


Interactions between Entomopathogenic Fungi and Insects and Prospects with Glycans



Abstract
:1. Introduction
2. EPF Pathogenicity and the Interaction between Fungi and Insect
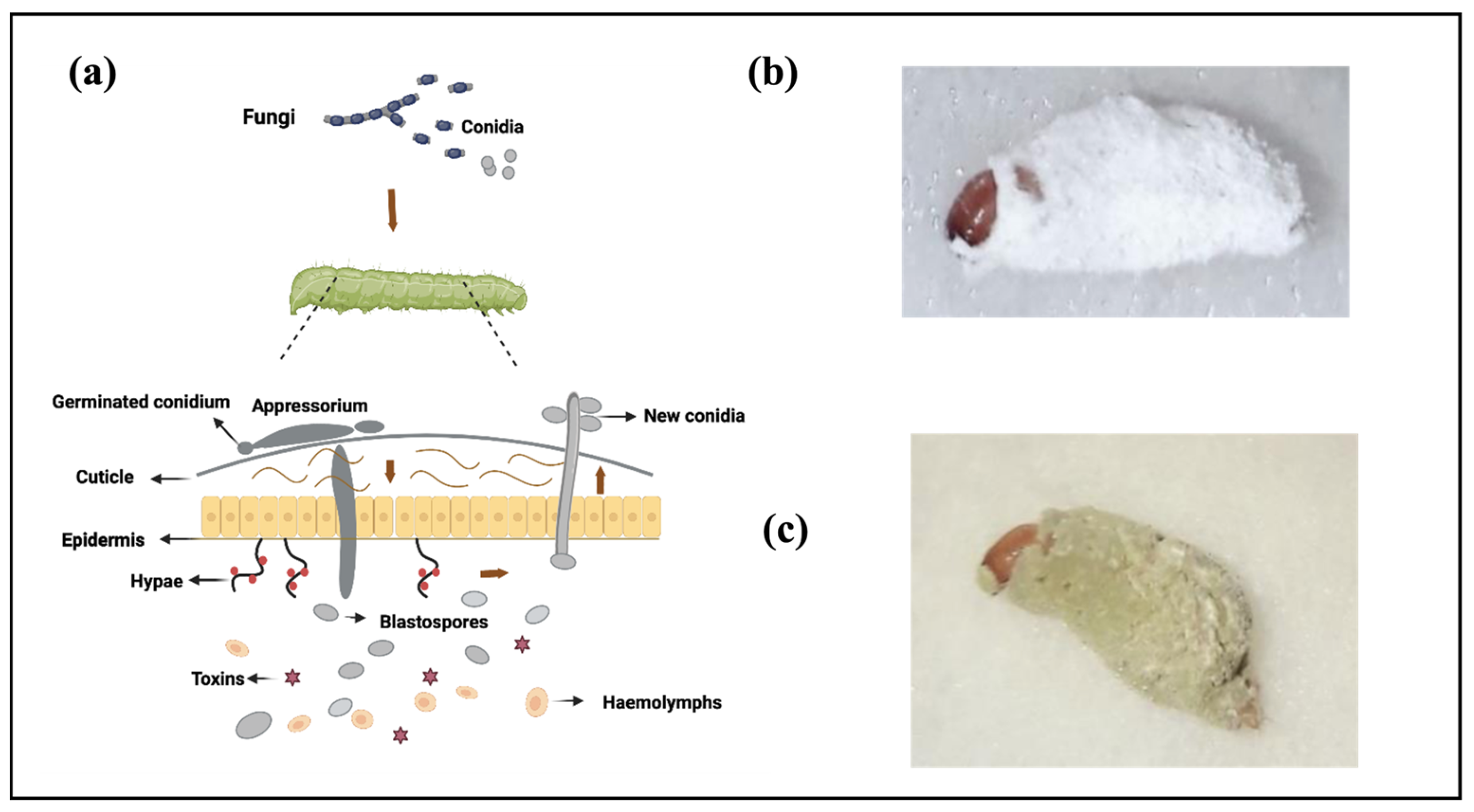
2.1. Fungi’s Infection Mode of Action in Insects: Penetration through the Cuticle

2.2. Ways of Interaction between Fungi and Insects
2.2.1. Interaction of the Fungus with the Microbiome of the Insect
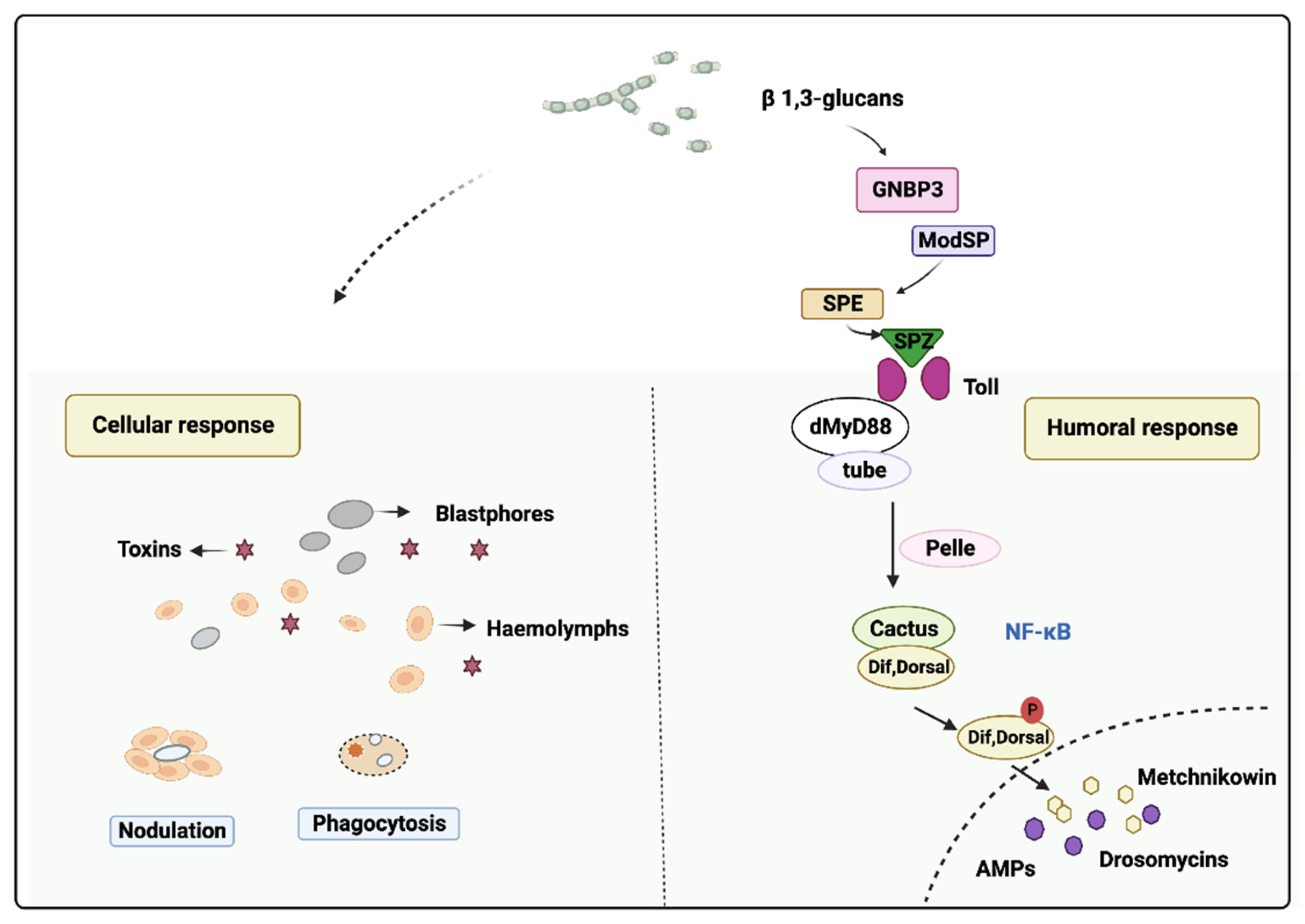
2.2.2. Stimulation of the Insect Immune-Competence by EPF
3. New Interesting Findings on the Interaction of Fungi in Insects
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Naqqash, M.N.; Gökçe, A.; Bakhsh, A.; Salim, M. Insecticide resistance and its molecular basis in urban insect pests. Parasitol. Res. 2016, 115, 1363–1373. [Google Scholar] [CrossRef]
- Venkatesan, T.; Chethan, B.R.; Mani, M. Insecticide resistance and its management in the insect pests of horticultural crops. In Trends in Horticultural Entomology; Mani, M., Ed.; Springer: Singapore, 2022; pp. 455–490. [Google Scholar] [CrossRef]
- Sparks, T.C.; Storer, N.; Porter, A.; Slater, R.; Nauen, R. Insecticide resistance management and industry: The origins and evolution of the Insecticide Resistance Action Committee (IRAC) and the mode of action classification scheme. Pest Manag. Sci. 2021, 77, 2609–2619. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef]
- Wakil, W.; Gulzar, S.; Prager, S.M.; Ghazanfar, M.U.; Shapiro-Ilan, D.I. Efficacy of entomopathogenic fungi, nematodes and spinetoram combinations for integrated management of Thrips tabaci: A two-year onion field study. Pest Manag. Sci. 2023. [Google Scholar] [CrossRef] [PubMed]
