
Characterization of Aspartic Proteases from Paracoccidioides brasiliensis and Their Role in Fungal Thermo-Dimorphism

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolate and Growth Conditions
2.2. Pepstatin A Inhibitor Susceptibility
2.3. Dimorphic Transition Assay
2.4. Spot Assay
2.5. Paracoccidioides RNA Isolation
2.6. Real-Time Quantitative RT-qPCR
2.7. Bioinformatic Analysis
2.8. Endoprotease Assay
2.9. Statistical Analysis
3. Results
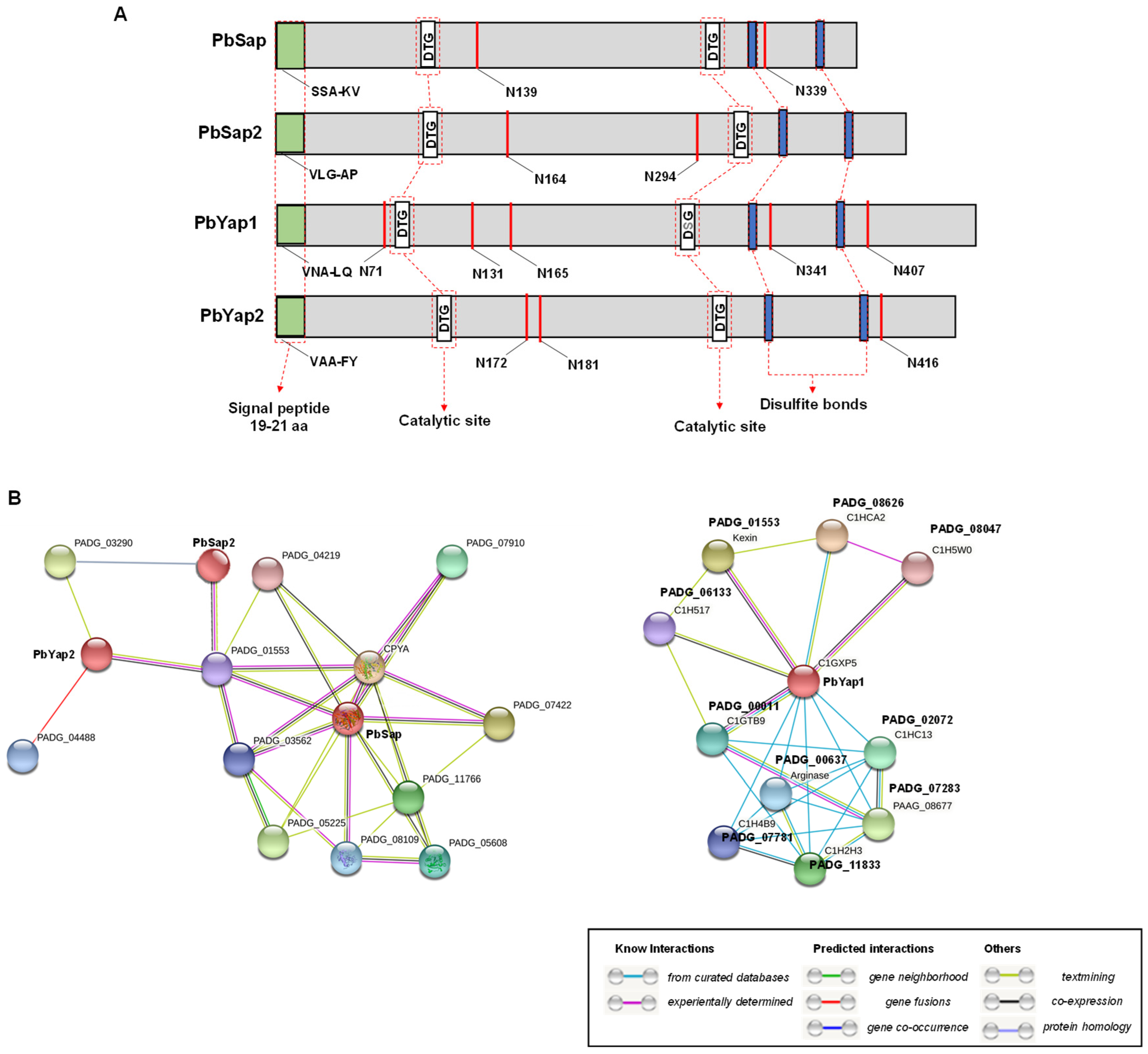
3.1. Identification and Characterization of Aspartic Proteases of P. brasiliensis
3.2. Interactome of Aspartic Protease Proteins of P. brasiliensis
3.3. Inhibition of Aspartic Protease Proteins Causes a Delay in the Mycelium-to-Yeast Transition
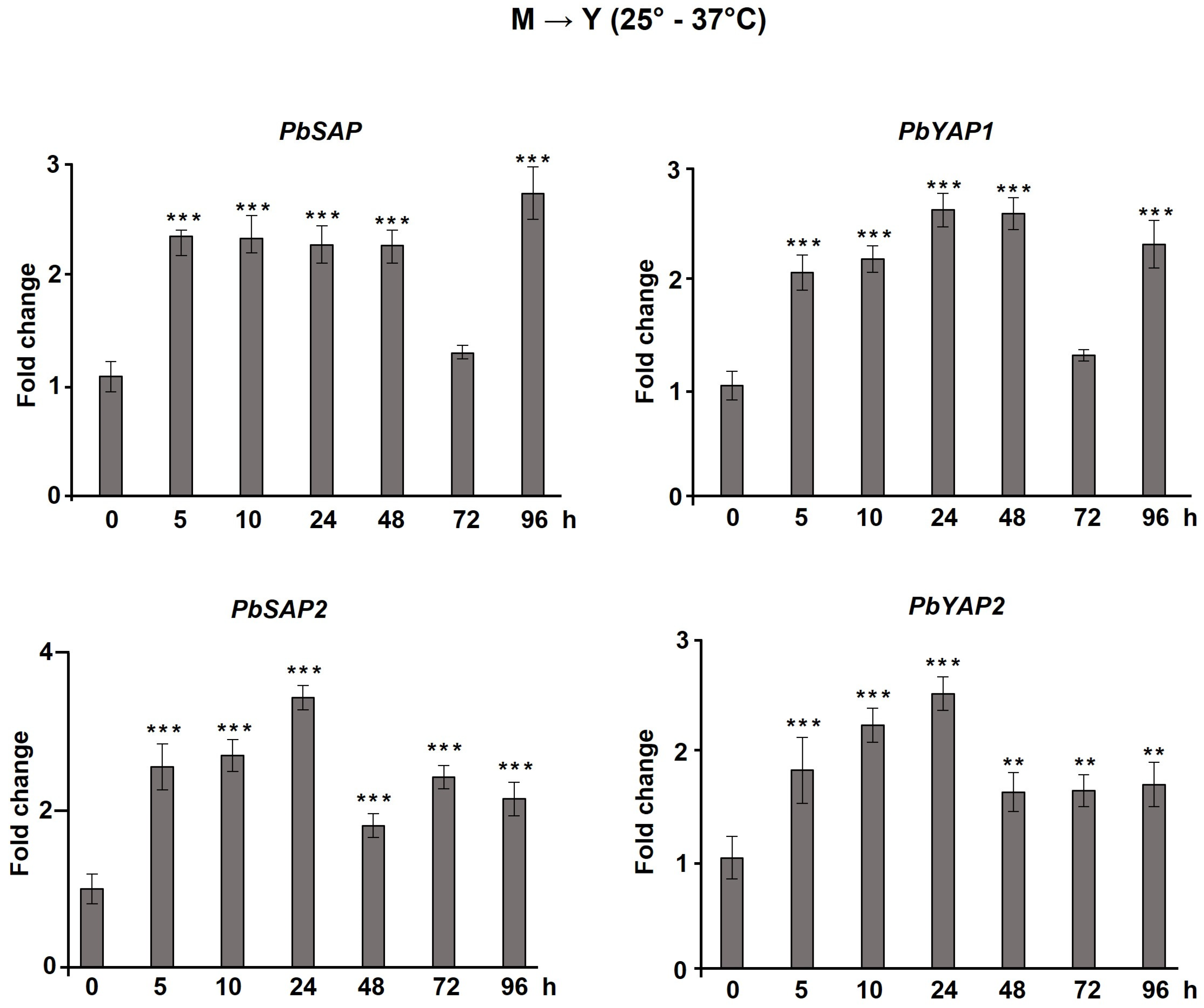
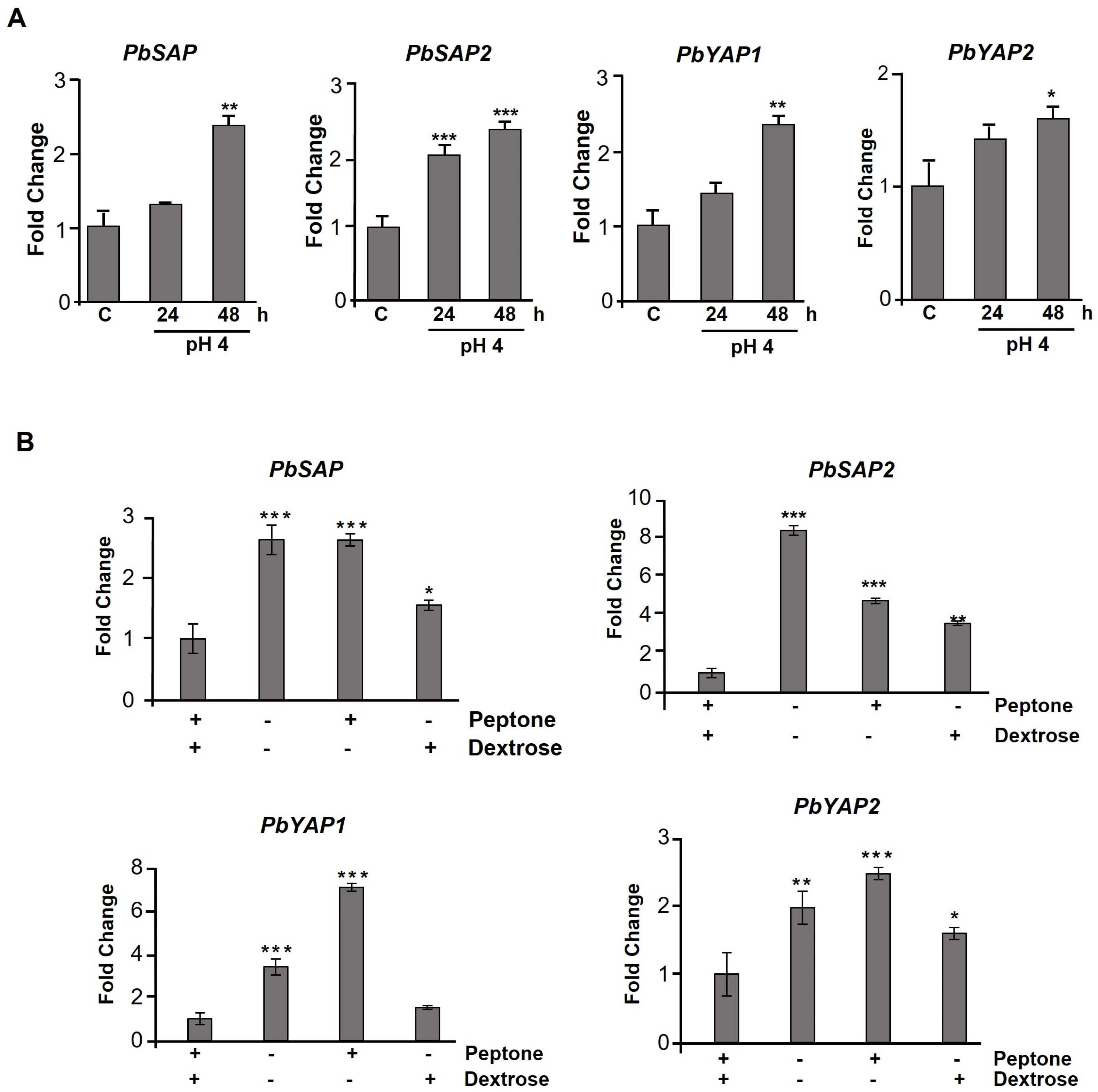
3.4. Aspartic Protease Genes Are Regulated during the Dimorphic Transition, at a Low pH, and in the Presence of Different Sources of Nutrients
3.5. Role of Aspartic Proteases in P. brasiliensis Cell Wall Maintenance and Response to Oxidative Stress
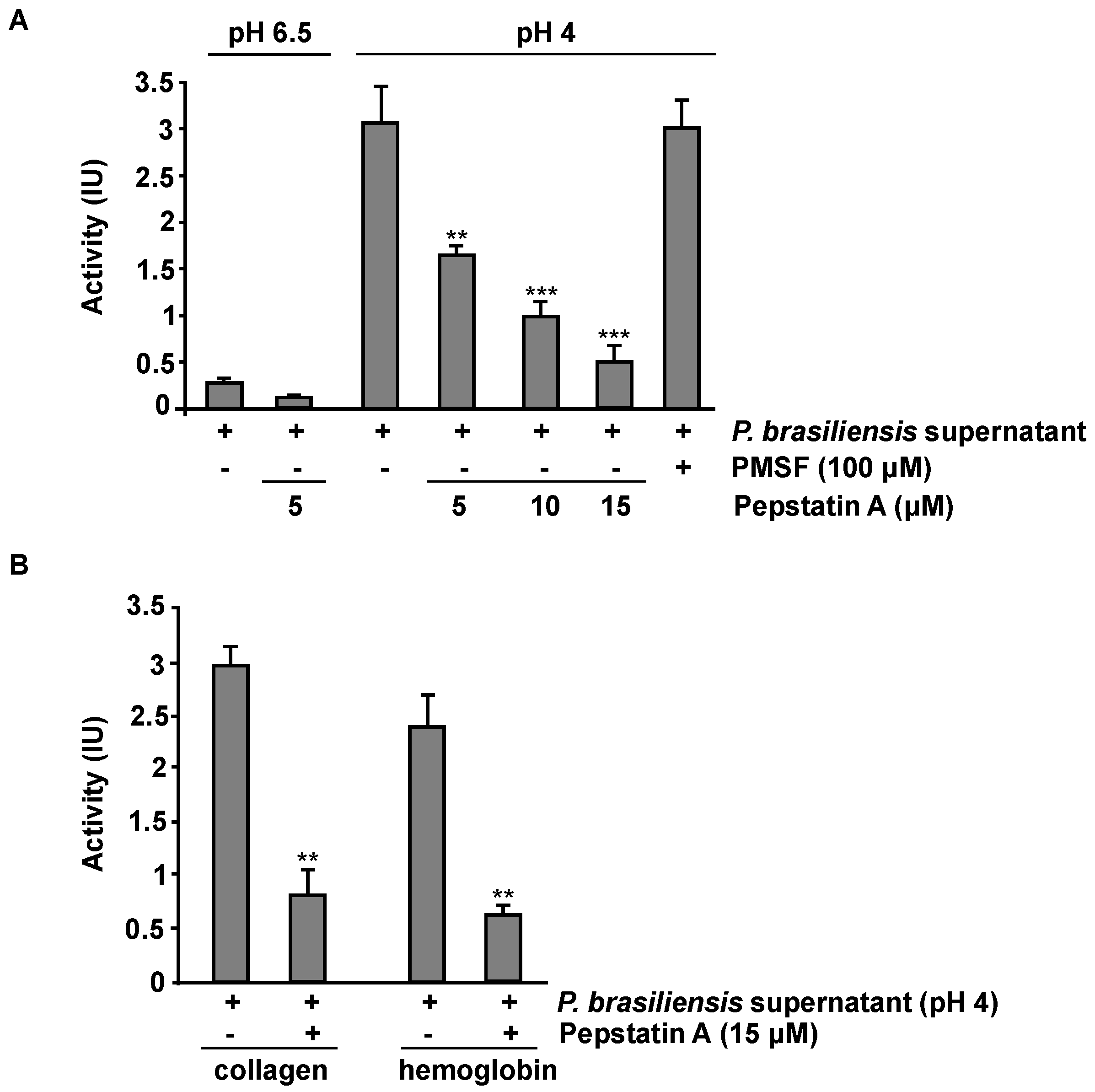
3.6. P. brasiliensis Secretes Aspartyl Proteases under Acidic Culture Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Denning, D.W.; Bromley, M.J. How to bolster the antifungal pipeline. Science 2015, 347, 1414–1416. [Google Scholar] [CrossRef] [Green Version]
- von Lilienfeld-Toal, M.; Wagener, J.; Einsele, H.; Cornely, O.A.; Kurzai, O. Invasive Fungal Infection. Dtsch. Ärzteblatt Int. 2019, 116, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Colombo, A.L.; Tobón, A.; Restrepo, A.; Queiroz-Telles, F.; Nucci, M. Epidemiology of endemic systemic fungal infections in Latin America. Med. Mycol. 2011, 49, 785–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queiroz-Telles, F.; Fahal, A.H.; Falci, D.R.; Caceres, D.H.; Chiller, T.; Pasqualotto, A.C. Neglected endemic mycoses. Lancet Infect. Dis. 2017, 17, e367–e377. [Google Scholar] [CrossRef] [PubMed]
- Rickerts, V. Klimawandel und Epidemiologie systemischer Pilzinfektionen. Bundesgesundheitsblatt Gesundh. Gesundh. 2019, 62, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Restrepo, A.; Tobón, A.M.; Cano, L.E. Paracoccidioidomycosis. In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases; Elsevier: Amsterdam, The Netherelands, 2015; pp. 2995–3002.e2992. [Google Scholar]
- Buccheri, R.; Khoury, Z.; Barata, L.C.B.; Benard, G. Incubation Period and Early Natural History Events of the Acute Form of Paracoccidioidomycosis: Lessons from Patients with a Single Paracoccidioides spp. Exposure. Mycopathologia 2016, 181, 435–439. [Google Scholar] [CrossRef]
- Chaves, A.F.A.; Navarro, M.V.; de Barros, Y.N.; Silva, R.S.; Xander, P.; Batista, W.L. Updates in Paracoccidioides Biology and Genetic Advances in Fungus Manipulation. J. Fungi 2021, 7, 116. [Google Scholar] [CrossRef]
- De Oliveira, H.C.; Assato, P.A.; Marcos, C.M.; Scorzoni, L.; Silva, A.C.A.D.P.E.; Silva, J.D.F.D.; Singulani, J.D.L.; Alarcon, K.M.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Paracoccidioides-host Interaction: An Overview on Recent Advances in the Paracoccidioidomycosis. Front. Microbiol. 2015, 6, 1319. [Google Scholar] [CrossRef] [Green Version]
