

Biocontrol of Diseases Caused by Phytophthora capsici and P. parasitica in Pepper Plants

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
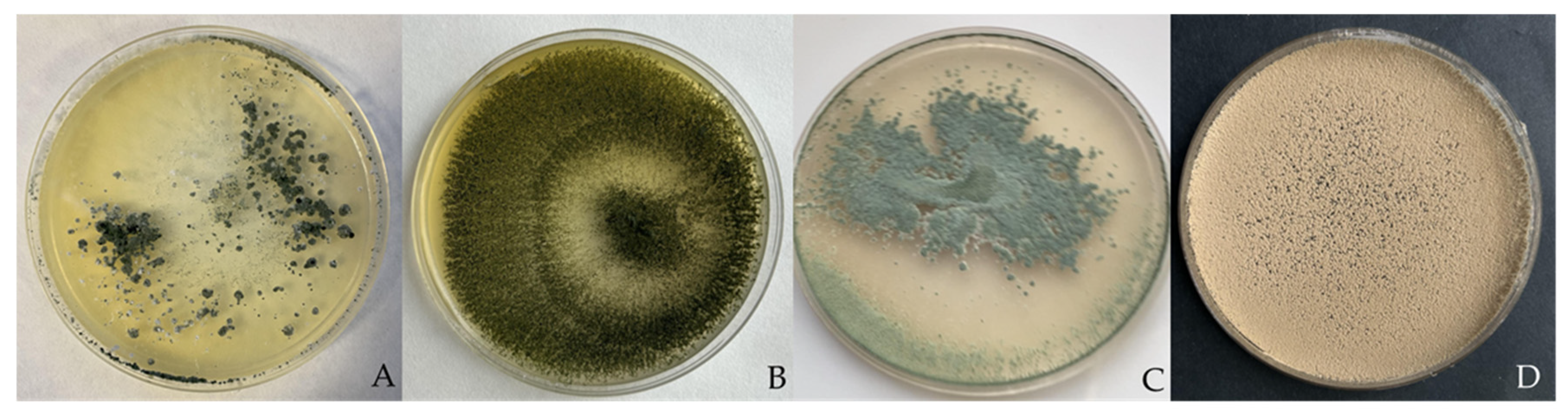
2.1. Fungal Isolates
2.2. Dual Culture Bioassays
2.3. Antifungal Volatile and Non-Volatile Metabolite Bioassay
2.4. Effects of TA, TS, TL, and PAE Isolates on the Severity of Phytophthora Blight in Pepper
2.5. Effect of Antagonists on the Chlorophyll Content of Peppers
2.6. RNA Extraction and Real-Time Polymerase Chain Reaction (RT-PCR)
2.7. Statistical Analysis
3. Results
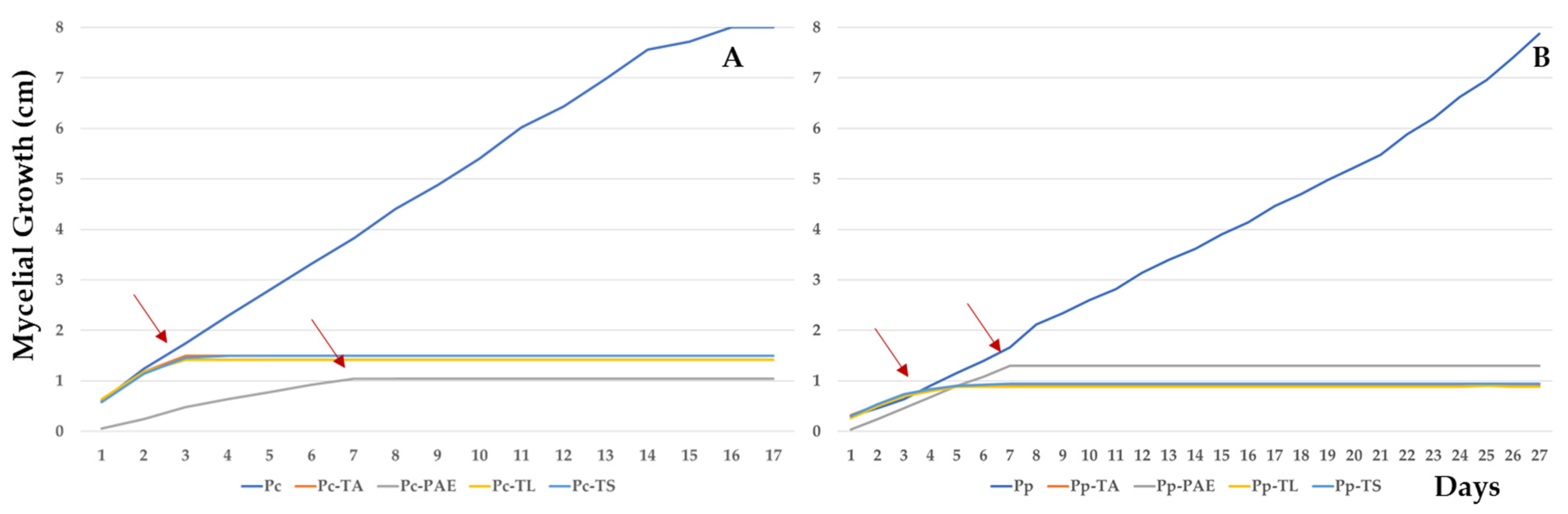
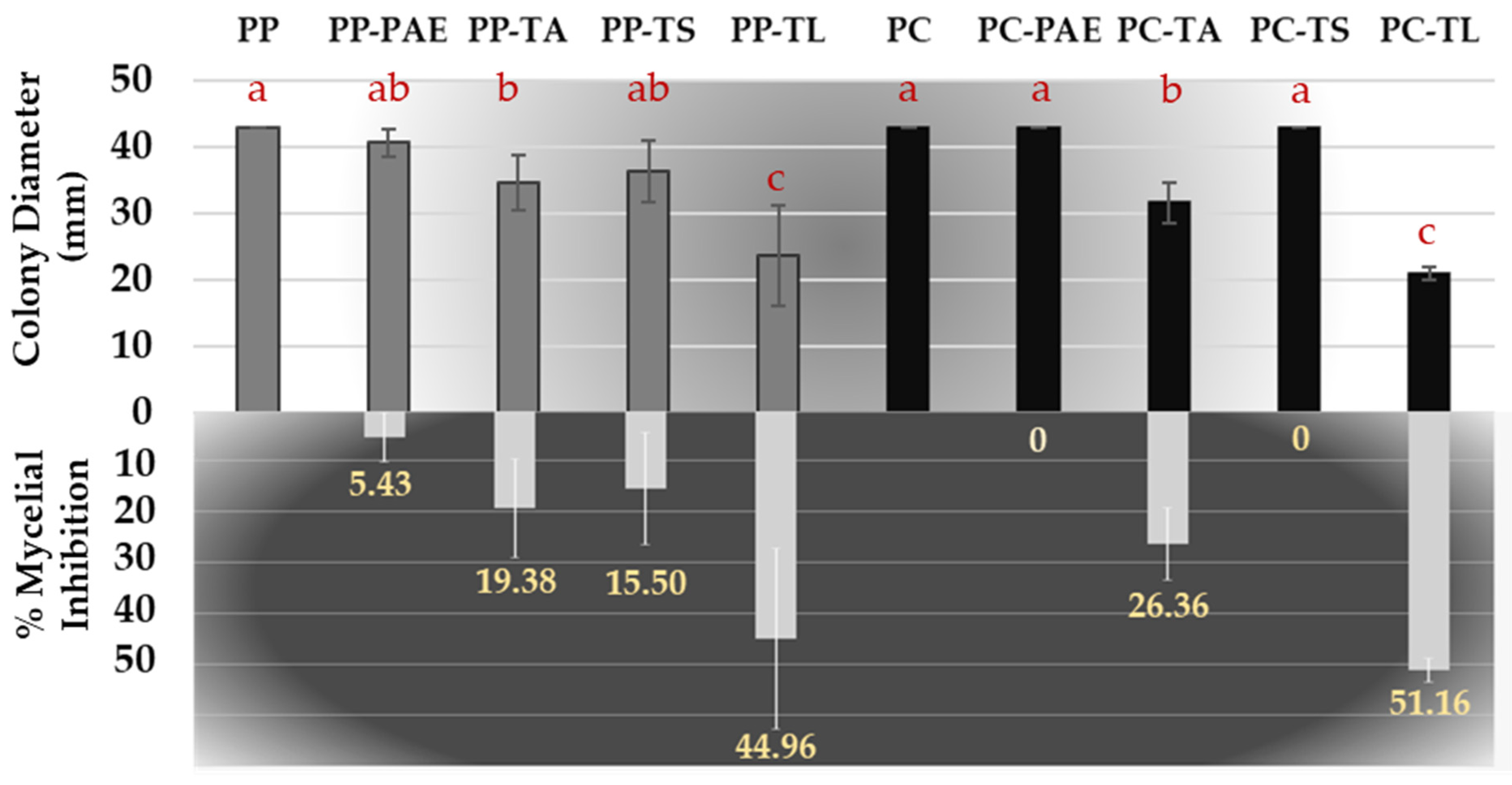
3.1. Dual Culture Bioassays
3.2. Antifungal Volatile and Non-Volatile Metabolite Bioassay
3.3. Effects of TA, TS, TL, and PAE Isolates on the Severity of Phytophthora Blight in Pepper
3.4. Effect of Antagonists on the Chlorophyll Content of Peppers
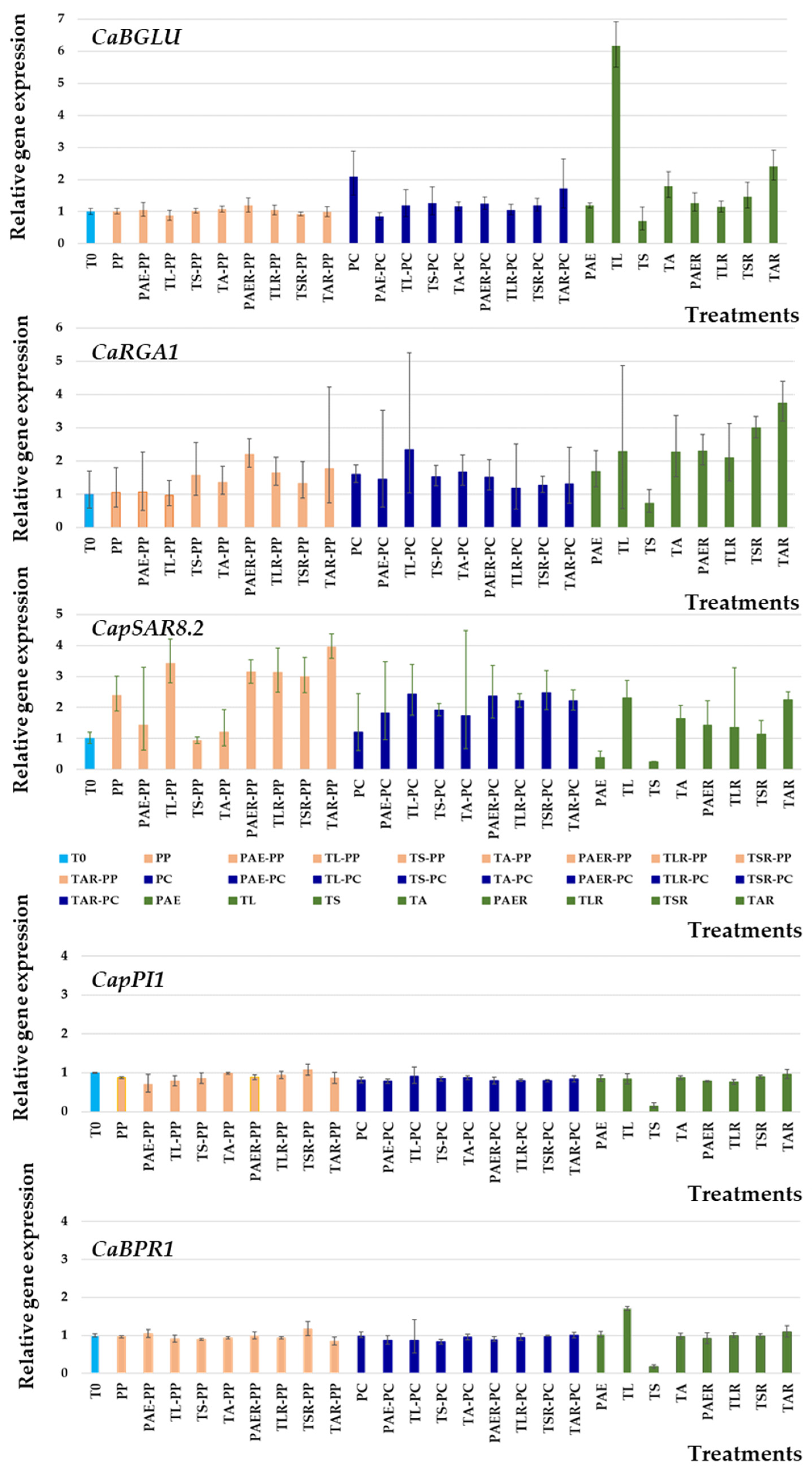
3.5. Molecular Responses of Pepper Leaves
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beakes, G.; Sekimoto, S. The evolutionary phylogeny of Oomycetes–insights gained from studies of holocarpic parasites of algae and invertebrates. In Oomycete Genetics and Genomics: Diversity, Interactions, and Research Tools; Lamour, K., Kamoun, S., Eds.; Wiley: New York, NY, USA, 2009; pp. 1–24. [Google Scholar] [CrossRef] [Green Version]
- Jayawardena, R.S.; Hyde, K.D.; Chen, Y.J.; Stadler, M.; Wang, Y. One stop shop IV: Taxonomic update with molecular phylogeny for important phytopathogenic genera. Fungal Divers. 2020, 103, 87–218. [Google Scholar] [CrossRef]
- Mitchell, D.J. Relationships of inoculum levels of several soilborne species of Phytophthora and Pythium to infection of severalhosts. Phytopathology 1978, 68, 1754–1759. [Google Scholar] [CrossRef]
- Bala, K.; Robideau, G.P.; Lévesque, C.A.; de Cock, A.; Abad, Z.; Lodhi, A.; ShahzadS, S.; Ghaffar, A.; Coffey, M. Persoonia—Molecular Phylogeny and Evolution of Fungi. Fungal Planet 2010, 24, 136–137. [Google Scholar] [CrossRef]
- Ho, H.H. The taxonomy and biology of Phytophthora and Pythium. J. Bacteriol. Mycol. 2018, 6, 40–45. [Google Scholar] [CrossRef] [Green Version]
- Tkaczyk, M. Phytopythium: Origin, differences and meaning in modern plant pathology. Folia For. Pol. A 2020, 62, 227–232. [Google Scholar] [CrossRef]
- de Cock, A.W.A.M.; Lodhi, A.M.; Rintoul, T.L.; Bala, K.; Robideau, G.P.; Abad, G.Z.; Coffey, M.D.; Shahzad, S.; Lévesque, C.A. Phytopythium: Molecular phylogeny and systematics. Persoonia 2015, 34, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C.M.; Cooke, D.E.; Duncan, J.M. Origin of a new Phytophthora pathogen through interspecific hybridization. Proc. Natl. Acad. Sci. USA 1999, 96, 5878–5883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goheen, E.M.; Frankel, S.J. Phytophthoras in forests and natural ecosystems. In Proceedings of the Fourth Meeting of the International Union of Forest Research Organizations (IUFRO) Working Party S07.02.09, Monterey, CA, USA, 27 September 2021. [Google Scholar]
- Martin, F.N.; Abad, Z.G.; Balci, Y.; Ivors, K. Identification and detection of Phytophthora: Reviewing our progress, identifying our needs. Plant Dis. 2012, 96, 1080–1103. [Google Scholar] [CrossRef] [Green Version]
- Božik, M.; Mrázková, M.; Novotná, K.; Klouček, P.; Černý, K. MALDI-TOF MS as a method for rapid identification of Phytophthora de Bary, 1876. Peer. J. 2021, 9, e11662. [Google Scholar] [CrossRef]
- Yang, X.; Tyler, B.M.; Hong, C. An expanded phylogeny for the genus Phytophthora. IMA Fungus 2017, 8, 355–384. [Google Scholar] [CrossRef] [Green Version]
- Blair, J.E.; Coffey, M.D.; Park, S.Y.; Geiser, D.M.; Kang, S. A multi-locus phylogeny for Phytophthora utilizing markers derived from complete genome sequences. Fungal Genet. Biol. 2008, 45, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Goss, E.M.; Cardenas, M.E.; Myers, K. The plant pathogen Phytophthora andina emerged via hybridization of an unknown Phytophthora species and the Irish potato famine pathogen, P. infestans. PLoS ONE 2011, 6, e24543. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C.; Scanu, B.; Cooke, D.; Jung, T. Phytophthora: An ancient, historic, biologically and structurally cohesive and evolutionarily successful generic concept in need of preservation. IMA Fungus 2022, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Richardson, P.A.; Hong, C. Phytophthora × stagnum notho sp. Nov., a new hybrid from irrigation reservoirs at ornamental plant nurseries in Virginia. PLoS ONE 2014, 9, e103450. [Google Scholar] [CrossRef]
- Brasier, C.M.; Kirk, S.A.; Delcan, J.; Cooke, D.E.L.; Jung, T.; Manin’t Veld, W.A. Phytophthora alni sp. nov. and its variants: Designation of emerging heteroploid hybrid pathogens spreading on Alnus trees. Mycol. Res. 2004, 108, 1172–1184. [Google Scholar] [CrossRef] [PubMed]
- Burgess, T.I. Molecular characterization of natural hybrids formed between five related indigenous clade 6 Phytophthora species. PLoS ONE 2015, 10, e0134225. [Google Scholar] [CrossRef] [Green Version]
