
Exploring the Mycovirus Universe: Identification, Diversity, and Biotechnological Applications

 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methodology
Literature Review and Acquisition of Genomic Data
3. Results and Discussion
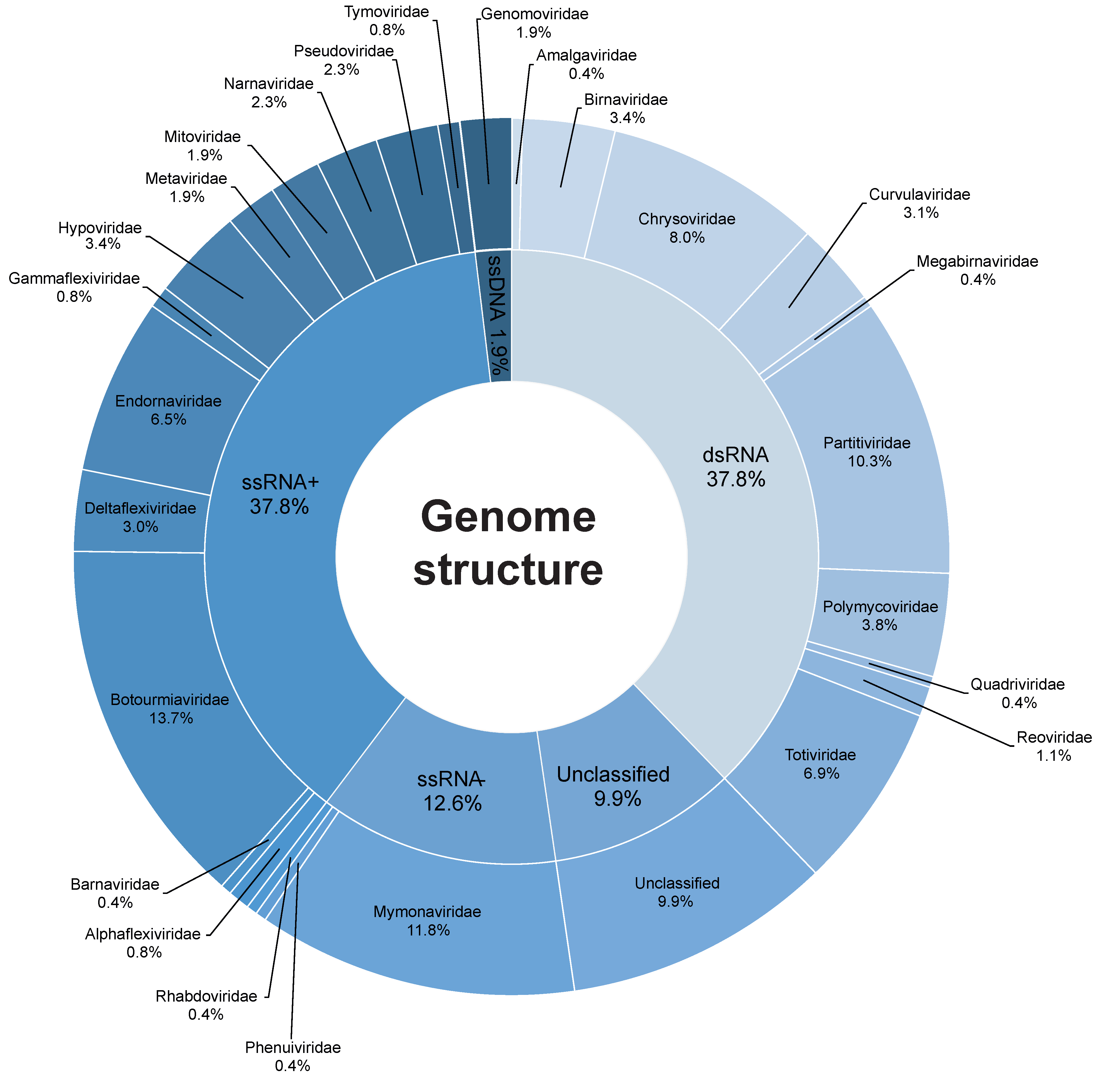
3.1. Data Curation
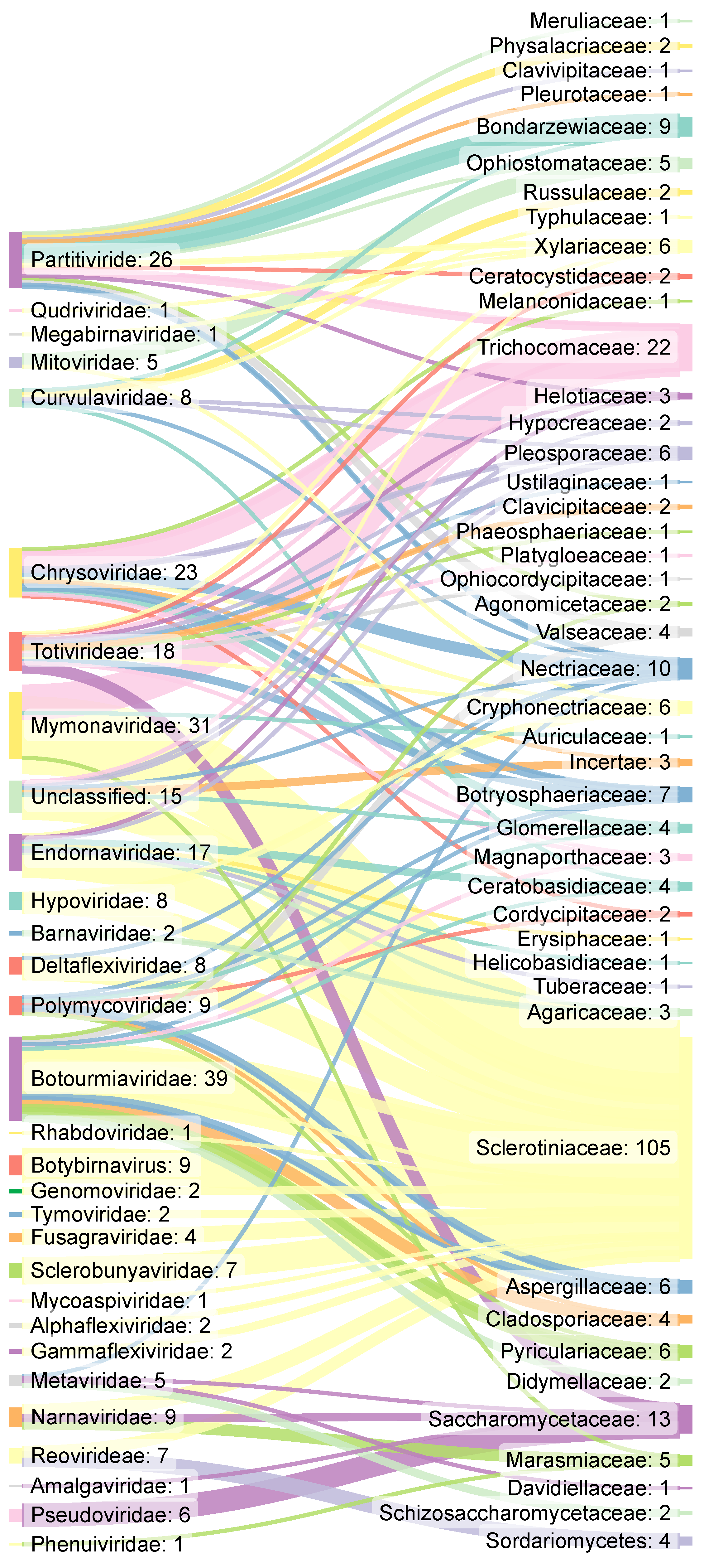
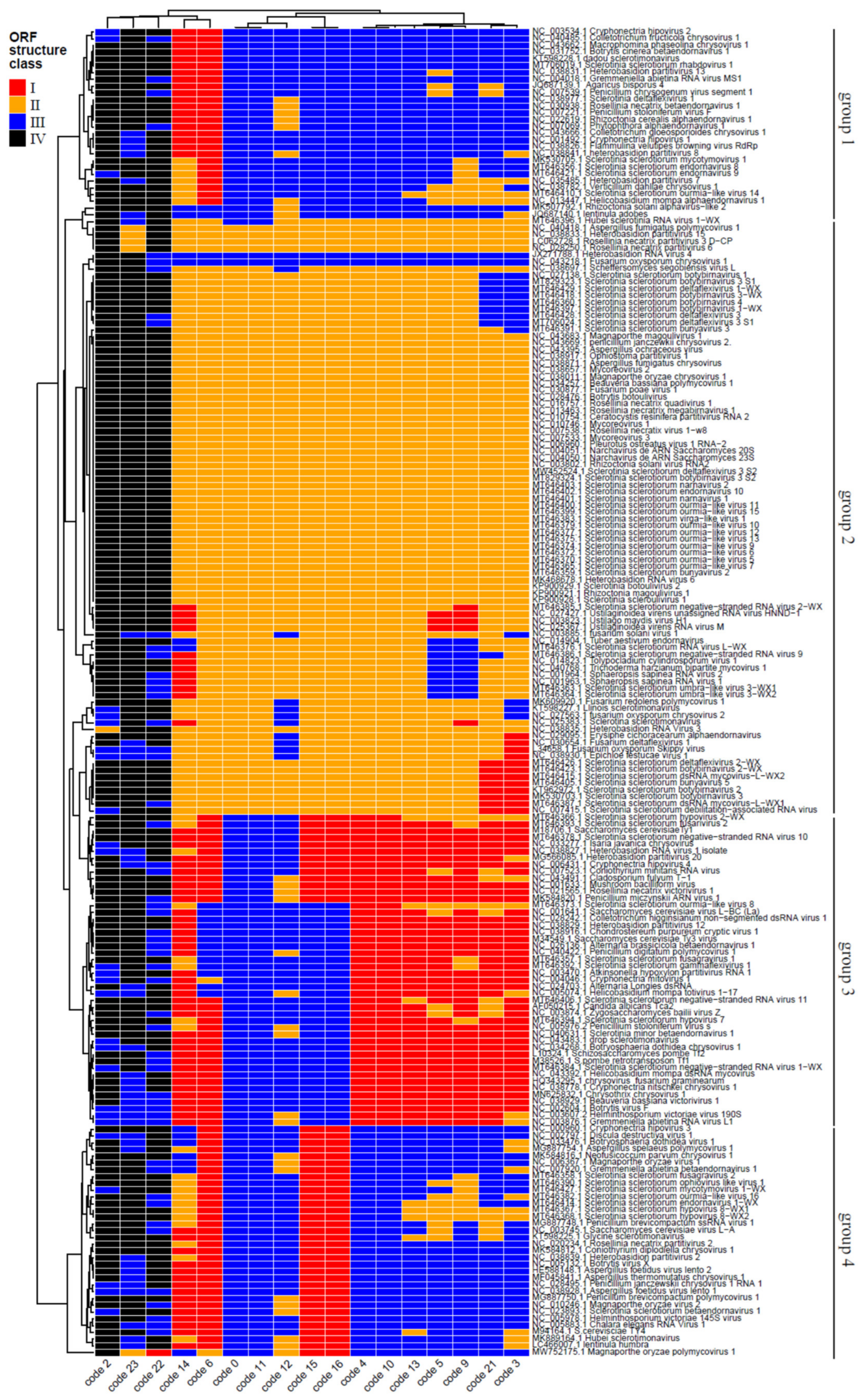
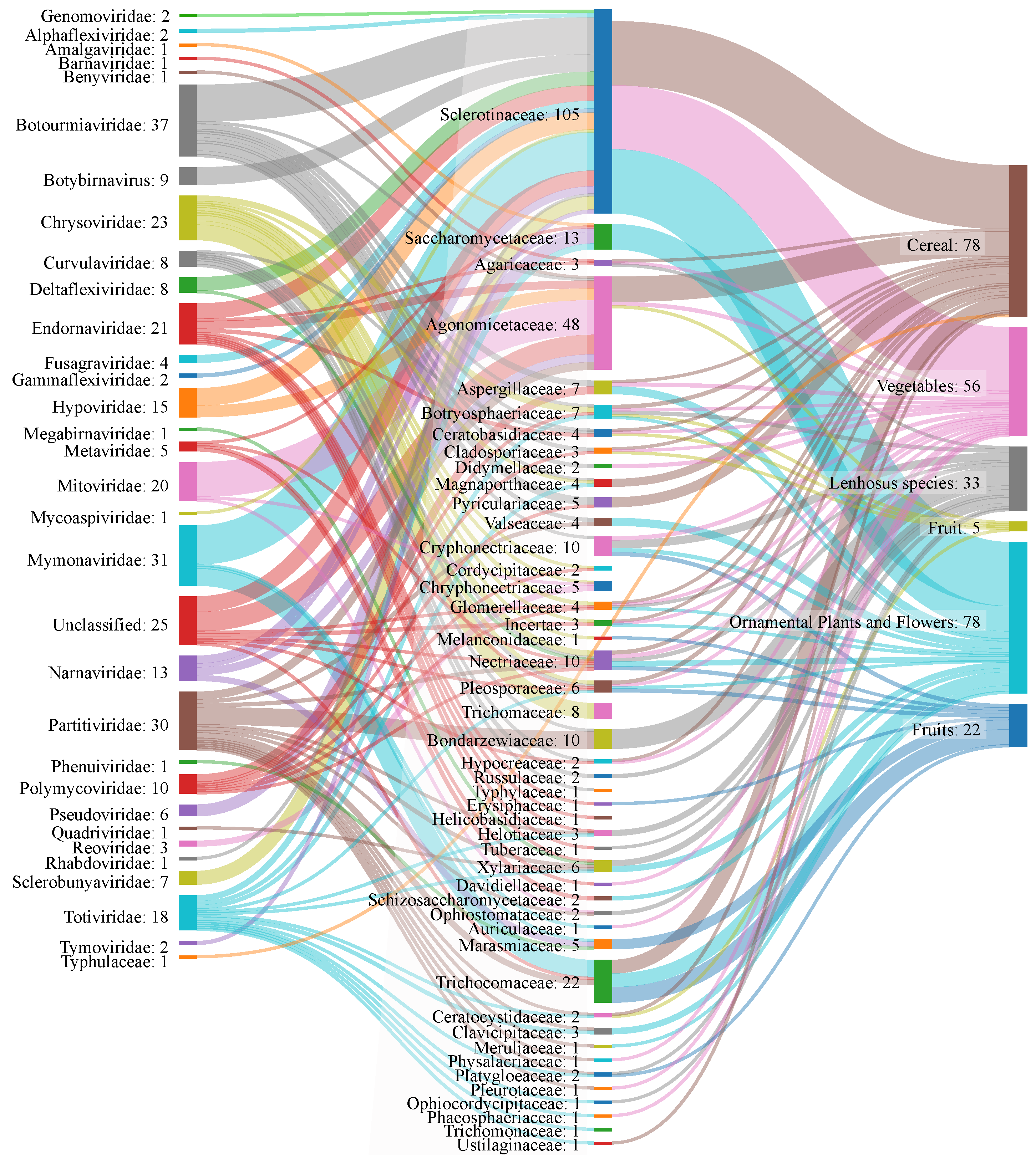
3.2. Mycovirus Diversity: Fungal Hosts, Main Viral Families, Genomic Aspects and Relationship with the Host Cell
3.3. Cases of Mycovirus as Hypervirulent Agents with Increased Pathogenicity
3.4. Cases of Mycovirus as Biological Control Agents (Hypovirulent Phenotypes)
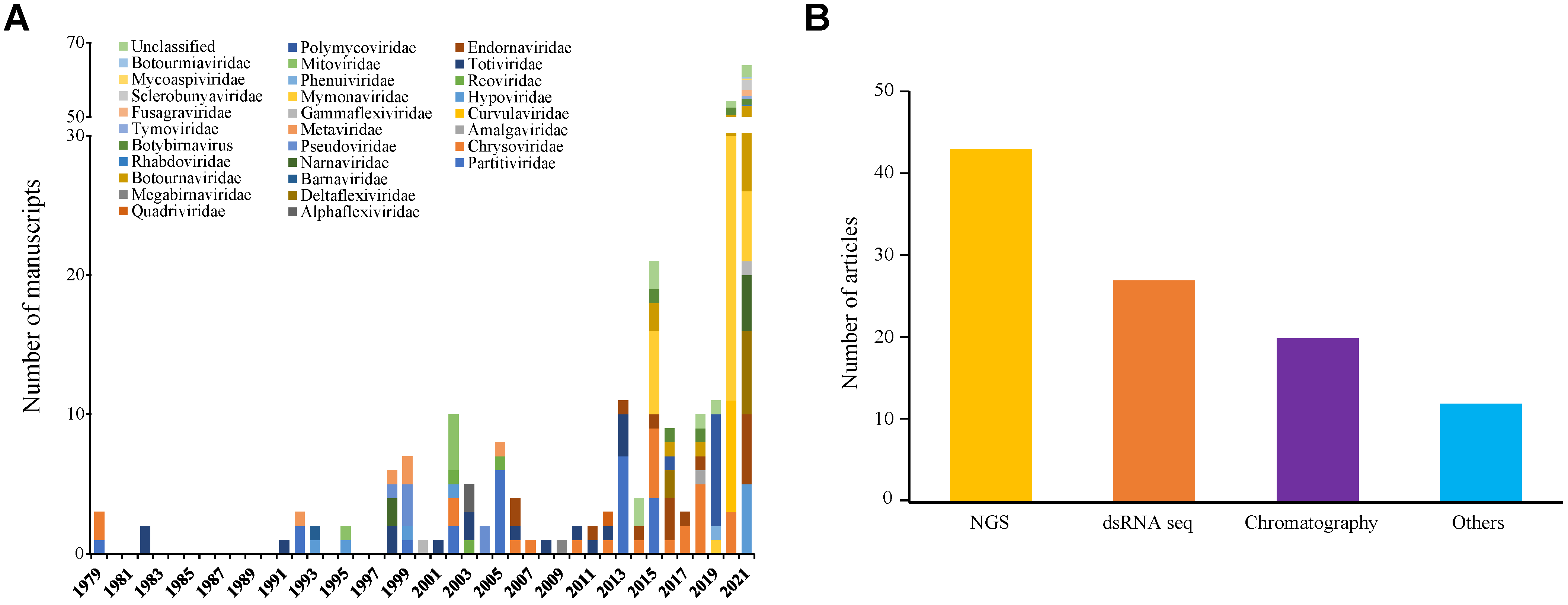
3.5. The Impact of High-Throughput Sequencing in Mycovirus Studies
3.6. Outlook and Perspectives in Mycoviral Research
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AfuCV | Aspergillus fumigatus chrysovirus |
| AfuPV-1 | Aspergillus fumigatus partitivirus-1 |
| AfuVmycovirus-1 | Aspergillus fumigatus tetramycovirus-1 |
| BbPmV-1 | Beauveria bassiana polymycovirus 1 |
| BbVV-1 | Beauveria bassiana victorivirus 1 |
| cDNA | complementary DNA |
| ChNRV1 | Colletotrichum higginsianum virus non-segmented dsRNA 1 |
| CHV1 | Cryphonectria hypovirus 1 |
| DNA | Deoxyribonucleic acid |
| DOI | Digital Object Identifier in scientific articles |
| DsRNA | Double-stranded RNA |
| ggplot2 | Open-source data visualization package for the R statistical programming language |
| ICTV | International Committee on Virus Taxonomy |
| LTR | Long terminal repeats: characteristic nucleotide sequence found at each end of a retroviral element that has been integrated into the host genome |
| MBV | Fungal bacilliform virus |
| Mycovirus | Virus that infects fungi |
| IPM | Integrated Pest Management |
| mRNA | messenger RNA |
| NCBI | National Center for Biotechnology Information |
| ORF | Open Reading Window |
| PubMed | an open-access search engine that allows searching primarily the contents of the MEDLINE database and other scientific journals |
| RdRp | RNA-dependent RNA polymerase |
| RNA | Ribonucleic acid |
| RNAi | RNA interference RNA |
| Sankeyplot | A specific type of flow chart, in which the width of the arrows is shown to be proportional to the amount of flow |
| ScNV-20S | Saccharomyces 20S RNA narnavirus |
| ScNV-23S | Saccharomyces 23S RNA narnavirus |
| Scopus | a bibliographic database of abstracts and citations of articles from scientific journals |
| SmEV1 | Sclerotinia minor endornavirus 1 |
| ssDNA | single-stranded DNA |
| ssRNA | single-stranded RNA |
| tRNAs | transporter RNA |
| TuVmycovirus-1 | Tetramycovirus-1 |
| VCG | vegetative compatibility groups |
| VLPs | virus-like particle |
| WoS | Web of Science is an online scientific information service |
References
- James, T.Y.; Stajich, J.E.; Hittinger, C.T.; Rokas, A. Toward a Fully Resolved Fungal Tree of Life. Annu. Rev. Microbiol. 2020, 74, 291–313. [Google Scholar] [CrossRef]
- Lutzoni, F.; Kauff, F.; Cox, C.J.; McLaughlin, D.; Celio, G.; Dentinger, B.; Padamsee, M.; Hibbett, D.; James, T.Y.; Baloch, E.; et al. Assembling the Fungal Tree of Life: Progress, Classification, and Evolution of Subcellular Traits. Am. J. Bot. 2004, 91, 1446–1480. [Google Scholar] [CrossRef]
- Richards, T.A.; Leonard, G.; Wideman, J.G. What Defines the “Kingdom” Fungi? Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Amorim, A.O.; Orlandelli, R.C.; Pamphile, J.A. Controle de patógenos do cacaueiro (Theobroma cacao L.) por fungos endofíticos dos gêneros trichoderma e clonostachys. Rev. UNINGÁ Rev. 2019, 34, 1–10. [Google Scholar]
