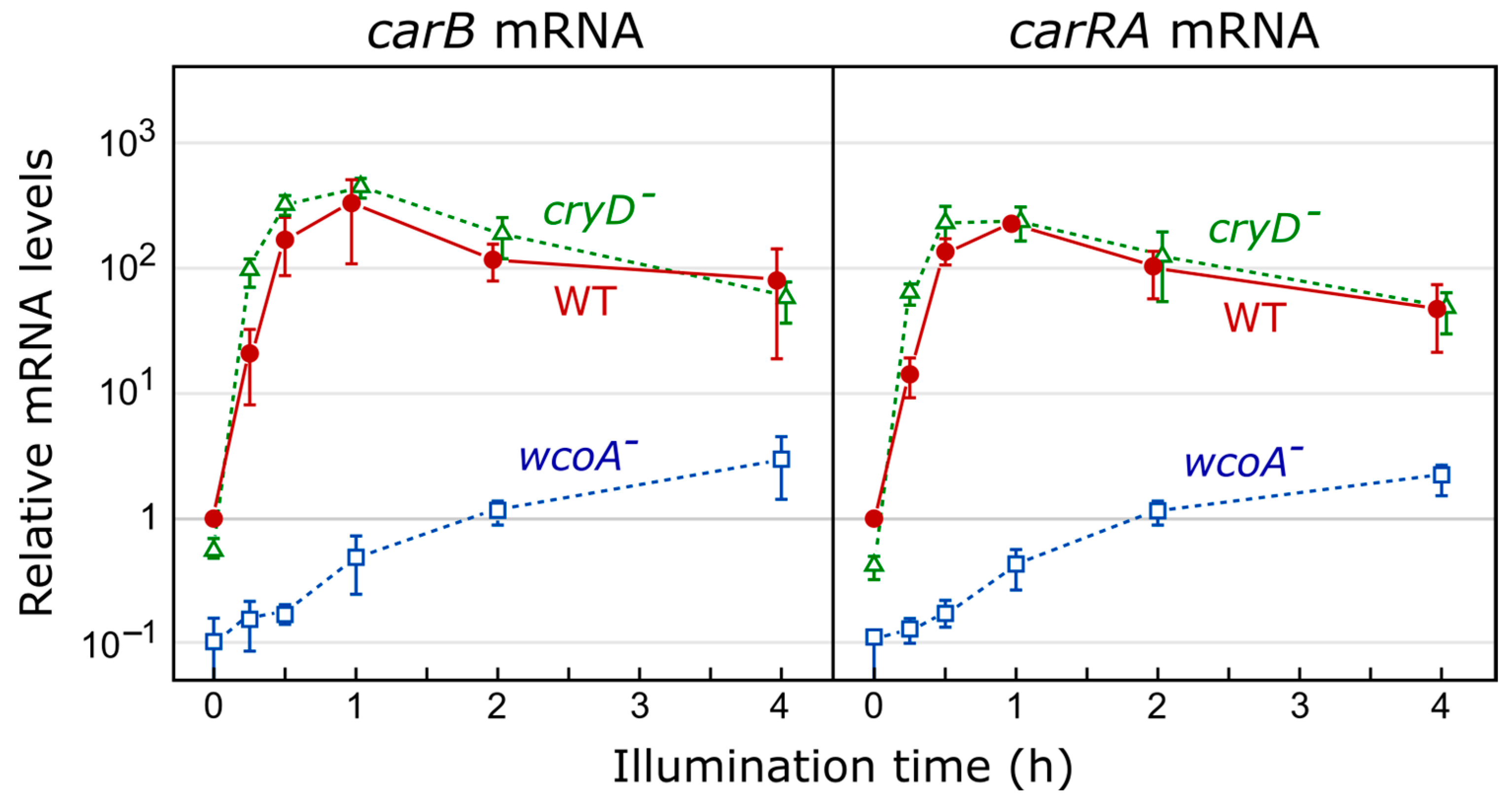
4.1. White Collar Complex
Fusarium White Collar System
4.2. VVD-Like Flavoproteins
Fusarium Vivid Protein VvdA
4.3. Other Flavin Photoreceptors
5. Cryptochromes
Fusarium Cryptochromes CryD and CryP
6. Rhodopsins
6.1. Fusarium Rhodopsins CarO and OpsA
7. Phytochromes
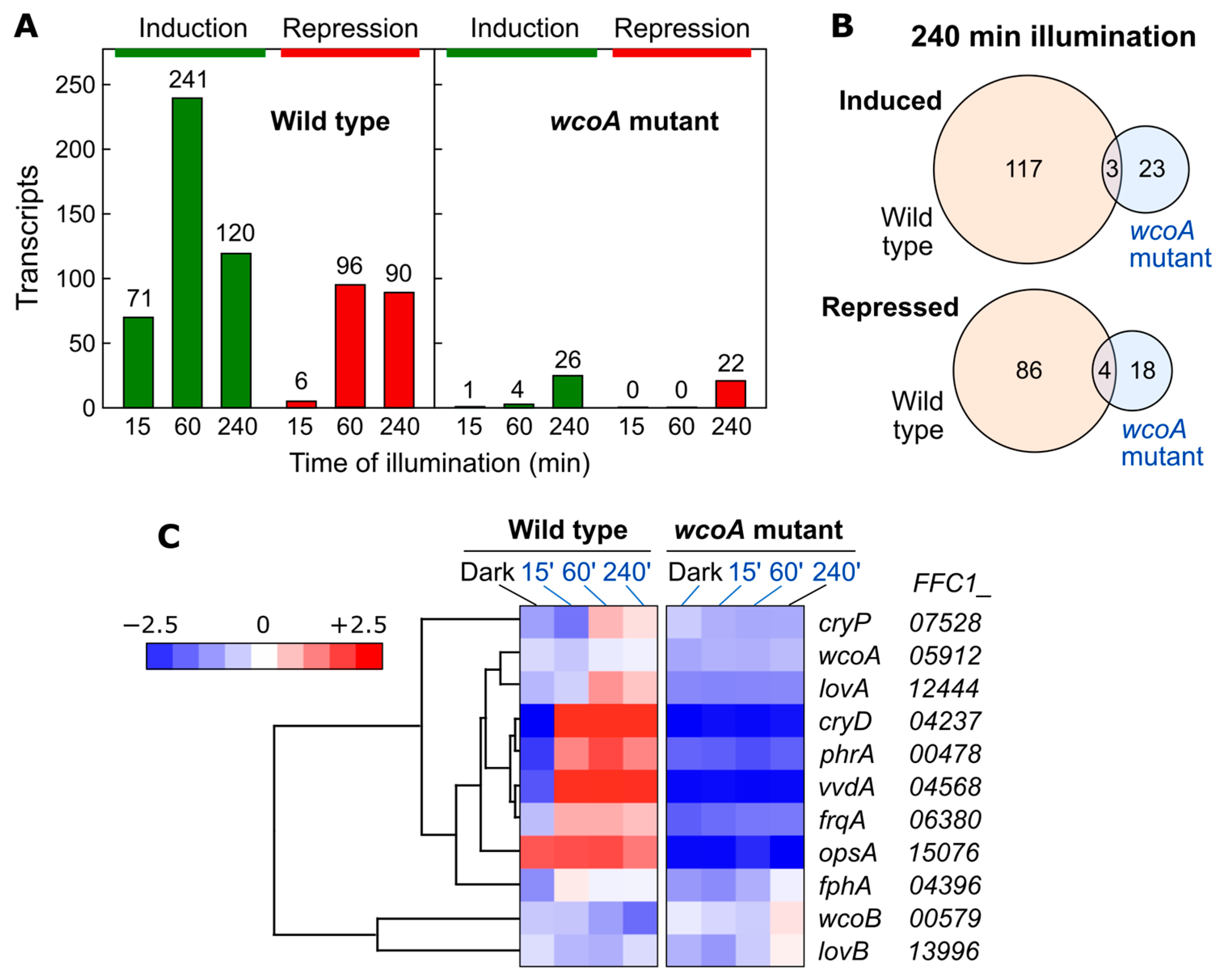
8. Study of Photoreceptor Functions through Global Transcriptomic Tools
8.1. Effect of wcoA Mutation on the Fusarium Transcriptome
8.2. Effect of cryD Deletion on the Fusarium Transcriptome
9. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fischer, R.; Aguirre, J.; Herrera-Estrella, A.; Corrochano, L.M. The Complexity of Fungal Vision. Microbiol. Spectr. 2016, 4, 1–22. [Google Scholar] [CrossRef]
- Tisch, D.; Schmoll, M. Light Regulation of Metabolic Pathways in Fungi. Appl. Microbiol. Biotechnol. 2010, 85, 1259–1277. [Google Scholar] [CrossRef]
- Yu, Z.; Fischer, R. Light Sensing and Responses in Fungi. Nat. Rev. Microbiol. 2019, 17, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Ringelberg, C.S.; Gross, R.H.; Dunlap, J.C.; Loros, J.J. Genome-Wide Analysis of Light-Inducible Responses Reveals Hierarchical Light Signalling in Neurospora. EMBO J. 2009, 28, 1029–1042. [Google Scholar] [CrossRef] [PubMed]
- Ruger-Herreros, C.; Rodríguez-Romero, J.; Fernández-Barranco, R.; Olmedo, M.; Fischer, R.; Corrochano, L.M.; Canovas, D. Regulation of Conidiation by Light in Aspergillus nidulans. Genetics 2011, 188, 809–822. [Google Scholar] [CrossRef]
- Ruger-Herreros, M.; Parra-Rivero, O.; Pardo-Medina, J.; Romero-Campero, F.J.; Limón, M.C.; Avalos, J. Comparative Transcriptomic Analysis Unveils Interactions between the Regulatory CarS Protein and Light Response in Fusarium. BMC Genom. 2019, 20, 67. [Google Scholar] [CrossRef]
- Tisch, D.; Schmoll, M. Targets of Light Signalling in Trichoderma reesei. BMC Genom. 2013, 14, 657. [Google Scholar] [CrossRef] [PubMed]
- Avalos, J.; Limón, M.C. Fungal Secondary Metabolism. Encyclopedia 2022, 2, 1–13. [Google Scholar] [CrossRef]
- Adrio, J.L.; Demain, A.L. Fungal Biotechnology. Int. Microbiol. 2003, 6, 191–199. [Google Scholar] [CrossRef]
- Gmoser, R.; Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Filamentous Ascomycetes Fungi as a Source of Natural Pigments. Fungal Biol. Biotechnol. 2017, 4, 4. [Google Scholar] [CrossRef]
- Lin, L.; Xu, J. Fungal Pigments and Their Roles Associated with Human Health. J. Fungi 2020, 6, 280. [Google Scholar] [CrossRef] [PubMed]
- Janik, E.; Niemcewicz, M.; Ceremuga, M.; Stela, M.; Saluk-Bijak, J.; Siadkowski, A.; Bijak, M. Molecular Aspects of Mycotoxins—A Serious Problem for Human Health. Int. J. Mol. Sci. 2020, 21, 8187. [Google Scholar] [CrossRef]
- Fisher, M.C.; Gurr, S.J.; Cuomo, C.A.; Blehert, D.S.; Jin, H.; Stukenbrock, E.H.; Stajich, J.E.; Kahmann, R.; Boone, C.; Denning, D.W.; et al. Threats Posed by the Fungal Kingdom to Humans, Wildlife, and Agriculture. mBio 2020, 11, e00449-20. [Google Scholar] [CrossRef] [PubMed]
- Cerón-Bustamante, M.; Balducci, E.; Beccari, G.; Nicholson, P.; Covarelli, L.; Benincasa, P. Effect of Light Spectra on Cereal Fungal Pathogens, a Review. Fungal Biol. Rev. 2022, 43, 100291. [Google Scholar] [CrossRef]
- Moss, M.O.; Smith, J.E. The Applied Mycology of Fusarium; Cambridge University Press: Cambridge, UK, 1982; ISBN 978-0-521-27924-6. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Professional: Ames, IA, USA, 2006; ISBN 0-8138-1919-9. [Google Scholar]
- Geiser, D.M.; Aoki, T.; Bacon, C.W.; Baker, S.E.; Bhattacharyya, M.K.; Brandt, M.E.; Brown, D.W.; Burgess, L.W.; Chulze, S.; Coleman, J.J.; et al. One Fungus, One Name: Defining the Genus Fusarium in a Scientifically Robust Way That Preserves Longstanding Use. Phytopathology 2013, 103, 400–408. [Google Scholar] [CrossRef]
- Rau, W. Über Den Einfluss Der Temperatur Auf Die Lichtabhängige Carotinoidbildung von Fusarium aquaeductuum. Planta 1962, 59, 123–137. [Google Scholar] [CrossRef]
- Ajmal, M.; Hussain, A.; Ali, A.; Chen, H.; Lin, H. Strategies for Controlling the Sporulation in Fusarium Spp. J. Fungi 2022, 9, 10. [Google Scholar] [CrossRef]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.-J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a Node or a Foot-Shaped Basal Cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef]
- Zemánková, M.; Lebeda, A. Fusarium Species, Their Taxonomy, Variability and Significance in Plant Pathology–a Review. Plant Protect. Sci. 2001, 37, 25–42. [Google Scholar] [CrossRef]
- Gordon, T.R. Fusarium oxysporum and the Fusarium Wilt Syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–39. [Google Scholar] [CrossRef]
- Egel, D.S.; Martyn, R.D. Fusarium Wilt of Watermelon and Other Cucurbits. Plant Health Instructor 2007, 10, 1094. [Google Scholar] [CrossRef]
- Avalos, J.; Estrada, A.F. Regulation by Light in Fusarium. Fungal Genet. Biol. 2010, 47, 930–938. [Google Scholar] [CrossRef]
- Fanelli, F.; Schmidt-Heydt, M.; Haidukowski, M.; Susca, A.; Geisen, R.; Logrieco, A.; Mulè, G. Influence of Light on Growth, Conidiation and Fumonisin Production by Fusarium verticillioides. Fungal Biol. 2012, 116, 241–248. [Google Scholar] [CrossRef]