- Van Poucke, K.; Haegeman, A.; Goedefroit, T.; Ruttink, T.; Heungens, K. Unravelling hybridization in Phytophthora using phylogenomics and genome size estimation. IMA Fungus 2021, 12, 1–24. [Google Scholar] [CrossRef]
- Rodríguez-Molina, M.C.; Morales-Rodríguez, M.C.; Palo Osorio, C.; Duarte Maya, M.S.; Picón-Toro, J. Phytophthora nicotianae, the causal agent of root and crown rot (Tristeza disease) of red pepper in La Vera region (Cáceres, Spain). Span. J. Agri. Res. 2010, 8, 770–774. [Google Scholar] [CrossRef] [Green Version]
- Lacasa, C.M.; Martínez, V.; Hernández, A.; Serrano-Pérez, P.; Larregla, S. Survival reduction of Phytophthora capsici oospores and P. nicotianae chlamydospores with Brassica green manures combined with solarization. Sci. Hortic. 2015, 197, 607–618. [Google Scholar] [CrossRef]
- Alcantara, T.P.; Bosland, P.W. An inexpensive disease screening technique for foliar blight of chile pepper seedlings. HortScience 1994, 29, 1182–1183. [Google Scholar] [CrossRef] [Green Version]
- Lamour, K.H.; Stam, R.; Jupe, J. The oomycete broad-host-range pathogen Phytophthora capsici. Mol. Plant Pathol. 2012, 13, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, Y.; Li, L. Infection of Phytophthora capsici on pepper-Models and affecting factors. Front. Agri. China 2008, 2, 66–71. [Google Scholar] [CrossRef]
- Café-Filho, A.C.; Duniway, J.M. Effect of location of drip irrigation emitters and position of Phytophthora capsici infections in roots on Phytophthora root rot of pepper. Phytopathology 1996, 86, 1364–1369. [Google Scholar]
- Hausbeck, M.K.; Lamour, K.H. Phytophthora capsici on vegetable crops: Research progress and management challenges. Plant Dis. 2004, 88, 1292–1303. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Regulation (EC) no. 1005/2009 of the European Parliament and of the council of 16 September 2009 on substances that deplete the ozone layer (recast). Off. J. Eur. Union 2009, 286, 1–30. [Google Scholar]
- Ploetz, R.C.; Heine, G.; Haynes, J.L.; Watson, M. Investigating biological attributes that may contribute to Phytophthora capsici importance as a vegetable pathogen in Florida. Ann. Appl. Biol. 2002, 140, 61–67. [Google Scholar] [CrossRef]
- Parra, G.; Ristaino, J.B. Resistance to mefenoxam and metalaxyl among field isolates of Phytophthora capsici causing Phytophthora blight of bell pepper. Plant Dis. 2001, 85, 1069–1075. [Google Scholar] [CrossRef] [Green Version]
- Gisbert, C.; Sánchez-Torres, P.; Raigón, M.D.; Nuez, F. Phytophthora capsici resistance evaluation in pepper hybrids: Agronomic performance and fruit quality of pepper grafted plants. J. Agric. Food Inf. 2010, 8, 116–121. [Google Scholar] [CrossRef]
- Ros, C.; Lacasa, C.M.; Martínez, V.; Bielza, P.; Lacasa, A. Response of pepper rootstocks to co-infection of Meloidogyne incognita and Phytophthora spp. Eur. J. Hortic. Sci. 2014, 79, 22–28. [Google Scholar]
- López-Marín, J.; Galvez, A.; Porras, I.; Brotons-Martínez, J.M. Pepper grafting (Capsicum annuum): Benefits and profitability. ITEA 2016, 112, 127–146. [Google Scholar]
- Guigón-López, C. Differential development of wilt and stem rot diseases in grafted bell pepper (Capsicum annuum, L.). Eur. J. Plant Pathol. 2019, 154, 347–357. [Google Scholar] [CrossRef]
- Kousik, C.S.; Ikerd, J.L.; Hassell, R. Grafting onto resistant rootstocks for managing Phytophthora crown rot of peppers. Acta Hort. 2021, 13, 163–168. [Google Scholar] [CrossRef]
- Guerrero, M.M.; Ros, C.; Guirao, P.; Martínez, M.A.; Martínez, M.C.; Barceló, N. Biofumigation plus solarisation efficacy for soil disinfestation in sweet pepper greenhouses in the Southeast of Spain. Acta Hort. 2005, 698, 293–297. [Google Scholar] [CrossRef]
- Guerrero, M.M.; Ros, C.; Lacasa, C.; Martínez, V.; Lacasa, A.; Fernández, P. Effect of biosolarization using pellets of Brassica carinata on soilborne pathogens in protected pepper crops. Acta Hort. 2010, 883, 337–344. [Google Scholar] [CrossRef]
- Butler, D.M.; Kokalis-Burelle, N.; Muramoto, J.; McCollum, T.G.; Rosskopf, E.N. Impact of anaerobic soil disinfestation combined with soil solarization on plant-parasitic nematodes and introduced inoculum of soilborne plant pathogens in raised-bed vegetable production. Crop. Prot. 2012, 39, 33–40. [Google Scholar] [CrossRef]
- Núñez-Zofío, M.; Larregla, S.; Garbisu, C. Repeated biodisinfection controls the incidence of Phytophthora root and crown rot of pepper while improving soil quality. Span. J. Agric. Res. 2012, 10, 794–805. [Google Scholar] [CrossRef]
- Larregla, S.; Núñez-Zofío, M.; Fernández-Molina, P.; Lacasa-Martinez, C.M.; Guerrero-Díaz, M.M. Reduction of Phytophthora capsici oospores viability by gases released during soil biosolarization of protected pepper crops in southeastern Spain. Acta Hort. 2014, 1044, 113–118. [Google Scholar] [CrossRef]
- Rodríguez-Molina, M.C.; Serrano-Pérez, P.; Palo, C. Effect of biofumigation with brassica pellets combined with Brassicaceae cover crops and plastic cover on the survival and infectivity of inoculum of Phytophthora nicotianae Breda de Haan. Pest Manag. Sci. 2016, 72, 1295–1301. [Google Scholar] [CrossRef]