- Muneret, L.; Auriol, A.; Thiéry, D.; Rusch, A. Organic Farming at Local and Landscape Scales Fosters Biological Pest Control in Vineyards. Ecol. Appl. Publ. Ecol. Soc. Am. 2019, 29, e01818. [Google Scholar] [CrossRef] [Green Version]
- Wyckhuys, K.A.G.; Lu, Y.; Zhou, W.; Cock, M.J.W.; Naranjo, S.E.; Fereti, A.; Williams, F.E.; Furlong, M.J. Ecological Pest Control Fortifies Agricultural Growth in Asia-Pacific Economies. Nat. Ecol. Evol. 2020, 4, 1522–1530. [Google Scholar] [CrossRef]
- Zeilinger, S.; Gupta, V.K.; Dahms, T.E.S.; Silva, R.N.; Singh, H.B.; Upadhyay, R.S.; Gomes, E.V.; Tsui, C.K.-M.; Nayak, S.C. Friends or Foes? Emerging Insights from Fungal Interactions with Plants. FEMS Microbiol. Rev. 2016, 40, 182–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuss, D.L. Hypovirulence: Mycoviruses at the Fungal-Plant Interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef]
- Mora, O.; Manuel, C. Control de Plagas En El Cultivo de Pepino Cucumis Sativus L. En El Cantón Yaguachi. Bachelor’s Thesis, Facultad de Ciencias Agrarias, Universidad de Guayaquil, Guayaquil, Ecuador, 2019. [Google Scholar]
- Shi, X.; Cordero, T.; Garrigues, S.; Marcos, J.F.; Daròs, J.; Coca, M. Efficient Production of Antifungal Proteins in Plants Using a New Transient Expression Vector Derived from Tobacco Mosaic Virus. Plant Biotechnol. J. 2019, 17, 1069–1080. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus Years of Fungal Viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Mu, F.; Xie, J.; Cheng, S.; You, M.P.; Barbetti, M.J.; Jia, J.; Wang, Q.; Cheng, J.; Fu, Y.; Chen, T.; et al. Virome Characterization of a Collection of S. sclerotiorum from Australia. Front. Microbiol. 2018, 8, 2540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic Reorganization of Family Partitiviridae and Other Recent Progress in Partitivirus Research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef]
- Domingo, E.; Holland, J.J. RNA Virus Mutations and Fitness for Survival. Annu. Rev. Microbiol. 1997, 51, 151–178. [Google Scholar] [CrossRef] [PubMed]
- Voth, P.D.; Mairura, L.; Lockhart, B.E.; May, G. Phylogeography of Ustilago Maydis Virus H1 in the USA and Mexico. J. Gen. Virol. 2006, 87, 3433–3441. [Google Scholar] [CrossRef]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of Filamentous Fungi and Their Relevance to Plant Pathology. Mol. Plant Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jiang, D. New Insights into Mycoviruses and Exploration for the Biological Control of Crop Fungal Diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [Green Version]
- van Diepeningen, A.; Debets, A.; Slakhorst, S.; Fekete, C.; Hornok, L.; Hoekstra, R. Interspecies Virus Transfer via Protoplast Fusions between Fusarium Poae and Black Aspergillus Strains. Fungal Genet. Rep. 2000, 47, 99–100. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s Conserved Domain Database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Eils, R.; Schlesner, M. Complex Heatmaps Reveal Patterns and Correlations in Multidimensional Genomic Data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Picarelli, M.A.S.C.; Forgia, M.; Rivas, E.B.; Nerva, L.; Chiapello, M.; Turina, M.; Colariccio, A. Extreme Diversity of Mycoviruses Present in Isolates of Rhizoctonia Solani AG2-2 LP From Zoysia Japonica From Brazil. Front. Cell. Infect. Microbiol. 2019, 9, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krupovic, M.; Ghabrial, S.A.; Jiang, D.; Varsani, A. Genomoviridae: A New Family of Widespread Single-Stranded DNA Viruses. Arch. Virol. 2016, 161, 2633–2643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, J.M.; Bonds, A.E.; Clemons, R.A.; Thapa, N.A.; Simmons, D.R.; Carter-House, D.; Ortanez, J.; Liu, P.; Miralles-Durán, A.; Desirò, A.; et al. Survey of Early-Diverging Lineages of Fungi Reveals Abundant and Diverse Mycoviruses. mBio 2020, 11, e02027-2. [Google Scholar] [CrossRef] [PubMed]