- Camacho, E.; Niño-Vega, G.A. Paracoccidioides Spp.: Virulence Factors and Immune-Evasion Strategies. Mediat. Inflamm. 2017, 2017, 5313691. [Google Scholar] [CrossRef] [Green Version]
- Monod, M.; Hube, B.; Hess, D.; Sanglard, D. Differential regulation of SAP8 and SAPS, which encode two new members of the secreted aspartic proteinase family in Candida albicans. Microbiology 1998, 144, 2731–2737. [Google Scholar] [CrossRef] [Green Version]
- Suárez-Rendueles, P.; Villa, L.; Arbesú, M.J.; Escudero, B. The proteolytic system of the fission yeast Schizosaccharomyces pombe. FEMS Microbiol. Lett. 1991, 81, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Vartivarian, S.E. Virulence Properties and Nonimmune Pathogenetic Mechanisms of Fungi. Clin. Infect. Dis. 1992, 14, S30–S36. [Google Scholar] [CrossRef] [PubMed]
- White, T.C.; Agabian, N. Candida albicans secreted aspartyl proteinases: Isoenzyme pattern is determined by cell type, and levels are determined by environmental factors. J. Bacteriol. 1995, 177, 5215–5221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yike, I. Fungal proteases and their pathophysiological effects. Mycopathologia 2011, 171, 299–323. [Google Scholar] [CrossRef]
- Castilho, D.G.; Chaves, A.F.A.; Navarro, M.V.; Conceição, P.M.; Ferreira, K.S.; da Silva, L.S.; Xander, P.; Batista, W.L. Secreted aspartyl proteinase (PbSap) contributes to the virulence of Paracoccidioides brasiliensis infection. PLOS Negl. Trop. Dis. 2018, 12, e0006806. [Google Scholar] [CrossRef] [Green Version]
- Castilho, D.G.; Navarro, M.V.; Chaves, A.F.A.; Xander, P.; Batista, W.L. Recovery of the Paracoccidioides brasiliensis virulence after animal passage promotes changes in the antioxidant repertoire of the fungus. FEMS Yeast Res. 2018, 18. [Google Scholar] [CrossRef]
- Davies, D.R. The Structure and Function of the Aspartic Proteinases. Annu. Rev. Biophys. Biophys. Chem. 1990, 19, 189–215. [Google Scholar] [CrossRef]
- Kyes, S.A.; Kraemer, S.M.; Smith, J.D. Antigenic Variation in Plasmodium falciparum: Gene Organization and Regulation of the var Multigene Family. Eukaryot. Cell 2007, 6, 1511–1520. [Google Scholar] [CrossRef] [Green Version]
- Silva, N.C.; Nery, J.M.; Dias, A.L.T. Aspartic proteinases of Candida spp.: Role in pathogenicity and antifungal resistance. Mycoses 2014, 57, 1–11. [Google Scholar] [CrossRef]
- Loaiza-Loeza, S.; Parra-Ortega, B.; Cancino-Díaz, J.C.; Illades-Aguiar, B.; Hernández-Rodríguez, C.H.; Villa-Tanaca, L. Differential expression of Candida dubliniensis-secreted aspartyl proteinase genes (CdSAP1–4) under different physiological conditions and during infection of a keratinocyte culture. FEMS Immunol. Med. Microbiol. 2009, 56, 212–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parra-Ortega, B.; Villa-Tanaca, L.; Hernandez-Rodriguez, C. Evolution of GPI-Aspartyl Proteinases (Yapsines) of Candida spp. In Gene Duplication; Friedberg, F., Ed.; InTech: London, UK, 2011. [Google Scholar]
