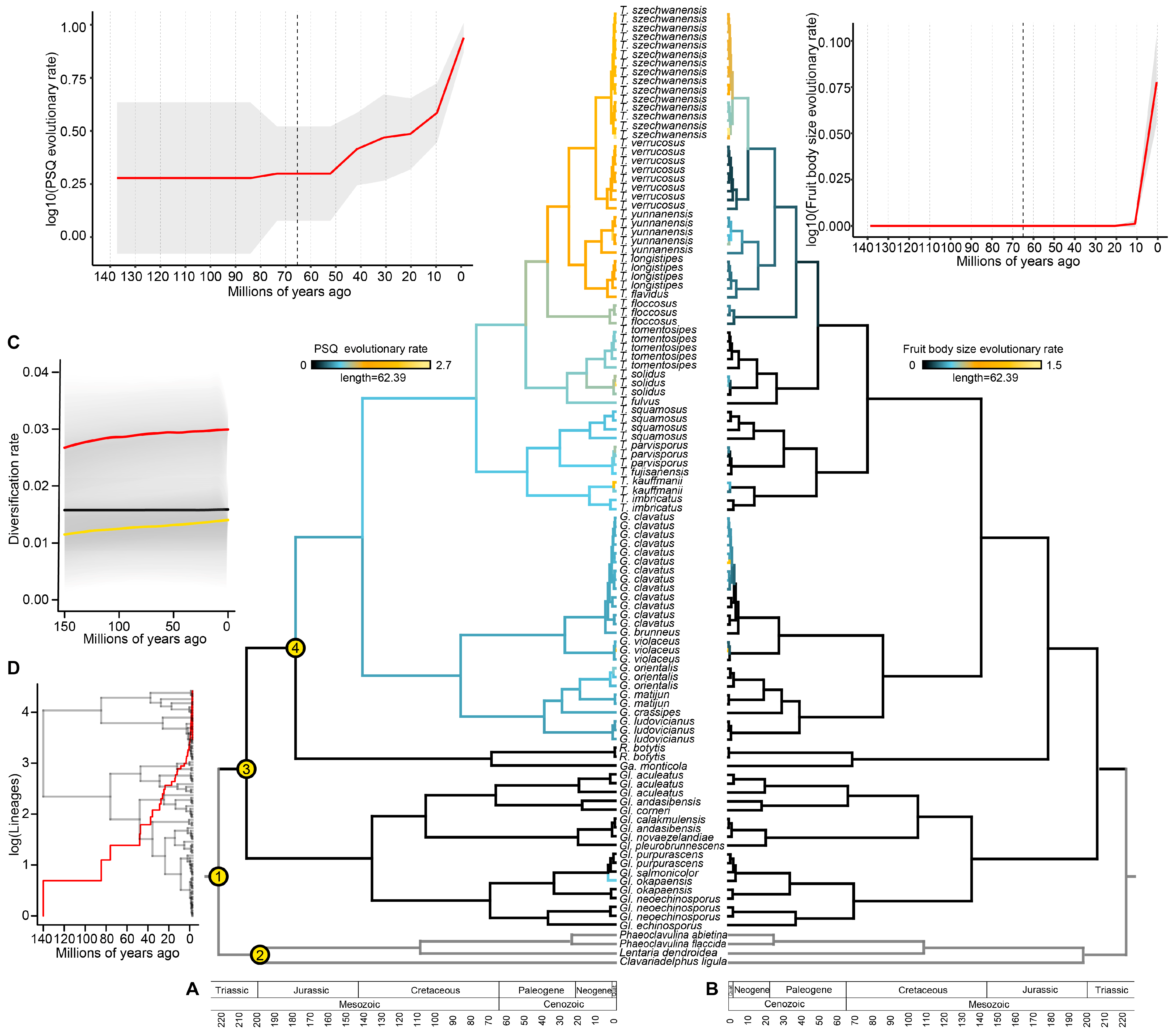
3.4. Taxonomy of Gomphoid Fungi in the Narrower Sense
Gomphus clavatus (Pers.) Gray, Nat. Arr. Brit. Pl. (London) 1: 638 (1821). (
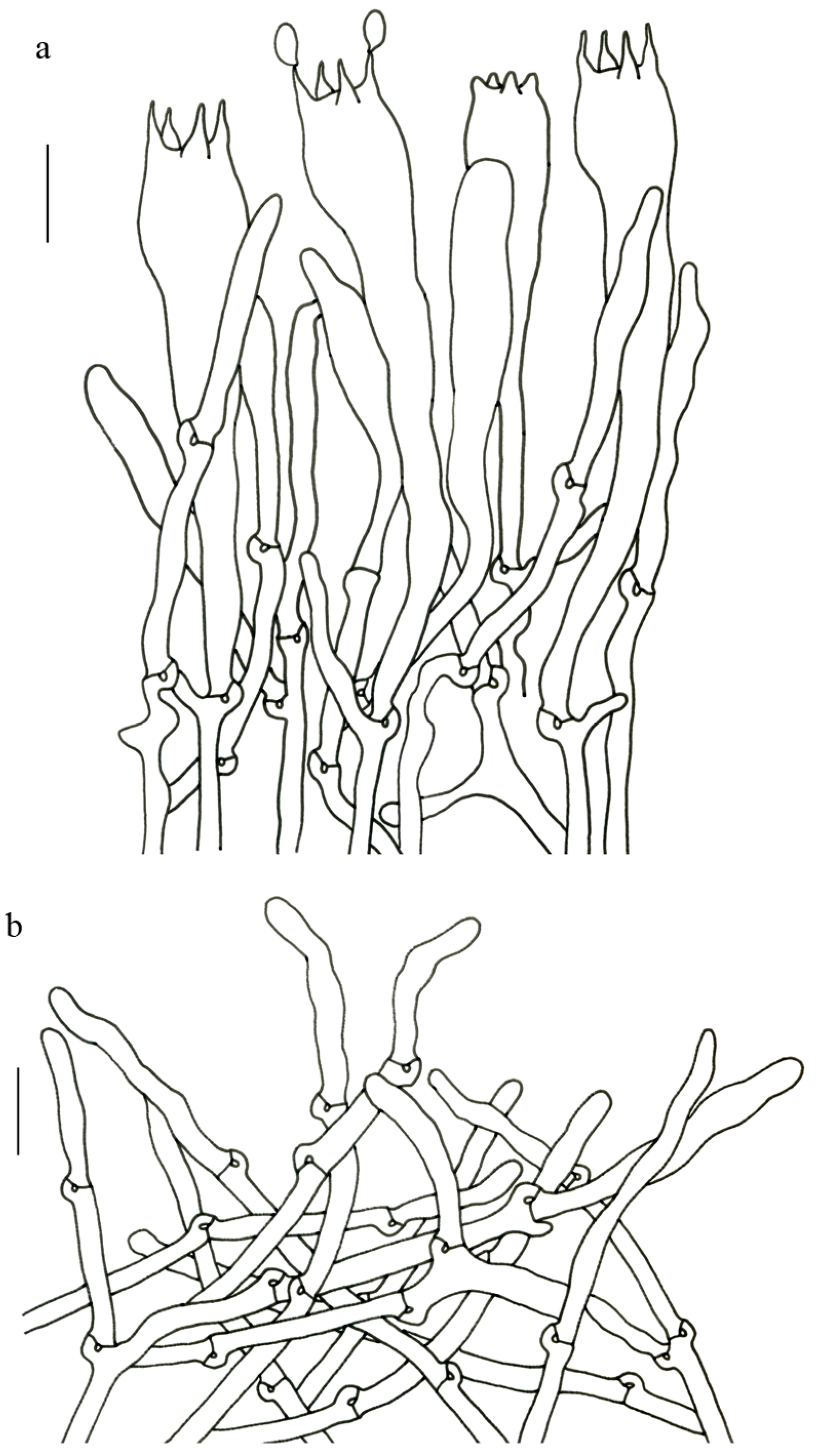
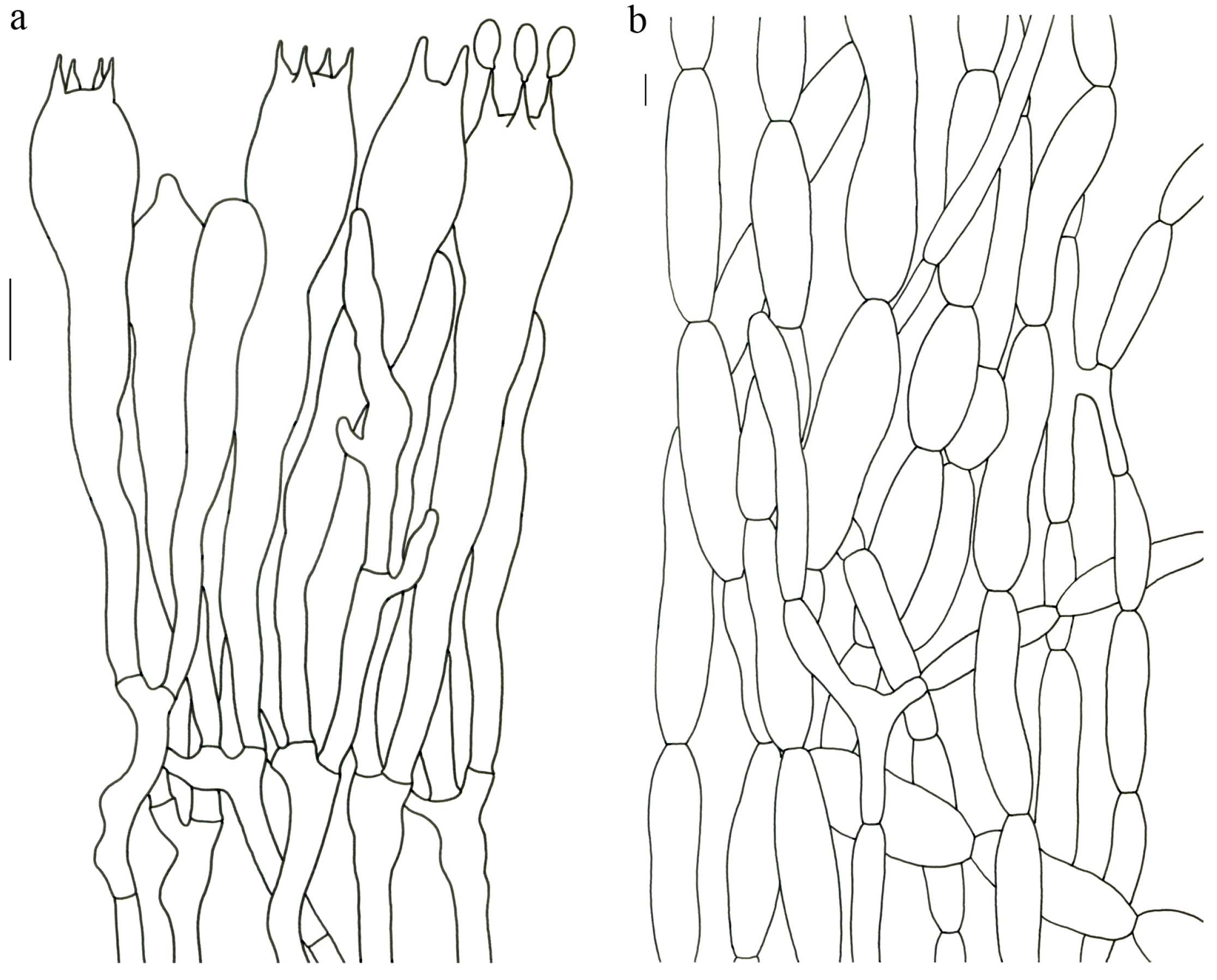
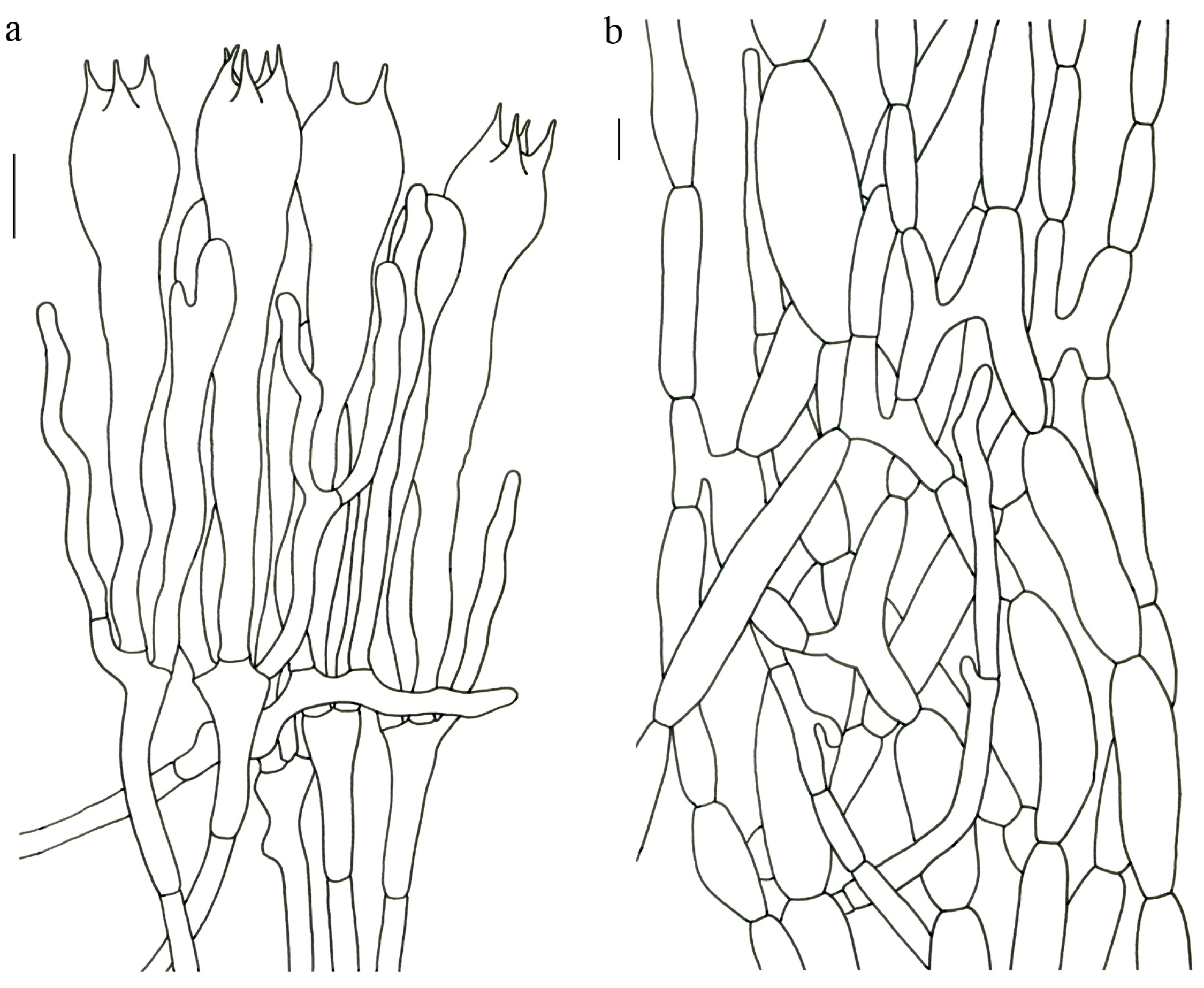
Figure 4).