- Marzano, S.-Y.L.; Nelson, B.D.; Ajayi-Oyetunde, O.; Bradley, C.A.; Hughes, T.J.; Hartman, G.L.; Eastburn, D.M.; Domier, L.L. Identification of Diverse Mycoviruses through Metatranscriptomics Characterization of the Viromes of Five Major Fungal Plant Pathogens. J. Virol. 2016, 90, 6846–6863. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Hisano, S.; Tani, A.; Kondo, H.; Kanematsu, S.; Suzuki, N. A Capsidless SsRNA Virus Hosted by an Unrelated DsRNA Virus. Nat. Microbiol. 2016, 1, 15001. [Google Scholar] [CrossRef]
- Azhar, A.; Mu, F.; Huang, H.; Cheng, J.; Fu, Y.; Hamid, M.R.; Jiang, D.; Xie, J. A Novel RNA Virus Related to Sobemoviruses Confers Hypovirulence on the Phytopathogenic Fungus Sclerotinia sclerotiorum. Viruses 2019, 11, 759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamid, M.R.; Xie, J.; Wu, S.; Maria, S.K.; Zheng, D.; Assane Hamidou, A.; Wang, Q.; Cheng, J.; Fu, Y.; Jiang, D. A Novel Deltaflexivirus That Infects the Plant Fungal Pathogen, Sclerotinia sclerotiorum, Can Be Transmitted Among Host Vegetative Incompatible Strains. Viruses 2018, 10, 295. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Wu, S.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. Molecular Characterization of Two Positive-Strand RNA Viruses Co-Infecting a Hypovirulent Strain of Sclerotinia sclerotiorum. Virology 2014, 464–465, 450–459. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Fu, Y.; Guoqing, L.; Ghabrial, S.A. Viruses of the Plant Pathogenic Fungus Sclerotinia sclerotiorum. Adv. Virus Res. 2013, 86, 215–248. [Google Scholar] [CrossRef]
- Mu, F.; Li, B.; Cheng, S.; Jia, J.; Jiang, D.; Fu, Y.; Cheng, J.; Lin, Y.; Chen, T.; Xie, J. Nine viruses from eight lineages exhibiting new evolutionary modes that co-infect a hypovirulent phytopathogenic fungus. PLOS Pathog. 2021, 17, e1009823. [Google Scholar] [CrossRef]
- Ran, H.; Liu, L.; Li, B.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. Co-Infection of a Hypovirulent Isolate of Sclerotinia sclerotiorum with a New Botybirnavirus and a Strain of a Mitovirus. Virol. J. 2016, 13, 92. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Cheng, S.; Xiao, X.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Xie, J. Discovery of Two Mycoviruses by High-Throughput Sequencing and Assembly of Mycovirus-Derived Small Silencing RNAs From a Hypovirulent Strain of Sclerotinia sclerotiorum. Front. Microbiol. 2019, 10, 1415. [Google Scholar] [CrossRef] [PubMed]
- Chiba, S.; Suzuki, N. Highly Activated RNA Silencing via Strong Induction of Dicer by One Virus Can Interfere with the Replication of an Unrelated Virus. Proc. Natl. Acad. Sci. USA 2015, 112, E4911–E4918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, J.; Fu, Y.; Jiang, D.; Mu, F.; Cheng, J.; Lin, Y.; Li, B.; Marzano, S.-Y.L.; Xie, J. Interannual Dynamics, Diversity and Evolution of the Virome in Sclerotinia sclerotiorum from a Single Crop Field. Virus Evol. 2021, 7, veab032. [Google Scholar] [CrossRef]
- Kreuze, J.F.; Vaira, A.M.; Menzel, W.; Candresse, T.; Zavriev, S.K.; Hammond, J.; Hyun Ryu, K.; Report Consortium, I. ICTV Virus Taxonomy Profile: Alphaflexiviridae. J. Gen. Virol. 2020, 101, 699–700. [Google Scholar] [CrossRef]