- Cortés-Acosta, E.; Ibarra, J.A.; Ramírez-Saad, H.C.; Vargas-Mendoza, C.F.; Villa-Tanaca, L.; Hernández-Rodríguez, C. Polymorphism in the regulatory regions of genes CgYPS1 and CgYPS7 encoding yapsins in Candida glabrata is associated with changes in expression levels. FEMS Yeast Res. 2017, 17, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnon-Arsenault, I.; Tremblay, J.; Bourbonnais, Y. Fungal yapsins and cell wall: A unique family of aspartic peptidases for a distinctive cellular function. FEMS Yeast Res. 2006, 6, 966–978. [Google Scholar] [CrossRef]
- Ammerer, G.; Hunter, C.P.; Rothman, J.H.; Saari, G.C.; Valls, L.A.; Stevens, T.H. PEP4 gene of Saccharomyces cerevisiae encodes proteinase A, a vacuolar enzyme required for processing of vacuolar precursors. Mol. Cell. Biol. 1986, 6, 2490–2499. [Google Scholar] [CrossRef]
- Hazel, H.B.V.D.; Kielland-Brandt, M.C.; Winther, J.R. Autoactivation of proteinase A initiates activation of yeast vacuolar zymogens. Eur. J. Biochem. 1992, 207, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.W. Three proteolytic systems in the yeast saccharomyces cerevisiae. J. Biol. Chem. 1991, 266, 7963–7966. [Google Scholar] [CrossRef]
- Juárez-Montiel, M.; Tesillo-Moreno, P.; Cruz-Angeles, A.; Soberanes-Gutiérrez, V.; Chávez-Camarillo, G.; Ibarra, J.A.; Hernández-Rodríguez, C.; Villa-Tanaca, L. Heterologous expression and characterization of the aspartic endoprotease Pep4um from Ustilago maydis, a homolog of the human Chatepsin D, an important breast cancer therapeutic target. Mol. Biol. Rep. 2018, 45, 1155–1163. [Google Scholar] [CrossRef]
- de Assis Tacco, B.A.C.; Parente, J.A.; Barbosa, M.S.; Báo, S.N.; de Souza Gsóes, T.; Pereira, M.; de Almeida Soares, C.M. Characterization of a secreted aspartyl protease of the fungal pathogen Paracoccidioides brasiliensis. Med. Mycol. 2009, 47, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Bastos, K.P.; Bailão, A.M.; Borges, C.L.; Faria, F.P.; Felipe, M.S.; Silva, M.G.; Martins, W.S.; Fiúza, R.B.; Pereira, M.; Soares, C.M. The transcriptome analysis of early morphogenesis in Paracoccidioides brasiliensis mycelium reveals novel and induced genes potentially associated to the dimorphic process. BMC Microbiol. 2007, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Castilho, D.G.; Chaves, A.F.A.; Xander, P.; Zelanis, A.; Kitano, E.S.; Serrano, S.M.T.; Tashima, A.K.; Batista, W.L. Exploring potential virulence regulators in Paracoccidioides brasiliensis isolates of varying virulence through quantitative proteomics. J. Proteome Res. 2014, 13, 4259–4271. [Google Scholar] [CrossRef]
- Braz, J.D.; Sardi, J.D.C.O.; Pitangui, N.D.S.; Voltan, A.R.; Almeida, A.M.F.; Mendes-Giannini, M.J.S. Gene expression of Paracoccidioides virulence factors after interaction with macrophages and fibroblasts. Mem. Inst. Oswaldo Cruz. 2021, 116, e200592. [Google Scholar] [CrossRef]
- Nakai, T.; Uno, J.; Ikeda, F.; Tawara, S.; Nishimura, K.; Miyaji, M. In Vitro Antifungal Activity of Micafungin (FK463) against Dimorphic Fungi: Comparison of Yeast-Like and Mycelial Forms. Antimicrob. Agents Chemother. 2003, 47, 1376–1381. [Google Scholar] [CrossRef] [Green Version]