- Gandariasbeitia, M.; Ojinaga, M.; Orbegozo, E.; Mendarte, S.; Larregla, S. Winter biodisinfestation with Brassica green manure is a promising management strategy for Phytophthora capsici control of protected pepper crops in humid temperate climate regions of northern Spain. Span. J. Agric. Res. 2019, 17, 1005. [Google Scholar] [CrossRef]
- Larregla, S.; Gandariasbeitia, M.; Ojinaga, M.; Guerrero, M.D.M.; Ortiz-Barredo, A. Gases Released During Soil Biodisinfestation of Pepper Greenhouses Reduce Survival of Phytophthora capsici Oospores in Northern Spain. Front. Sustain. Food Syst. 2021, 5, 663–915. [Google Scholar] [CrossRef]
- Guerrero, M.D.M.; Lacasa, C.M.; Martínez, V.; Monserrat, A.; Larregla, S. Low Temperature Biodisinfection Effectiveness for Phytophthora capsici Control of Protected Sweet Pepper Crops in the Southeast of Spain. Front. Sustain. Food Syst. 2021, 5, 659290. [Google Scholar] [CrossRef]
- Babadoost, M.; Pavón, C. Survival of Oospores of Phytophthora capsici in Soil. Plant Dis. 2013, 97, 1478–1483. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Zhang, D.; Ren, L.; Wang, Q.; Cao, A. Bio-activation of soil with beneficial microbes after soil fumigation reduces soil-borne pathogens and increases tomato yield. Environ. Pollut. 2021, 283, 117–160. [Google Scholar] [CrossRef] [PubMed]
- Lorito, M.; Farkas, V.; Rebuffat, S.; Bodo, B.; Kubicek, C.P. Cell wall synthesis is a major target of mycoparasitic antagonism by Trichoderma harzianum. J. Bacteriol. 1996, 178, 6382–6385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, M.; Diánez, F. Los antagonistas microbianos en el manejo de micosis de la parte aérea de la planta. In Organismos Para el Control de Patógenos en los Cultivos Protegidos. Prácticas Culturales Para una Agricultura Sostenible, 2nd ed.; Tello, J.C., Camacho, F., Eds.; Fundación Cajamar: Almería, España, 2010; pp. 523–528. [Google Scholar]
- Wang, Q.; Ma, Y.; Wang, G.; Gu, Z.; Sun, D.; Andyz, X.; Chang, Z. Integration of biofumigation with antagonistic microorganism can control Phytophthora blight of pepper plants by regulating soil bacterial community structure. Eur. J. Soil Biol. 2014, 61, 58–67. [Google Scholar] [CrossRef]
- Galletti, S.; Sala, E.; Leoni, O.; Burzi, P.L.; Cerato, C. Trichoderma spp. tolerance to Brassica carinata seed meal for a combined use in biofumigation. Biol. Cont. 2008, 45, 319–327. [Google Scholar] [CrossRef]
- Joo, G.J. Production of an anti-fungal substance for biological control of Phytophthora capsici causing phytophthora blight in red-peppers by Streptomyces halstedii. Biotechnol. Lett. 2005, 27, 201–205. [Google Scholar] [CrossRef]
- Nguyen, X.H.; Naing, K.W.; Lee, Y.S.; Kim, Y.H.; Moon, J.H.; Kim, K.Y. Antagonism of antifungal metabolites from Streptomyces griseus H7602 against Phytophthora capsici. J. Basic Microbiol. 2015, 5, 45–53. [Google Scholar] [CrossRef]
- Nguyen, X.H.; Naing, K.W.; Lee, Y.S.; Tindwa, H.; Lee, G.H.; Jeong, B.K.; Kim, K.Y. Biocontrol potential of Streptomyces griseus H7602 against root rot disease (Phytophthora capsici) in pepper. Plant Pathol. J. 2012, 28, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Sakineh, A.; Ayme, S.; Akram, S.; Naser, S. Streptomyces strains modulate dynamics of soil bacterial communities and their efficacy in disease suppression caused by Phytophthora capsici. Sci. Rep. 2021, 11, 9317. [Google Scholar] [CrossRef]
- Trinidad-Cruz, J.R.; Rincón-Enríquez, G.; Evangelista-Martínez, Z.; Quiñones-Aguilar, E.E. Biorational control of Phytophthora capsici in pepper plants using Streptomyces spp. Rev. Chapingo Ser. Hort. 2021, 27, 85–99. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Chen, P.C.; Tsay, T.T. The biocontrol efficacy and antibiotic activity of Streptomyces plicatus on the oomycete Phytophthora capsici. Biol. Cont. 2016, 98, 34–42. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, S.W.; Li, K.T. Antagonistic activity and mechanism of an isolated Streptomyces corchorusii stain AUH-1 against phytopathogenic fungi. World J. Microbiol. Biotechnol. 2019, 35, 145. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, S.; Safaie, N.; Sadeghi, A.; Shamsbakhsh, M. Tissue-specific synergistic bio-priming of pepper by two Streptomyces species against Phytophthora capsici. PLoS ONE 2020, 15, 230–531. [Google Scholar] [CrossRef] [Green Version]
- Sid, A.; Ezziyyani, M.; Egea-Gilabert, C.; Candela, M.E. Selecting bacterial strains for use in the biocontrol of diseases caused by Phytophthora capsici and Alternaria alternata in sweet pepper plants. Biol. Plant 2003, 47, 569–574. [Google Scholar] [CrossRef]
- Jiang, Z.; Guo, Y.; Li, S.; Qi, H.; Guo, J. Evaluation of biocontrol efficiency of different Bacillus preparations and field application methods against Phytophthora blight of bell pepper. Biol Cont. 2006, 36, 216–223. [Google Scholar] [CrossRef]
- Zhang, S.; White, T.L.; Martinez, M.C.; Mcinroy, J.A.; Kloepper, J.W.; Klassen, W. Evaluation of plant growth-promoting rhizobacteria for control of Phytophthora blight on squash under greenhouse conditions. Biol Cont. 2010, 53, 129–135. [Google Scholar] [CrossRef]
- Akgül, D.S.; Mirik, M. Biocontrol of Phytophthora capsici on pepper plants by Bacillus megaterium strains. J. Plant Pathol. 2008, 90, 29–34. [Google Scholar] [CrossRef]