Basionym—Merulius clavatus Pers., Observationes Mycologicae 1:21 (1796).
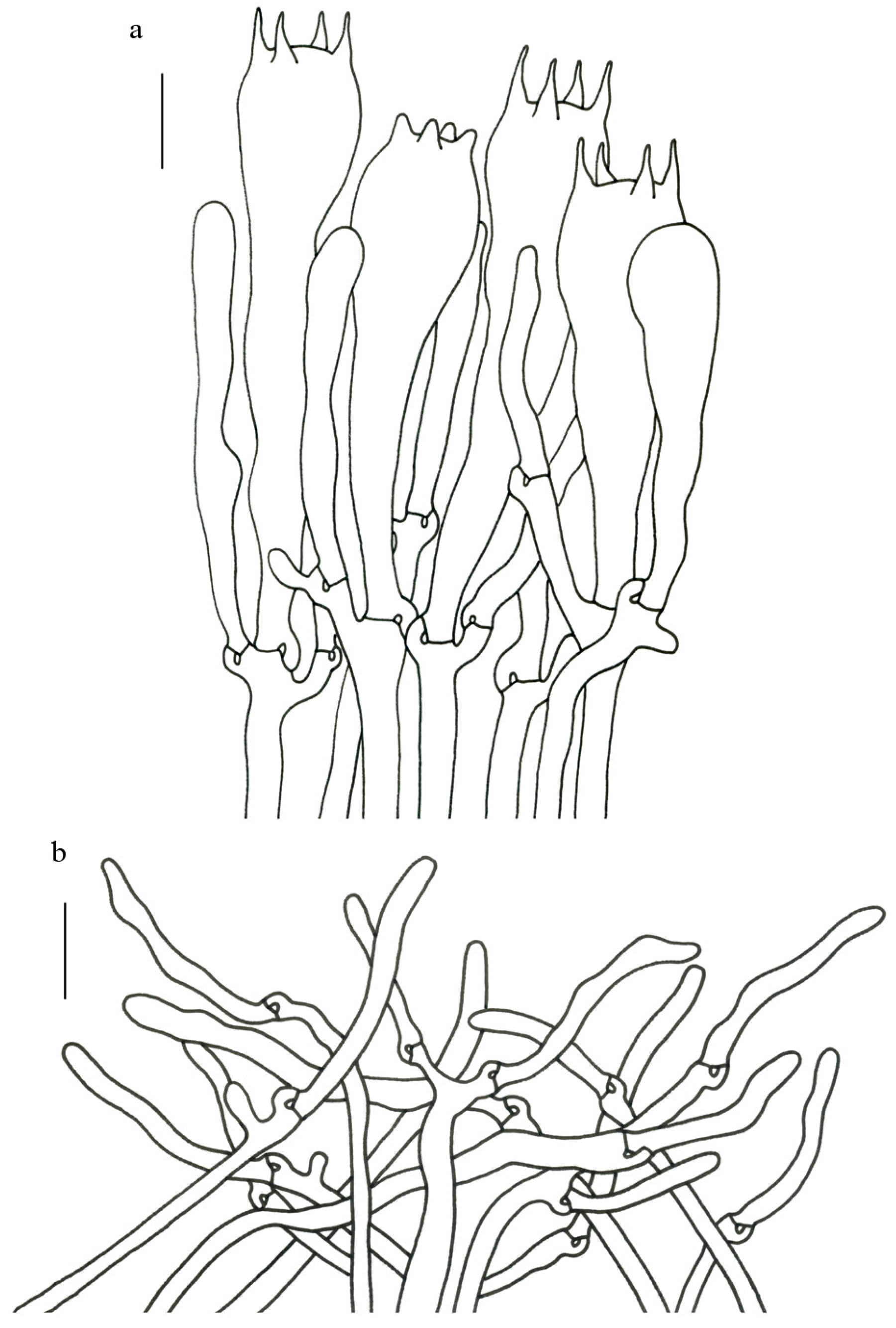
Description—Basidioma up to 17 cm tall, unipileate at base and then merismatoid with 5–15 subpilei. Pileus up to 15 cm wide; irregularly fan-shaped; surface orangish brown to vinaceous brown to creamy violet; glabrous or covered with minute patches toward the crenate or undulate margin. Hymenium surface gray-violet to violet or vinaceous brown; wrinkled; irregularly reticulate to almost poroid and extending to the upper stipe. Stipe is off-white to pale violet; glabrous at apical part but tomentose to hispid toward the white base; context off-white.
Basidiospores (9–) 10–15 (–17) × (4–) 5–7.5 μm; oblong in side view; ellipsoid in ventral view; non-amyloid; surface covered with verrucose ornamentation. Basidia 60–95 × 8.5–12 μm, clavate, (2–) 4-spored. Pileipellis is composed of uninflated hyphae with pileocystidia 3–4.5 (–5.5) μm, protruding 50–120 μm from the surface. Pileus and stipe context composed of profusely interwoven and uninflated hyphae 2.5–6 µm wide. Clamp connections present in all parts of basidioma; up to 12 µm wide.
Habitat and locality—Gregarious in forests of Abies, Picea, Pinus densata and Quercus semecarpifolia. It is common in southwestern China at 2700–3600 m altitude and widely distributed in Europe and North America.
Specimens examined—AUSTRIA. Klein Walsertal: in forests with Abies and Picea, alt. 1100 m, 27 September 2016, Zhu-Liang Yang 5933 (HKAS 96161). CHINA. GANSU: Gannan Prefecture, Zhouqu County, Shatan Forest Park, in forests with Abies, 16 August 2012, Xue-Tai Zhu 725 (HKAS 76574). SICHUAN: Garzê Prefecture, Daofu County, Geka Township, alt. 3544 m, 17 July 2014, Jian-Wei 82 (HKAS 90876). Garzê Prefecture, Daofu County, Kongse Township, 439 km milestone of Provincial Highway 303, in forests with Picea, alt. 3500 m, 18 August 2013, Bang Feng 1441 (HKAS 82547). TIBET: Bomi County, alt. 2760 m, 17 July 2019, Zhu-Liang Yang 6187 (HKAS 106799). Changdu County, Gongxi Village, 7 August 2013, Kuan Zhao 324 (HKAS 80749). Gongbujiangda County, Bahe Town, in forests with Picea, Pinus densata and Quercus semecarpifolia, alt. 3458 m, 29 July 2014, Bang Feng 1646 (HKAS 94030). Linzhi County, Bayi District, Tunbudanggang Village, 29°39′ N, 94°44′ E, 23 July 2019, Geng-Shen Wang 532 (HKAS 116246). Linzhi County, Bujiu Township, Lamaling, 31 July 2014, Jian-Wei Liu 164 (HKAS 90958). YUNNAN: Diqing Prefecture, Xianggelila City, Haba Snow Mountain, Mianshaba, alt. 3100 m, 13 August 2003, Li-Ping Tang 621 (HKAS 54852). Diqing Prefecture, Xianggelila City, Xiaozhongdian Town, Xiaotianchi Scenic Spot, in forests of Abies, 9 May 2018, Jian-Wei Liu 1623 (HKAS 122603). Lijiang City, Yulong County, Alpine Garden, 22 September 2019, Jian-Wei Liu 2085 (HKAS 122961).
Notes—This species was reported from southwestern China under either the name of
G. orientalis or
G. clavatus [
57,
58]. Our molecular and morphological study of 13 collections of this group confirmed the occurrence of
G. clavatus in southwestern China, which was often confused with
G. orientalis (see below) due to their similar characteristics of merismatoid basidiomata, irregularly fan-shaped pileus and purplish hymenium. However,
G. clavatus generally possesses an off-white context, smaller basidiospores (10–15 × 5–7.5 μm) and the common presence of pileocystidia in pileipellis, while
G. orientalis has a grayish violet context, larger basidiospores (15–18 × 7–9 μm) and the rare presence of pileocystidia in pileipellis. Our phylogenetic analysis resolved
G. orientalis as a lineage independent of
G. clavatus (
Figure 1).