- Batista, W.L.; Barros, T.F.; Goldman, G.H.; Morais, F.V.; Puccia, R. Identification of transcription elements in the 5′ intergenic region shared by LON and MDJ1 heat shock genes from the human pathogen Paracoccidioides brasiliensis. Evaluation of gene expression. Fungal Genet. Biol. 2007, 44, 347–356. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Teufel, F.; Armenteros, J.J.A.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Hamby, S.E.; Hirst, J.D. Prediction of glycosylation sites using random forests. BMC Bioinform. 2008, 9, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fankhauser, N.; Mäser, P. Identification of GPI anchor attachment signals by a Kohonen self-organizing map. Bioinformatics 2005, 21, 1846–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutmanas, A.; Alhroub, Y.; Battle, G.M.; Berrisford, J.M.; Bochet, E.; Conroy, M.J.; Dana, J.M.; Montecelo, M.A.F.; van Ginkel, G.; Gore, S.P.; et al. PDBe: Protein Data Bank in Europe. Nucleic Acids Res. 2014, 42, D285–D291. [Google Scholar] [CrossRef] [Green Version]
- Camargo, Z.P.; Taborda, C.P.; Rodrigues, E.G.; Travassos, L.R. The use of cell-free antigens of Paracoccidioides brasiliensis in serological tests. J. Med. Vet. Mycol. Bi-Mon. Publ. Int. Soc. Hum. Anim. Mycol. 1991, 29, 31–38. [Google Scholar]
- O’Donoghue, A.J.; Knudsen, G.M.; Beekman, C.; Perry, J.A.; Johnson, A.D.; DeRisi, J.L.; Craik, C.S.; Bennett, R.J. Destructin-1 is a collagen-degrading endopeptidase secreted by Pseudogymnoascus destructans, the causative agent of white-nose syndrome. Proc. Natl. Acad. Sci. USA 2015, 112, 7478–7483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Tu, T.; Zheng, J.; Bai, Y.; Huang, H.; Su, X.; Wang, Y.; Wang, Y.; Yao, B.; Luo, H. Improvement of BsAPA Aspartic Protease Thermostability via Autocatalysis-Resistant Mutation. J. Agric. Food Chem. 2019, 67, 10505–10512. [Google Scholar] [CrossRef]
- Li, S.C.; Kane, P.M. The yeast lysosome-like vacuole: Endpoint and crossroads. Biochim. Biophys. Acta BBA Mol. Cell Res. 2009, 1793, 650–663. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, G.F.; Góes, C.G.d.; Onorio, D.S.; Campos, C.B.L.d.; Morais, F.V. Autophagy in Paracoccidioides brasiliensis under normal mycelia to yeast transition and under selective nutrient deprivation. PLoS ONE 2018, 13, e0202529. [Google Scholar] [CrossRef] [Green Version]
- Felk, A.; Kretschmar, M.; Albrecht, A.; Schaller, M.; Beinhauer, S.; Nichterlein, T.; Sanglard, D.; Korting, H.C.; Schäfer, W.; Hube, B. Candida albicans Hyphal Formation and the Expression of the Efg1-Regulated Proteinases Sap4 to Sap6 Are Required for the Invasion of Parenchymal Organs. Infect. Immun. 2002, 70, 3689–3700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, M.; Weerasinghe, H.; Tedja, I.; Andrianopoulos, A. A unique aspartyl protease gene expansion in Talaromyces marneffei plays a role in growth inside host phagocytes. Virulence 2019, 10, 277–291. [Google Scholar] [CrossRef] [Green Version]
- Soberanes-Gutiérrez, C.V.; Juárez-Montiel, M.; Olguín-Rodríguez, O.; Hernández-Rodríguez, C.; Ruiz-Herrera, J.; Villa-Tanaca, L. The pep4 gene encoding proteinase A is involved in dimorphism and pathogenesis of U stilago maydis: Role of vacuole proteinase A in Ustilago maydis. Mol. Plant Pathol. 2015, 16, 837–846. [Google Scholar] [CrossRef]