- Wakil, W.; Ghazanfar, M.U.; Usman, M.; Hunter, D.; Shi, W. Fungal-based biopesticide formulations to control nymphs and adults of the desert locust, Schistocerca gregaria Forskål (Orthoptera: Acrididae): A laboratory and field cage study. Agronomy 2022, 12, 1160. [Google Scholar] [CrossRef]
- Wakil, W.; Usman, M.; Piñero, J.C.; Wu, S.; Toews, M.D.; Shapiro-Ilan, D.I. Combined application of entomopathogenic nematodes and fungi against fruit flies, Bactrocera zonata and B. dorsalis (Diptera: Tephritidae): Laboratory cups to field study. Pest Manag. Sci. 2022, 78, 2779–2791. [Google Scholar] [CrossRef]
- Wakil, W.; Tahir, M.; Al-Sadi, A.M.; Shapiro-Ilan, D. Interactions between two invertebrate pathogens: An endophytic fungus and an externally applied bacterium. Front. Microbiol. 2020, 11, 2624. [Google Scholar] [CrossRef]
- Qayyum, M.A.; Saleem, M.A.; Saeed, S.; Wakil, W.; Ishtiaq, M.; Ashraf, W.; Ahmed, N.; Ali, M.; Ikram, R.M.; Yasin, M.; et al. Integration of entomopathogenic fungi and eco-friendly insecticides for management of red palm weevil, Rhynchophorus ferrugineus (Olivier). Saudi J. Biol. Sci. 2020, 27, 1811–1817. [Google Scholar] [CrossRef]
- Gulzar, S.; Wakil, W.; Shapiro-Ilan, D.I. Combined effect of entomopathogens against Thrips tabaci Lindeman (Thysanoptera: Thripidae): Laboratory, greenhouse and field trials. Insects 2021, 12, 456. [Google Scholar] [CrossRef]
- Marrone, P.G. Status of the biopesticide market and prospects for new bioherbicides. Pest Manag. Sci. 2023. [Google Scholar] [CrossRef]
- AgroPages. Bioinput Market Grows 67% during 2021–22 Crop in Brazil-Agricultural News. Available online: https://news.agropages.com/News/NewsDetail---45387.htm (accessed on 29 March 2023).