- Lim, J.H.; Sang, D.K. Biocontrol of Phytophthora blight of red pepper caused by Phytophthora capsici using Bacillus subtilis AH18 and B. licheniformis K11 formulations. J. Korean Soc. Appl. Biol. Chem. 2010, 53, 766–773. [Google Scholar] [CrossRef]
- Yau, J.A.; Diánez, F.; Marín, F.; Carretero, C.; Santos, M. Screening and evaluation of potential biocontrol fungi and bacteria foliar endophytes against Phytophthora capsici and Phytophthora parasitica on pepper. J. Food Agric. Environ. 2013, 11, 490–495. [Google Scholar]
- Li, Y.; Feng, X.; Wang, X.; Zheng, L.; Liu, H. Inhibitory effects of Bacillus licheniformis BL06 on Phytophthora capsici in pepper by multiple modes of action. Biol. Cont. 2020, 144, 104–210. [Google Scholar] [CrossRef]
- Bhusal, B.; Mmbaga, M.T. Biological control of Phytophthora blight and growth promotion in sweet pepper by Bacillus species. Biol. Cont. 2020, 150, 104–373. [Google Scholar] [CrossRef]
- Naing, K.W.; Anees, M.; Nguyen, X.H.; Lee, Y.S.; Jeon, S.W.; Kim, S.J.; Kim, M.H.; Kim, K.Y. Biocontrol of late blight disease (Phytophthora capsici) of pepper and the plant growth promotion by Paenibacillus ehimensis KWN38. J. Phytopathol. 2014, 162, 367–376. [Google Scholar] [CrossRef]
- Mei, X.L.; Zhao, Q.Y.; Tan, S.Y.; Shen, B.; Shen, Q.R. Screening, identification, and biocontrol effect of antagonistic bacteria against Phytophthora capsici. Chin. J. Appl. Ecol. 2010, 21, 2652–2658. [Google Scholar]
- Gilardi, G.; Vasileiadou, A.; Garibaldi, A.; Gullino, M.L. Biocontrol agents and resistance inducers reduce Phytophthora crown rot (Phytophthora capsici) of sweet pepper in closed soilless culture. Phytopathol. Mediterr. 2021, 60, 149–163. [Google Scholar] [CrossRef]
- Plett, J.M. Order of microbial succession affects rhizobia-mediated biocontrol efforts against Phytophthora root rot. Microbiol. Res. 2021, 242, 126–628. [Google Scholar] [CrossRef]
- Okamoto, H.; Sato, M.; Sato, Z.; Isaka, M. Biocontrol of Phytophthora capsici by Serratia marcescens F-1-1 and analysis of biocontrol mechanisms using transposon-insertion mutants. Ann. Phytopathol. Soc. Jpn. 1998, 64, 287–293. [Google Scholar] [CrossRef]
- Segarra, G.; Avilés, M.; Casanova, E.; Borrero, A.; Trillas, I. Effectiveness of biological control of Phytophthora capsici in pepper by Trichoderma asperellum strain T34. Phytopathol. Mediterr. 2013, 52, 77–83. [Google Scholar] [CrossRef]
- Diánez, F.; Santos, M.; Carretero, F.; Marín, F. Trichoderma saturnisporum, a new biological control agent. J. Sci. Food Agric. 2016, 96, 1934–1944. [Google Scholar] [CrossRef]
- Chemeltorit, P.P.; Mutaqin, K.H.; Widodo, W. Combining Trichoderma hamatum THSW13 and Pseudomonas aeruginosa BJ10–86: A synergistic chili pepper seed treatment for Phytophthora capsici infested soil. Eur. J. Plant Pathol. 2017, 147, 157–166. [Google Scholar] [CrossRef]
- de Oliveira, T.A.S.; Blum, L.E.B.; Duarte, E.A.A.; Luz, E.D.M.N. Control of Phytophthora palmivora on postharvest papaya with Trichoderma asperellum, T. virens, T. harzianum and T. longibrachiatum. Bioscience 2018, 34, 1513–1521. [Google Scholar] [CrossRef] [Green Version]
- Tomah, A.A.; Abd Alamer, I.S.; Li, B.; Zhang, J.Z. A new species of Trichoderma and gliotoxin role: A new observation in enhancing biocontrol potential of T. virens against Phytophthora capsici on chili pepper. Biol. Cont. 2020, 145, 104–261. [Google Scholar] [CrossRef]
- Gilardi, G.; Pugliese, M.; Gullino, M.L.; Garibaldi, A. Effect of biocontrol agents and potassium phosphite against Phytophthora crown rot, caused by Phytophthora capsici, on zucchini in a closed soilless system. Sci. Hortic. 2020, 265, 109–207. [Google Scholar] [CrossRef]
- La Spada, F.; Stracquadanio, C.; Riolo, M.; Pane, A.; Cacciola, S.O. Trichoderma counteracts the challenge of Phytophthora nicotianae cnfections on tomato by modulating plant defense mechanisms and the expression of crinkler, necrosis-inducing Phytophthora Protein 1, and cellulose-Binding Elicitor Lectin Pathogenic Effectors. Front. Plant Sci. 2020, 11, 539–583. [Google Scholar] [CrossRef]
- Kang, S.W.; Kim, S.W. New antifungal activity of penicillic acid against Phytophthora species. Biotechnol. Lett. 2004, 26, 695–698. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; Ali, G.S. Aspergillus flavipes is a novel efficient biocontrol agent of Phytophthora parasitica. Biol. Cont. 2020, 140, 72–104. [Google Scholar] [CrossRef]
- Wang, H.; Shi, J.; Luo, Z.; Chen, Q.; Wang, C. Isolation and identification of Penicillium strain QMYCS-2 and its effect on tobacco black shank Tobacco. Sci. Technol. Stud. 2020, 49, 2663. [Google Scholar] [CrossRef]
- Mondol, M.A.M.; Farthouse, J.; Islam, M.T.; Schüffler, A.; Laatsch, H. Metabolites from the Endophytic Fungus Curvularia sp. M12 Act as Motility Inhibitors against Phytophthora capsici zoospores. J. Nat. Prod. 2017, 80, 347–355. [Google Scholar] [CrossRef]
- Chen, J.T.; Su, H.J.; Huang, J.W. Isolation and identification of secondary metabolites of Clitocybe nuda responsible for inhibition of zoospore germination of Phytophthora capsici. J. Agric. Food Chem. 2012, 60, 7341–7344. [Google Scholar] [CrossRef]