- Martelli, G.P.; Sabanadzovic, S.; Abou-Ghanem Sabanadzovic, N.; Edwards, M.C.; Dreher, T. The Family Tymoviridae. Arch. Virol. 2002, 147, 1837–1846. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Blasdell, K.R.; Calisher, C.H.; Dietzgen, R.G.; Kondo, H.; Kurath, G.; Longdon, B.; Stone, D.M.; Tesh, R.B.; Tordo, N.; et al. ICTV Virus Taxonomy Profile: Rhabdoviridae. J. Gen. Virol. 2018, 99, 447–448. [Google Scholar] [CrossRef]
- Mateus, D.D.; Paredes, J.A.; Español, Y.; Ribas de Pouplana, L.; Moura, G.R.; Santos, M.A.S. Molecular Reconstruction of a Fungal Genetic Code Alteration. RNA Biol. 2013, 10, 968–980. [Google Scholar] [CrossRef] [Green Version]
- Crick, F.H.C. The Origin of the Genetic Code. J. Mol. Biol. 1968, 38, 367–379. [Google Scholar] [CrossRef]
- Luque, D.; Mata, C.P.; Suzuki, N.; Ghabrial, S.A.; Castón, J.R. Capsid Structure of DsRNA Fungal Viruses. Viruses 2018, 10, 481. [Google Scholar] [CrossRef] [Green Version]
- Hantula, J.; Mäkelä, S.; Xu, P.; Brusila, V.; Nuorteva, H.; Kashif, M.; Hyder, R.; Vainio, E.J. Multiple Virus Infections on Heterobasidion Sp. Fungal Biol. 2020, 124, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Kashif, M.; Hyder, R.; De Vega Perez, D.; Hantula, J.; Vainio, E.J. Heterobasidion Wood Decay Fungi Host Diverse and Globally Distributed Viruses Related to Helicobasidium Mompa Partitivirus V70. Virus Res. 2015, 195, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Bian, R.; Andika, I.B.; Pang, T.; Lian, Z.; Wei, S.; Niu, E.; Wu, Y.; Kondo, H.; Liu, X.; Sun, L. Facilitative and Synergistic Interactions between Fungal and Plant Viruses. Proc. Natl. Acad. Sci. USA 2020, 117, 3779–3788. [Google Scholar] [CrossRef]
- Vainio, E.J.; Jurvansuu, J.; Hyder, R.; Kashif, M.; Piri, T.; Tuomivirta, T.; Poimala, A.; Xu, P.; Mäkelä, S.; Nitisa, D.; et al. The Partitivirus HetPV13-An1 Mediates Severe Growth Debilitation and Major Alterations in the Gene Expression of a Fungal Forest Pathogen. J. Virol. 2017, 95, JVI.01744-17. [Google Scholar] [CrossRef] [Green Version]
- Vainio, E.J.; Hakanpää, J.; Dai, Y.-C.; Hansen, E.; Korhonen, K.; Hantula, J. Species of Heterobasidion Host a Diverse Pool of Partitiviruses with Global Distribution and Interspecies Transmission. Fungal Biol. 2011, 115, 1234–1243. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, A.; Urayama, S.; Suo, R.; Itoi, S.; Fuji, S.; Moriyama, H.; Hagiwara, D. Mycovirus-Induced Tenuazonic Acid Production in a Rice Blast Fungus Magnaporthe Oryzae. Front. Microbiol. 2020, 11, 1641. [Google Scholar] [CrossRef]
- Glass, N.L.; Dementhon, K. Non-Self Recognition and Programmed Cell Death in Filamentous Fungi. Curr. Opin. Microbiol. 2006, 9, 553–558. [Google Scholar] [CrossRef]
- Thapa, V.; Roossinck, M.J. Determinants of Coinfection in the Mycoviruses. Front. Cell. Infect. Microbiol. 2019, 9, 169. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Peisley, A.; Tetrault, D.; Li, Z.; Egelman, E.H.; Magor, K.E.; Walz, T.; Penczek, P.A.; Hur, S. Molecular Imprinting as a Signal-Activation Mechanism of the Viral RNA Sensor RIG-I. Mol. Cell 2014, 55, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal Effectors and Plant Susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef]
- Nagy, P.D. Yeast as a Model Host to Explore Plant Virus-Host Interactions. Annu. Rev. Phytopathol. 2008, 46, 217–242. [Google Scholar] [CrossRef] [Green Version]
- Andika, I.B.; Wei, S.; Cao, C.; Salaipeth, L.; Kondo, H.; Sun, L. Phytopathogenic Fungus Hosts a Plant Virus: A Naturally Occurring Cross-Kingdom Viral Infection. Proc. Natl. Acad. Sci. USA 2017, 114, 12267–12272. [Google Scholar] [CrossRef] [Green Version]