- Niehaus, E.-M.; Kim, H.-K.; Münsterkötter, M.; Janevska, S.; Arndt, B.; Kalinina, S.A.; Houterman, P.M.; Ahn, I.-P.; Alberti, I.; Tonti, S.; et al. Comparative Genomics of Geographically Distant Fusarium fujikuroi Isolates Revealed Two Distinct Pathotypes Correlating with Secondary Metabolite Profiles. PLoS Pathog. 2017, 13, e1006670. [Google Scholar] [CrossRef]
- Costa, T.P.C.; Rodrigues, E.M.; Dias, L.P.; Pupin, B.; Ferreira, P.C.; Rangel, D.E.N. Different Wavelengths of Visible Light Influence the Conidial Production and Tolerance to Ultra-Violet Radiation of the Plant Pathogens Colletotrichum acutatum and Fusarium fujikuroi. Eur. J. Plant Pathol. 2020, 159, 105–115. [Google Scholar] [CrossRef]
- Son, H.; Kim, M.-G.; Min, K.; Seo, Y.-S.; Lim, J.Y.; Choi, G.J.; Kim, J.-C.; Chae, S.-K.; Lee, Y.-W. AbaA Regulates Conidiogenesis in the Ascomycete Fungus Fusarium graminearum. PLoS ONE 2013, 8, e72915. [Google Scholar] [CrossRef] [PubMed]
- Niehaus, E.-M.; Schumacher, J.; Burkhardt, I.; Rabe, P.; Spitzer, E.; Münsterkötter, M.; Güldener, U.; Sieber, C.M.K.; Dickschat, J.S.; Tudzynski, B. The GATA-Type Transcription Factor Csm1 Regulates Conidiation and Secondary Metabolism in Fusarium fujikuroi. Front. Microbiol. 2017, 8, 1175. [Google Scholar] [CrossRef]
- Janevska, S.; Güldener, U.; Sulyok, M.; Tudzynski, B.; Studt, L. Set1 and Kdm5 Are Antagonists for H3K4 Methylation and Regulators of the Major Conidiation-Specific Transcription Factor Gene ABA1 in Fusarium fujikuroi. Environ. Microbiol. 2018, 20, 3343–3362. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Myung, K.; Guse, D.; Donkin, B.; Proctor, R.H.; Grayburn, W.S.; Calvo, A.M. FvVE1 Regulates Filamentous Growth, the Ratio of Microconidia to Macroconidia and Cell Wall Formation in Fusarium verticillioides. Mol. Microbiol. 2006, 62, 1418–1432. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, H.-K.; Lee, S.; Yun, S.-H. The White Collar Complex Is Involved in Sexual Development of Fusarium graminearum. PLoS ONE 2015, 10, e0120293. [Google Scholar] [CrossRef]
- Tschanz, A.T.; Horst, R.K.; Nelson, P.E. The Effect of Environment on Sexual Reproduction of Gibberella zeae. Mycologia 1976, 68, 327. [Google Scholar] [CrossRef]
- Trail, F.; Xu, H.; Loranger, R.; Gadoury, D. Physiological and Environmental Aspects of Ascospore Discharge in Gibberella zeae (Anamorph Fusarium graminearum). Mycologia 2002, 94, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Avalos, J.; Pardo-Medina, J.; Parra-Rivero, O.; Ruger-Herreros, M.; Rodríguez-Ortiz, R.; Hornero-Méndez, D.; Limón, M. Carotenoid Biosynthesis in Fusarium. J. Fungi 2017, 3, 39. [Google Scholar] [CrossRef]
- Rau, W. Untersuchungen Über Die Lichtabhängige Carotinoidsynthese: II. Ersatz Der Lichtinduktion Durch Mercuribenzoat. Planta 1967, 74, 263–277. [Google Scholar] [CrossRef]
- Rau, W. Untersuchungen über die lichtabhängige Carotinoidsynthese. VII. Reversible Unterbrechung der Reaktionskette durch Cycloheximid und Anaerobe Bedingungen. Planta 1971, 101, 251–264. [Google Scholar] [CrossRef]
- Rau, W.; Feuser, B.; Rau-Hund, A. Substitution of P-Chloro- or p-Hydroxymercuribenzoate for Light in Carotenoid Synthesis by Fusarium aquaeductuum. Biochim. Biophys. Acta 1967, 136, 589–590. [Google Scholar] [CrossRef]
- Theimer, R.R.; Rau, W. Untersuchungen über die Lichtabhängige Carotinoidsynthese V. Aufhebung der Lichtinduktion Dutch Reduktionsmittel und Ersatz des Lichts Durch Wasserstoffperoxid. Planta 1970, 92, 129–137. [Google Scholar] [CrossRef]
- Theimer, R.R.; Rau, W. Untersuchungen über die lichtabhängige Carotinoidsynthese. VIII. Die unterschiedlichen Wirkungsmechanismen von Licht und Mercuribenzoat. Planta 1972, 106, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Ávalos, J.; Cerdá-Olmedo, E. Chemical Modification of Carotenogenesis in Gibberella fujikuroi. Phytochemistry 1986, 25, 1837–1841. [Google Scholar] [CrossRef]
- Rodríguez-Ortiz, R.; Michielse, C.; Rep, M.; Limón, M.C.; Avalos, J. Genetic Basis of Carotenoid Overproduction in Fusarium oxysporum. Fungal Genet. Biol. 2012, 49, 684–696. [Google Scholar] [CrossRef] [PubMed]
- Adám, A.L.; García-Martínez, J.; Szucs, E.P.; Avalos, J.; Hornok, L. The MAT1-2-1 Mating-Type Gene Upregulates Photo-Inducible Carotenoid Biosynthesis in Fusarium verticillioides. FEMS Microbiol. Lett. 2011, 318, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Sánchez, V.; Limón, M.C.; Schaub, P.; Al-Babili, S.; Avalos, J. A RALDH-like Enzyme Involved in Fusarium verticillioides Development. Fungal Genet. Biol. 2016, 86, 20–32. [Google Scholar] [CrossRef]
- Mende, K.; Homann, V.; Tudzynski, B. The Geranylgeranyl Diphosphate Synthase Gene of Gibberella fujikuroi: Isolation and Expression. Mol. Genet. Genom. 1997, 255, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Sánchez, V.; Estrada, A.F.; Trautmann, D.; Al-Babili, S.; Avaard, J. The Gene carD Encodes the Aldehyde Dehydrogenase Responsible for Neurosporaxanthin Biosynthesis in Fusarium fujikuroi. FEBS J. 2011, 278, 3164–3176. [Google Scholar] [CrossRef] [PubMed]
- Wiemann, P.; Brown, D.W.; Kleigrewe, K.; Bok, J.W.; Keller, N.P.; Humpf, H.-U.; Tudzynski, B. FfVel1 and FfLae1, Components of a Velvet-like Complex in Fusarium fujikuroi, Affect Differentiation, Secondary Metabolism and Virulence. Mol. Microbiol. 2010, 77, 972–994. [Google Scholar] [CrossRef]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.-J.; Keller, N.P.; Yu, J.-H.; et al. VelB/VeA/LaeA Complex Coordinates Light Signal with Fungal Development and Secondary Metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef]