Gomphus matijun J.W. Liu & F.Q. Yu, Mycoscience 63: 293–297 (2022). (
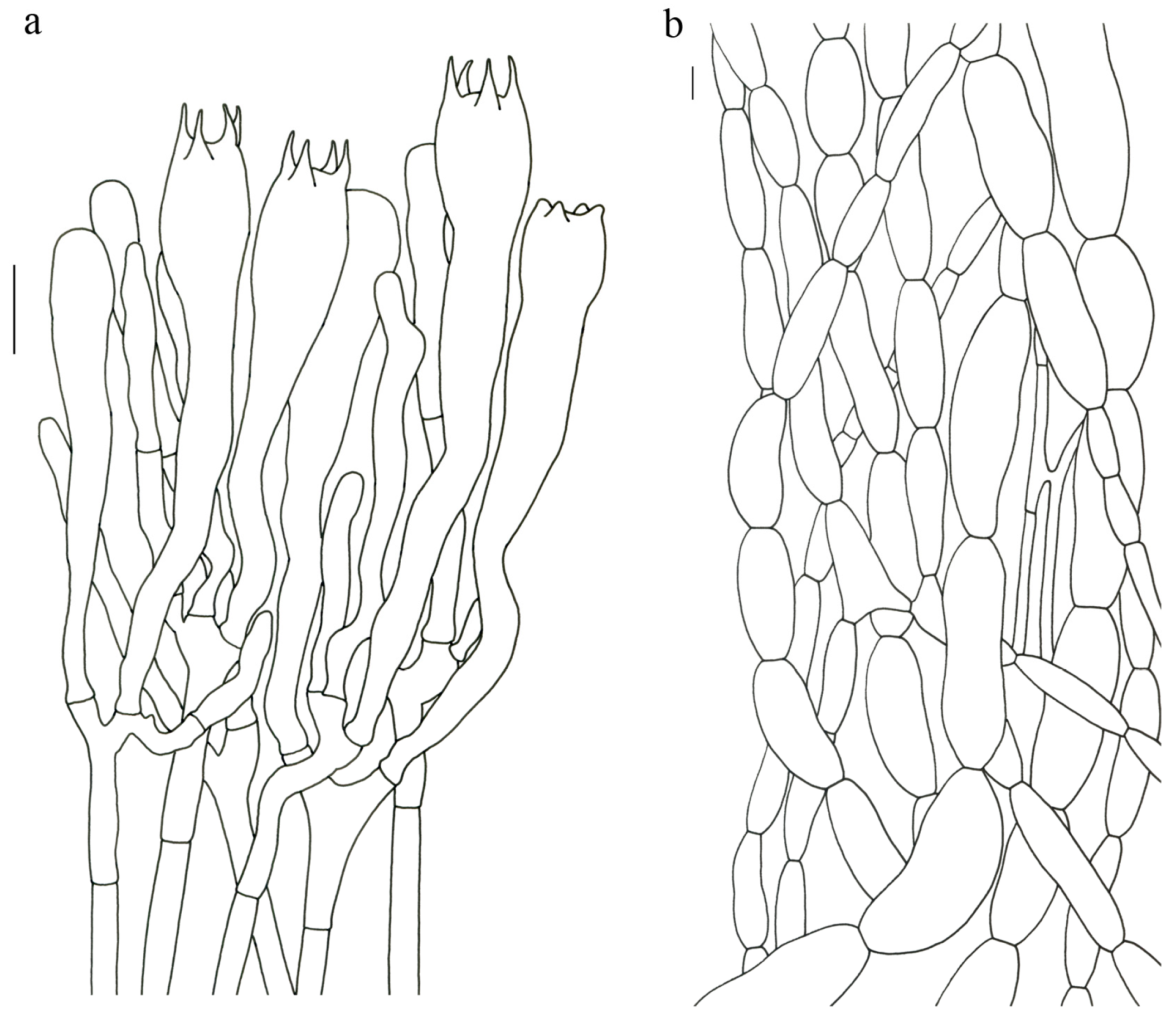
Figure 4).
Description—Basidioma up to 14 cm tall; unipileate to merismatoid; clavate or urceiform when young and hippocrepiform with age. Pileus up to 9 cm in diam.; gradually inflated upwards and depressed at center when mature; grayish purple; pileal margin undulate; rarely lobed. Hymenium surface grayish blue to bluish purple when young but fading to vinaceous gray or pale lilac when mature; deeply wrinkled; irregularly reticulate and extending to the stipe base. Stipe about 2–7 cm long; 1–3.5 cm in diam.; subclavate; upper part vinaceous brown or grayish purple, becoming whitish toward the base; context white to gray or slightly grayish blue.
Basidiospores [40/2/2] 9–11 (–12) × (5–) 6–7 (–8) μm; Q = (1.38–) 1.40–1.8 (–2.0); Qm = 1.65 ± 0.16, ellipsoid to oblong; non-amyloid; ornamentation verrucous. Basidia 60–90 × 7–11 μm, 1–3 spored and 2 spored common; hymenial cystidia absent. Pileipellis composed of uninflated hyphae 4–6 μm wide. Clamp connections present in all parts of basidioma.
Habitat and locality—Solitary to scattered in subalpine fagaceous forests at 1100–1200 m altitude during summer and autumn. Currently known from southwestern China.
Specimens examined—CHINA. GUIZHOU: Southwest Guizhou Autonomous Prefecture, Xingyi City, bought from Xiangjiaba Market, 24°59′ N, 104°56′ E, alt. 1163 m, 19 June 2020, Jian-Wei Liu 2287 (HKAS 122604, holotypus). Southwest Guizhou Autonomous Prefecture, Xingyi City, Zerong Town, bought from Xiangjiaba Market, alt. 1170 m, 30 August 2021, Jian-Wei Liu 2407 (HKAS 122605).
Notes—This species is close to
G. ludovicianus R.H. Petersen, J. Justice and D.P. Lewis,
G. crassipes and
G. orientalis (
Figure 1). However, it can be distinguished from the latter three by its grayish blue to blue or bluish purple and rarely lobed pileus, smaller basidiospores (9–11 × 6–7 μm) and basidia with 1–3 sterigmata. Furthermore, they occur in different habitats:
Gomphus matijun occurs in fagaceous forests in southwestern China (Guizhou Province);
G. ludovicianus produces basidiomata in forests of
Carya,
Pinus palustris and
Quercus in central Louisiana and southeastern Texas;
G. crassipes in coniferous forests in Algeria, Morocco and Spain; and
G. orientalis in forests with
Abies,
Picea,
Pinus and
Betula in southwestern China (Tibet and Yunnan).
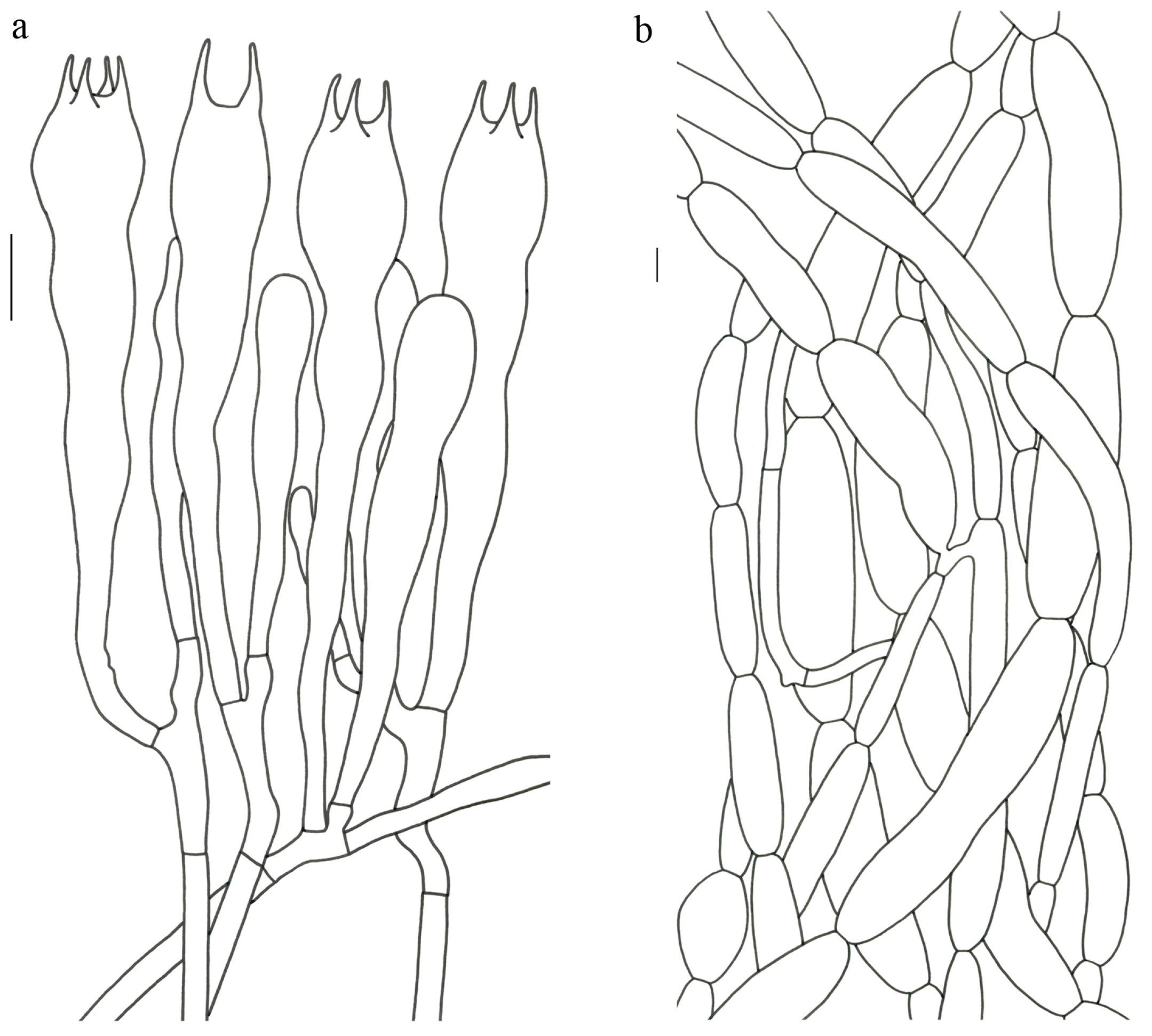
Gomphus orientalis R.H. Petersen et M. Zang, in Zang, Li and Xi, Fungi of the Hengduan Mountains 181 (1996). (
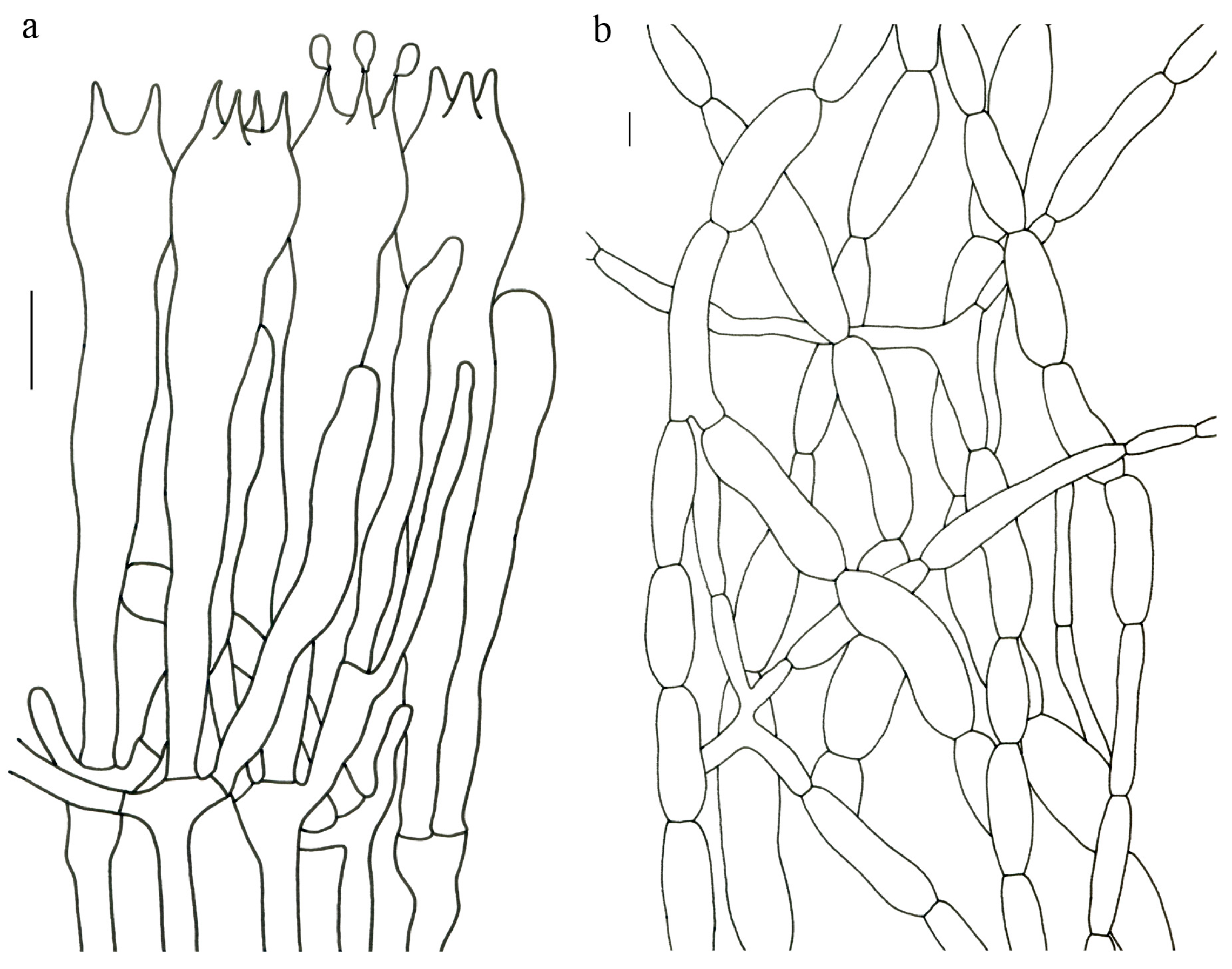
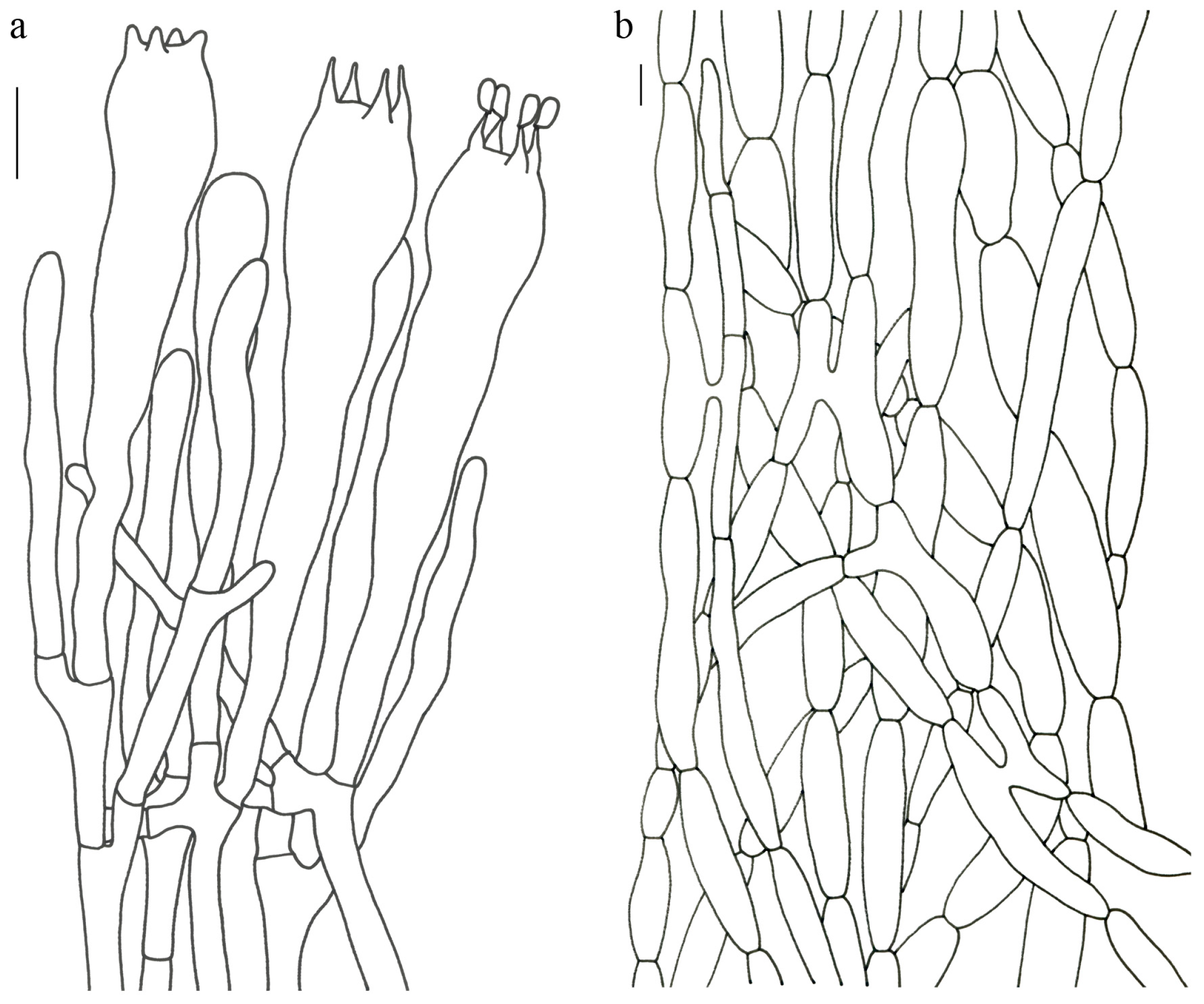
Figure 4,
Figure 5 and
Figure 6).
Description—Basidioma 7–13 cm tall; unipileate or merismatoid; tapering downward. Pileus 4–13 cm in diam.; lightly funnel-shaped or irregularly fan-shaped; depressed at center; surface yellowish brown (5D3) to purplish brown (8D3); pubescent; pileal margin uplifted and undulate. Hymenium surface grayish violet (14D3); deeply wrinkled; irregularly reticulate and extending longitudinally to the stipe base. Stipe about 3–5 cm long; 2–4 cm in diam.; tapering downward; solid; grayish violet (14D3); context grayish violet (14D3); unchanging on exposure; basal mycelium white (1A1).
Basidiospores [60/3/3] 15–18 (–20) × (6–) 7–9 µm; Q = (1.67–) 1.94–2.43 (–2.67); Qm = 2.18 ± 0.18; elliptic-fusiform to cylindrical and inequilateral in side view with distinct suprahilar depression; oblong in ventral view; non-amyloid; light yellowish in KOH; ornamentation strongly verrucous arranged in longitudinal ridges. Basidia 55–75 × 11–14 µm; clavate; 4-spored; sterigmata 4–7 µm long. Hymenial cystidia absent. Pileipellis composed of uninflated hyphae; usually 3–5 um wide; irregularly erect at surface; more or less parallel arranged toward pileal context; gloeoplerous hyphae abundant; yellow in water. Stipitipellis composed of some basidia-like cells and narrow hyphae (1.5–4 μm in diam.). Pileus and stipe context composed of simple-branched, narrow hyphae; usually 2–5 um wide; hyaline to yellow in KOH. Hymenial trama composed of subparallel to interwoven, undifferentiated and narrow hyphae. Clamp connections present in all parts of basidioma.
Habitat and distribution—Scattered in forests of Abies, Picea, Pinus and Betula in southwestern China (Tibet and Yunnan); fruiting from June to September between 2450 and 3400 m altitude.
Specimens examined—CHINA. YUNNAN: Lijiang City, Yulong County, Yulong Snow Mountain, in forests of Abies and Picea, alt. 3000–3400 m, 6 September 1986, R.H. Petersen 56,817 (HKAS 18144, isotype). Nujiang Prefecture, Lanping County, Tongdian Town, Luoguqing Scenic Spot, in forests with Pinus and Betula, alt. 2490 m, 16 August 2010, Bang Feng 865 (HKAS 68646). TIBET: Bomi County, near National Highway 318, in forests with Pinus densata, Pinus armandii and Betula platyphylla, 29°48′ N, 95°47′ E, alt. 2762 m, 30 June 2014, Qing Cai 1107 (HKAS 83567).