- Santos, M.; Diánez, F.; Moreno-Gavíra, A.; Sánchez-Montesinos, B.; Gea, F.J. Cladobotryum mycophilum as potential biocontrol agent. Agronomy 2019, 9, 891. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.; Jamshaid, M.; Zahid, M.A.; Vetukuri, R.R.; Stenberg, J.A. Biological control of strawberry crown rot, root rot and grey mould by the beneficial fungus Aureobasidium pullulans. Bio. Cont. 2021, 66, 535–545. [Google Scholar] [CrossRef]
- Safitri, D.; Wiyono, S.; Soekarno, B.P.W. Mode of action of the endophytic yeast Rhodotorula mucilaginosa in controlling basal stem rot caused by Phytopthora capsici. IOP Conf. Ser. Earth Environ. Sci. 2021, 667, 12–50. [Google Scholar] [CrossRef]
- Mercier, D.C. Manker Biocontrol of soil-borne diseases and plant growth enhancement in greenhouse soilless mix by the volatile producing fungus Muscodor albus. Crop. Prot. 2005, 24, 355–362. [Google Scholar] [CrossRef]
- Pozo, M.J.; Cordier, C.; Dumas-Gaudot, E.; Barea, J.M.; Azcón-Aguilar, C. Localized versus systemic effect of arbuscular mycorrhizal fungi on defence responses to Phytophthora infection in tomato plants. J. Exp. Bot. 2002, 53, 525–534. [Google Scholar] [CrossRef]
- Hu, J.; Hou, S.; Li, M.; Wu, F.; Lin, X. The better suppression of pepper Phytophthora blight by arbuscular mycorrhizal (AM) fungus than Purpureocillium lilacinum alone or combined with AM fungus. J. Soils Sed. 2020, 20, 792–800. [Google Scholar] [CrossRef]
- González-Hernández, A.I.; Suárez-Fernández, M.B.; Pérez-Sánchez, R.; Gómez-Sánchez, M.Á.; Morales-Corts, M.R. Compost tea induces growth and resistance against Rhizoctonia solani and Phytophthora capsici in pepper. Agronomy 2021, 11, 781. [Google Scholar] [CrossRef]
- Manasfi, Y.; Cannesan, M.A.; Riah, W.; Vicré, M.; Trinsoutrot-Gattin, I. Potential of combined biological control agents to cope with Phytophthora parasitica, a major pathogen of Choisya ternata. Eur. J. Plant Pathol. 2018, 152, 1011–1025. [Google Scholar] [CrossRef]
- Marín, F.; Diánez, F.; Santos, M.; Navarro, M.J.; Yau, J.A. Control of Phytophthora capsici and Phytophthora parasitica on pepper (Capsicum annuum L.) with compost teas from different sources, and their effects on plant growth promotion. Phytopathol. Med. 2014, 53, 216–228. [Google Scholar] [CrossRef]
- Diánez, F.; Santos, M.; Tello, J.C. Suppressive effects of grape marc compost on phytopathogenic oomycetes. Arch. Phytopathol. Pflanzenschutz 2007, 40, 1–18. [Google Scholar] [CrossRef]
- Fontana, D.C.; de Paula, S.; Torres, A.G.; Schmidt, D.; Neto, D.D. Endophytic fungi: Biological control and induced resistance to phytopathogens and abiotic stresses. Pathogens 2021, 10, 570. [Google Scholar] [CrossRef]
- Yyani, M.E.; Requena, C.; Egea-Gilabert, M.E. Candela biological control of Phytophthora root rot of pepper using Trichoderma harzianum and Streptomyces rochei in combination. J. Phytopathol. 2007, 155, 342–349. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, L.; Zhang, J.Z.; Ojaghian, M.R.; Hyde, K.D. Antagonistic interaction between Trichoderma asperellum and Phytophthora capsici in vitro. J. Zhejiang Univ. Sci. B 2016, 17, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Trotta, A.; Varese, G.C.; Gnavi, E.; Fusconi, A.; Sampò, S.; Berta, G. Interactions between the soilborne root pathogen Phytophthora nicotianae var.parasitica and the arbuscular mycorrhizal fungus Glomus mosseae in tomato plants. Plant Soil 1996, 185, 199–209. [Google Scholar] [CrossRef]
- Phoka, N.; Suwannarach, N.; Lumyong, S.; Ito, S.; Matsui, K.; Arikit, S.; Sunpapao, A. Role of volatiles from the endophytic fungus Trichoderma asperelloides PSU-P1 in biocontrol potential and in promoting the plant growth of Arabidopsis thaliana. J. Fungi 2020, 6, 341. [Google Scholar] [CrossRef]
- Jin, J.H.; Zhang, H.X.; Tan, J.Y.; Yan, M.J.; Li, D.W.; Khan, A. A new ethylene-responsive factor CaPTI1 gene of pepper (Capsicum annuum L.) involved in the regulation of defense response to Phytophthora capsici. Front. Plant Sci. 2015, 6, 1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.C.; Hwang, B.K. Induction of some defense-related genes and oxidative burst is required for the establishment of systemic acquired resistance in Capsicum annuum. Planta 2005, 221, 790–800. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Akbar, A.; Iqrar, I. A glucanolytic Pseudomonas sp. associated with Smilax bona-nox L. displays strong activity against Phytophthora parasitica. Microbiol. Res. 2018, 207, 140–152. [Google Scholar] [CrossRef]
- Shobha, M.S.; Mahadeva, M.S. Effect of Endophytic and plant growth promoting rhizobacteria against foot rot disease of Piper nigrum L. Int. J. Environ. Agric. Biotechnol. 2018, 3, 6. [Google Scholar] [CrossRef]
- Spence, C.; Alff, E.; Johnson, C.; Ramos, C.; Donofrio, N.; Sundaresan, V.; Bais, H. Natural rice rhizospheric microbes suppress rice blast infections. BMC Plant Biol. 2014, 14, 130. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhou, J.M. Plant immunity triggered by microbial molecular signatures. Mol. Plant 2010, 3, 783–793. [Google Scholar] [CrossRef] [Green Version]