- Nerva, L.; Varese, G.C.; Falk, B.W.; Turina, M. Mycoviruses of an Endophytic Fungus Can Replicate in Plant Cells: Evolutionary Implications. Sci. Rep. 2017, 7, 1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Li, J.; Yu, B.; Liu, L.; Zhang, X.; Liu, J.; Pan, H.; Zhang, Y. Transcription Factor SsSte12 Was Involved in Mycelium Growth and Development in Sclerotinia sclerotiorum. Front. Microbiol. 2018, 9, 2476. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Li, S.J.; Yan, J.; Tang, Y.; Cheng, J.P.; Gao, A.J.; Yao, X.; Ruan, J.J.; Xu, B.L. Research Progress on Phytopathogenic Fungi and Their Role as Biocontrol Agents. Front. Microbiol. 2021, 12, 670135. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, Y.; Li, B.; Chen, T.; Tian, S. Reactive Oxygen Species: A Generalist in Regulating Development and Pathogenicity of Phytopathogenic Fungi. Comput. Struct. Biotechnol. J. 2020, 18, 3344–3349. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Qian, L.; Disi, J.O.; Yang, X.; Li, Q.; Li, J.; Frauen, M.; Cai, D.; Qian, W. Identification of Resistant Sources against Sclerotinia sclerotiorum in Brassica Species with Emphasis on B. oleracea. Euphytica 2011, 177, 393–399. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P.H.J.; Nelson, B.D. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and Molecular Traits of a Cosmopolitan Pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef]
- Abbas, A. A Review Paper on Mycoviruses. J. Plant Pathol. Microbiol. 2016, 7, 1000390. [Google Scholar] [CrossRef] [Green Version]
- Abid, M.; Khan, M.A.U.; Mushtaq, S.; Afzaal, S.; Haider, M.S. A Comprehensive review on mycoviruses as biological control agent. World J. Biol. Biotechnol. 2018, 3, 187–192. [Google Scholar] [CrossRef]
- Urayama, S.; Ohta, T.; Onozuka, N.; Sakoda, H.; Fukuhara, T.; Arie, T.; Teraoka, T.; Moriyama, H. Characterization of Magnaporthe Oryzae Chrysovirus 1 Structural Proteins and Their Expression in Saccharomyces Cerevisiae. J. Virol. 2012, 86, 8287–8295. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, K.B.; Holcomb, E.E.; Allscheid, R.L.; Carrington, J.C. Hiding in plain sight: New virus genomes discovered via a systematic analysis of fungal public transcriptomes. PLoS ONE 2019, 14, e0219207. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S.; Lin, Y.-H.; Kondo, H.; Kanematsu, S.; Suzuki, N. Effects of Defective Interfering RNA on Symptom Induction by, and Replication of, a Novel Partitivirus from a Phytopathogenic Fungus, Rosellinia Necatrix. J. Virol. 2013, 87, 2330–2341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukša, J.; Ravoitytė, B.; Konovalovas, A.; Aitmanaitė, L.; Butenko, A.; Yurchenko, V.; Serva, S.; Servienė, E. Different Metabolic Pathways Are Involved in Response of Saccharomyces Cerevisiae to L-A and M Viruses. Toxins 2017, 9, 233. [Google Scholar] [CrossRef] [Green Version]
- Filippou, C.; Garrido-Jurado, I.; Meyling, N.V.; Quesada-Moraga, E.; Coutts, R.H.A.; Kotta-Loizou, I. Mycoviral Population Dynamics in Spanish Isolates of the Entomopathogenic Fungus Beauveria bassiana. Viruses 2018, 10, 665. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S.; Lin, Y.-H.; Kondo, H.; Kanematsu, S.; Suzuki, N. A Novel Victorivirus from a Phytopathogenic Fungus, Rosellinia necatrix, Is Infectious as Particles and Targeted by RNA Silencing. J. Virol. 2013, 87, 6727–6738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, U.A.; Kotta-Loizou, I.; Fitt, B.D.L.; Coutts, R.H.A. Identification, Molecular Characterization, and Biology of a Novel Quadrivirus Infecting the Phytopathogenic Fungus Leptosphaeria biglobosa. Viruses 2018, 11, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magae, Y.; Sunagawa, M. Characterization of a Mycovirus Associated with the Brown Discoloration of Edible Mushroom, Flammulina velutipes. Virol. J. 2010, 7, 342. [Google Scholar] [CrossRef] [Green Version]