Notes—Gomphus orientalis is distinguished by the combination characters of the yellowish brown and lightly funnel-shaped or irregularly fan-shaped pubescent pileus, and the violet hymenium, stipe and context. Gomphus orientalis is similar and closely related to G. matijun. However, the former possesses a violet context, 4-spored basidia and larger basidiospores (15–18 × 7–9 µm), while the latter has a white to gray or slightly grayish blue context, generally 2-spored basidia and smaller basidiospores (9–11 × 6–7 μm). In addition, G. orientalis and G. matijun occur in different habitats: Gomphus orientalis is distributed in forests of Abies, Picea, Pinus and Betula at 2450–3400 m altitude, while G. matijun is found in fagaceous forests at 1100–1200 m altitude.
Zang et al. [
59] described basidiospores of
G. orientalis as 10.3–15.5 (–16.8) × 4.3–7.5 µm, significantly smaller than those of the isotype measured by us (15–17 × 7–9 µm), which were consistent with other two collections and were adopted here.
Gomphus orientalis has a relative wide range of habitats. The isotype was collected in forests of
Abies and
Picea at 3000–3400 m altitude and the other two collections were collected in forests of
Pinus and
Betula at 2450–2800 m altitude. Nevertheless, these collections were treated here as the same species because molecular evidence indicated that they represented the same species (
Figure 1).
MycoBank: MB 843958
Etymology—From the Latin adjective, violaceus, referring to the violet colored basidioma and context of the new species.
Holotypus—CHINA. YUNNAN: Lufeng City, Heping Town, beside Daluqing Reservoir, in forests with Pinus, alt. 2000 m, 18 August 2018, Zhu-Liang Yang 6154 (HKAS 104313; Genbank OQ858428/OQ858495/OQ847591).
Diagnosis—Gomphus violaceus is similar and closely related to G. clavatus. However, the former possesses a merismatoid basidioma with 2–5 subpilei, a bluish violet pileus, a violet context and the rare presence of pileocystidia in pileipellis; the latter has a merismatoid basidioma with 5–15 subpilei, a brown or grayish purple pileus, an off-white context and the common presence of pileocystidia in pileipellis. G. violaceus has an allopatric distribution with G. clavatus, in which G. violaceus occurs in forests with Pinus in 2000–2150 m altitude, while G. clavatus is found in forests with Abies and Picea at 2700–3600 m altitude.
Description—Basidioma 6–13 cm tall; unipileate at base and then merismatoid with 2–5 subpilei; and two subpilei common. Pileus 2–8 cm in diam.; irregularly fan- or funnel-shaped; depressed to one side; surface bluish violet (18A5) to violet (16C4); thick in the center; thin toward the ascending; pileal margin uplifted and undulate. Hymenium surface bluish violet (18A4–B4); deeply wrinkled; irregularly reticulate to almost poroid and extending to the stipe base. Stipe about 3–5 cm long; 1–3 cm in diam.; tapering downward; dilating obconical into the pileus; solid; bluish violet (18A5) to violet (16C4) or dark violet (14D4); context bright violet (18B4); unchanging on exposure; basal mycelium white (1A1).
Basidiospores [57/3/3] (11.5–) 12–14.5 (–15) × 6–8.5 (–9) µm, Q = (1.60–) 1.64–2.17 (–2.33), Qm = 1.94 ± 0.18, oblong and inequilateral in side view with distinct suprahilar depression; ellipsoid in ventral view; non-amyloid; light yellowish in KOH; ornamentation strongly verrucose with fine warts. Basidia 50–70 × 8–11 µm; clavate; generally 4-spored; sterigmata 5–9 µm long. Hymenial cystidia absent. Pileipellis composed of uninflated hyphae with simple branches that are usually 2–4 um wide and 2–7 um wide downward; irregularly erect at surface; more or less parallel arranged toward the pileal context; gloeoplerous hyphae abundant; yellow in KOH. Stipitipellis composed of some basidia-like cells and narrow hyphae (2–5 μm in diam.). Pileus and stipe context composed of profusely parallel-arranged, narrow and uninflated hyphae; 2–5 µm wide; hyaline to yellow in KOH. Hymenial trama composed of subparallel to interwoven, undifferentiated and narrow hyphae; 2–4 µm wide. Clamp connections present in all parts of basidioma.
Habitat and distribution—Solitary to scattered in subtropical forests dominated by plants of Pinus; endemic to Yunnan Province, China; fruiting from June to August in 2000–2150 m altitude.
Additional specimens examined—CHINA. YUNNAN: Kunming City, Qiongzhu Temple, alt. 2047 m, 8 August 2007, Li-Ping Tang 224 (HKAS 53267). Lufeng City, Heping Town, beside Daluqing Reservoir, alt. 2129 m, 18 August 2018, Jian-Wei Liu 1156 (HKAS 122947).