- Saijo, Y.; Loo, E.P.; Yasuda, S. Pattern recognition receptors and signaling in plant–microbe interactions. Plant J. 2018, 93, 592–613. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Ai, G.; Xia, C.; Pan, W.; Yin, Z.; Dou, D. Different requirement of immunity pathway components by oomycete effectors-induced cell death. Phytopathol. Res. 2022, 4, 4. [Google Scholar] [CrossRef]
- Dou, D.L.; Zhou, J.M. Phytopathogen effectors subverting host immunity: Different foes, similar battleground. Cell Host Microbe 2012, 12, 484–495. [Google Scholar] [CrossRef] [Green Version]
- Silvar, C.; Merino, F.; Díaz, J. Resistance in pepper plants induced by Fusarium oxysporum f. sp. lycopersici involves different defence-related genes. Plant Biol. 2009, 11, 68–74. [Google Scholar] [CrossRef]
- Stamler, R.A.; Holguin, O.; Dungan, B.; Goldberg, N.; Randall, J.J. BABA and Phytophthora nicotianae induce resistance to Phytophthora capsici in chile pepper (Capsicum annuum). PLoS ONE 2015, 10, e0128327. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Zamioudis, C.B.R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced systemic resistance by beneficial microbes. Ann. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Savitha, M.J.; Sriram, S. Induced systemic resistance (ISR) in hot pepper against Phytophthora capsici infection triggered by cell wall oligosaccharide elicitors from Trichoderma species. Indian J. Hortic. 2017, 74, 233–239. [Google Scholar] [CrossRef]
- Sriram, S.; Manasa, S.B.; Savitha, M.J. Potential use of elicitors from Trichoderma in induced systemic resistance for the management of Phytophthora capsici in red pepper. J. Biol. Cont. 2009, 23, 449–456. [Google Scholar] [CrossRef]
- Umadevi, P.; Anandaraj, M. Proteomic analysis of the tripartite interaction between black pepper, Trichoderma harzianum and Phytophthora capsici provides insights into induced systemic resistance mediated by Trichoderma spp. Eur. J. Plant Pathol. 2019, 154, 607–620. [Google Scholar] [CrossRef]
- Bellini, A.; Pugliese, M.; Guarnaccia, V.; Meloni, G.R.; Gullino, L.M. Calcium oxide, potassium phosphite and a Trichoderma enriched compost water suspension protect Capsicum annuum against Phytophthora capsici by priming the immune system. Pest Manag. Sci. 2021, 77, 3484–3490. [Google Scholar] [CrossRef] [PubMed]
- Chowdappa, P.; Mohan Kumar, S.P.; Jyothi Lakshmi, M.; Upreti, K.K. Growth stimulation and induction of systemic resistance in tomato against early and late blight by Bacillus subtilis OTPB1 or Trichoderma harzianum OTPB3. Biol. Cont. 2013, 65, 109–117. [Google Scholar] [CrossRef]
- Qiu, Y.; Yan, H.H.; Sun, S.M.; Wang, Y.Q.; Zhao, X.R.; Wang, H.Y. Use of Bacillus velezensis SDTB022 against tobacco black shank (TBS) and the biochemical mechanism involved. Biol. Cont. 2022, 165, 104–785. [Google Scholar] [CrossRef]
- Yarullina, L.G.; Sorokan, A.V.; Tsvetkov, V.O.; Burkhanova, G.F.; Kalatskaja, J.N. Influence of the genus Bacillus bacteria on the content of H2O2 and the activity of hydrolases and their inhibitors in potato plants during Phytophthora infestans Mont. de Bary infection. BIO Web Conf. 2020, 23, 02010. [Google Scholar] [CrossRef]
- Sorokan, A.; Benkovskaya, G.; Burkhanova, G.; Blagova, D.; Maksimov, I. Endophytic strain Bacillus subtilis 26DCryChS producing Cry1Ia toxin from Bacillus thuringiensis promotes multifaceted potato defense against Phytophthora infestans (Mont.) de Bary and pest Leptinotarsa decemlineata Say. Plants 2020, 9, 1115. [Google Scholar] [CrossRef]
- Park, J.W.; Balaraju, K.; Kim, J.W.; Lee, S.W.; Park, K. Systemic resistance and growth promotion of chili pepper induced by an antibiotic producing Bacillus vallismortis strain BS07. Biol. Cont. 2013, 65, 246–257. [Google Scholar] [CrossRef]
- Lee, B.D.; Dutta, S.; Ryu, H.; Suh, D.S.; Park, K. Induction of systemic resistance in panax ginseng against Phytophthora cactorum by native Bacillus amyloliquefaciens HK34. J. Ginseng Res. 2015, 39, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.M.; Hamayun, M.; Waqas, M.; Shinwari, Z.K.; Lee, I.J. Burkholderia sp. KCTC 11096BP modulates pepper growth and resistance against Phytophthora capsici. Pak. J. Bot. 2016, 48, 1965–1970. [Google Scholar]
- Sang, M.K.; Kim, J.G.; Kim, K.D. Biocontrol activity and induction of systemic resistance in pepper by compost water extracts against Phytophthora capsici. Phytopathology 2010, 100, 774–783. [Google Scholar] [CrossRef] [Green Version]
- Ferrusquía-Jiménez, N.I.; Serrano-Jamaica, L.M.; Martínez-Camacho, J.E.; Sáenz de la O, D.; Villagomez-Aranda, A.L.; González-Chavira, M.M.; Guerrero-Aguilar, B.Z.; Torres-Pacheco, I.; Feregrino-Pérez, A.A.; Medina-Ramos, G.; et al. Extracellular self-DNA plays a role as a damage-associated molecular pattern (DAMP) delaying zoospore germination rate and inducing stress-related responses in Phytophthora capsici. Plant Pathol. 2022, 71, 1066–1075. [Google Scholar] [CrossRef]
- Analisis de la Campaña Hortofruticola. Campaña 2020/21. Cajamar. Available online: https://publicacionescajamar.es/publicacionescajamar/public/pdf/series-tematicas/informes-coyuntura-analisis-de-campana/informe-71-campana-almeria-20-21.pdf (accessed on 10 March 2023).