- Shah, P.A.; Pell, J.K. Entomopathogenic fungi as biological control agents. Appl. Microbiol. Biotechnol. 2003, 61, 413–423. [Google Scholar] [CrossRef]
- Pucheta, D.M.; Macias, A.F.; Navarro, S.R.; Mayra, D.L.T. Mechanism of action of entomopathogenic fungi. Interciencia 2016, 156, 2164–2171. [Google Scholar]
- Tulloch, M. The genus Metarhizium. Trans. Br. Mycol. Soc. 1976, 3, 407–411. [Google Scholar] [CrossRef]
- Dolinski, C.; Lacey, L.A. Microbial control of arthropod pests of tropical tree fruits. Neotrop. Entomol. 2007, 36, 161–179. [Google Scholar] [CrossRef]
- Lacey, L.A.; Shapiro-Ilan, D.I. Microbial control of insect pests in temperate orchard systems: Potential for incorporation into IPM. Annu. Rev. Entomol. 2008, 53, 121–144. [Google Scholar] [CrossRef]
- Qayyum, M.A.; Wakil, W.; Arif, M.J.; Sahi, S.T.; Dunlap, C.A. Infection of Helicoverpa armigera by endophytic Beauveria bassiana colonizing tomato plants. Biol. Control 2015, 90, 200–207. [Google Scholar] [CrossRef]
- Roberts, D.W.; St Leger, R. Metarhizium spp., cosmopolitan insect-pathogenic fungi: Mycological aspects. Adv. Appl. Microbiol. 2004, 54, 1–70. [Google Scholar] [CrossRef]
- Tahir, M.; Wakil, W.; Ali, A.; Gen, S.S.E. Pathogenicity of Beauveria bassiana and Metarhizium anisopliae isolates against larvae of the polyphagous pest Helicoverpa armigera. Entomol. Gen. 2019, 38, 225–242. [Google Scholar] [CrossRef]
- Veen, K.H. Recherches Sur La Maladie, Due Al Metarrhizium anisopliae Chez Le Criquet Pelerin; Wageningen University and Research: Wageningen, The Netherlands, 1968. [Google Scholar]
- Hussein, K.A.; Abdel Rahman, M.A.A.; Abdel-Mallek, A.Y.; El Maraghy, S.S.; Joo, J.H. Climatic factors interference with the occurrence of Beauveria bassiana and Metarhizium anisopliae in cultivated soil. Afr. J. Biotechnol. 2010, 9, 7674–7682. [Google Scholar] [CrossRef]
- Khan, S.; Nadir, S.; Lihua, G.; Xu, J.; Holmes, K.A.; Dewen, Q. Identification and characterization of an insect toxin protein, Bb70p, from the entomopathogenic fungus, Beauveria bassiana, using Galleria mellonella as a model system. J. Invertebr. Pathol. 2016, 133, 87–94. [Google Scholar] [CrossRef]
- Wang, J.; Ying, S.H.; Hu, Y.; Feng, M.G. Mas5, a homologue of bacterial DnaJ, is indispensable for the host infection and environmental adaptation of a filamentous fungal insect pathogen. Environ. Microbiol. 2015, 18, 1037–1047. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.J.; Sun, H.H.; Zhu, X.G.; Ying, S.H.; Feng, M.G. Discovery of a new intravacuolar protein required for the autophagy, development and virulence of Beauveria bassiana. Environ. Microbiol. 2017, 19, 2806–2818. [Google Scholar] [CrossRef] [PubMed]
- Sufyan, M.; Abbasi, A.; Wakil, W.; Gogi, M.D.; Arshad, M.; Nawaz, A.; Shabbir, Z. Efficacy of Beauveria bassiana and Bacillus thuringiensis against maize stem borer Chilo partellus (Swinhoe) (Lepidoptera: Pyralidae). Gesunde Pflanzen. 2019, 71, 197–204. [Google Scholar] [CrossRef]
- Nakahara, Y.; Shimura, S.; Ueno, C.; Kanamori, Y.; Mita, K.; Kiuchi, M.; Kamimura, M. Purification and characterization of silkworm hemocytes by flow cytometry. Dev. Comp. Immunol. 2009, 33, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Chelico, L.; Khachatourians, G.G. Isolation and characterization of nucleotide excision repair deficient mutants of the entomopathogenic fungus, Beauveria bassiana. J. Invertebr. Pathol. 2008, 98, 93–100. [Google Scholar] [CrossRef]
- Abdel-Raheem, M.A.; Ismail, I.A.; Abdel Rahman, R.S.; Farag, N.A.; Abdel Rhman, I.E. Entomopathogenic fungi, Beauveria bassiana (Bals.) and Metarhizium anisopliae (Metsch.) as biological control agents on some stored product insects. J. Entomol. Zool. Stud. 2015, 3, 316–320. [Google Scholar]
- Mantzoukas, S.; Lagogiannis, I.; Kitsiou, F.; Eliopoulos, P.A. Entomopathogenic action of wild fungal strains against stored product beetle pests. Insects 2023, 14, 91. [Google Scholar] [CrossRef]
- Meikle, W.; Mercadier, G.; Holst, N.; Nansen, C.; Girod, V. Impact of a treatment of Beauveria bassiana (Deuteromycota: Hyphomycetes) on honeybee (Apis mellifera) colony health and on Varroa destructor mites. Apidologie 2008, 39, 247–259. [Google Scholar] [CrossRef]
- Kanga, L.H.B.; Jones, A.W.; James, R.R. Field trials using the fungal pathogen, Beauveria bassiana (Deuteromycetes: Hyphomycetes) to control the ectoparasitic mite, Varroa destructor (Acari: Varroidae) in honey bee, Apis mellifera (Hymenoptera: Apidae) colonies. J. Econ. Entomol. 2003, 96, 1091–1099. [Google Scholar] [CrossRef]
- Ummidi, V.R.S.; Vadlamani, P. Preparation and use of oil formulations of Beauveria bassiana and Metarrhizium anisopliae against Spodoptera litura larvae. Afr. J. Microbiol. Res. 2014, 8, 1638–1644. [Google Scholar] [CrossRef]
- Freed, S.; Saleem, M.; Khan, M.; Naeem, M. Prevalence and effectiveness of Metarhizium anisopliae against Spodoptera exigua (Lepidoptera: Noctuidae) in southern Punjab, Pakistan. Pak. J. Zool. 2012, 44, 753–758. [Google Scholar]
- Petlamul, W.; Prasertsan, P. Evaluation of strains of Metarhizium anisopliae and Beauveria bassiana against Spodoptera litura on the basis of their virulence, germination rate, conidia production, radial growth and enzyme activity. Mycobiology 2012, 40, 111–116. [Google Scholar] [CrossRef]