Notes—Gomphus violaceus is easily distinguished by a simply merismatoid basidioma; a bluish violet and irregularly funnel-shaped pileus; a violet context; and small basidiospores (12–14.5 × 6–8.5 µm), some of which are shared with G. orientalis. However, they have different habitats with G. violaceus in forests with Pinus in 2000–2150 m altitude and G. orientalis in forests with Abies, Picea, Pinus and Betula in 2450–3400 m altitude.
MycoBank: MB 843946
Etymology—From the Latin adjective, flavidus, referring to the yellowish basidioma of the new species.
Holotypus—CHINA. HUNAN: Zhou City, Xuanzhang County, Mangshan Forest Park, near Zeziping, in forests of Castanopsis, Pinus kwangtungensis and Schima superba, alt. 1340–1460 m, 14 September 2016, Gang Wu 1826 (HKAS 99946; Genbank OQ858428/OQ858495/OQ847591).
Diagnosis—Turbinellus flavidus is very similar to T. longistipes in having sparsely scaly pileal surface, yellow hymenium, yellow cylindrical stipe and large basidiospores. However, T. flavidus possesses a smaller basidioma (less than 5 cm tall) with a yellow pileus, a short stipe and a yellow context, while T. longistipes has a larger basidioma (up to 15 cm tall) with an orange pileus, a long stipe and a white context.
Description—Basidioma 3.5–5 cm tall; unipileate; tapering downward. Pileus 0.9–2.3 cm in diam.; funnel-shaped; depressed at center; surface light yellow (3A4) to orange-yellow (4A4–6); sparsely covered with minute appressed scales; scales more or less radially arranged; pileal margin uplifted and undulate. Hymenium surface creamy (3A2–3); wrinkled; irregularly reticulate and extending to the upper stipe; sparse; reaching to margin upwards. Stipe about 1.5–1.7 cm long; 0.5–0.8 cm in diam.; approximately cylindrical; solid; orange-yellow (4A4); glabrous above the base; context pliable; thin to 0.1–0.2 cm; creamy (3A2); unchanging on exposure; basal mycelium white (1A1).
Basidiospores [22/2/1] (15.5–) 16–20 (–21) × 8–10 µm; Q = (1.82–) 1.84–2.12 (–2.21); Qm = 1.99 ± 0.1; elliptic-fusiform and inequilateral in side view with distinct suprahilar depression; ellipsoid in ventral view; non-amyloid; light yellowish in KOH; ornamentation strongly verrucose with coarse warts. Basidia 55–100 × 10–15 µm; clavate; 2–4-spored; sterigmata 5–10 µm long. Hymenial cystidia absent. Pileipellis composed of numerously inflated hyphae (7–30 μm in diam.), mostly with ampulliform swellings and little uninflated hyphae (3–6 μm in diam.); simple branched; constricted at septa; repent; more or less parallel arranged toward the pileal context; gloeoplerous hyphae abundant; yellow in water. Stipitipellis composed of some basidia-like cells; plentifully uninflated hyphae (2–6 μm in diam.) and little inflated hyphae (7–13 μm in diam.); gloeoplerous hyphae abundant; yellow in water. Pileus and stipe context composed of thin- to thick-walled hyphae; approximately parallel; hyaline to yellow in KOH. Hymenial trama composed of subparallel to interwoven, undifferentiated and narrow hyphae. Clamp connections absent in all parts of basidioma.
Habitat and distribution—In forests with Castanopsis, Pinus kwangtungensis and Schima superba in central China (Hunan Province) between 1300 m and 1500 m altitude.
Notes—This species almost has the smallest basidioma in the genus and is closely related to
T. tomentosipes and
T. solidus in our phylogenetic analysis (
Figure 1). However,
T. flavidus differs from the latter two due its smaller yellow basidioma with cream-colored context and large basidiospores. Moreover,
T. flavidus has only been collected in forests with
Castanopsis,
Pinus kwangtungensis and
Schima superba in central China, while the latter two species occur in forests with Fagaceae and
Pinus yunnanensis in southwestern China (see below).
MycoBank: MB 843949
Etymology—From the Latin adjective, fulvus, referring to the fulvous pileus and stipe of the new species.
Holotypus—CHINA. YUNNAN: Chuxiong Prefecture, Nanhua County, 15 km of Provincial Highway 217, in the mixed coniferous broad-leaved forest, alt. 2200 m, 3 August 2009, Gang Wu 93 (HKAS 57625; Genbank OQ858452/OQ858520/OQ847610).
Diagnosis—Turbinellus fulvus is very similar to T. solidus and T. tomentosipes in that it has a sparsely scaly pileal surface, sparse hymenium, and solid stipe. However, T. fulvus possesses a smaller basidioma (less than 9 cm tall), a khaki pileus, a short and pubescent stipe and smaller basidiospores (11.5–14 × 6–7 µm), while T. tomentosipes has a larger basidioma up to 15 cm tall, an orange-yellow pileus and a tomentose cylindrical stipe. T. solidus has a glabrous stipe and larger basidiospores (13.5–16 × 6.5–9 µm).
Description—Basidioma 5–9 cm tall; unipileate; tapering downward. Pileus 1.5–3.5 cm in diam.; funnel-shaped; depressed at center; surface khaki (4B3–4); sparsely covered with minute appressed scales; scales usually darker (5B5) than the pileal surface and more or less radially arranged; pileal margin extensional and undulate. Hymenium surface off-white (1A1); weakly wrinkled; longitudinal ridges extending to the stipe base and branching simply upwards toward the margin; sparse. Stipe about 3.5–4 cm long; 0.6–1.2 cm in diam.; tapering downward; dilating into the pileus; solid; khaki (5B3); covered with short pubescence; context pliable; creamy (1A2); unchanging on exposure; basal mycelium white (1A1).
Basidiospores [26/2/1] 11.5–14 × (5.5–) 6–7 (–7.5) µm; Q = 1.71–2.17 (–2.18); Qm = 1.96 ± 0.15; oblong and inequilateral in side view with distinct suprahilar depression; ellipsoid in ventral view; non-amyloid; light yellowish in KOH; ornamentation strongly verrucose arranged in longitudinal ridges. Basidia 65–85 × 10–13 µm; clavate; 4-spored; sterigmata 4–7 µm long. Hymenial cystidia absent. Pileipellis composed of profusely inflated hyphae (7–27 μm in diam.) mostly with ampulliform swellings and little uninflated hyphae (3–6 μm in diam.); simple branched; constricted at septa; repent; more or less parallel arranged toward the pileal context; gloeoplerous hyphae abundant; hyaline to yellow in water. Stipitipellis composed of some basidia-like cells; plentifully uninflated hyphae (2–6 μm in diam.) and some inflated hyphae (7–12 μm in diam.); gloeoplerous hyphae abundant; yellow in KOH. Pileus and stipe context composed of simple branched hyphae; approximately parallel; hyaline to yellow in KOH. Hymenial trama composed of subparallel to interwoven, undifferentiated and narrow hyphae. Clamp connections absent in all parts of basidioma.
Habitat and distribution—In the mixed coniferous broad-leaved forests; fruiting from June to August in southwestern China (Yunnan Province) at about 2200 m altitude.
MycoBank: MB 843959
Etymology—From the Latin adjective, imbricatus, referring to the pileal surface of the new species densely covered with large scales.
Holotypus—CHINA. YUNNAN: Wenshan Prefecture, Guangnan County, Nanping Town, Shuigou Mountain, in forests with fagaceous trees, 23°45′ N, 105°12′ E, alt. 1323 m, 1 August 2014, Jing Li 81 (HKAS 85905; Genbank OQ858443/OQ858509/OQ847603).
Diagnosis—
Turbinellus imbricatus is similar to
T. kauffmanii in having large scales on the pileal surface, but the former has a smaller basidioma less than 11 cm, a narrower pileus of less than 7 cm, a short and thin stipe (1.8–4.1 × 0.8–1.5 cm), a 4- or 5-spored basidia and the common presence of ampulliform hyphae in pileipellis. The latter has a larger basidioma up to 40 cm, a broader pileus up to 35 cm, a long and stout stipe up to 15 × 6 cm, a 2- or 4-spored basidia and the rare presence of ampulliform hyphae in pileipellis [
1]. Moreover,
T. imbricatus occurs in fagaceous forests only known to southwestern China, while
T. kauffmanii grows in coniferous forests only known to North America.
Description—Basidioma 6–11 cm tall; unipileate; tapering downward. Pileus 1–7 cm in diam.; subcylindrical when young and narrow funnel-shaped with age; depressed at center; surface light brown (5B4); radial bulge of the surface densely covered with large scales; scales with radial stripes and rolling up in waves; pileal margin extending over hymenium and undulate. Hymenium surface off-white (1A1); slightly wrinkled; irregularly reticulate and extending to the upper stipe; sparse; thin to 0.2–0.4 cm. Stipe about 1.8–4.1 cm long; 0.8–1.5 cm in diam.; tapering downward; dilating obconical into the pileus; solid; white (1A1); glabrous above the base; context pliable; thin to 0.1–0.2 cm; white (1A1); unchanging on exposure; basal mycelium white (1A1).
Basidiospores [40/2/2] 11.5–14 × 6–8 µm; Q = 1.71–2.15 (–2.17); Qm = 1.91 ± 0.13; oblong and inequilateral in side view with distinct suprahilar depression; ellipsoid in ventral view; non-amyloid; light yellowish in KOH; ornamentation slightly verrucose with fine warts. Basidia 60–85 × 9–14 µm; clavate; generally 4- or 5-spored; sterigmata 4.5–9 µm long. Hymenial cystidia absent. Pileipellis composed of profusely inflated hyphae (7–20 μm in diam.), mostly with ampulliform swellings and barely uninflated hyphae (2–6 μm in diam.); simple branched; constricted at septa; repent; more or less parallel arranged toward the pileal context; gloeoplerous hyphae abundant; yellow in KOH. Stipitipellis composed of some basidia-like cells and profusely uninflated hyphae (2–8 μm in diam.); gloeoplerous hyphae abundant; yellow in water. Pileus and stipe context composed of thin- to thick-walled hyphae; approximately parallel; hyaline to yellow in KOH. Hymenial trama composed of subparallel to interwoven, undifferentiated, thin-walled and narrow hyphae. Clamp connections absent in all parts of basidioma.
Habitat and distribution—In mingled forests dominated by fagaceous plants; fruiting from June to August in southwestern China (Yunnan Province) in 1300–1400 m altitude.
Additional specimen examined—CHINA. YUNNAN: Wenshan Prefecture, Guangnan County, Nanping Town, Shuigou Mountain, in mingled forest dominated by fagaceous plants, 23°40′ N, 105°12′ E, alt. 1398 m, 1 August 2014, Pan-Meng Wang 18 (HKAS 95099).
MycoBank: MB 843960
Etymology—From the Latin longistipes, referring to the long stipe of the new species.
Holotypus—CHINA. YUNNAN: Lijiang City, Yulong County, 99 Longtan Scenic Spot, alt. 3600 m, 27 August 2020, Fei-Fei Liu 316 (HKAS 113226; Genbank OQ858464/OQ858533/OQ847621).
Diagnosis—Turbinellus longistipes is very similar to T. yunnanensis in having orange pileus with minute appressed scales. However, the former possesses a creamy to light orange basidioma, a long stipe (5–10 cm) with the slightly swollen base and larger basidiospores (15–20 × 7–9.5 µm). The latter has a reddish basidioma, a cylindrical and short stipe (2–4 cm) and smaller basidiospores (13–17 × 6.5–7.5 µm). Ecologically, T. longistipes occurs at 3200–3600 m altitude, while T. yunnanensis is distributed at 2800–3200 m altitude (see below).
Description—Basidioma 4–15 cm tall; unipileate; tapering downward. Pileus 2–10 cm in diam.; funnel-shaped; depressed at center; surface orange (5A7–6A7); sparsely covered with minute appressed scales; pileal margin undulate or entire. Hymenium surface creamy (2A2) to creamy orange (5A2); wrinkled; irregularly reticulate and extending to the upper stipe; sparse; thin to 0.2 cm. Stipe 5–10 cm long; 0.5–1 cm in diam.; subcylindrical; base slightly swollen; nearly solid; creamy (1A2–3) to creamy orange (5A2); glabrous; context pliable; thick to 0.2–0.4 cm; white (1A1); unchanging on exposure; basal mycelium white (1A1).