- Niehaus, E.-M.; Rindermann, L.; Janevska, S.; Münsterkötter, M.; Güldener, U.; Tudzynski, B. Analysis of the Global Regulator Lae1 Uncovers a Connection between Lae1 and the Histone Acetyltransferase HAT1 in Fusarium fujikuroi. Appl. Microbiol. Biotechnol. 2018, 102, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, M.; Avalos, J. The Flavoproteins CryD and VvdA Cooperate with the White Collar Protein WcoA in the Control of Photocarotenogenesis in Fusarium fujikuroi. PLoS ONE 2015, 10, e0119785. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, M.; García-Martínez, J.; Avalos, J. Light-Dependent Functions of the Fusarium fujikuroi CryD DASH Cryptochrome in Development and Secondary Metabolism. Appl. Environ. Microbiol. 2013, 79, 2777–2788. [Google Scholar] [CrossRef]
- Audhya, T.K.; Russell, D.W. Production of Enniatins by Fusarium sambucinum: Selection of High-Yield Conditions from Liquid Surface Cultures. J. Gen. Microbiol. 1974, 82, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Bayram, Ö.S.; Dettmann, A.; Karahoda, B.; Moloney, N.M.; Ormsby, T.; McGowan, J.; Cea-Sánchez, S.; Miralles-Durán, A.; Brancini, G.T.P.; Luque, E.M.; et al. Control of Development, Secondary Metabolism and Light-Dependent Carotenoid Biosynthesis by the Velvet Complex of Neurospora crassa. Genetics 2019, 212, 691–710. [Google Scholar] [CrossRef] [PubMed]
- Bayram, O.; Braus, G.H.; Fischer, R.; Rodriguez-Romero, J. Spotlight on Aspergillus nidulans Photosensory Systems. Fungal Genet. Biol. 2010, 47, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Gil-Sánchez, M.D.M.; Cea-Sánchez, S.; Luque, E.M.; Cánovas, D.; Corrochano, L.M. Light Regulates the Degradation of the Regulatory Protein VE-1 in the Fungus Neurospora crassa. BMC Biol. 2022, 20, 149. [Google Scholar] [CrossRef]
- Jiang, J.; Liu, X.; Yin, Y.; Ma, Z. Involvement of a Velvet Protein FgVeA in the Regulation of Asexual Development, Lipid and Secondary Metabolisms and Virulence in Fusarium graminearum. PLoS ONE 2011, 6, e28291. [Google Scholar] [CrossRef] [PubMed]
- Merhej, J.; Urban, M.; Dufresne, M.; Hammond-Kosack, K.E.; Richard-Forget, F.; Barreau, C. The Velvet Gene, FgVe1, Affects Fungal Development and Positively Regulates Trichothecene Biosynthesis and Pathogenicity in Fusarium graminearum. Mol. Plant Pathol. 2012, 13, 363–374. [Google Scholar] [CrossRef]
- López-Berges, M.S.; Hera, C.; Sulyok, M.; Schäfer, K.; Capilla, J.; Guarro, J.; Di Pietro, A. The Velvet Complex Governs Mycotoxin Production and Virulence of Fusarium oxysporum on Plant and Mammalian Hosts. Mol. Microbiol. 2013, 87, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Beattie, G.A.; Hatfield, B.M.; Dong, H.; McGrane, R.S. Seeing the Light: The Roles of Red- and Blue-Light Sensing in Plant Microbes. Annu. Rev. Phytopathol. 2018, 56, 41–66. [Google Scholar] [CrossRef]
- Purschwitz, J.; Müller, S.; Kastner, C.; Schöser, M.; Haas, H.; Espeso, E.A.; Atoui, A.; Calvo, A.M.; Fischer, R. Functional and Physical Interaction of Blue- and Red-Light Sensors in Aspergillus nidulans. Curr. Biol. 2008, 18, 255–259. [Google Scholar] [CrossRef]
- Schumacher, J. How Light Affects the Life of Botrytis. Fungal Genet. Biol. 2017, 106, 26–41. [Google Scholar] [CrossRef]
- Corrochano, L.M. Light in the Fungal World: From Photoreception to Gene Transcription and Beyond. Annu. Rev. Genet. 2019, 53, 149–170. [Google Scholar] [CrossRef]
- Estrada, A.F.; Avalos, J. The White Collar Protein WcoA of Fusarium fujikuroi Is Not Essential for Photocarotenogenesis, but Is Involved in the Regulation of Secondary Metabolism and Conidiation. Fungal Genet. Biol. 2008, 45, 705–718. [Google Scholar] [CrossRef]
- Kim, H.; Son, H.; Lee, Y.-W. Effects of Light on Secondary Metabolism and Fungal Development of Fusarium graminearum. J. Appl. Microbiol. 2014, 116, 380–389. [Google Scholar] [CrossRef]
- Ruiz-Roldán, M.C.; Garre, V.; Guarro, J.; Mariné, M.; Roncero, M.I. Role of the White Collar 1 Photoreceptor in Carotenogenesis, UV Resistance, Hydrophobicity, and Virulence of Fusarium oxysporum. Eukaryot. Cell 2008, 7, 1227–1230. [Google Scholar] [CrossRef]
- Tang, Y.; Zhu, P.; Lu, Z.; Qu, Y.; Huang, L.; Zheng, N.; Wang, Y.; Nie, H.; Jiang, Y.; Xu, L. The Photoreceptor Components FaWC1 and FaWC2 of Fusarium asiaticum Cooperatively Regulate Light Responses but Play Independent Roles in Virulence Expression. Microorganisms 2020, 8, 365. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, M.; Avalos, J. Light-Mediated Participation of the VIVID-like Protein of Fusarium fujikuroi VvdA in Pigmentation and Development. Fungal Genet. Biol. 2014, 71, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, M.; Bernhardt, A.; Avalos, J.; Batschauer, A.; Pokorny, R. Biochemical Characterization of the DASH-Type Cryptochrome CryD from Fusarium fujikuroi. Photochem. Photobiol. 2015, 91, 1356–1367. [Google Scholar] [CrossRef]