- Sánchez-Montesinos, B.; Diánez, F.; Moreno-Gavíra, A.; Gea, F.J.; Santos, M. Plant growth promotion and biocontrol of Pythium ultimum by saline tolerant Trichoderma isolates under salinity stress. Int. J. Environ. Res. Public Health 2019, 16, 2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Montesinos, B.; Santos, M.; Moreno-Gavíra, A.; Marín-Rodulfo, T.; Gea, F.J.; Diánez, F. Biological control of fungal diseases by Trichoderma aggressivum f. europaeum and its compatibility with fungicides. J. Fungi 2021, 7, 598. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Gavíra, A.; Diánez, F.; Sánchez-Montesinos, B.; Santos, M. Paecilomyces variotii as a plant-growth promoter in horticulture. Agronomy 2020, 10, 597. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.; Dianez, F.; Gonzalez del Valle, M.; Tello, J.C. Grape marc compost: Microbial studies and suppression of soil-borne mycosis in vegetable seedlings. World J. Microbiol. Biotechnol. 2008, 24, 1493–1505. [Google Scholar] [CrossRef]
- Diánez, F.; Boix, A.; de Cara, M.; Trillas, I.; Avilés, M.; Tello, J.C. Grape marc compost tea suppressiveness to plant pathogenic fungi: Role of siderophores. Compost. Sci. Util. 2006, 14, 48–53. [Google Scholar] [CrossRef]
- Sánchez-Montesinos, B.; Diánez, F.; Moreno-Gavira, A.; Gea, F.J.; Santos, M. Role of Trichoderma aggressivum f. europaeum as plant-growth promoter in horticulture. Agronomy 2020, 10, 1004. [Google Scholar] [CrossRef]
- Ling, Q.; Huang, W.; Jarvis, P. Use of a SPAD-502 meter to measure leaf chlorophyll concentration in Arabidopsis thaliana. Photosyn. Res. 2011, 107, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wang, J.E.; Li, D.W.; Zhang, Y.L.; Zhao, Q.; He, Y.M.; Gong, Z.H. Defence responses of pepper (Capsicum annuum L.) infected with incompatible and compatible strains of Phytophthora capsici. Eur. J. Plant Pathol. 2013, 136, 625–638. [Google Scholar] [CrossRef]
- Gayoso, C.; Martinez de ilarduya, O.; Pomar, F.; Merino, F. Assessment of real-time PCR as a method for determining the presence of Verticillium dahliae in different Solanaceae cultivars. Eur. J. Plant Pathol. 2007, 118, 199–209. [Google Scholar] [CrossRef]
- Lee, S.C.; Hwang, B.K. Identification of the pepper SAR8.2 gene as a molecular marker for pathogen infection, abiotic elicitors and environmental stresses in Capsicum annuum. Planta 2003, 216, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Pang, X.; Wan, H.; Ahammed, G.J.; Yu, J.; Yao, Z.; Ruan, M.; Ye, Q.; Li, Z.; Wang, R.; et al. Identification of optimal reference genes for normalization of qPCR analysis during pepper fruit development. Front. Plant Sci. 2017, 8, 1128. [Google Scholar] [CrossRef] [Green Version]
- Volynchikova, E.; Kim, K.D. Biological control of oomycete soilborne diseases caused by Phytophthora capsici, Phytophthora infestans, and Phytophthora nicotianae in Solanaceous crops. Mycobiology 2022, 50, 269–293. [Google Scholar] [CrossRef]
- Ren, Y.; Armstrong, M.; Qi, Y.; Mclellan, H.; Zhong, C.; Du, B. Phytophthora infestans RXLR effectors target parallel steps in an immune signal transduction pathway. Plant Physiol. 2019, 180, 2227–2239. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Gavira, A.; Huertas, V.; Diánez, F.; Sánchez-Montesinos, B.; Santos, M. Paecilomyces and its importance in the biological control of agricultural pests and diseases. Plants 2020, 9, 1746. [Google Scholar] [CrossRef]
- Bononi, L.; Chiaramonte, J.B.; Pansa, C.C.; Moitinho, M.A.; Melo, I.S. Phosphorus-solubilizing Trichoderma spp. from Amazon soils improve soybean plant growth. Sci. Rep. 2020, 10, 2858. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.A.A.; Najeeb, S.; Hussain, S.; Xie, B.; Li, Y. Bioactive secondary metabolites from Trichoderma spp. against phytopathogenic fungi. Microorganisms 2020, 8, 817. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; He, P.; He, P.; Tian, Y.; He, Y. Potential biocontrol efficiency of Trichoderma species against oomycete pathogens. Front. Microbiol. 2022, 13, 974024. [Google Scholar] [CrossRef] [PubMed]
- Ros, M.; Raut, I.; Santisima-Trinidad, A.B.; Pascual, J.A. Relationship of microbial communities and suppressiveness of Trichoderma fortified composts for pepper seedlings infected by Phytophthora nicotianae. PLoS ONE 2017, 12, e0174069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, A.K.; Singh, N.; Singh, D. Evaluation of the bioformulation of potent native strains of Trichoderma spp. against the foot rot/gummosis of Kinnow mandarin. Egypt J. Biol. Pest Cont. 2021, 31, 90. [Google Scholar] [CrossRef]
- Roberts, D.P.; Maul, J.E.; McKenna, L.F.; Emche, S.E.; Meyer, S.L.F.; Collins, R.T.; Bowers, J.H. Selection of genetically diverse Trichoderma spp. isolates for suppression of Phytophthora capsici on bell pepper. Can. J. Microbiol. 2010, 56, 864–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezziyyani, M.; Pérez Sánchez, C.; Sid Ahmed, A.; Requena, M.E.; Candela, M.E. Trichoderma harzianum como biofungicida para el biocontrol de Phytophthora capsici en plantas de pimiento (Capsicum annuum L.). Anal. Biol. 2004, 26, 35–45. [Google Scholar]
- Li, M.F.; Li, G.H.; Zhang, K.Q. Non-volatile metabolites from Trichoderma spp. Metabolites 2019, 9, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Yap, M.; Behringer, G.; Hung, R.; Joan, W.B. Volatile organic compounds emitted by Trichoderma species mediate plant growth. Fungal Biol. Biotechnol. 2016, 3, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.Q.; Kuo, X.; Liu, X.M.; Zhang, P. A systematic review on secondary metabolites of paecilomyces species: Chemical diversity and biological activity. Planta Med. 2020, 86, 805–821. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.J.; Mohanta, T.K.; Chung, J.Y.; Ryu, M.; Park, G.; Shim, S.; Hong, S.B.; Seo, H.; Bae, D.W.; Bae, I.; et al. Trichoderma metabolites as biological control agents against Phytophthora pathogens. Biol. Cont. 2016, 92, 128–138. [Google Scholar] [CrossRef]
- Dai, Z.B.; Wang, X.; Li, G.H. Secondary metabolites and their bioactivities produced by Paecilomyces. Molecules 2020, 25, 5077. [Google Scholar] [CrossRef]