Basidiospores [82/4/4] 15–20 (–21) × 7–9.5 (–10) µm; Q = (1.76–) 1.82–2.33 (–2.5); Qm = 2.06 ± 0.16; oblong to cylindrical in side view with distinct suprahilar depression; ellipsoid in ventral view; non-amyloid; light yellowish in KOH; ornamentation strongly verrucose arranged in longitudinal ridges. Basidia 65–120 × 11–20 µm; clavate, 2–4-spored; sterigmata 5–10 µm long. Hymenial cystidia absent. Pileipellis composed of profusely inflated hyphae (7–24 μm in diam.), mostly with ampulliform swellings and little uninflated hyphae (3–6 μm in diam.); simple branched; constricted at septa; repent; more or less parallel arranged toward the pileal context; gloeoplerous hyphae abundant; hyaline to yellow in KOH. Stipitipellis composed of some basidia-like cells, inflated hyphae (7–18 μm in diam.) and uninflated hyphae (2–6 μm in diam.); gloeoplerous hyphae abundant; yellow in KOH. Pileus and stipe context composed of thin- to thick-walled hyphae; approximately parallel; hyaline to yellow in KOH. Hymenial trama composed of subparallel to interwoven, undifferentiated and narrow hyphae. Clamp connections absent in all parts of basidioma.
Habitat and distribution—In subtropical forests dominated by plants of Abies, Picea, Pinus densata and Quercus semicarpifolia; fruiting from June to September in southwestern China (Yunnan Province) between 3200 and 3600 m altitude.
Additional specimens examined—CHINA. YUNNAN: Diqing Prefecture, Xianggelila City, Xiaozhongdian Town, in forests with Picea and Abies, alt. 3300 m, 21 August 2013, Qi Zhao 2190 (HKAS 87956). Lijiang City, Yulong County, Yuhu Reservoir, in forests with Pinus densata and Quercus semicarpifolia, alt. 3200 m, 30 August 2013, Bang Feng 1463 (HKAS 82569). Nujiang Prefecture, Lanping County, Tongdian Town, 1 September 2018, Jian-Wei Liu 1472 (HKAS 122957).
Notes—Turbinellus longistipes is easily distinguished by its creamy to light orange and large basidioma, long stipe (5–10 cm) with slightly swollen base and large basidiospores. This species may also be confused with T. verrucosus in the wild. However, T. longistipes differs from T. verrucosus by its minute appressed scales sparsely covering the pileal surface and long stipe with slightly swollen base (see below).
MycoBank: MB 843961
Etymology—From the Latin prefix, parvi-, and the Latin suffix, -sporus, referring to the small basidiospores of the new species.
Holotypus—CHINA. YUNNAN: Kunming City, Panlong District, Wild Duck Lake Park, alt. 2000–2200 m, 18 August 2012, Yan-Chun Li 2832 (HKAS 89475; Genbank OQ858450/OQ858516/OQ847607).
Diagnosis—Turbinellus parvisporus is similar to T. fujisanensis in having large deltoid scales on pileal surface and small basidiospores, but the former has a smaller basidioma less than 9 cm tall, with the reticulation of hymenium becoming stronger upwards and the common presence of inflated hyphae (7–24 μm in diam.) in pileipellis and stipitipellis. The latter has a larger basidioma up to 15 cm tall, an uniformly forked hymenium and the rare presence of inflated hyphae in all parts of the basidioma.
Description—Basidioma 4–9 cm tall; unipileate; tapering downward. Pileus 3–7 cm in diam.; funnel-shaped; depressed at center; surface flesh-colored (6A4–5); radial bulge of the surface densely covered with large deltoid scales especially in the center; scales rolling up in waves; pileal margin uplifted and undulate. Hymenium surface white (1A1) to creamy (3A2); strongly wrinkled; irregularly reticulate and extending to the upper stipe. Stipe about 1.5–3.5 cm long; 0.6–1.2 cm in diam.; tapering downward; dilating obconical into the pileus; hollow; khakis (4B3) to flesh-colored (6A5); glabrous above the base; context pliable; thin to 0.1–0.2 cm; white (1A1); unchanging on exposure; basal mycelium white (1A1).
Basidiospores [41/3/2] 10–13 × 5.5–7 µm, Q = 1.43–2.09 (–2.18); Qm = 1.78 ± 0.2, ellipsoid to oblong and inequilateral in side view with distinct suprahilar depression; ellipsoid in ventral view; non-amyloid; light yellowish in KOH; ornamentation moderately verrucose with coarse warts. Basidia 50–80 × 9–13 µm; clavate; generally 4-spored; sterigmata 4–8 µm long. Hymenial cystidia absent. Pileipellis composed of profusely inflated hyphae (7–24 μm in diam.), mostly with ampulliform swellings and little uninflated hyphae (3–6 μm in diam.); simple branched; constricted at septa; repent; more or less parallel arranged toward the pileal context; gloeoplerous hyphae abundant; hyaline to yellow in KOH. Stipitipellis composed of some basidia-like cells; plenty of inflated hyphae (7–15 μm in diam.) and uninflated hyphae (2–6 μm in diam.); gloeoplerous hyphae abundant; hyaline to yellow in KOH. Pileus and stipe context composed of thin- to thick-walled hyphae; approximately parallel; hyaline to yellow in KOH. Hymenial trama composed of subparallel to interwoven, undifferentiated and narrow hyphae. Clamp connections absent in all parts of basidioma.
Habitat and distribution—On humid, heavy metal-contaminated soils; fruiting from June to August in southwestern China (Yunnan Province) between 1800 m and 2000 m altitude.
Additional specimens examined—CHINA. YUNNAN: Dali Prefecture, Heqing County, Xiyi Town, Beiya Village, 27 August 2018, Jian-Wei Liu 1241 (HKAS 122952). Dali Prefecture, Heqing County, Xiyi Town, Beiya Village, alt. 1800–1900 m, 18 August 2018, Jian-Wei Liu 1263 (HKAS 122954).
MycoBank: MB 843963
Etymology—From the Latin adjective, solidus, referring to the solid stipe of the new species.
Holotypus—CHINA. YUNNAN: Wenshan Prefecture, Guangnan County, Nanping Town, in forests with Keteleeria fortunei and Pinus yunnanensis, 23°45′ N, 105°12′ E, alt. 1350 m, 7 August 2015, Kuan Zhao 820 (HKAS 92450; Genbank OQ858517/OQ847608).
Diagnosis—Turbinellus solidus is similar to T. yunnanensis due to their small basidiomata with the orange-yellow to orange and scaly pileal surface and medium-sized basidiospores. However, the former possesses a white to creamy basidioma and a solid stipe, while the latter has a reddish basidioma and a hollow stipe. Ecologically, T. solidus is distributed in subtropical forests dominated by Fagaceae, Keteleeria fortunei and Pinus yunnanensis at 1300–1600 m altitude, while T. yunnanensis occurs in forests of Abies, Picea, Pinus densata and Quercus at 2800–3200 m altitude (see below).
Description—Basidioma 6–10 cm tall; unipileate; tapering downward. Pileus 1.8–7.5 cm in diam.; funnel-shaped; depressed at center; cup depth 3–6 cm; surface orange-yellow (4A3–4) to orange (6A7–8); sparsely covered with minute appressed scales; scales more or less radially arranged; pileal margin uplifted and undulate. Hymenium surface off-white (1A1) to creamy (3A2); wrinkled; irregularly reticulate and extending longitudinally to the upper stipe; sparse; thin to 0.2 cm. Stipe about 2–5.5 cm long; 0.5–1.5 cm in diam.; tapering downward; dilating obconically into the pileus; solid; white (1A1) to creamy (1A2); glabrous above the base; context pliable; thin to 0.1–0.2 cm; white (1A1); unchanging on exposure; basal mycelium white (1A1).
Basidiospores [59/3/3] (12–) 13.5–16 (–17.5) × (6–) 6.5–9 µm; Q = (1.67–) 1.75–2.22 (–2.33); Qm = 1.96 ± 0.15; oblong and inequilateral in side view with distinct suprahilar depression; ellipsoid in ventral view; non-amyloid; light yellowish in KOH; ornamentation moderately verrucose with coarse warts. Basidia 50–90 × 9–14 µm; clavate; 2–4-spored; sterigmata 4–9 µm long. Hymenial cystidia absent. Pileipellis composed of profusely inflated hyphae (7–20 μm in diam.), mostly with ampulliform swellings and little uninflated hyphae (3–6 μm in diam.); simple branched; constricted at septa; repent; more or less parallel arranged toward the pileal context; gloeoplerous hyphae abundant; yellow in water. Stipitipellis composed of some basidia-like cells; plenty of inflated hyphae (7–15 μm in diam.) and many uninflated hyphae (2–6 μm in diam.); gloeoplerous hyphae abundant; hyaline to yellow in KOH. Pileus and stipe context composed of moderately thin- to thick-walled hyphae; approximately parallel; hyaline to yellow in KOH. Hymenial trama composed of subparallel to interwoven, undifferentiated, thin-walled and narrow hyphae. Clamp connections absent in all parts of basidioma.
Habitat and distribution—Scattered in subtropical forests with Fagaceae, Keteleeria fortunei and Pinus yunnanensis; on loamy and humid soils; fruiting from June to August in southwestern China (Yunnan Province) between 1300 and 1600 m altitude.
Additional specimens examined—CHINA. YUNNAN: Wenshan Prefecture, Guangnan County, Nanping Town, Shuigou Mountain, in fagaceous forests, 23°45′ N, 105°12′ E, alt. 1323 m, 1 August 2014, Jing Li 78 (HKAS 140801). Wenshan Prefecture, Qiubei County, Jinping Town, Xiangqi Village, in forests with Fagaceae and Pinus yunnanensis, 24°05′ N, 104°14′ E, alt. 1569 m, 10 August 2014, Jing Li 180 (HKAS 86006).
MycoBank: MB 843964
Etymology—From the Latin adjective, squamosus, referring to the pileal surface of the new species covered with large scales.
Holotypus—CHINA. YUNNAN: Dali Prefecture, Binchuan County, Jizushan Town, Shanqiansi Village, in forests with Pinus yunnanensis, alt. 2000–2100 m, 11 August 2011, Qing Cai 654 (HKAS 70249; Genbank OQ858444/OQ858510/OQ847604).
Diagnosis—Turbinellus squamosus is very similar to T. fujisanensis in having large deltoid scales densely covering the pileal surface, but the former has a smaller basidioma less than 9 cm tall; a narrower pileus (1–6 cm in diam.); the reticulation of hymenium becoming stronger upwards; larger basidiospores (11.5–14.5 × 6–7 µm); and the common presence of inflated hyphae (7–25 μm in diam.) in pileipellis. The latter has a larger basidioma up to 15 cm tall, a broader pileus (5–10 cm in diam.), a uniformly forked hymenium, smaller basidiospores (9.5–11.5 × 5–6 µm) and the rare presence of inflated hyphae in all parts of the basidioma. Furthermore, T. squamosus occurs in forests of Pinus yunnanensis in southwestern China, while T. fujisanensis grows in mixed coniferous forests in Japan.
Description—Basidioma 4–9 cm tall; unipileate; tapering downward. Pileus 1–6 cm in diam.; fan- to funnel-shaped; depressed at center; surface creamy (2A3) to flesh-colored (6A3–4); densely covered with large triangular scales; scales rolling up in waves; often loose near the margin with small ones and then filling the center of the basidioma with large ones; darker (6B4–5) than the pileal surface; annularly arranged; pileal margin uplifted and undulate. Hymenium surface white (1A1) when young and creamy (2A3) with age; wrinkled; irregular ridges extending longitudinally to the upper stipe; thick to 0.2–0.5 cm. Stipe about 2–3 cm long; 0.8–1 cm in diam.; tapering downward; dilating obconically into the pileus; hollow; white (1A1) to creamy (2A3); glabrous above the base; context pliable; white (1A1); unchanging on exposure; basal mycelium white (1A1).