- Aoki, W.; Kitahara, N.; Miura, N.; Morisaka, H.; Yamamoto, Y.; Kuroda, K.; Ueda, M. Candida albicans Possesses Sap7 as a Pepstatin A-Insensitive Secreted Aspartic Protease. PLoS ONE 2012, 7, e32513. [Google Scholar] [CrossRef] [Green Version]
- Purushothaman, K.; Bhat, S.K.; Singh, S.A.; Marathe, G.K.; Rao, A.R.G.A. Aspartic protease from Aspergillus niger: Molecular characterization and interaction with pepstatin A. Int. J. Biol. Macromol. 2019, 139, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Mandujano-González, V.; Villa-Tanaca, L.; Anducho-Reyes, M.A.; Mercado-Flores, Y. Secreted fungal aspartic proteases: A review. Rev. Iberoam. Micol. 2016, 33, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Naglik, J.R.; Challacombe, S.J.; Hube, B. Candida albicans Secreted Aspartyl Proteinases in Virulence and Pathogenesis. Microbiol. Mol. Biol. Rev. 2003, 67, 400–428. [Google Scholar] [CrossRef] [Green Version]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.M. Three Extracellular Proteases from Cochliobolus carbonum: Cloning and Targeted Disruption of ALP1. Mol. Plant-Microbe Interact. 1996, 9, 290. [Google Scholar] [CrossRef] [PubMed]
- Steiner, K.; Abigail, A.; Dixie, S.A.; Whitt, D.; Pathogenesis, B. A Molecular Approach. XXVII + 418 S., 137 Abb., 22 Tab. Washington D.C. 1994. ASM Press. L 24.95. ISBN: 1-55581-094-2. J. Basic Microbiol. 1996, 36, 148. [Google Scholar] [CrossRef]
- Albrecht, A.; Felk, A.; Pichova, I.; Naglik, J.R.; Schaller, M.; de Groot, P.; MacCallum, D.; Odds, F.C.; Schäfer, W.; Klis, F.; et al. Glycosylphosphatidylinositol-anchored Proteases of Candida albicans Target Proteins Necessary for Both Cellular Processes and Host-Pathogen Interactions. J. Biol. Chem. 2006, 281, 688–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krysan, D.J.; Ting, E.L.; Abeijon, C.; Kroos, L.; Fuller, R.S. Yapsins Are a Family of Aspartyl Proteases Required for Cell Wall Integrity in Saccharomyces cerevisiae. Eukaryot. Cell 2005, 4, 1364–1374. [Google Scholar] [CrossRef] [Green Version]
- Yoshimi, A.; Miyazawa, K.; Abe, K. Function and Biosynthesis of Cell Wall α-1,3-Glucan in Fungi. J. Fungi 2017, 3, 63. [Google Scholar] [CrossRef]
- Fleck, C.B.; Schöbel, F.; Brock, M. Nutrient acquisition by pathogenic fungi: Nutrient availability, pathway regulation, and differences in substrate utilization. Int. J. Med. Microbiol. 2011, 301, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.; Casadevall, A. Phagosome Extrusion and Host-Cell Survival after Cryptococcus neoformans Phagocytosis by Macrophages. Curr. Biol. 2006, 16, 2161–2165. [Google Scholar] [CrossRef] [Green Version]
- Bliska, J.B.; Casadevall, A. Intracellular pathogenic bacteria and fungi—A case of convergent evolution? Nat. Rev. Microbiol. 2009, 7, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, D.E. Hemoglobin Degradation. In Malaria: Drugs, Disease and Post-Genomic Biology; Compans, R.W., Cooper, M.D., Honjo, T., Koprowski, H., Melchers, F., Oldstone, M.B.A., Olsnes, S., Potter, M., Vogt, P.K., Wagner, H., et al., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 295, pp. 275–291. [Google Scholar]