- Alejandre-Durán, E.; Roldán-Arjona, T.; Ariza, R.R.; Ruiz-Rubio, M. The Photolyase Gene from the Plant Pathogen Fusarium oxysporum f. Sp. lycopersici Is Induced by Visible Light and Alpha-Tomatine from Tomato Plant. Fungal Genet. Biol. 2003, 40, 159–165. [Google Scholar] [CrossRef]
- Adam, A.; Deimel, S.; Pardo-Medina, J.; García-Martínez, J.; Konte, T.; Limón, M.C.; Avalos, J.; Terpitz, U. Protein Activity of the Fusarium fujikuroi Rhodopsins CarO and OpsA and Their Relation to Fungus-Plant Interaction. Int. J. Mol. Sci. 2018, 19, 215. [Google Scholar] [CrossRef]
- Prado, M.M.; Prado-Cabrero, A.; Fernández-Martín, R.; Avalos, J. A Gene of the Opsin Family in the Carotenoid Gene Cluster of Fusarium fujikuroi. Curr. Genet. 2004, 46, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Estrada, A.F.; Avalos, J. Regulation and Targeted Mutation of opsA, Coding for the NOP-1 Opsin Orthologue in Fusarium fujikuroi. J. Mol. Biol. 2009, 387, 59–73. [Google Scholar] [CrossRef]
- Iwata, T.; Masuda, S. Photoreaction Mechanisms of Flavoprotein Photoreceptors and Their Applications. Adv. Exp. Med. Biol. 2021, 1293, 189–206. [Google Scholar] [CrossRef]
- Harding, R.W.; Turner, R.V. Photoregulation of the Carotenoid Biosynthetic Pathway in Albino and White Collar Mutants of Neurospora crassa. Plant Physiol. 1981, 68, 745–749. [Google Scholar] [CrossRef]
- Ballario, P.; Vittorioso, P.; Magrelli, A.; Talora, C.; Cabibbo, A.; Macino, G. White Collar-1, a Central Regulator of Blue Light Responses in Neurospora, Is a Zinc Finger Protein. EMBO J. 1996, 15, 1650–1657. [Google Scholar] [CrossRef] [PubMed]
- Linden, H.; Macino, G. White Collar 2, a Partner in Blue-Light Signal Transduction, Controlling Expression of Light-Regulated Genes in Neurospora crassa. EMBO J. 1997, 16, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Degli-Innocenti, F.; Russo, V.E. Isolation of New White Collar Mutants of Neurospora crassa and Studies on Their Behavior in the Blue Light-Induced Formation of Protoperithecia. J. Bacteriol. 1984, 159, 757–761. [Google Scholar] [CrossRef]
- van der Horst, M.A.; Hellingwerf, K.J. Photoreceptor Proteins, “Star Actors of Modern Times”: A Review of the Functional Dynamics in the Structure of Representative Members of Six Different Photoreceptor Families. Acc. Chem. Res. 2004, 37, 13–20. [Google Scholar] [CrossRef]
- Crosson, S.; Moffat, K. Photoexcited Structure of a Plant Photoreceptor Domain Reveals a Light-Driven Molecular Switch. Plant Cell 2002, 14, 1067–1075. [Google Scholar] [CrossRef]
- Pfeifer, A.; Majerus, T.; Zikihara, K.; Matsuoka, D.; Tokutomi, S.; Heberle, J.; Kottke, T. Time-Resolved Fourier Transform Infrared Study on Photoadduct Formation and Secondary Structural Changes within the Phototropin LOV Domain. Biophys. J. 2009, 96, 1462–1470. [Google Scholar] [CrossRef]
- Briggs, W.R. The LOV Domain: A Chromophore Module Servicing Multiple Photoreceptors. J. Biomed. Sci. 2007, 14, 499–504. [Google Scholar] [CrossRef]
- Taylor, B.L.; Zhulin, I.B. PAS Domains: Internal Sensors of Oxygen, Redox Potential, and Light. Microbiol. Mol. Biol. Rev. 1999, 63, 479–506. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Yang, F.; Smith, K.M.; Peterson, M.; Dekhang, R.; Zhang, Y.; Zucker, J.; Bredeweg, E.L.; Mallappa, C.; Zhou, X.; et al. Genome-Wide Characterization of Light-Regulated Genes in Neurospora crassa. G3 Genes Genomes Genet. 2014, 4, 1731–1745. [Google Scholar] [CrossRef]
- Sancar, C.; Ha, N.; Yilmaz, R.; Tesorero, R.; Fisher, T.; Brunner, M.; Sancar, G. Combinatorial Control of Light Induced Chromatin Remodeling and Gene Activation in Neurospora. PLoS Genet. 2015, 11, e1005105. [Google Scholar] [CrossRef] [PubMed]
- Malzahn, E.; Ciprianidis, S.; Káldi, K.; Schafmeier, T.; Brunner, M. Photoadaptation in Neurospora by Competitive Interaction of Activating and Inhibitory LOV Domains. Cell 2010, 142, 762–772. [Google Scholar] [CrossRef]
- Brenna, A.; Grimaldi, B.; Filetici, P.; Ballario, P. Physical Association of the WC-1 Photoreceptor and the Histone Acetyltransferase NGF-1 Is Required for Blue Light Signal Transduction in Neurospora crassa. Mol. Biol. Cell 2012, 23, 3863–3872. [Google Scholar] [CrossRef]
- Grimaldi, B.; Coiro, P.; Filetici, P.; Berge, E.; Dobosy, J.R.; Freitag, M.; Selker, E.U.; Ballario, P. The Neurospora crassa White Collar-1 Dependent Blue Light Response Requires Acetylation of Histone H3 Lysine 14 by NGF-1. Mol. Biol. Cell 2006, 17, 4576–4583. [Google Scholar] [CrossRef] [PubMed]
- Káldi, K.; González, B.H.; Brunner, M. Transcriptional Regulation of the Neurospora Circadian Clock Gene wc-1 Affects the Phase of Circadian Output. EMBO Rep. 2006, 7, 199–204. [Google Scholar] [CrossRef]
- Schwerdtfeger, C.; Linden, H. Localization and Light-Dependent Phosphorylation of White Collar 1 and 2, the Two Central Components of Blue Light Signaling in Neurospora crassa. Eur. J. Biochem. 2000, 267, 414–422. [Google Scholar] [CrossRef]
- He, Q.; Liu, Y. Molecular Mechanism of Light Responses in Neurospora: From Light-Induced Transcription to Photoadaptation. Genes Dev. 2005, 19, 2888–2899. [Google Scholar] [CrossRef]
- Schafmeier, T.; Haase, A.; Káldi, K.; Scholz, J.; Fuchs, M.; Brunner, M. Transcriptional Feedback of Neurospora Circadian Clock Gene by Phosphorylation-Dependent Inactivation of Its Transcription Factor. Cell 2005, 122, 235–246. [Google Scholar] [CrossRef]