- Hussein, K.A.; Abdel-Rahman, M.A.A.; Abdel-Mallek, A.Y.; El-Maraghy, S.S.; Joo, J.H. Pathogenicity of Beauveria bassiana and Metarhizium anisopliae against Galleria mellonella. Phytoparasitica 2012, 40, 117–126. [Google Scholar] [CrossRef]
- Bukhari, T.; Takken, W.; Koenraadt, C.J.M. Development of Metarhizium anisopliae and Beauveria bassiana formulations for control of Malaria mosquito larvae. Parasites Vectors 2011, 4, 23. [Google Scholar] [CrossRef]
- Lee, J.Y.; Woo, R.M.; Choi, C.J.; Shin, T.Y.; Gwak, W.S.; Woo, S.D. Beauveria bassiana for the simultaneous control of Aedes albopictus and Culex pipiens mosquito adults shows high conidia persistence and productivity. AMB Express 2019, 9, 206. [Google Scholar] [CrossRef]
- Bitencourt, R.D.O.B.; Santos-Mallet, J.R.D.; Lowenberger, C.; Ventura, A.; Gôlo, P.S.; Bittencourt, V.R.E.P.; Angelo, I.D.C. A novel model of pathogenesis of Metarhizium anisopliae propagules through the midguts of Aedes aegypti larvae. Insect 2023, 14, 328. [Google Scholar] [CrossRef]
- Dong, C.; Zhang, J.; Huang, H.; Chen, W.; Hu, Y. Pathogenicity of a new China variety of Metarhizium anisopliae (M. anisopliae Var. Dcjhyium) to subterranean termite Odontotermes formosanus. Microbiol. Res. 2009, 164, 27–35. [Google Scholar] [CrossRef]
- Rios-Velasco, C.; Pérez-Corral, D.A.; Salas-Marina, M.Á.; Berlanga-Reyes, D.I.; Ornelas-Paz, J.J.; Muñiz, C.H.A.; Cambero-Campos, J.; Jacobo-Cuellar, J.L. Pathogenicity of the hypocreales fungi Beauveria bassiana and Metarhizium anisopliae against insect pests of tomato. Southwest. Entomol. 2014, 39, 739–750. [Google Scholar] [CrossRef]
- Rivero-Borja, M.; Guzmán-Franco, A.W.; Rodríguez-Leyva, E.; Santillán-Ortega, C.; Pérez-Panduro, A. Interaction of Beauveria bassiana with chlorpyrifos ethyl and spinosad in Spodoptera frugiperda larvae. Pest Manag. Sci. 2018, 74, 2047–2052. [Google Scholar] [CrossRef]
- Quintela, E.; Mccoy, C.W. Pathogenicity enhancement of Metarhizium anisopliae and Beauveria bassiana to first instars of Diaprepes abbreviatus (Coleoptera: Curculionidae) with sublethal doses of imidacloprid. Environ. Entomol. 1997, 26, 1173–1182. [Google Scholar] [CrossRef]
- Brito, E.S.; De Paula, A.R.; Vieira, L.P.; Dolinski, C.; Samuels, R.I. Combining vegetable oil and sub-lethal concentrations of imidacloprid with Beauveria bassiana and Metarhizium anisopliae against adult Guava weevil Conotrachelus psidii (Coleoptera: Curculionidae). Biocontrol Sci. Technol. 2008, 18, 665–673. [Google Scholar] [CrossRef]
- Wendel, J.; Cisneros, J.; Jaronski, S.; Vitek, C.; Ciomperlik, M.; Flores, D. Screening commercial entomopathogenic fungi for the management of Diaphorina citri Populations in the lower Rio Grande Valley, Texas, USA. BioControl 2022, 67, 225–235. [Google Scholar] [CrossRef]
- Bava, R.; Castagna, F.; Piras, C.; Musolino, V.; Lupia, C.; Palma, E.; Britti, D.; Musella, V. Entomopathogenic fungi for pests and predators control in beekeeping. Vet. Sci. 2022, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.C.; Wu, S.S.; Liu, Y.C.; Yang, Y.H.; Tsai, Y.F.; Li, Y.H.; Tseng, C.T.; Tang, L.C.; Nai, Y.S. Construction and selection of an entomopathogenic fungal library from soil samples for controlling Spodoptera litura. Front. Sustain. Food Syst. 2021, 5, 596316. [Google Scholar] [CrossRef]
- De Souza, D.A.; Lopes, R.B.; Humber, R.; Faria, M. Assessment of the diversity of Brazilian entomopathogenic fungi in the genus Beauveria. J. Invertebr. Pathol. 2020, 171, 107339. [Google Scholar] [CrossRef]
- Mathulwe, L.L.; Malan, A.P.; Stokwe, N.F. Mass production of entomopathogenic fungi, Metarhizium robertsii and Metarhizium pingshaense, for commercial application against insect pests. J. Vis. Exp. 2022, 181, e63246. [Google Scholar] [CrossRef]
- Rice, S.J.; Baker, D.K.; Mayer, D.G.; Leemon, D.M. Mycoinsecticide formulations of Beauveria bassiana and Metarhizium anisopliae reduce populations of lesser mealworm, Alphitobius diaperinus, in chicken-broiler houses. Biol. Control 2020, 144, 104234. [Google Scholar] [CrossRef]
- Kassa, A.; Brownbridge, M.; Parker, B.L.; Skinner, M.; Gouli, V.; Gouli, S.; Guo, M.; Lee, F.; Hata, T. Whey for mass production of Beauveria bassiana and Metarhizium anisopliae. Mycol. Res. 2008, 112, 583–591. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Babalola, O.O. Exploring the potentialities of beneficial endophytes for improved plant growth. Saudi J. Biol. Sci. 2020, 27, 3622–3633. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Siddiqui, J.A.; Akutse, K.S.; Aguila, L.C.R.; Xu, Y. General limitations to endophytic entomopathogenic fungi use as plant growth promoters, pests and pathogens biocontrol agents. Plants 2021, 10, 2119. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Akutse, K.S.; Siddiqui, J.A.; Xu, Y. Model application of entomopathogenic fungi as alternatives to chemical pesticides: Prospects, challenges, and insights for next-generation sustainable agriculture. Front. Plant Sci. 2021, 12, 2132. [Google Scholar] [CrossRef]
- Biedermann, P.H.W. Cooperative breeding in the Ambrosia beetle Xyleborus affinis and management of its fungal symbionts. Front. Ecol. Evol. 2020, 8, 518954. [Google Scholar] [CrossRef]
- Hulcr, J.; Stelinski, L.L. The ambrosia symbiosis: From evolutionary ecology to practical management. Annu. Rev. Entomol. 2017, 62, 285–303. [Google Scholar] [CrossRef]