- Karkowska-Kuleta, J.; Rapala-Kozik, M.; Kozik, A. Fungi pathogenic to humans: Molecular bases of virulence of Candida albicans, Cryptococcus neoformans and Aspergillus fumigatus. Acta Biochim. Pol. 2009, 56, 211–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polke, M.; Hube, B.; Jacobsen, I.D. Candida Survival Strategies. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 91, pp. 139–235. [Google Scholar]
- Köhler, J.R.; Hube, B.; Puccia, R.; Casadevall, A.; Perfect, J.R. Fungi that Infect Humans. Microbiol. Spectr. 2017, 5, 813–843. [Google Scholar] [CrossRef]
- Schaller, M.; Borelli, C.; Korting, H.C.; Hube, B. Hydrolytic enzymes as virulence factors of Candida albicans. Mycoses 2005, 48, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Pietrella, D.; Pandey, N.; Gabrielli, E.; Pericolini, E.; Perito, S.; Kasper, L.; Bistoni, F.; Cassone, A.; Hube, B.; Vecchiarelli, A. Secreted aspartic proteases of Candida albicans activate the NLRP3 inflammasome. Eur. J. Immunol. 2013, 43, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, M.; Battu, A.; Kaur, R. Aspartyl proteases in Candida glabrata are required for suppression of the host innate immune response. J. Biol. Chem. 2018, 293, 6410–6433. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.K.; Németh, T.; Papp, A.; Tóth, R.; Lukácsi, S.; Heidingsfeld, O.; Dostal, J.; Vágvölgyi, C.; Bajtay, Z.; Józsi, M.; et al. Functional Characterization of Secreted Aspartyl Proteases in Candida parapsilosis. mSphere 2019, 4, e00484-19. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Downs, D.; Ghosh, K.; Ghosh, A.K.; Staib, P.; Monod, M.; Tang, J. Candida albicans secreted aspartic proteases 4–6 induce apoptosis of epithelial cells by a novel Trojan horse mechanism. FASEB J. 2013, 27, 2132–2144. [Google Scholar] [CrossRef] [PubMed]
- Parra-Ortega, B.; Cruz-Torres, H.; Villa-Tanaca, L.; Hernández-Rodríguez, C. Phylogeny and evolution of the aspartyl protease family from clinically relevant Candida species. Memórias Do Inst. Oswaldo Cruz 2009, 104, 505–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soberanes-Gutiérrez, C.V.; Vázquez-Carrada, M.; López-Villegas, E.O.; Vega-Arreguín, J.C.; Villa-Tanaca, L.; Ruiz-Herrera, J. Autophagosomes accumulation in the vacuoles of the fungus Ustilago maydis and the role of proteases in their digestion. FEMS Microbiol. Lett. 2019, 366, fnz108. [Google Scholar] [CrossRef]
- Bryant, N.J.; Stevens, T.H. Vacuole Biogenesis in Saccharomyces cerevisiae: Protein Transport Pathways to the Yeast Vacuole. Microbiol. Mol. Biol. Rev. 1998, 62, 230–247. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Xu, H.; Wang, P.; Liu, L.; Cai, J.; Chen, Y.; Zhao, X.; You, X.; Liu, J.; Guo, X.; et al. The entry of unclosed autophagosomes into vacuoles and its physiological relevance. PLoS Genet. 2022, 18, e1010431. [Google Scholar] [CrossRef]
- Azzopardi, M.; Farrugia, G.; Balzan, R. Cell-cycle involvement in autophagy and apoptosis in yeast. Mech. Ageing Dev. 2017, 161, 211–224. [Google Scholar] [CrossRef]