- Dunlap, J.C.; Loros, J.J. Making Time: Conservation of Biological Clocks from Fungi to Animals. In The Fungal Kingdom; Heitman, J., Howlett, B.J., Crous, P.W., Stukenbrock, E.H., James, T.Y., Gow, N.A., Eds.; American Society of Microbiology: Washington, DC, USA, 2017; pp. 515–534. ISBN 978-1-55581-957-6. [Google Scholar]
- Franchi, L.; Fulci, V.; Macino, G. Protein Kinase C Modulates Light Responses in Neurospora by Regulating the Blue Light Photoreceptor WC-1. Mol. Microbiol. 2005, 56, 334–345. [Google Scholar] [CrossRef]
- Cheng, P.; Yang, Y.; Gardner, K.H.; Liu, Y. PAS Domain-Mediated WC-1/WC-2 Interaction Is Essential for Maintaining the Steady-State Level of WC-1 and the Function of Both Proteins in Circadian Clock and Light Responses of Neurospora. Mol. Cell. Biol. 2002, 22, 517–524. [Google Scholar] [CrossRef]
- Schafmeier, T.; Diernfellner, A.C.R. Light Input and Processing in the Circadian Clock of Neurospora. FEBS Lett. 2011, 585, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Idnurm, A.; Verma, S.; Corrochano, L.M. A Glimpse into the Basis of Vision in the Kingdom Mycota. Fungal Genet. Biol. 2010, 47, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Sanz, C.; Rodríguez-Romero, J.; Idnurm, A.; Christie, J.M.; Heitman, J.; Corrochano, L.M.; Eslava, A.P. Phycomyces MADB Interacts with MADA to Form the Primary Photoreceptor Complex for Fungal Phototropism. Proc. Natl. Acad. Sci. USA 2009, 106, 7095–7100. [Google Scholar] [CrossRef]
- Silva, F.; Torres-Martínez, S.; Garre, V. Distinct White Collar-1 Genes Control Specific Light Responses in Mucor circinelloides. Mol. Microbiol. 2006, 61, 1023–1037. [Google Scholar] [CrossRef]
- Idnurm, A.; Heitman, J. Light Controls Growth and Development via a Conserved Pathway in the fungal Kingdom. PLoS Biol. 2005, 3, e95. [Google Scholar] [CrossRef]
- Brych, A.; Mascarenhas, J.; Jaeger, E.; Charkiewicz, E.; Pokorny, R.; Bölker, M.; Doehlemann, G.; Batschauer, A. White Collar 1-induced Photolyase Expression Contributes to UV -tolerance of Ustilago maydis. Microbiologyopen 2016, 5, 224–243. [Google Scholar] [CrossRef]
- Qi, Y.; Sun, X.; Ma, L.; Wen, Q.; Qiu, L.; Shen, J. Identification of Two Pleurotus ostreatus Blue Light Receptor Genes (PoWC-1 and PoWC-2) and in Vivo Confirmation of Complex PoWC-12 Formation through Yeast Two Hybrid System. Fungal Biol. 2020, 124, 8–14. [Google Scholar] [CrossRef]
- Castellanos, F.; Schmoll, M.; Martínez, P.; Tisch, D.; Kubicek, C.P.; Herrera-Estrella, A.; Esquivel-Naranjo, E.U. Crucial Factors of the Light Perception Machinery and Their Impact on Growth and Cellulase Gene Transcription in Trichoderma reesei. Fungal Genet. Biol. 2010, 47, 468–476. [Google Scholar] [CrossRef]
- Canessa, P.; Schumacher, J.; Hevia, M.A.; Tudzynski, P.; Larrondo, L.F. Assessing the Effects of Light on Differentiation and Virulence of the Plant Pathogen Botrytis cinerea: Characterization of the White collar Complex. PLoS ONE 2013, 8, e84223. [Google Scholar] [CrossRef] [PubMed]
- Corrochano, L.M.; Garre, V. Photobiology in the Zygomycota: Multiple Photoreceptor Genes for Complex Responses to Light. Fungal Genet. Biol. 2010, 47, 893–899. [Google Scholar] [CrossRef]
- Silva, F.; Navarro, E.; Peñaranda, A.; Murcia-Flores, L.; Torres-Martínez, S.; Garre, V. A RING-Finger Protein Regulates Carotenogenesis via Proteolysis-Independent Ubiquitylation of a White Collar-1-like Activator. Mol. Microbiol. 2008, 70, 1026–1036. [Google Scholar] [CrossRef]
- Brandhoff, B.; Simon, A.; Dornieden, A.; Schumacher, J. Regulation of Conidiation in Botrytis cinerea Involves the Light-Responsive Transcriptional Regulators BcLTF3 and BcREG1. Curr. Genet. 2017, 63, 931–949. [Google Scholar] [CrossRef]
- Schumacher, J.; Simon, A.; Cohrs, K.C.; Viaud, M.; Tudzynski, P. The Transcription Factor BcLTF1 Regulates Virulence and Light Responses in the Necrotrophic Plant Pathogen Botrytis cinerea. PLoS Genet. 2014, 10, e1004040. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M. The VeA Regulatory System and Its Role in Morphological and Chemical Development in Fungi. Fungal Genet. Biol. 2008, 45, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- De Fabo, E.C.; Harding, R.W.; Shropshire, W. Action Spectrum between 260 and 800 Nanometers for the Photoinduction of Carotenoid Biosynthesis in Neurospora crassa. Plant Physiol. 1976, 57, 440–445. [Google Scholar] [CrossRef]
- Rau, W. Untersuchungen Über Die Lichtabhängige Carotinoidsynthese. I. Das Wirkungsspektrum von Fusarium aquaeductuum. Planta 1967, 72, 14–28. [Google Scholar] [CrossRef]
- Schwerdtfeger, C.; Linden, H. Blue Light Adaptation and Desensitization of Light Signal Transduction in Neurospora crassa. Mol. Microbiol. 2001, 39, 1080–1087. [Google Scholar] [CrossRef]
- Zoltowski, B.D.; Crane, B.R. Light Activation of the LOV Protein Vivid Generates a Rapidly Exchanging Dimer. Biochemistry 2008, 47, 7012–7019. [Google Scholar] [CrossRef]
- Schwerdtfeger, C.; Linden, H. VIVID Is a Flavoprotein and Serves as a Fungal Blue Light Photoreceptor for Photoadaptation. EMBO J. 2003, 22, 4846–4855. [Google Scholar] [CrossRef] [PubMed]
- Zoltowski, B.D.; Schwerdtfeger, C.; Widom, J.; Loros, J.J.; Bilwes, A.M.; Dunlap, J.C.; Crane, B.R. Conformational Switching in the Fungal Light Sensor Vivid. Science 2007, 316, 1054–1057. [Google Scholar] [CrossRef]