- Revill, P.A.; Davidson, A.D.; Wright, P.J. The Nucleotide Sequence and Genome Organization of Mushroom Bacilliform Virus: A Single-Stranded RNA Virus of Agaricus Bisporus (Lange) Imbach. Virology 1994, 202, 904–911. [Google Scholar] [CrossRef]
- Magae, Y. Molecular Characterization of a Novel Mycovirus in the Cultivated Mushroom, Lentinula Edodes. Virol. J. 2012, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Zhang, M.; Chen, Q.; Zhu, M.; Zhou, E. A Novel Mycovirus Closely Related to Viruses in the Genus Alphapartitivirus Confers Hypovirulence in the Phytopathogenic Fungus Rhizoctonia Solani. Virology 2014, 456–457, 220–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashif, M.; Jurvansuu, J.; Vainio, E.J.; Hantula, J. Alphapartitiviruses of Heterobasidion Wood Decay Fungi Affect Each Other’s Transmission and Host Growth. Front. Cell. Infect. Microbiol. 2019, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Özkan, S.; Mohorianu, I.; Xu, P.; Dalmay, T.; Coutts, R.H.A. Profile and Functional Analysis of Small RNAs Derived from Aspergillus Fumigatus Infected with Double-Stranded RNA Mycoviruses. BMC Genom. 2017, 18, 416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Trenas, A.; Prieto, P.; Cañizares, M.C.; García-Pedrajas, M.D.; Pérez-Artés, E. Mycovirus Fusarium Oxysporum f. Sp. Dianthi Virus 1 Decreases the Colonizing Efficiency of Its Fungal Host. Front. Cell. Infect. Microbiol. 2019, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Jiang, J.; Wang, Y.; Hong, N.; Zhang, F.; Xu, W.; Wang, G. Hypovirulence of the Phytopathogenic Fungus Botryosphaeria Dothidea: Association with a Coinfecting Chrysovirus and a Partitivirus. J. Virol. 2014, 88, 7517–7527. [Google Scholar] [CrossRef] [Green Version]
- Suryanarayanan, T.S. Endophyte Research: Going beyond Isolation and Metabolite Documentation. Fungal Ecol. 2013, 6, 561–568. [Google Scholar] [CrossRef]
- Zhai, L.; Zhang, M.; Hong, N.; Xiao, F.; Fu, M.; Xiang, J.; Wang, G. Identification and Characterization of a Novel Hepta-Segmented DsRNA Virus From the Phytopathogenic Fungus Colletotrichum Fructicola. Front. Microbiol. 2018, 9, 754. [Google Scholar] [CrossRef] [Green Version]
- Harmsen, M.C.; Tolner, B.; Kram, A.; Go, S.J.; de Haan, A.; Wessels, J.G. Sequences of Three DsRNAs Associated with La France Disease of the Cultivated Mushroom (Agaricus bisporus). Curr. Genet. 1991, 20, 137–144. [Google Scholar] [CrossRef]
- Aoki, N.; Moriyama, H.; Kodama, M.; Arie, T.; Teraoka, T.; Fukuhara, T. A Novel Mycovirus Associated with Four Double-Stranded RNAs Affects Host Fungal Growth in Alternaria alternata. Virus Res. 2009, 140, 179–187. [Google Scholar] [CrossRef]
- Chun, J.; Ko, Y.-H.; Kim, D.-H. Transcriptome Analysis of Cryphonectria parasitica Infected With Cryphonectria hypovirus 1 (CHV1) Reveals Distinct Genes Related to Fungal Metabolites, Virulence, Antiviral RNA-Silencing, and Their Regulation. Front. Microbiol. 2020, 11, 1711. [Google Scholar] [CrossRef]
- Rigling, D.; Prospero, S. Cryphonectria Parasitica, the Causal Agent of Chestnut Blight: Invasion History, Population Biology and Disease Control. Mol. Plant Pathol. 2017, 19, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Eusebio-Cope, A.; Suzuki, N. Mycoreovirus Genome Rearrangements Associated with RNA Silencing Deficiency. Nucleic Acids Res. 2015, 43, 3802–3813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bologna, N.G.; Voinnet, O. The Diversity, Biogenesis, and Activities of Endogenous Silencing Small RNAs in Arabidopsis. Annu. Rev. Plant Biol. 2014, 65, 473–503. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Feng, F.; Zhou, Q.; Zhao, F.; Du, R.; Zhou, Z.; Han, Y. Isolation, Purification and Characterization of Exopolysaccharide Produced by Leuconostoc pseudomesenteroides YF32 from Soybean Paste. Int. J. Biol. Macromol. 2018, 114, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Osaki, H.; Nakamura, H.; Sasaki, A.; Matsumoto, N.; Yoshida, K. An Endornavirus from a Hypovirulent Strain of the Violet Root Rot Fungus, Helicobasidium mompa. Virus Res. 2006, 118, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Jiāng, D.; Ayllón, M.A.; Marzano, S.-Y.L. ICTV Report ConsortiumYR 2019 ICTV Virus Taxonomy Profile: Mymonaviridae. J. Gen. Virol. 2019, 100, 1343–1344. [Google Scholar] [CrossRef]
- Sato, Y.; Miyazaki, N.; Kanematsu, S.; Xie, J.; Ghabrial, S.A.; Hillman, B.I.; Suzuki, N. ICTV Report ConsortiumYR 2019 ICTV Virus Taxonomy Profile: Megabirnaviridae. J. Gen. Virol. 2019, 100, 1269–1270. [Google Scholar] [CrossRef]
- Kim, J.-M.; Jung, J.-E.; Park, J.-A.; Park, S.-M.; Cha, B.-J.; Kim, D.-H. Biological Function of a Novel Chrysovirus, CnV1-BS122, in the Korean Cryphonectria nitschkei BS122 Strain. J. Biosci. Bioeng. 2013, 115, 1–3. [Google Scholar] [CrossRef]
- Petrzik, K.; Koloniuk, I.; Sehadová, H.; Sarkisova, T. Chrysoviruses Inhabited Symbiotic Fungi of Lichens. Viruses 2019, 11, 1120. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Liu, C.; Jiang, X.; Tan, Z.; Li, H.; Xu, S.; Zhang, S.; Shang, Q.; Deising, H.B.; Behrens, S.-E.; et al. The Newly Identified Trichoderma Harzianum partitivirus (ThPV2) Does Not Diminish Spore Production and Biocontrol Activity of Its Host. Viruses 2022, 14, 1532. [Google Scholar] [CrossRef]
- Sahin, E.; Akata, I. Viruses Infecting Macrofungi. VirusDisease 2018, 29, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Milgroom, M.G.; Hillman, B.I. The Ecology and Evolution of Fungal Viruses. Stud. Viral Ecol. Microb. Bot. Host Syst. 2011, 1, 217–253. [Google Scholar] [CrossRef]
- Pasin, F.; Menzel, W.; Daròs, J. Harnessed Viruses in the Age of Metagenomics and Synthetic Biology: An Update on Infectious Clone Assembly and Biotechnologies of Plant Viruses. Plant Biotechnol. J. 2019, 17, 1010–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decker, C.J.; Steiner, H.R.; Hoon-Hanks, L.L.; Morrison, J.H.; Haist, K.C.; Stabell, A.C.; Poeschla, E.M.; Morrison, T.E.; Stenglein, M.D.; Sawyer, S.L.; et al. DsRNA-Seq: Identification of Viral Infection by Purifying and Sequencing DsRNA. Viruses 2019, 11, 943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urayama, S.; Sakoda, H.; Takai, R.; Katoh, Y.; Minh Le, T.; Fukuhara, T.; Arie, T.; Teraoka, T.; Moriyama, H. A DsRNA Mycovirus, Magnaporthe oryzae chrysovirus 1-B, Suppresses Vegetative Growth and Development of the Rice Blast Fungus. Virology 2014, 448, 265–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urayama, S.-I.; Takaki, Y.; Nunoura, T. FLDS: A Comprehensive DsRNA Sequencing Method for Intracellular RNA Virus Surveillance. Microbes Environ. 2016, 31, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Klenner, J.; Kohl, C.; Dabrowski, P.W.; Nitsche, A. Comparing Viral Metagenomic Extraction Methods. Curr. Issues Mol. Biol. 2017, 24, 59–70. [Google Scholar] [CrossRef]
- Schmitt, M.J.; Breinig, F. The Viral Killer System in Yeast: From Molecular Biology to Application. FEMS Microbiol. Rev. 2002, 26, 257–276. [Google Scholar] [CrossRef]
- Rodríguez-Cousiño, N.; Gómez, P.; Esteban, R. L-A-Lus, a New Variant of the L-A Totivirus Found in Wine Yeasts with Klus Killer Toxin-Encoding Mlus Double-Stranded RNA: Possible Role of Killer Toxin-Encoding Satellite RNAs in the Evolution of Their Helper Viruses. Appl. Environ. Microbiol. 2013, 79, 4661–4674. [Google Scholar] [CrossRef] [Green Version]