- Bartoszewska, M.; Kiel, J.A. The Role of Macroautophagy in Development of Filamentous Fungi. Antioxid. Redox Signal. 2011, 14, 2271–2287. [Google Scholar] [CrossRef] [PubMed]
- Kohda, T.A.; Tanaka, K.; Konomi, M.; Sato, M.; Osumi, M.; Yamamoto, M. Fission yeast autophagy induced by nitrogen starvation generates a nitrogen source that drives adaptation processes. Genes Cells 2007, 12, 155–170. [Google Scholar] [CrossRef]
- Liu, X.-H.; Lu, J.-P.; Lin, F.-C. Autophagy During Conidiation, Conidial Germination and Turgor Generation in Magnaporthe grisea. Autophagy 2007, 3, 472–473. [Google Scholar] [CrossRef] [Green Version]
- López-Berges, M.S.; Rispail, N.; Prados-Rosales, R.C.; Di Pietro, A. A Nitrogen Response Pathway Regulates Virulence Functions in Fusarium oxysporum via the Protein Kinase TOR and the bZIP Protein MeaB. Plant Cell 2010, 22, 2459–2475. [Google Scholar] [CrossRef] [Green Version]
- Umekawa, M.; Ujihara, M.; Makishima, K.; Yamamoto, S.; Takematsu, H.; Wakayama, M. The signaling pathways underlying starvation-induced upregulation of α-mannosidase Ams1 in Saccharomyces cerevisiae. Biochim. Biophys. Acta 2016, 1860, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Kerstens, W.; Van Dijck, P. A Cinderella story: How the vacuolar proteases Pep4 and Prb1 do more than cleaning up the cell’s mass degradation processes. Microb. Cell 2018, 5, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Hube, B. Candida albicans secreted aspartyl proteinases. Curr. Top. Med. Mycol. 1996, 7, 55–69. [Google Scholar]
- Palmer, G.E. Vacuolar trafficking and Candida albicans pathogenesis. Commun. Integr. Biol. 2011, 4, 240–242. [Google Scholar] [CrossRef] [Green Version]
- Palmer, G.E.; Kelly, M.N.; Sturtevant, J.E. The Candida albicans Vacuole Is Required for Differentiation and Efficient Macrophage Killing. Eukaryot. Cell 2005, 4, 1677–1686. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Ma, B.; Cormack, B.P. A family of glycosylphosphatidylinositol-linked aspartyl proteases is required for virulence of Candida glabrata. Proc. Natl. Acad. Sci. USA 2007, 104, 7628–7633. [Google Scholar] [CrossRef] [Green Version]
- Bairwa, G.; Rasheed, M.; Taigwal, R.; Sahoo, R.; Kaur, R. GPI (glycosylphosphatidylinositol)-linked aspartyl proteases regulate vacuole homoeostasis in Candida glabrata. Biochem. J. 2014, 458, 323–334. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′→3) | |
|---|---|---|
| PbSAP * | Sense | GATGACTCTGAGGCTACCTTTG |
| Anti Sense | ATCGAGATCAACCTCCCAGTA | |
| PbSAP2 | Sense | CCGTCTTCACCGCTCAATTA |
| Anti Sense | CCACAGGGACATCAACCATATC | |
| PbYAP1 | Sense | GTCAACATGAGCGAGCTAGT |
| Anti Sense | GAGATGCCGAAGATACAGGTT | |
| PbYAP2 | Sense | CCCGGTTATCTGTGAGAAAGTC |
| Anti Sense | TGCGGATGACGTAGACAAAC | |
| α-TUB* | Sense | GTGGACCAGGTGATCGATGT |
| Anti Sense | ACCCTGGAGGCAGTCACA | |
| 18S * | Sense | CGGAGAGAGGGAGCCTGAGAA |
| Anti Sense | GGGATTGGGTAATTTGCGC |
| Gene | Assession Number | Gene with Intron (bp) | Gene without Intron (bp) | Protein (aa) | Pep4 Similarity (%) * |
|---|---|---|---|---|---|
| PbSAP | PADG_00634 | 1460 | 1203 | 400 | 59 |
| PbSAP2 ** | PADG_03432 | 1571 | 1281 | 426 | 30 |