Basidiospores [80/4/4] (11–) 11.5–14.5 (–15) × 6–7 (–8) µm; Q = (1.64–) 1.78–2.33 (–2.50); Qm = 2.01 ± 0.17; oblong to cylindrical and inequilateral in side view with distinct suprahilar depression; ellipsoid in ventral view; non-amyloid; light yellowish in KOH; ornamentation strongly verrucose with fine warts. Basidia 50–105 × 7–14 µm; clavate; generally 4-spored; sterigmata 5–9 µm long. Hymenial cystidia absent. Pileipellis composed of profusely inflated hyphae (7–25 μm in diam.), mostly with ampulliform swellings and rarely uninflated hyphae (3–6 μm in diam.); simple branched; constricted at septa; repent; more or less parallel arranged toward the pileal context; gloeoplerous hyphae abundant; hyaline to yellow in KOH. Stipitipellis composed of some basidia-like cells; plenty of uninflated hyphae (2–6 μm in diam.) and little inflated hyphae (7–11 μm in diam.); gloeoplerous hyphae abundant; hyaline to yellow in KOH. Pileus and stipe context composed of thin- to thick-walled hyphae; approximately parallel; hyaline to yellow in KOH. Hymenial trama composed of subparallel to interwoven, undifferentiated and narrow hyphae. Clamp connections absent in all parts of basidioma.
Habitat and distribution—Scattered in subtropical forests mainly with Pinus yunnanensis; moderately common in southwestern China (Yunnan Province); fruiting in June to August between 1800 and 2100 m altitude.
Additional specimens examined—CHINA. YUNNAN: Dali Prefecture, Heqing County, Xiyi Town, Beiya Village, alt. 1800–1900 m, 26 August 2018, Jian-Wei Liu 1207 (HKAS 122950). Dali Prefecture, Heqing County, Xiyi Town, Beiya Village, alt. 1800–1900 m, 28 August 2018, Jian-Wei Liu 1281 (HKAS 122955). Dali Prefecture, Binchuan County, Jizushan Town, Shanqiansi Village, in forests with Pinus yunnanensis, alt. 2000 m, 11 August 2011, Ting Guo 438 (HKAS 71334).
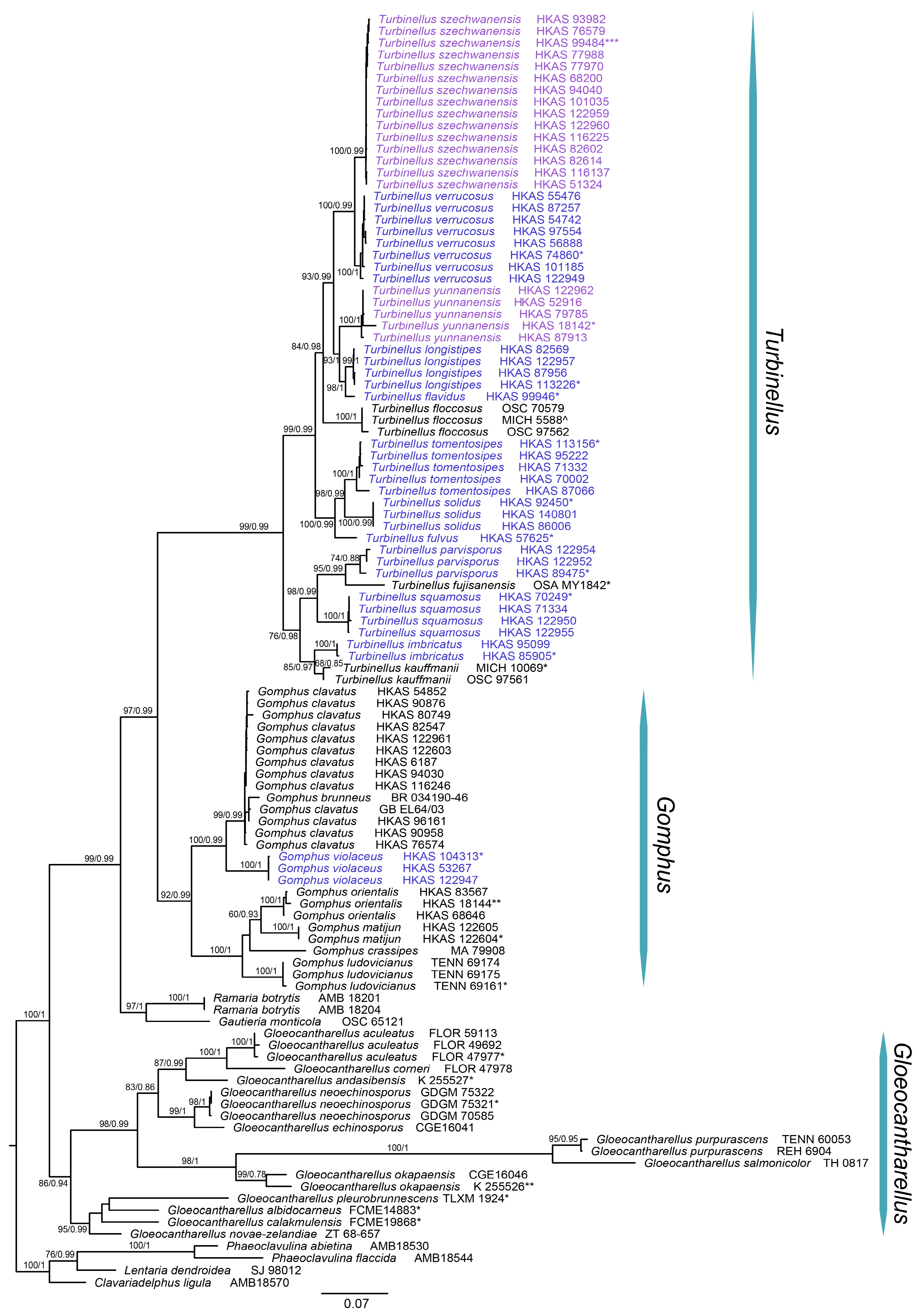
Notes—Our phylogenetic analysis showed that
T. squamosus shares, with high support, a most recent common ancestor with
T. fujisanensis,
T. imbricatus,
T. kauffmanii and
T. parvisporus (
Figure 1). The large deltoid scales on pileal surface are a synapomorphy for these five species and appear to have evolved independently in gomphoid fungi.
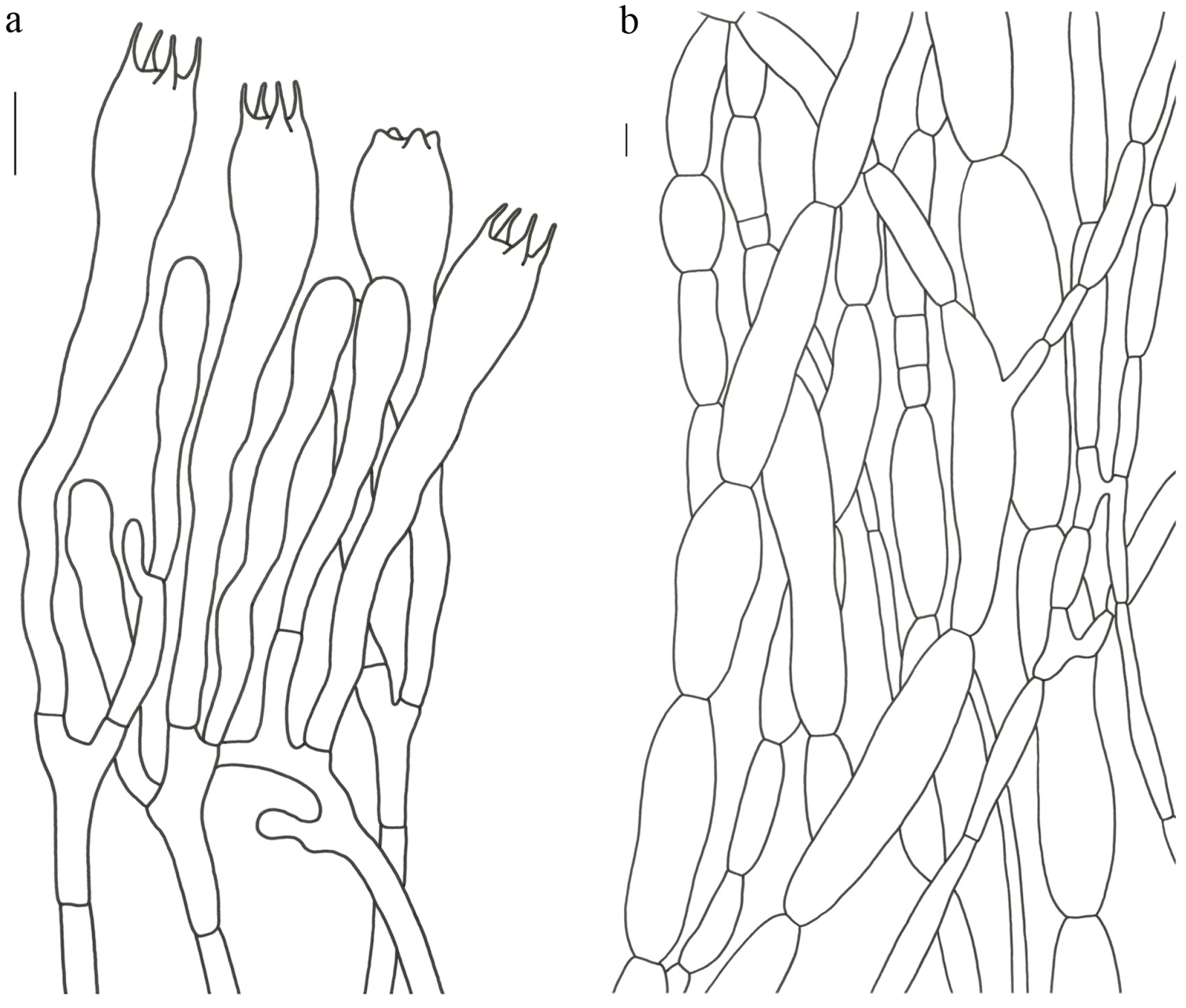
Turbinellus szechwanensis (R.H. Petersen) Xue-Ping Fan & Zhu L. Yang,
comb. nov. (
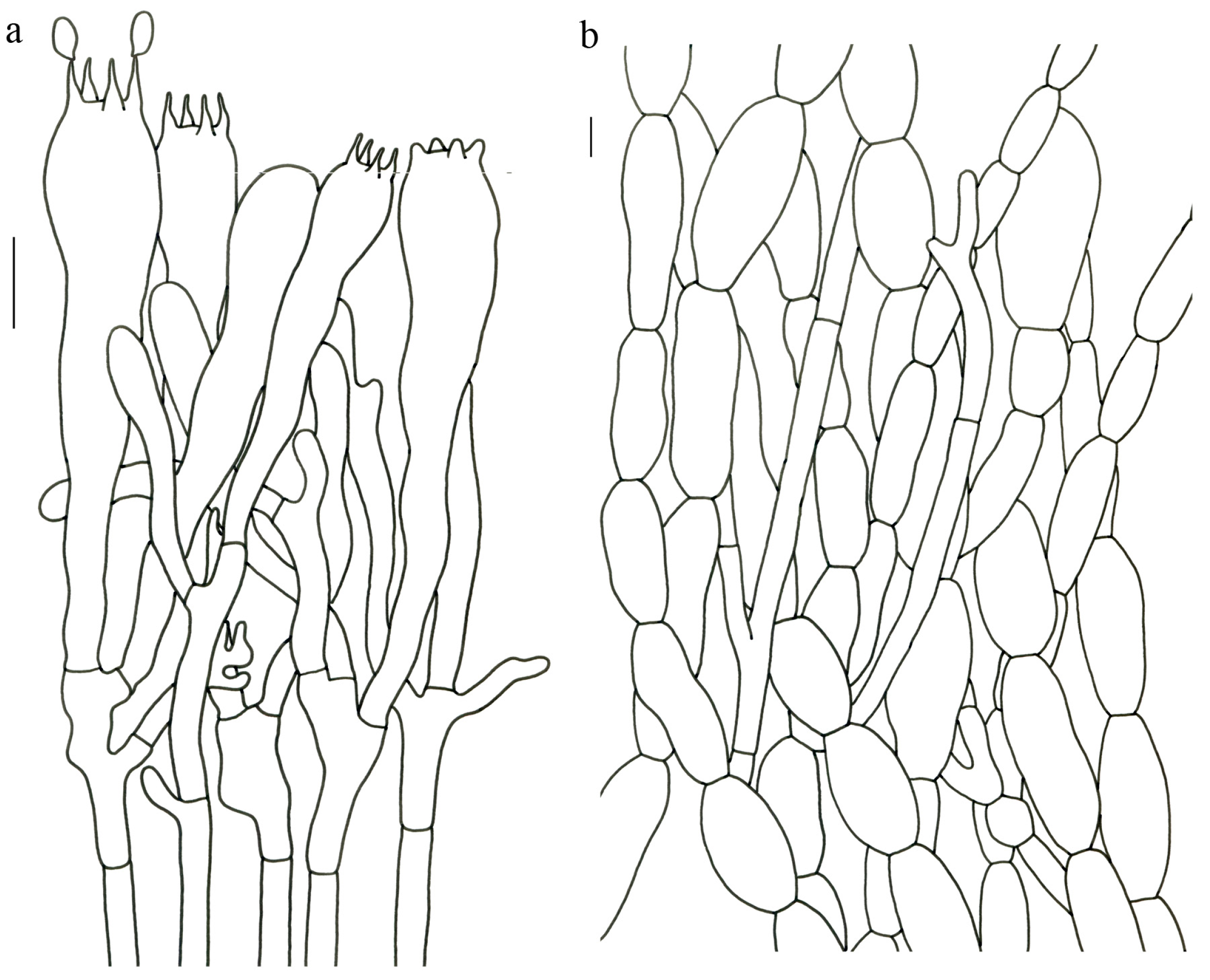
Figure 4 and
Figure 5).