- Sobhy, I.S.; Baets, D.; Goelen, T.; Herrera-Malaver, B.; Bosmans, L.; Van den Ende, W.; Verstrepen, K.J.; Wäckers, F.; Jacquemyn, H.; Lievens, B. Sweet scents: Nectar specialist yeasts enhance nectar attraction of a generalist aphid parasitoid without affecting survival. Front. Plant Sci. 2018, 9, 1009. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Pozo, M.I.; ÁLvarez-Pérez, S.A.; Lievens, B.; Fukami, T. Yeast–nectar interactions: Metacommunities and effects on pollinators. Curr. Opin. Insect Sci. 2021, 44, 35–40. [Google Scholar] [CrossRef]
- Nicoletti, R.; Becchimanzi, A. Ecological and molecular interactions between insects and fungi. Microorganisms 2022, 10, 96. [Google Scholar] [CrossRef]
- Samaras, K.; Mourtiadou, S.; Arampatzis, T.; Kakagianni, M.; Feka, M.; Wäckers, F.; Papadopoulou, K.K.; Broufas, G.D.; Pappas, M.L. Plant-mediated effects of beneficial microbes and a plant strengthener against spider mites in tomato. Plants 2023, 12, 938. [Google Scholar] [CrossRef]
- Coppola, M.; Cascone, P.; Di Lelio, I.; Woo, S.L.; Lorito, M.; Rao, R.; Pennacchio, F.; Guerrieri, E.; Digilio, M.C. Trichoderma atroviride P1 colonization of tomato plants enhances both direct and indirect defense barriers against insects. Front. Physiol. 2019, 10, 813. [Google Scholar] [CrossRef]
- Coppola, M.; Cascone, P.; Chiusano, M.L.; Colantuono, C.; Lorito, M.; Pennacchio, F.; Rao, R.; Woo, S.L.; Guerrieri, E.; Digilio, M.C. Trichoderma harzianum enhances tomato indirect defense against aphids. Insect Sci. 2017, 24, 1025–1033. [Google Scholar] [CrossRef]
- González-Mas, N.; Gutiérrez-Sánchez, F.; Sánchez-Ortiz, A.; Grandi, L.; Turlings, T.C.J.; Manuel Muñoz-Redondo, J.; Moreno-Rojas, J.M.; Quesada-Moraga, E. Endophytic colonization by the entomopathogenic fungus Beauveria bassiana affects plant volatile emissions in the presence or absence of chewing and sap-sucking insects. Front. Plant Sci. 2021, 12, 660460. [Google Scholar] [CrossRef]
- Dicke, M.; Baldwin, I.T. The evolutionary context for herbivore-induced plant volatiles: Beyond the ‘cry for help’. Trends Plant Sci. 2010, 15, 167–175. [Google Scholar] [CrossRef]
- Bezemer, T.M.; van Dam, N.M. Linking aboveground and belowground interactions via induced plant defenses. Trends Ecol. Evol. 2005, 20, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Zarate, S.I.; Kempema, L.A.; Walling, L.L. Silverleaf whitefly induces salicylic acid defenses and suppresses effectual jasmonic acid defenses. Plant Physiol. 2007, 143, 866–875. [Google Scholar] [CrossRef]
- Kuśnierczyk, A.; Winge, P.E.R.; Jørstad, T.S.; Troczyńska, J.; Rossiter, J.T.; Bones, A.M. Towards global understanding of plant defence against aphids–timing and dynamics of early Arabidopsis defence responses to cabbage aphid (Brevicoryne brassicae) attack. Plant Cell Environ. 2008, 31, 1097–1115. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone crosstalk in plant disease and defense: More than just more than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Giron, D.; Frago, E.; Glevarec, G.; Pieterse, C.M.; Dicke, M. Cytokinins as key regulators in plant–microbe–insect interactions: Connecting plant growth and defence. Funct. Ecol. 2013, 27, 599–609. [Google Scholar] [CrossRef]
- Navarro, L.; Bari, R.; Achard, P.; Lisón, P.; Nemri, A.; Harberd, N.P.; Jones, J.D. DELLAs control plant immune responses by modulating the balance of jasmonic acid and salicylic acid signaling. Curr. Biol. 2008, 18, 650–655. [Google Scholar] [CrossRef]
- Ryu, H.; Cho, H.; Choi, D.; Hwang, I. Plant hormonal regulation of nitrogen-fixing nodule organogenesis. Mol. Cells 2012, 34, 117–126. [Google Scholar] [CrossRef]
- Cosme, M.; Wurst, S. Interactions between arbuscular mycorrhizal fungi, rhizobacteria, soil phosphorus and plant cytokinin deficiency change the root morphology, yield and quality of tobacco. Soil Biol. Biochem. 2013, 57, 436–443. [Google Scholar] [CrossRef]
- Liu, J.; Lovisolo, C.; Schubert, A.; Cardinale, F. Signaling role of strigolactones at the interface between plants, (micro)organisms, and a changing environment. J. Plant Interact. 2013, 8, 17–33. [Google Scholar] [CrossRef]
- Pangesti, N.; Pineda, A.; Pieterse, C.M.J.; Dicke, M.; Loon, J.J.A. Two-way plant-mediated interactions between root-associated microbes and insects: From ecology to mechanisms. Front. Plant Sci. 2013, 4, 414. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, S.; Yadav, P.K. Entomopathogenic fungi and their relevance in sustainable agriculture: A review. Cogent Food Agric. 2023, 9, 2180857. [Google Scholar] [CrossRef]
- Halder, J.; Rai, A.B.; Kodandaram, M.H. Compatibility of neem oil and different entomopathogens for the management of major vegetable sucking pests. Natl. Acad. Sci. Lett. 2013, 36, 19–25. [Google Scholar] [CrossRef]
- Padilla-Guerrero, I.E.; Barelli, L.; González-Hernández, G.A.; Torres-Guzmán, J.C.; Bidochka, M.J. Flexible metabolism in Metarhizium anisopliae and Beauveria bassiana: Role of the glyoxylate cycle during insect pathogenesis. Microbiology 2011, 157, 199–208. [Google Scholar] [CrossRef]
- Sutanto, K.D.; Husain, M.; Rasool, K.G.; Al-Qahtani, W.H.; Aldawood, A.S. Pathogenicity of local and exotic entomopathogenic fungi isolates against different life stages of red palm weevil (Rhynchophorus ferrugineus). PLoS ONE 2021, 16, e0255029. [Google Scholar] [CrossRef]