- Moreno-Gavíra, A.; Diánez, F.; Sánchez-Montesinos, B.; Santos, M. Biocontrol effects of Paecilomyces variotii against fungal plant diseases. J. Fungi 2021, 7, 415. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Jung, H.W.; Hwang, B.K. Pepper gene encoding a basic b-1,3-glucanase is differentially expressed in pepper tissues upon pathogen infection and ethephon or methyl jasmonate treatment. Plant Sci. 2000, 156, 23–34. [Google Scholar] [CrossRef]
- Kim, Y.J.; Hwang, B.K. Pepper gene encoding a basic pathogenesis-related 1 protein is pathogen and ethylene inducible. Physiol. Plantarum 2000, 108, 51–60. [Google Scholar] [CrossRef]
- Kang, D.S.; Min, K.J.; Kwak, A.M.; Lee, S.Y.; Kang, H.W. Defense response and suppression of Phytophthora blight disease of pepper by water extract from spent mushroom substrate of Lentinula edodes. Plant Pathol. J. 2017, 33, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, N.; Giménez, M.J.; Torres, A.M.; Atienza, S.G.; Avila, C.M.; Palomino, C. Up-regulation of resistance gene analogs (RGA) in chickpea in the early response to Fusarium wilt. Euphytica 2012, 186, 793–804. [Google Scholar] [CrossRef]
- Silvar, C.; Merino, F.; Díaz, J. Differential activation of defense-related genes in susceptible and resistant pepper cultivars infected with Phytophthora capsici. J. Plant Physiol. 2008, 165, 1120–1124. [Google Scholar] [CrossRef]
- Sarowar, S.; Kim, Y.J.; Kim, E.N. Overexpression of a pepper basic pathogenesis-related protein 1 gene in tobacco plants enhances resistance to heavy metal and pathogen stresses. Plant Cell Rep. 2005, 24, 216–224. [Google Scholar] [CrossRef]
- Shi, R.; Jin, J.; Nifong, J.M.; Shew, D.; Lewis, R.S. Homoeologous chromosome exchange explains the creation of a QTL affecting soil-borne pathogen resistance in tobacco. Plant Biotechnol. J. 2022, 20, 47–58. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primers | Sequences (5′→3′) | Reference |
|---|---|---|---|
| CaBGLU | CABGLU-F | ACAGGCACATCTTCACTTACC | [107] |
| CABGLU-R | CGAGCAAAGGCGAATTTATCC | ||
| CaRGA1 | CARGA-F | ATGAGAAGGGAATAGGACGAG | [133] |
| CARGA-R | ACATCCAATGGCAGGAAACT | ||
| CaBPR1 | CaBPR1-F | GTTGTGCTAGGGTTCGGTGT | [99] |
| CaBPR1-R | CAAGCAATTATTTAAACGATCCA | ||
| CaPTI1 | CapPI1-F | TTTGAAACGGCCGAAGAAGC | [98,134] |
| CapPI1-R | TGCACGATTCTGTCTTAGCGT | ||
| CaSAR8.2 | CapSAR8.2-F | TGTTGCCAGGGAGATGACTTC | [135] |
| CapSAR8.2-R | ACAACGGCCATGACAAGTTT | ||
| ACT | ACT-F | TGTTATGGTAGGGATGGGTC | [136] |
| ACT-R | TTCTCTCTATTTGCCTTGGG |
| Colony Diameter (cm) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Phytophthora capsici | Phytophthora parasitica | |||||||
| Incubation Time (days) | ||||||||
| Treatments | 7 | 15 | 21 | 30 | 7 | 15 | 21 | 30 |
| Control | 2.7 ± 0 a* | 2.68 ± 0.06 a | 2.7 ± 0 a | 2.7 ± 0 a | 2.7 ± 0.00 a | 2.69 ± 0.03 a | 2.69 ± 0.08 a | 2.7 ± 0.00 a |
| PAE 5% | 2.65 ± 0.00 a | 2.68 ± 0.06 a | 2.66 ± 0.07 ab | 2.68 ± 0.06 a | 2.51 ± 0.25 cd | 2.54 ± 0.22 bcd | 2.54 ± 0.22 bcd | 2.62 ± 0.11 b |
| PAE 10% | 2.61 ± 0.10 ab | 2.61 ± 0.18 ab | 2.61 ± 0.18 abc | 2.61 ± 0.18 bc | 2.62 ± 0.09 b | 2.62 ± 0.09 ab | 2.60 ± 0.10 abc | 2.59 ± 0.09 b |
| PAE 15% | 2.66 ± 0.18 a | 2.62 ± 0.15 ab | 2.63 ± 0.15 abcd | 2.63 ± 0.15 ab | 2.58 ± 0.08 bc | 2.57 ± 0.08 bc | 2.52 ± 0.08 bcd | 2.58 ± 0.08 |
| TA 5% | 2.5 ± 0.14 cd | 2.43 ± 0.1 cd | 2.53 ± 0.19 cde | 2.60 ± 0.09 bc | 2.58 ± 0.08 bc | 2.45 ± 0.07 de | 2.61 ± 0.04 ab | 2.58 ± 0.05 bcd |
| TA 10% | 2.51 ± 0.07 cd | 2.46 ± 0.07 cd | 2.62 ± 0.07 bcde | 2.58 ± 0.06 bcd | 2.51 ± 0.12 cd | 2.43 ± 0.11 e | 2.52 0.0 bcd | 2.58 ± 0.06 bc |
| TA 15% | 2.55 ± 0.09 bc | 2.54 ± 0.05 bc | 2.52 ± 0.04 de | 2.56 ± 0.05 bcd | 2.54 ± 0.05 bcd | 2.56 ± 0.05 bcd | 2.49 ± 0.06 cd | 2.52 ± 0.06 bcd |
| TS 5% | 2.54 ± 0.09 bc | 2.51 ± 0.08 bcd | 2.54 ± 0.16 cde | 2.52 ± 0.07 d | 2.7 ± 0.00 a | 2.65 ± 0.07 ab | 2.43 ± 0.06 d | 2.48 ± 0.1 e |
| TS 10% | 2.47 ± 0.07 cd | 2.39 ± 0.16 de | 2.45 ± 0.08 ef | 2.43 ± 0.07 e | 2.57 ± 0.05 bc | 2.45 ± 0.13 de | 2.46 ± 0.05 d | 2.48 ± 0.06 e |
| TS 15% | 2.34 ± 0.08 e | 2.41 ± 0.09 de | 2.48 ± 0.11 ef | 2.43 ± 0.07 e | 2.52 ± 0.07 cd | 2.46 ± 0.19 cde | 2.48 ± 0.11 d | 2.45 ± 0.05 ef |
| TL 5% | 2.45 ± 0.13 cd | 2.58 ± 0.14 de | 2.5 ± 0.12 de | 2.55 ± 0.08 cd | 2.57 ± 0.05 bc | 2.54 ± 0.10 bcd | 2.43 ± 0.06 d | 2.51 ± 0.07 de |
| TL 10% | 2.43 ± 0.08 de | 2.31 ± 0.25 ef | 2.46 ± 0.13 ef | 2.43 ± 0.1 e | 2.54 ± 0.05 bcd | 2.28 ± 0.19 f | 2.44 ± 0.09 d | 2.48 ± 0.13 e |
| TL 15% | 2.18 ± 0.13 f | 2.22 ± 0.11 f | 2.38 ± 0.14 f | 2.43 ± 0.06 e | 2.48 ± 0.08 d | 2.42 ± 0.11 e | 2.44 ± 0.1 d | 2.4 ± 0.09 f |
| p | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| P. parasitica | P. capsici | |||
|---|---|---|---|---|
| Treatment | Severity | Plants without Symptoms (%) | Severity | Plants without Symptoms (%) |
| T0 | 1.00 ± 0.00 c* | 100% | 1.00 ± 0.00 c | 100% |
| TI | 4.20 ± 1.13 a | 0% | 5.00 ± 0.00 a | 0% |
| TA | 1.40 ± 0.69 c | 70% | 2.30 ± 1.76 b | 60% |
| TS | 2.60 ± 2.06 b | 60% | 4.10 ± 1.37 a | 5% |
| TL | 1.80 ± 1.31 bc | 70% | 1.20 ± 0.42 c | 80% |
| PAE | 1.50 ± 1.08 c | 80% | 1.50 ± 0.84 bc | 65% |
| TAR | 1.20 ± 0.42 c | 80% | 1.50 ± 0.84 bc | 75% |
| TSR | 1.10 ± 0.31 c | 90% | 2.40 ± 1.57 b | 50% |
| TLR | 1.20 ± 0.42 c | 80% | 1.10 ± 0.31 c | 90% |
| PAER | 1.30 ± 0.67 c | 80% | 1.50 ± 1.26 bc | 70% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, M.; Diánez, F.; Sánchez-Montesinos, B.; Huertas, V.; Moreno-Gavira, A.; Esteban García, B.; Garrido-Cárdenas, J.A.; Gea, F.J. Biocontrol of Diseases Caused by Phytophthora capsici and P. parasitica in Pepper Plants. J. Fungi 2023, 9, 360. https://doi.org/10.3390/jof9030360
Santos M, Diánez F, Sánchez-Montesinos B, Huertas V, Moreno-Gavira A, Esteban García B, Garrido-Cárdenas JA, Gea FJ. Biocontrol of Diseases Caused by Phytophthora capsici and P. parasitica in Pepper Plants. Journal of Fungi. 2023; 9(3):360. https://doi.org/10.3390/jof9030360
Chicago/Turabian StyleSantos, Mila, Fernando Diánez, Brenda Sánchez-Montesinos, Victoria Huertas, Alejandro Moreno-Gavira, Belén Esteban García, José A. Garrido-Cárdenas, and Francisco J. Gea. 2023. "Biocontrol of Diseases Caused by Phytophthora capsici and P. parasitica in Pepper Plants" Journal of Fungi 9, no. 3: 360. https://doi.org/10.3390/jof9030360
APA StyleSantos, M., Diánez, F., Sánchez-Montesinos, B., Huertas, V., Moreno-Gavira, A., Esteban García, B., Garrido-Cárdenas, J. A., & Gea, F. J. (2023). Biocontrol of Diseases Caused by Phytophthora capsici and P. parasitica in Pepper Plants. Journal of Fungi, 9(3), 360. https://doi.org/10.3390/jof9030360