- Chen, C.-H.; DeMay, B.S.; Gladfelter, A.S.; Dunlap, J.C.; Loros, J.J. Physical Interaction between VIVID and White Collar Complex Regulates Photoadaptation in Neurospora. Proc. Natl. Acad. Sci. USA 2010, 107, 16715–16720. [Google Scholar] [CrossRef]
- Hunt, S.M.; Thompson, S.; Elvin, M.; Heintzen, C. VIVID Interacts with the WHITE COLLAR Complex and FREQUENCY-Interacting RNA Helicase to Alter Light and Clock Responses in Neurospora. Proc. Natl. Acad. Sci. USA 2010, 107, 16709–16714. [Google Scholar] [CrossRef]
- Dasgupta, A.; Chen, C.-H.; Lee, C.; Gladfelter, A.S.; Dunlap, J.C.; Loros, J.J. Biological Significance of Photoreceptor Photocycle Length: VIVID Photocycle Governs the Dynamic VIVID-White Collar Complex Pool Mediating Photo-Adaptation and Response to Changes in Light Intensity. PLoS Genet. 2015, 11, e1005215. [Google Scholar] [CrossRef]
- Ruger-Herreros, C.; Gil-Sánchez, M.D.M.; Sancar, G.; Brunner, M.; Corrochano, L.M. Alteration of Light-Dependent Gene Regulation by the Absence of the RCO-1/RCM-1 Repressor Complex in the Fungus Neurospora crassa. PLoS ONE 2014, 9, e95069. [Google Scholar] [CrossRef] [PubMed]
- Ruesch, C.E.; Ramakrishnan, M.; Park, J.; Li, N.; Chong, H.S.; Zaman, R.; Joska, T.M.; Belden, W.J. The Histone H3 Lysine 9 Methyltransferase DIM-5 Modifies Chromatin at Frequency and Represses Light-Activated Gene Expression. G3 Genes Genomes Genet. 2014, 5, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Schmoll, M.; Franchi, L.; Kubicek, C.P. Envoy, a PAS/LOV Domain Protein of Hypocrea jecorina (Anamorph Trichoderma reesei), Modulates Cellulase Gene Transcription in Response to Light. Eukaryot. Cell 2005, 4, 1998–2007. [Google Scholar] [CrossRef] [PubMed]
- Schmoll, M.; Zeilinger, S.; Mach, R.L.; Kubicek, C.P. Cloning of Genes Expressed Early during Cellulase Induction in Hypocrea jecorina by a Rapid Subtraction Hybridization Approach. Fungal Genet. Biol. 2004, 41, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, T.; Guo, D.; Gao, J.; Zhang, J.; Cai, F.; Fischer, R.; Shen, Q.; Yu, Z. Comprehensive Analysis of the Regulatory Network of Blue-Light-Regulated Conidiation and Hydrophobin Production in Trichoderma guizhouense. Environ. Microbiol. 2021, 23, 6241–6256. [Google Scholar] [CrossRef]
- Schmoll, M. Light, Stress, Sex and Carbon–The Photoreceptor ENVOY as a Central Checkpoint in the Physiology of Trichoderma reesei. Fungal Biol. 2018, 122, 479–486. [Google Scholar] [CrossRef]
- Tong, S.-M.; Zhang, A.-X.; Guo, C.-T.; Ying, S.-H.; Feng, M.-G. Daylight Length-Dependent Translocation of VIVID Photoreceptor in Cells and Its Essential Role in Conidiation and Virulence of Beauveria bassiana. Environ. Microbiol. 2018, 20, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tang, W.; Liu, K.; Huang, Q.; Zhang, X.; Yan, X.; Chen, Y.; Wang, J.; Qi, Z.; Wang, Z.; et al. Eight RGS and RGS-like Proteins Orchestrate Growth, Differentiation, and Pathogenicity of Magnaporthe oryzae. PLoS Pathog. 2011, 7, e1002450. [Google Scholar] [CrossRef]
- Ozturk, N. Phylogenetic and Functional Classification of the Photolyase/Cryptochrome Family. Photochem. Photobiol. 2017, 93, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Sancar, A. Photolyase and Cryptochrome Blue-Light Photoreceptors. Adv. Protein Chem. 2004, 69, 73–100. [Google Scholar] [CrossRef]
- Kiontke, S.; Göbel, T.; Brych, A.; Batschauer, A. DASH-Type Cryptochromes–Solved and Open Questions. Biol. Chem. 2020, 401, 1487–1493. [Google Scholar] [CrossRef]
- Navarro, E.; Niemann, N.; Kock, D.; Dadaeva, T.; Gutiérrez, G.; Engelsdorf, T.; Kiontke, S.; Corrochano, L.M.; Batschauer, A.; Garre, V. The DASH-Type Cryptochrome from the Fungus Mucor circinelloides Is a Canonical CPD-Photolyase. Curr. Biol. 2020, 30, 4483–4490.e4. [Google Scholar] [CrossRef]
- Tagua, V.G.; Pausch, M.; Eckel, M.; Gutiérrez, G.; Miralles-Durán, A.; Sanz, C.; Eslava, A.P.; Pokorny, R.; Corrochano, L.M.; Batschauer, A. Fungal Cryptochrome with DNA Repair Activity Reveals an Early Stage in Cryptochrome Evolution. Proc. Natl. Acad. Sci. USA 2015, 112, 15130–15135. [Google Scholar] [CrossRef]
- Bayram, O.; Biesemann, C.; Krappmann, S.; Galland, P.; Braus, G.H. More than a Repair Enzyme: Aspergillus nidulans Photolyase-like CryA Is a Regulator of Sexual Development. Mol. Biol. Cell 2008, 19, 3254–3262. [Google Scholar] [CrossRef] [PubMed]
- Froehlich, A.C.; Chen, C.-H.; Belden, W.J.; Madeti, C.; Roenneberg, T.; Merrow, M.; Loros, J.J.; Dunlap, J.C. Genetic and Molecular Characterization of a Cryptochrome from the Filamentous Fungus Neurospora crassa. Eukaryot. Cell 2010, 9, 738–750. [Google Scholar] [CrossRef] [PubMed]
- Olmedo, M.; Ruger-Herreros, C.; Luque, E.M.; Corrochano, L.M. A Complex Photoreceptor System Mediates the Regulation by Light of the Conidiation Genes con-10 and con-6 in Neurospora crassa. Fungal Genet. Biol. 2010, 47, 352–363. [Google Scholar] [CrossRef]
- Cohrs, K.C.; Schumacher, J. The Two Cryptochrome/Photolyase Family Proteins Fulfill Distinct Roles in DNA Photorepair and Regulation of Conidiation in the Gray Mold Fungus Botrytis cinerea. Appl. Environ. Microbiol. 2017, 83, e00812-17. [Google Scholar] [CrossRef]