- Boucias, D.G.; Zhou, Y.; Huang, S.; Keyhani, N.O. Microbiota in insect fungal pathology. Appl. Microbiol. Biotechnol. 2018, 102, 5873–5888. [Google Scholar] [CrossRef]
- You, L.; Simmons, D.R.; Bateman, C.C.; Short, D.P.G.; Kasson, M.T.; Rabaglia, R.J.; Hulcr, J. New fungus-insect symbiosis: Culturing, molecular, and histological methods determine Saprophytic Polyporales mutualists of Ambrosiodmus Ambrosia beetles. PLoS ONE 2015, 11, e0147305. [Google Scholar] [CrossRef]
- Blackwell, M. Made for each other: Ascomycete yeasts and insects. Microbiol. Spectr. 2017, 5, FUNK-0081-2016. [Google Scholar] [CrossRef]
- Wei, G.; Lai, Y.; Wang, G.; Chen, H.; Li, F.; Wang, S. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc. Natl. Acad. Sci. USA 2017, 114, 5994–5999. [Google Scholar] [CrossRef]
- Xu, L.; Deng, J.; Zhou, F.; Cheng, C.; Zhang, L.; Zhang, J.; Lu, M. Gut microbiota in an invasive bark beetle infected by a pathogenic fungus accelerates beetle mortality. J. Pest Sci. 2019, 92, 343–351. [Google Scholar] [CrossRef]
- Wang, Z.; Cheng, Y.; Wang, Y.; Yu, X. Topical fungal infection induces shifts in the gut microbiota structure of brown planthopper, Nilaparvata lugens (Homoptera: Delphacidae). Insects 2022, 13, 528. [Google Scholar] [CrossRef] [PubMed]
- Stączek, S.; Zdybicka-barabas, A.; Wojda, I.; Wiater, A.; Mak, P.; Suder, P.; Skrzypiec, K.; Cytryńska, M. Fungal α-1,3-Glucan as a new pathogen-associated molecular pattern in the insect model host Galleria mellonella. Molecules 2021, 26, 5097. [Google Scholar] [CrossRef] [PubMed]
- Mburu, D.M.; Ochola, L.; Maniania, N.K.; Njagi, P.G.N.; Gitonga, L.M.; Ndung’u, M.W.; Wanjoya, A.K.; Hassanali, A. Relationship between virulence and repellency of entomopathogenic isolates of Metarhizium anisopliae and Beauveria bassiana to the termite Macrotermes michaelseni. J. Insect Physiol. 2009, 55, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Meyling, N.V.; Pell, J.K.; Eilenberg, J. Dispersal of Beauveria bassiana by the activity of nettle insects. J. Invertebr. Pathol. 2006, 93, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Blomquist, G.J.; Tillman, J.A.; Mpuru, S.; Seybold, S.J. The cuticle and cuticular hydrocarbons of insects: Structure, function, and biochemistry. In Pheromone Communication in Social Insects; Vander Meer, R.K., Breed, M.D., Winston, M., Espelie, K.E., Eds.; CRC Press: New York, NY, USA, 1998; pp. 34–54. [Google Scholar]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef]
- Charnley, A.K. Fungal pathogens of insects: Cuticle degrading enzymes and toxins. Adv. Bot. Res. 2003, 40, 241–321. [Google Scholar] [CrossRef]
- Roberts, D.W.; Hajek, A.E. Entomopathogenic fungi as bioinsecticides. In Frontiers in Industrial Mycology; Leatham, G.F., Ed.; Springer Science & Business Media: Berlin/Heildelberg, Germany, 1992; pp. 144–159. [Google Scholar]
- Uchida, R.; Imasato, R.; Yamaguchi, Y.; Masuma, R.; Shiomi, K.; Tomoda, H.; Ōmura, S. New insecticidal antibiotics, hydroxyfungerins A and B, produced by Metarhizium sp. FKI-1079. J. Antibiot. 2005, 58, 804–809. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungus Metarhizium anisopliae. Biocontrol Sci. Technol. 2007, 17, 879–920. [Google Scholar] [CrossRef]
- Stleger, R.; Bidochka, M.J.; Roberts, D.W. Isoforms of the cuticle-degrading Pr1 proteinase and production of a metalloproteinase by Metarhizium anisopliae. Arch. Biochem. Biophys. 1994, 313, 1–7. [Google Scholar] [CrossRef]
- Schrank, A.; Vainstein, M.H. Beauveria bassiana enzymes and toxins. Toxicon 2010, 56, 1267–1274. [Google Scholar] [CrossRef]
- Strand, M.R. The insect cellular immune response. Insect Sci. 2008, 15, 1–14. [Google Scholar] [CrossRef]
- Ali Mohammadie Kojour, M.; Han, Y.S.; Jo, Y.H. An overview of insect innate immunity. Entomol. Res. 2020, 50, 282–291. [Google Scholar] [CrossRef]
- Kingsolver, M.B.; Huang, Z.; Hardy, R.W. Insect antiviral innate immunity: Pathways, effectors, and connections. J. Mol. Biol. 2013, 425, 4921–4936. [Google Scholar] [CrossRef]
- Tsakas, S.; Marmaras, V.J. Insect immunity and its signalling: An overview. Invertebr. Surviv. J. 2010, 7, 228–238. [Google Scholar]
- Zhang, Z.T.; Zhu, S.Y. Drosomycin, an essential component of antifungal defence in Drosophila. Wiley Online Libr. 2009, 18, 549–556. [Google Scholar] [CrossRef]
- Cherry, S.; Silverman, N. Host-pathogen interactions in Drosophila: New tricks from an old friend. Nat. Immunol. 2006, 7, 911–917. [Google Scholar] [CrossRef]
- Elrod-Erickson, M.; Mishra, S.; Schneider, D. Interactions between the cellular and humoral immune responses in Drosophila. Curr. Biol. 2000, 10, 781–784. [Google Scholar] [CrossRef]
- Haine, E.R.; Moret, Y.; Siva-Jothy, M.T.; Rolff, J. Antimicrobial defense and persistent infection in insects. Science 2008, 322, 1257–1259. [Google Scholar] [CrossRef]
- Wang, J.L.; Yang, K.H.; Wang, S.S.; Li, X.L.; Liu, J.; Yu, Y.X.; Liu, X.S. Infection of the entomopathogenic fungus Metarhizium rileyi suppresses cellular immunity and activates Humoral antibacterial immunity of the host Spodoptera frugiperda. Pest Manag. Sci. 2022, 78, 2828–2837. [Google Scholar] [CrossRef]
- Li, S.; Liu, F.; Kang, Z.; Li, X.; Lu, Y.; Li, Q.; Pang, Y.; Zheng, F.; Yin, X. Cellular immune responses of the yellow peach moth, Conogethes punctiferalis (Lepidoptera: Crambidae), to the entomopathogenic fungus, Beauveria bassiana (Hypocreales: Cordycipitaceae). J. Invertebr. Pathol. 2022, 194, 107826. [Google Scholar] [CrossRef]