| PbYAP1 ** | PADG_12056# | 1681 | 1485 | 495 | 27 |
| PbYAP2 ** | PADG_08282 | 1601 | 1446 | 481 | 29 |
| Protein | Signal Peptide | Protein Molecular Weight (kDa) | Isoeletric Point (pI) | N-glycosylation Sites | PDB Similarity |
|---|---|---|---|---|---|
| PbSap | SSA-KV | 43.8 | 5.36 | N139, N339 | Saccharopepsin—64% |
| PbSap2 * | VLG-AP | 57 | 5.21 | N164, N294 | Aspergillopepsin—46% |
| PbYap1 * | VNG-LQ | 51 | 5.03 | N71, N131, N165, N341, N407 | Candidapepsin—38% |
| PbYap2 * | VAA-FY | 52.5 | 5.1 | N172, N181, N416 | Endopeptidase—30% |
| Gene ID | Function | Interaction |
|---|---|---|
| PADG_06314 | Carboxypeptidase Y homolog A; vacuolar carboxypeptidase | PbSap |
| PADG_07422 | Uncharacterized protein; belongs to the peptidase S8 family | PbSap |
| PADG_05225 | Orotidine 5′-phosphate decarboxylase; belongs to the OMP decarboxylase family | PbSap |
| PADG_11766 | Alkaline phosphatase | PbSap |
| PADG_07910 | Proteinase T [B. dermatitidis ATCC 18188] | PbSap |
| PADG_05608 | Ras-like protein Rab7 [B. dermatitidis ATCC 18188] | PbSap |
| PADG_08109 | Autophagy-related protein; belongs to the ATG8 family | PbSap |
| PADG_03562 | Glucose-regulated protein [B. dermatitidis ATCC 18188] | PbSap |
| PADG_01553 | Kexin [B. dermatitidis ATCC 18188] | PbSap, PbSap2, PbYap1 and PbYap2 |
| PADG_04219 | Glycosyl hydrolase [B. dermatitidis ATCC 18188] | PbSap |
| PADG_03290 | Tripeptidyl peptidase SED3 [B. dermatitidis ATCC 18188] | PbSap2 and PbYap2 |
| PADG_04488 | Lipoate-protein ligase A [B. dermatitidis ATCC 18188] | PbYap2 |
| PADG_08626 | Uncharacterized protein | PbYap1 |
| PADG_07283 | Uncharacterized protein | PbYap2 |
| PADG_11833 | Hydrolase | PbYap2 |
| PADG_02072 | Uncharacterized protein | PbYap2 |
| PADG_00011 | Actin binding protein | PbYap2 |
| PADG_00637 | Arginase; belongs to the arginase family. | PbYap2 |
| PADG_07781 | Pyridoxine kinase | PbYap2 |
| PADG_06133 | Mating-type alpha-pheromone receptor PreB | PbYap2 |
| PADG_08047 | Serine/threonine-protein kinase ppk4 | PbYap2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, R.d.S.; Segura, W.D.; Oliveira, R.S.; Xander, P.; Batista, W.L. Characterization of Aspartic Proteases from Paracoccidioides brasiliensis and Their Role in Fungal Thermo-Dimorphism. J. Fungi 2023, 9, 375. https://doi.org/10.3390/jof9030375
Silva RdS, Segura WD, Oliveira RS, Xander P, Batista WL. Characterization of Aspartic Proteases from Paracoccidioides brasiliensis and Their Role in Fungal Thermo-Dimorphism. Journal of Fungi. 2023; 9(3):375. https://doi.org/10.3390/jof9030375
Chicago/Turabian StyleSilva, Rafael de Souza, Wilson Dias Segura, Reinaldo Souza Oliveira, Patricia Xander, and Wagner Luiz Batista. 2023. "Characterization of Aspartic Proteases from Paracoccidioides brasiliensis and Their Role in Fungal Thermo-Dimorphism" Journal of Fungi 9, no. 3: 375. https://doi.org/10.3390/jof9030375
APA StyleSilva, R. d. S., Segura, W. D., Oliveira, R. S., Xander, P., & Batista, W. L. (2023). Characterization of Aspartic Proteases from Paracoccidioides brasiliensis and Their Role in Fungal Thermo-Dimorphism. Journal of Fungi, 9(3), 375. https://doi.org/10.3390/jof9030375