- García-Pedrajas, M.D.; Cañizares, M.C.; Sarmiento-Villamil, J.L.; Jacquat, A.G.; Dambolena, J.S. Mycoviruses in Biological Control: From Basic Research to Field Implementation. Phytopathology 2019, 109, 1828–1839. [Google Scholar] [CrossRef]
- Dawe, A.L.; Nuss, D.L. Hypovirus Molecular Biology: From Koch’s Postulates to Host Self-Recognition Genes That Restrict Virus Transmission. Adv. Virus Res. 2013, 86, 109–147. [Google Scholar] [CrossRef]
- Chen, Y.-S.; Wu, H.-C.; Liu, C.-H.; Chen, H.-C.; Yanagida, F. Isolation and Characterization of Lactic Acid Bacteria from Jiang-Sun (Fermented Bamboo Shoots), a Traditional Fermented Food in Taiwan. J. Sci. Food Agric. 2010, 90, 1977–1982. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.N.; Bailey, A.M. Viruses of Botrytis. Adv. Virus Res. 2013, 86, 249–272. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, C.S.; da Fonseca, R.R.; Batista, T.M.; Barreto, M.A.; Argolo, C.S.; Carvalho, M.R.D.; Amaral, D.O.J.D.; Silva, E.M.D.A.; Arévalo-Gardini, E.; Hidalgo, K.S.; et al. Genome Sequence and Effectorome of Moniliophthora perniciosa and Moniliophthora roreri subpopulations. BMC Genom. 2018, 19, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, B.A.; Evans, H.C.; Phillips-Mora, W.; Ali, S.S.; Meinhardt, L.W. Moniliophthora Roreri, Causal Agent of Cacao Frosty Pod Rot. Mol. Plant Pathol. 2018, 19, 1580–1594. [Google Scholar] [CrossRef] [Green Version]
- Loguercio, L.L.; de Carvalho, A.C.; Niella, G.R.; De Souza, J.T.; Pomella, A.W.V. Selection of Trichoderma Stromaticum Isolates for Efficient Biological Control of Witches’ Broom Disease in Cacao. Biol. Control 2009, 51, 130–139. [Google Scholar] [CrossRef]
- Loguercio, L.L.; Santos, L.S.; Niella, G.R.; Miranda, R.A.C.; de Souza, J.T.; Collins, R.T.; Pomella, A.W.V. Canopy-Microclimate Effects on the Antagonism between Trichoderma stromaticum and Moniliophthora perniciosa in Shaded Cacao. Plant Pathol. 2009, 58, 1104–1115. [Google Scholar] [CrossRef]
- Liu, C.; Li, M.; Redda, E.T.; Mei, J.; Zhang, J.; Wu, B.; Jiang, X. A Novel Double-Stranded RNA Mycovirus Isolated from Trichoderma harzianum. Virol. J. 2019, 16, 113. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villan Larios, D.C.; Diaz Reyes, B.M.; Pirovani, C.P.; Loguercio, L.L.; Santos, V.C.; Góes-Neto, A.; Fonseca, P.L.C.; Aguiar, E.R.G.R. Exploring the Mycovirus Universe: Identification, Diversity, and Biotechnological Applications. J. Fungi 2023, 9, 361. https://doi.org/10.3390/jof9030361
Villan Larios DC, Diaz Reyes BM, Pirovani CP, Loguercio LL, Santos VC, Góes-Neto A, Fonseca PLC, Aguiar ERGR. Exploring the Mycovirus Universe: Identification, Diversity, and Biotechnological Applications. Journal of Fungi. 2023; 9(3):361. https://doi.org/10.3390/jof9030361
Chicago/Turabian StyleVillan Larios, Diana Carolina, Brayan Maudiel Diaz Reyes, Carlos Priminho Pirovani, Leandro Lopes Loguercio, Vinícius Castro Santos, Aristóteles Góes-Neto, Paula Luize Camargos Fonseca, and Eric Roberto Guimarães Rocha Aguiar. 2023. "Exploring the Mycovirus Universe: Identification, Diversity, and Biotechnological Applications" Journal of Fungi 9, no. 3: 361. https://doi.org/10.3390/jof9030361
APA StyleVillan Larios, D. C., Diaz Reyes, B. M., Pirovani, C. P., Loguercio, L. L., Santos, V. C., Góes-Neto, A., Fonseca, P. L. C., & Aguiar, E. R. G. R. (2023). Exploring the Mycovirus Universe: Identification, Diversity, and Biotechnological Applications. Journal of Fungi, 9(3), 361. https://doi.org/10.3390/jof9030361