- Söderhäll, K.; Ajaxon, R. Effect of quinones and melanin on mycelial growth of Aphanomyces spp. and extracellular protease of Aphanomyces astaci, a parasite on crayfish. J. Invertebr. Pathol. 1982, 39, 105–109. [Google Scholar] [CrossRef]
- Lu, H.L.; Leger, R.S. Insect immunity to entomopathogenic fungi. Adv. Genet. 2016, 94, 251–285. [Google Scholar] [CrossRef] [PubMed]
- Thak, E.J.; Lee, S.B.; Xu-Vanpala, S.; Lee, D.J.; Chung, S.Y.; Bahn, Y.S.; Oh, D.B.; Shinohara, M.L.; Kang, H.A. Core N-glycan structures are critical for the pathogenicity of Cryptococcus neoformans by modulating host cell death. mBio 2020, 11, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.R.; Lenardon, M.D. Architecture of the dynamic fungal cell wall. Nat. Rev. Microbiol. 2022, 21, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, J.; Ortiz-Castellanos, L. Cell wall glucans of fungi. A review. Cell Surf. 2019, 5, 100022. [Google Scholar] [CrossRef]
- Fernando, L.D.; Dickwella Widanage, M.C.; Penfield, J.; Lipton, A.S.; Washton, N.; Latgé, J.P.; Wang, P.; Zhang, L.; Wang, T. Structural polymorphism of chitin and chitosan in fungal cell walls from solid-state NMR and principal component analysis. Front. Mol. Biosci. 2021, 8, 814. [Google Scholar] [CrossRef]
- Gow, N.A.R.; Latge, J.P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5, 5–13. [Google Scholar] [CrossRef]
- Lipke, P.N.; Ovalle, R. Cell wall architecture in yeast: New structure and new challenges. J. Bacteriol. 1998, 180, 3735–3740. [Google Scholar] [CrossRef]
- Schäffer, C.; Messner, P. Emerging facets of prokaryotic glycosylation. FEMS Microbiol. Rev. 2017, 41, 49–91. [Google Scholar] [CrossRef]
- De Pourcq, K.; De Schutter, K.; Callewaert, N. Engineering of glycosylation in yeast and other fungi: Current state and perspectives. Appl. Microbiol. Biotechnol. 2010, 87, 1617–1631. [Google Scholar] [CrossRef]
- Chung, C.Y.; Majewska, N.I.; Wang, Q.; Paul, J.T.; Betenbaugh, M.J. SnapShot: N-glycosylation processing pathways across kingdoms. Cell 2017, 171, 258. [Google Scholar] [CrossRef]
- Yan, A.; Lennarz, W.J. Unraveling the mechanism of protein N-glycosylation. J. Biol. Chem. 2005, 280, 3121–3124. [Google Scholar] [CrossRef]
- Garcia-Rubio, R.; De Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The Fungal cell wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef]
- Liu, D.D.; De Schutter, K.; Smargiass, N.; De Pauw, E.; Van Damme, E.J.M.; Smagghe, G. The N-glycan profile of the peritrophic membrane in the Colorado potato beetle larvae (Leptinotarsa decemlineata). J. Insect Physiol. 2019, 115, 27–32. [Google Scholar] [CrossRef]
- Van de Veerdonk, F.L.; Marijnissen, R.J.; Kullberg, B.J.; Koenen, H.J.; Cheng, S.C.; Joosten, I.; Van den Berg, W.B.; Williams, D.L.; Van der Meer, J.W.; Joosten, L.A.; et al. The macrophage mannose receptor induces IL-17 in response to Candida albicans. Cell Host Microbe 2009, 5, 329–340. [Google Scholar] [CrossRef]
- Schirmer, M.; Smeekens, S.P.; Vlamakis, H.; Jaeger, M.; Oosting, M.; Franzosa, E.A.; Jansen, T.; Jacobs, L.; Bonder, M.J.; Kurilshikov, A.; et al. Linking the human gut microbiome to inflammatory cytokine production capacity. Cell 2016, 167, 1125–1136. [Google Scholar] [CrossRef]
- Walski, T.; Van Damme, E.J.M.; Smargiasso, N.; Christiaens, O.; De Pauw, E.; Smagghe, G. Protein N-glycosylation and N-glycan trimming are required for postembryonic development of the pest beetle Tribolium castaneum. Sci. Rep. 2016, 6, 35151. [Google Scholar] [CrossRef]
- Liu, D.; De Schutter, K.; Far, J.; Staes, A.; Dewettinck, K.; Quinton, L.; Gevaert, K.; Smagghe, G. RNAi of Mannosidase-Ia in the Colorado potato beetle and changes in the midgut and peritrophic membrane. Pest Manag. Sci. 2022, 78, 5071–5079. [Google Scholar] [CrossRef]
- Hakim, R.S.; Baldwin, K.; Smagghe, G. Regulation of midgut growth, development, and metamorphosis. Annu. Rev. Entomol. 2010, 55, 593–608. [Google Scholar] [CrossRef]
- Liu, D.; De Schutter, K.; Chen, P.; Smagghe, G. The N-glycosylation-related genes as potential targets for RNAi-mediated pest control of the Colorado potato beetle (Leptinotarsa decemlineata). Pest Manag. Sci. 2021, 78, 3815–3822. [Google Scholar] [CrossRef]
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA uptake in insects and potentials of RNAi for pest control: A review. J. Insect Physiol. 2010, 56, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-J.; Shi, Y.; Wu, J.-N.; Li, H.; Smagghe, G.; Liu, T.-X. CRISPR/Cas9 in Lepidopteran insects: Progress, application and prospects. J. Insect Physiol. 2021, 135, 104325. [Google Scholar] [CrossRef] [PubMed]

 —GlcNAc, N-acetylglucosamine,
—GlcNAc, N-acetylglucosamine,  —Man, mannose,
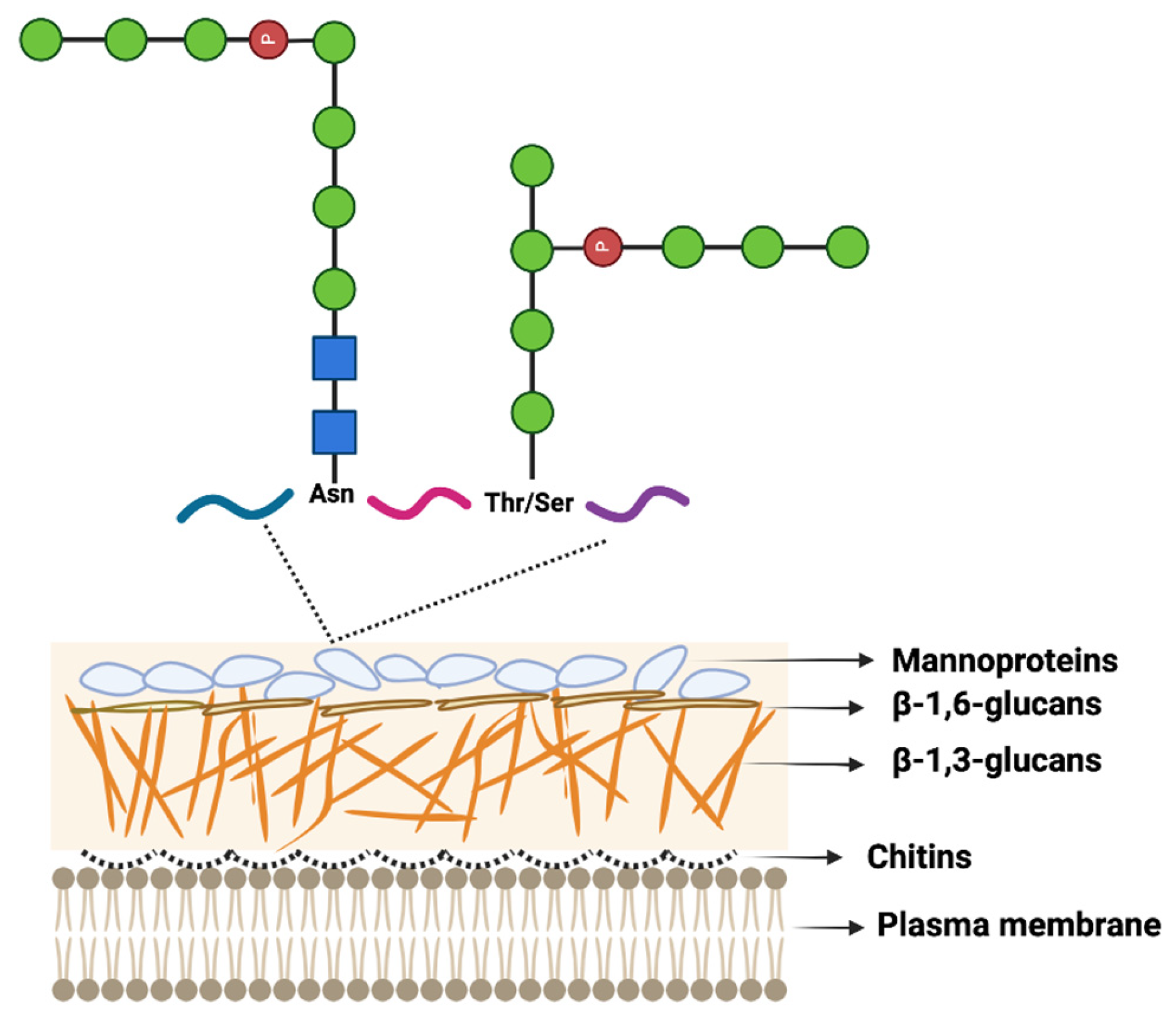
—Man, mannose,  —phosphate group. The fungal cell wall is composed of chitins (1.5–6% by weight), β-linked glucans (β-1,3-glucans (30–45% by weight), β-1,6 glucan (5–10% by weight)), and mannoproteins (30–50% by weight). Chitin is a structurally important component of the fungal cell wall located closest to the plasma membrane. Branched β-1,3 glucan cross-links to chitin and is covalently linked to other polysaccharides (e.g., galactomannan and β-1,6 glucan). Mannoproteins are N- and O-glycosylated proteins. The Figure was modified from [118] and drawn with BioRender.com.