- Veluchamy, S.; Rollins, J.A. A CRY-DASH-Type Photolyase/Cryptochrome from Sclerotinia sclerotiorum Mediates Minor UV-A-Specific Effects on Development. Fungal Genet. Biol. 2008, 45, 1265–1276. [Google Scholar] [CrossRef]
- Wang, F.; Song, X.; Dong, X.; Zhang, J.; Dong, C. DASH-Type Cryptochromes Regulate Fruiting Body Development and Secondary Metabolism Differently than CmWC-1 in the Fungus Cordyceps militaris. Appl. Microbiol. Biotechnol. 2017, 101, 4645–4657. [Google Scholar] [CrossRef] [PubMed]
- Pardo-Medina, J. Regulatory Roles of Photoreceptors and Non-Coding RNA in Fusarium fujikuro. Ph.D. Thesis, Universidad de Sevilla, Seville, Spain, 2021. [Google Scholar]
- Pardo-Medina, J.; Gutiérrez, G.; Limón, M.C.; Avalos, J. Impact of the White Collar Photoreceptor WcoA on the Fusarium fujikuroi Transcriptome. Front. Microbiol. 2021, 11, 619474. [Google Scholar] [CrossRef]
- Milo-Cochavi, S.; Adar, S.; Covo, S. Developmentally Regulated Oscillations in the Expression of UV Repair Genes in a Soilborne Plant Pathogen Dictate UV Repair Efficiency and Survival. mBio 2019, 10, e02623-19. [Google Scholar] [CrossRef] [PubMed]
- Nagata, T.; Inoue, K. Rhodopsins at a Glance. J. Cell Sci. 2021, 134, jcs258989. [Google Scholar] [CrossRef]
- Brown, L.S. Fungal Rhodopsins and Opsin-Related Proteins: Eukaryotic Homologues of Bacteriorhodopsin with Unknown Functions. Photochem. Photobiol. Sci. 2004, 3, 555–565. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, J.; Li, N.; Li, J.; Trail, F.; Dunlap, J.C.; Townsend, J.P. Light Sensing by Opsins and Fungal Ecology: NOP-1 Modulates Entry into Sexual Reproduction in Response to Environmental Cues. Mol. Ecol. 2018, 27, 216–232. [Google Scholar] [CrossRef] [PubMed]
- García-Martínez, J.; Brunk, M.; Avalos, J.; Terpitz, U. The CarO Rhodopsin of the Fungus Fusarium fujikuroi Is a Light-Driven Proton Pump That Retards Spore Germination. Sci. Rep. 2015, 5, 7798. [Google Scholar] [CrossRef]
- Rodriguez-Romero, J.; Hedtke, M.; Kastner, C.; Müller, S.; Fischer, R. Fungi, Hidden in Soil or up in the Air: Light Makes a Difference. Annu. Rev. Microbiol. 2010, 64, 585–610. [Google Scholar] [CrossRef]
- Panzer, S.; Brych, A.; Batschauer, A.; Terpitz, U. Opsin 1 and Opsin 2 of the Corn Smut Fungus Ustilago maydis Are Green Light-Driven Proton Pumps. Front. Microbiol. 2019, 10, 735. [Google Scholar] [CrossRef] [PubMed]
- Keszthelyi, A.; Jeney, A.; Kerényi, Z.; Mendes, O.; Waalwijk, C.; Hornok, L. Tagging Target Genes of the MAT1-2-1 Transcription Factor in Fusarium verticillioides (Gibberella fujikuroi MP-A). Antonie Leeuwenhoek 2007, 91, 373–391. [Google Scholar] [CrossRef]
- Rensing, S.A.; Sheerin, D.J.; Hiltbrunner, A. Phytochromes: More Than Meets the Eye. Trends Plant Sci. 2016, 21, 543–546. [Google Scholar] [CrossRef]
- Blumenstein, A.; Vienken, K.; Tasler, R.; Purschwitz, J.; Veith, D.; Frankenberg-Dinkel, N.; Fischer, R. The Aspergillus nidulans Phytochrome FphA Represses Sexual Development in Red Light. Curr. Biol. 2005, 15, 1833–1838. [Google Scholar] [CrossRef]
- Froehlich, A.C.; Noh, B.; Vierstra, R.D.; Loros, J.; Dunlap, J.C. Genetic and Molecular Analysis of Phytochromes from the Filamentous Fungus Neurospora crassa. Eukaryot. Cell 2005, 4, 2140–2152. [Google Scholar] [CrossRef]
- Schumacher, J.; Gorbushina, A.A. Light Sensing in Plant- and Rock-Associated Black Fungi. Fungal Biol. 2020, 124, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Pires, S.; Espeso, E.A.; Rasiukevičiūtė, N.; Melgarejo, P.; De Cal, A. Light-Photoreceptors and Proteins Related to Monilinia laxa Photoresponses. J. Fungi 2021, 7, 32. [Google Scholar] [CrossRef]
- Streng, C.; Hartmann, J.; Leister, K.; Krauß, N.; Lamparter, T.; Frankenberg-Dinkel, N.; Weth, F.; Bastmeyer, M.; Yu, Z.; Fischer, R. Fungal Phytochrome Chromophore Biosynthesis at Mitochondria. EMBO J. 2021, 40, e108083. [Google Scholar] [CrossRef] [PubMed]
- Röhrig, J.; Kastner, C.; Fischer, R. Light Inhibits Spore Germination through Phytochrome in Aspergillus nidulans. Curr. Genet. 2013, 59, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Armant, O.; Fischer, R. Fungi Use the SakA (HogA) Pathway for Phytochrome-Dependent Light Signalling. Nat. Microbiol. 2016, 1, 16019. [Google Scholar] [CrossRef]
- Hedtke, M.; Rauscher, S.; Röhrig, J.; Rodríguez-Romero, J.; Yu, Z.; Fischer, R. Light-Dependent Gene Activation in Aspergillus nidulans Is Strictly Dependent on Phytochrome and Involves the Interplay of Phytochrome and White Collar-Regulated Histone H3 Acetylation. Mol. Microbiol. 2015, 97, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Rauscher, S.; Pacher, S.; Hedtke, M.; Kniemeyer, O.; Fischer, R. A Phosphorylation Code of the Aspergillus nidulans Global Regulator VelvetA (VeA) Determines Specific Functions. Mol. Microbiol. 2016, 99, 909–924. [Google Scholar] [CrossRef]
- Igbalajobi, O.; Yu, Z.; Fischer, R. Red- and Blue-Light Sensing in the Plant Pathogen Alternaria alternata Depends on Phytochrome and the White-Collar Protein LreA. mBio 2019, 10, e00371-19. [Google Scholar] [CrossRef]
- Igbalajobi, O.; Gao, J.; Fischer, R. The HOG Pathway Plays Different Roles in Conidia and Hyphae during Virulence of Alternaria alternata. Mol. Plant Microbe Interact. 2020, 33, 1405–1410. [Google Scholar] [CrossRef]