—GlcNAc, N-acetylglucosamine, —Man, mannose, —phosphate group. The fungal cell wall is composed of chitins (1.5–6% by weight), β-linked glucans (β-1,3-glucans (30–45% by weight), β-1,6 glucan (5–10% by weight)), and mannoproteins (30–50% by weight). Chitin is a structurally important component of the fungal cell wall located closest to the plasma membrane. Branched β-1,3 glucan cross-links to chitin and is covalently linked to other polysaccharides (e.g., galactomannan and β-1,6 glucan). Mannoproteins are N- and O-glycosylated proteins. The Figure was modified from [118] and drawn with BioRender.com.
—phosphate group. The fungal cell wall is composed of chitins (1.5–6% by weight), β-linked glucans (β-1,3-glucans (30–45% by weight), β-1,6 glucan (5–10% by weight)), and mannoproteins (30–50% by weight). Chitin is a structurally important component of the fungal cell wall located closest to the plasma membrane. Branched β-1,3 glucan cross-links to chitin and is covalently linked to other polysaccharides (e.g., galactomannan and β-1,6 glucan). Mannoproteins are N- and O-glycosylated proteins. The Figure was modified from [118] and drawn with BioRender.com.
—GlcNAc, N-acetylglucosamine, —Man, mannose, —phosphate group. The fungal cell wall is composed of chitins (1.5–6% by weight), β-linked glucans (β-1,3-glucans (30–45% by weight), β-1,6 glucan (5–10% by weight)), and mannoproteins (30–50% by weight). Chitin is a structurally important component of the fungal cell wall located closest to the plasma membrane. Branched β-1,3 glucan cross-links to chitin and is covalently linked to other polysaccharides (e.g., galactomannan and β-1,6 glucan). Mannoproteins are N- and O-glycosylated proteins. The Figure was modified from [118] and drawn with BioRender.com.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EPF Species | Commercial Name | Strain | Producer | Target Pests | Regions to Use |
|---|---|---|---|---|---|
| B. bassiana * | BotaniGardTM ES | GHA | Laverlam | Whiteflies, aphids, weevils, mealybugs | Worldwide |
| BotaniGardTM MAXX | |||||
| MycotrolTM ESO | Certis USA | USA | |||
| MycotrolTM WPO | USA | ||||
| AprehendTM | Conidia Bioscience | USA and Canada | |||
| BroadbandTM | Bioworks | Whiteflies, thrips, moths, stinkbugs, red spider mite, red scale | Worldwide | ||
| BioBeeTM | PPRI 5339 | BioBee Biological Systems | Worldwide | ||
| BioforestTM | ANT-03 | BioForest Technologies | Spruce budworm | USA and Canada | |
| BioPalmTM | PL11 | BioPalm Manufacturing | Bagworms | Malaysia | |
| BoverinTM | ATCC 74040 | Boverin Europe | Europe | ||
| Boveril TM | ESALQ Tec | Whiteflies, weevils | Brazil | ||
| NaturalisTM | Koppert Biological Systems | Whiteflies, thrips, mites, aphids, tingids | Worldwide | ||
| BioCeresTM | ANT-03 | Bioceres | Whiteflies, aphids, thrips | Argentina, Chile, Peru, Mexico | |
| M. anisopliae * | ESALQ-E9 MetarrilTM | ESALQ-E9 | EMBRAPA | Several insect and mites | Brazil |
| NCIM 1311 PacerTM | NCIM 1311 | Chema Industries | Termites, root grub, locusts, root weevils, ants, beetles, caterpillar | Worldwide | |
| Bio-Blast Biological TermiticideTM | F52 | BioProdex | Ticks, weevils, mites, thrips | USA | |
| Tick-Ex TM ECTM | ICIPE 69 | Kenyan Biologics | Worldwide | ||
| Met52 TM ECTM | F52 | Novozymes BioAg | Worldwide | ||
| MAS-01TM | Sor-1 | Bio-Insumos Naturales | Coffee berry borer | USA | |
| Green MuscleTM | F52 | Bioworks Inc | thrips, whiteflies, and aphids | Worldwide | |
| Metarhizium ICIPE 7TM | ICIPE 7 | Kenya Biologics | East Africa | ||
| Metarhizium AQTM | CQMa421 | Sinochem | Citrus root weevil | China |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Smagghe, G.; Liu, T.-X. Interactions between Entomopathogenic Fungi and Insects and Prospects with Glycans. J. Fungi 2023, 9, 575. https://doi.org/10.3390/jof9050575
Liu D, Smagghe G, Liu T-X. Interactions between Entomopathogenic Fungi and Insects and Prospects with Glycans. Journal of Fungi. 2023; 9(5):575. https://doi.org/10.3390/jof9050575
Chicago/Turabian StyleLiu, Dongdong, Guy Smagghe, and Tong-Xian Liu. 2023. "Interactions between Entomopathogenic Fungi and Insects and Prospects with Glycans" Journal of Fungi 9, no. 5: 575. https://doi.org/10.3390/jof9050575
APA StyleLiu, D., Smagghe, G., & Liu, T. -X. (2023). Interactions between Entomopathogenic Fungi and Insects and Prospects with Glycans. Journal of Fungi, 9(5), 575. https://doi.org/10.3390/jof9050575