- Hayes, S.; Schachtschabel, J.; Mishkind, M.; Munnik, T.; Arisz, S.A. Hot Topic: Thermosensing in Plants. Plant. Cell Environ. 2021, 44, 2018–2033. [Google Scholar] [CrossRef] [PubMed]
- Njimona, I.; Lamparter, T. Temperature Effects on Agrobacterium Phytochrome Agp1. PLoS ONE 2011, 6, e25977. [Google Scholar] [CrossRef]
- Yu, Z.; Ali, A.; Igbalajobi, O.A.; Streng, C.; Leister, K.; Krauß, N.; Lamparter, T.; Fischer, R. Two Hybrid Histidine Kinases, TcsB and the Phytochrome FphA, Are Involved in Temperature Sensing in Aspergillus nidulans. Mol. Microbiol. 2019, 112, 1814–1830. [Google Scholar] [CrossRef]
- Yu, Z.; Streng, C.; Seibeld, R.F.; Igbalajobi, O.A.; Leister, K.; Ingelfinger, J.; Fischer, R. Genome-Wide Analyses of Light-Regulated Genes in Aspergillus nidulans Reveal a Complex Interplay between Different Photoreceptors and Novel Photoreceptor Functions. PLoS Genet. 2021, 17, e1009845. [Google Scholar] [CrossRef] [PubMed]
- Casal, J.J. Photoreceptor Signaling Networks in Plant Responses to Shade. Annu. Rev. Plant Biol. 2013, 64, 403–427. [Google Scholar] [CrossRef]
- Virtanen, O.; Constantinidou, E.; Tyystjärvi, E. Chlorophyll Does Not Reflect Green Light–How to Correct a Misconception. J. Biol. Educ. 2022, 56, 552–559. [Google Scholar] [CrossRef]
- Fuller, K.K.; Dunlap, J.C.; Loros, J.J. Fungal Light Sensing at the Bench and Beyond. Adv. Genet. 2016, 96, 1–51. [Google Scholar] [CrossRef]
- Fuller, K.K.; Ringelberg, C.S.; Loros, J.J.; Dunlap, J.C. The Fungal Pathogen Aspergillus fumigatus Regulates Growth, Metabolism, and Stress Resistance in Response to Light. MBio 2013, 4, e00142-13. [Google Scholar] [CrossRef] [PubMed]
- Tagua, V.G.; Navarro, E.; Gutiérrez, G.; Garre, V.; Corrochano, L.M. Light Regulates a Phycomyces blakesleeanus Gene Family Similar to the Carotenogenic Repressor Gene of Mucor circinelloides. Fungal Biol. 2020, 124, 338–351. [Google Scholar] [CrossRef] [PubMed]
- Cetz-Chel, J.E.; Balcázar-López, E.; Esquivel-Naranjo, E.U.; Herrera-Estrella, A. The Trichoderma atroviride Putative Transcription Factor Blu7 Controls Light Responsiveness and Tolerance. BMC Genom. 2016, 17, 327. [Google Scholar] [CrossRef]
- Brenna, A.; Talora, C. WC-1 and the Proximal GATA Sequence Mediate a Cis-/Trans-Acting Repressive Regulation of Light-Dependent Gene Transcription in the Dark. Int. J. Mol. Sci. 2019, 20, 2854. [Google Scholar] [CrossRef] [PubMed]
- Pola-Sánchez, E.; Villalobos-Escobedo, J.M.; Carreras-Villaseñor, N.; Martínez-Hernández, P.; Beltrán-Hernández, E.B.; Esquivel-Naranjo, E.U.; Herrera-Estrella, A. A Global Analysis of Photoreceptor-Mediated Transcriptional Changes Reveals the Intricate Relationship between Central Metabolism and DNA Repair in the Filamentous Fungus Trichoderma atroviride. Front. Microbiol. 2021, 12, 724676. [Google Scholar] [CrossRef]
- Krobanan, K.; Liang, S.-W.; Chiu, H.-C.; Shen, W.-C. The Blue-Light Photoreceptor sfwc-1 Gene Regulates the Phototropic Response and Fruiting-Body Development in the Homothallic Ascomycete Sordaria fimicola. Appl. Environ. Microbiol. 2019, 85, e02206-18. [Google Scholar] [CrossRef]
- Idnurm, A.; Heitman, J. Ferrochelatase Is a Conserved Downstream Target of the Blue Light-Sensing White Collar Complex in Fungi. Microbiology 2010, 156, 2393–2407. [Google Scholar] [CrossRef]
- Parra-Rivero, O.; Paes de Barros, M.; Prado, M.D.M.; Gil, J.V.; Hornero-Méndez, D.; Zacarías, L.; Rodrigo, M.J.; Limón, M.C.; Avalos, J. Neurosporaxanthin Overproduction by Fusarium fujikuroi and Evaluation of Its Antioxidant Properties. Antioxidants 2020, 9, 528. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Chromophore 1 | Protein 2 | References 3 | F. fujikuroi 4 | F. oxysporum 5 | F. graminearum 6 | |
|---|---|---|---|---|---|---|---|
| Flavoproteins | Flavin (blue) | WCC 7 | WcoA | [32,63,64,65,66] | FFUJ_13691 | FOXG_03727 | FGSG_07941 |
| WcoB | [32,65,66] | FFUJ_00530 | FOXG_01037 | FGSG_00710 | |||
| VvdA | [50,67] | FFUJ_06055 | FOXG_03254 | FGSG_08456 | |||
| LovA | - | FFUJ_11713 | FOXG_12253 | FGSG_02972 | |||
| LovB | - | FFUJ_08848 | FOXG_09176 | FGSG_04991 | |||
| Cryptochrome photolyase family | MTHF/flavin (blue) | CryD | [50,51,68] | FFUJ_05732 | FOXG_03570 | FGSG_08852 | |
| CryP | - | FFUJ_03105 | FOXG_02060 | FGSG_06765 | |||
| PhrA | [69] | FFUJ_00436 | FOXG_01134 | FGSG_00797 | |||
| Rhodopsins | Retinal (green) | CarO | [70,71] | FFUJ_11804 | FOXG_12142 | FGSG_03064 | |
| OpsA | [70,72] | FFUJ_02352 | FOXG_15406 | FGSG_07554 | |||
| Phytochromes | Biliverdin (red) | FphA | [32] | FFUJ_05887 | FOXG_03424 | FGSG_08608 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pardo-Medina, J.; Limón, M.C.; Avalos, J. Fusarium Photoreceptors. J. Fungi 2023, 9, 319. https://doi.org/10.3390/jof9030319
Pardo-Medina J, Limón MC, Avalos J. Fusarium Photoreceptors. Journal of Fungi. 2023; 9(3):319. https://doi.org/10.3390/jof9030319
Chicago/Turabian StylePardo-Medina, Javier, M. Carmen Limón, and Javier Avalos. 2023. "Fusarium Photoreceptors" Journal of Fungi 9, no. 3: 319. https://doi.org/10.3390/jof9030319
APA StylePardo-Medina, J., Limón, M. C., & Avalos, J. (2023). Fusarium Photoreceptors. Journal of Fungi, 9(3), 319. https://doi.org/10.3390/jof9030319