MycoBank: MB 843943
Basionym—Gomphus szechwanensis R.H. Petersen, Nova Hedwigia 21: 102 (1972) [“1971”].
Epitype—CHINA. SICHUAN: Garzê Prefecture, Kangding County, Yala Township, Mugecuo Scenic Spot, 9 September 2016, Bang Feng 130 (HKAS 99484, here designated! Genbank OQ858484/OQ858556/OQ847641).
Description—Basidioma 14–25 (–30) cm tall; unipileate; tapering downward. Pileus up to 17 cm in diam.; funnel-shaped; surface orange-ochre to brownish orange; pellicle-like. Hymenium surface clay colored; strongly wrinkled; irregularly reticulate and extending longitudinally to the upper stipe; dense. Stipe approximately cylindrical; rounded at base; hollow; upper part off-white and becoming reddish toward the base; glabrous above the base; context white.
Basidiospores 16–19.5 (–20) × 7.5–11 µm; oblong and inequilateral in side view with distinct suprahilar depression; ellipsoid in ventral view; non-amyloid; dull ochre in KOH; ornamentation of very coarse ridges or warts. Basidia 85–110 × 10–13 µm; clavate; generally 2–4-spored. Hymenial cystidia absent. Pileipellis composed of profusely inflated hyphae (8–21 μm in diam.), mostly with ampulliform swellings and little uninflated hyphae; simple branched; strictly parallel arranged toward pileal context; gloeoplerous hyphae abundant. Clamp connections absent in all parts of basidioma.
Habitat and locality—Gregarious in forests of Abies, Picea, Pinus densata and Quercus semicarpifolia in Southwest and central China at (2400–) 3200–4000 m altitude.
Specimens examined—CHINA. SICHUAN: Garzê Prefecture, Kangding County, locality unknown, 1894, M. Farges (Herb. Patouillard, HF, type of G. szechwanensis). Garzê Prefecture, Daofu County, Geka Township, 17 July 2014, Bang Feng 1598 (HKAS 93982). Garzê Prefecture, Xiangcheng County, Great Snow Mountain, in forests with Abies and Rhododendron, alt. 4000 m, Li-Song Wang 942 (HKAS 7871). HUBEI: Yichang City, Xingshan County, Shennongding Nature Reserve, 17 July 2012, Jiao Qin 569 (HKAS 77970). Yichang City, Xingshan County, Shennongding Nature Reserve, alt. 2400 m, 18 July 2012, Jiao Qin 586 (HKAS 77988). GANSU: Gannan Prefecture, Zhouqu County, Shatan Forest Park, in forests with Abies, 16 August 2012, Xue-Tai Zhu 730 (HKAS 76579). YUNNAN: Diqing Prefecture, Deqin County, Baima Snow Mountain, alt. 3700 m, Xing-Jiang Li 1142 (HKAS 8159). Diqing Prefecture, Xianggelila City, Xiaozhongdian Town, alt. 3500 m, 3 September 2018, Jian-Wei Liu 1597 (HKAS 122959). Diqing Prefecture, Xianggelila City, Xiaozhongdian Town, in forests with Abies, 2 September 2013, Bang Feng 1496 (HKAS 82602). Diqing Prefecture, Xianggelila City, Zhongdian Town, Bitahai Natural Reserve, in forests with Picea, alt. 3628 m, 3 September 2013, Bang Feng 1508 (HKAS 82614). Lijiang City, Yulong County, Alpine Garden, alt. 3525 m, 28 August 2019, Jian-Wei Liu 1711 (HKAS 122960). Lijiang City, Yulong County, Yulong Snow Mountain, in forests with Pinus densata, alt. 3231 m, 12 July 2010, Xue-Tai Zhu 24 (HKAS 68200). TIBET: Bomi County, on the way from Bomi to 24K, alt. 3257 m, 26 July 2017, Si-Peng Jian 46 (HKAS 101035). Bomi County, on the way from Bomi to Linzhi, alt. 2400 m, 18 July 2019, Zhu-Liang Yang 6201 (HKAS 106813). Gongbujiangda County, Niangdang Village, in forests with Abies, alt. 3812 m, 30 July 2014, Bang Feng 1656 (HKAS 94040). Rikaze City, Nielamu County, alt. 2400–2700 m, 3 September 2006, Jun-Feng Liang 613 (HKAS 51324). Linzhi County, National Highway 318, 757 m of northeast of Lulang Military Station, in forests of Picea, 29°49’ N, 94°44’ E, 22 July 2019, Geng-Shen Wang 511 (HKAS 116225). Linzhi County, Lulang Town, in forests of Quercus semicarpifolia, 29°58’ N, 94°51’ E, 18 July 2019, Geng-Shen Wang 436 (HKAS 116137).
Notes—
Gomphus floccosus (Schwein.) Singer, under the name of
Cantharellus floccosus Schwein., was first reported in China by Patouillard (1895), based on a collection made from Tchen-KéouTin (currently called Kangding) by M. Farges in 1894. This collection was treated as a new species by Petersen [
6] and Zang et al. [
59], namely
Gomphus szechwanensis. Giachini [
1] and Giachini and Castellano [
60] synonymized
G. szechwanensis under
T. floccosus, it was originally described from North America, based on morphological analysis. However, our morphological and molecular data indicated that it is a species of
Turbinellus and indeed differs from
T. floccosus (
Figure 1,
Figure 4 and
Figure 5); thus, a new combination was proposed. In order to fix the concept of the species, an epitype for the species was selected in this paper.
As mentioned by Petersen [
6],
T. szechwanensis is characterized by its larger basidioma exceeding 30 cm in height, broader pileus up to 17 cm in diam., pellicle-like surface of the pileus and coarsely decorated larger basidiospores.
Turbinellus floccosus differs from this species by its significantly smaller basidiospores (10–16 × 5–8 µm in Arora [
61]; 11.5–14.5 × 7–8 µm in Bessette et al. [
62]). This species is closely related to
T. verrucosus in the phylogenetic analysis (
Figure 1), but the latter has a smaller basidioma less than 7 cm tall, a narrower pileus (2–4.5 cm in diam.) with verrucous scales and smaller basidiospores measuring 13–15.5 × 6–8 µm. Ecologically,
T. szechwanensis mainly scatters in forests of
Abies,
Picea and
Pinus densata at 3200–4000 m altitude. This is in contrast to several other species that occur at lower elevations, such as
T. verrucosus,
T. squamosus,
T. solidus and so on. To date, only
T. szechwanensis and
T. flavidus of the genus have been founded in central parts of China.
MycoBank: MB 843965
Etymology—From the Latin tomentosipes, referring to the stipe of the new species covered with long tomentum.
Holotypus—CHINA. YUNNAN: Kunming City, Panlong District, Wild Duck Lake Park, alt. 2000–2200 m, 11 August 2020, Fei-Fei Liu 251 (HKAS 113156; Genbank OQ858453/OQ858521/OQ847611).
Diagnosis—Turbinellus tomentosipes is similar to T. solidus, but the former possesses a larger basidioma up to 15 cm tall, a densely tomentose stipe and profusely inflated hyphae in stipitipellis. The latter has a smaller basidioma less than 10 cm tall, a glabrous stipe and little inflated hyphae in stipitipellis.
Description—Basidioma 5–15 cm tall; unipileate or bipileate; tapering downward. Pileus 2–8 cm in diam.; funnel-shaped; depressed at center; surface orange-yellow (5A7–8) when young and faint yellow (1A2–4) with age; rough and sparsely covered with minute appressed scales; scales more or less radially arranged and usually darker than the pileal surface; appearing glutinous; pileal margin extensional and undulate. Hymenium surface white (1A1) to faint yellow (1A2–4) or creamy orange (4A2–3); weakly wrinkled; longitudinal ridges extending to the upper stipe; branching irregularly upwards; sparse; thin to 0.2 cm. Stipe about 3–6 cm long, 0.7–2 cm in diam.; approximately cylindrical; solid; creamy orange (4A3) when young and creamy (1A3) with age; densely covered with long creamy yellow (4A2) tomentum; context pliable; thin to 0.2–0.4 cm; white (1A1) to creamy (1A2); unchanging on exposure; basal mycelium white (1A1).
Basidiospores [63/3/3] 11–14 × 5.5–7.5 (–8) µm; Q = (1.53–) 1.58–2.15 (–2.36); Qm = 1.91 ± 0.17; oblong and inequilateral in side view with distinct suprahilar depression; ellipsoid in ventral view; non-amyloid; light yellowish in KOH; ornamentation of very coarse ridges or warts. Basidia 50–75 × 8–13.5 µm; clavate, 2–5-spored; sterigmata 4–7 µm long. Hymenial cystidia absent. Pileipellis composed of profusely inflated hyphae (7–24 μm in diam.); mostly with ampulliform swellings and little uninflated hyphae (3–6 μm in diam.); simple branched; constricted at septa; repent; more or less parallel arranged toward the pileal context; gloeoplerous hyphae abundant; yellow in KOH. Stipitipellis composed of some basidia-like cells; profusely uninflated hyphae (2–6 μm in diam.) and some inflated hyphae (7–12 μm in diam.); gloeoplerous hyphae abundant; yellow in KOH. Pileus and stipe context composed of simple branched hyphae; approximately parallel; hyaline to yellow in KOH. Hymenial trama composed of subparallel to interwoven, undifferentiated and narrow hyphae. Clamp connections absent in all parts of basidioma.
Habitat and distribution—Scattered in subtropical forests dominated by plants of Pinus yunnanensis, Quercus and Rhododendron decorum; fruiting from June to August in southwestern China (Yunnan Province) between 1000 and 2200 m altitude.
Additional specimens examined—CHINA. YUNNAN: Chuxiong Prefecture, Lufeng City, Heijing Town, in broad-leaved forests, alt. 1028 m, 10 July 2014, Xiao-Bin Liu 419 (HKAS 87066). Dali Prefecture, Binchuan County, Jizushan Town, Shanqiansi Village, 23°45′ N, 105°12′ E, alt. 1350 m, 11 August 2011, Ting Guo 436 (HKAS 71332). Dali Prefecture, Binchuan County, Jizushan Town, Siqian Village, in forests of Pinus yunnanensis, Quercus and Rhododendron decorum, 25°56′ N, 100°23′ E, alt. 1988 m, 11 August 2011, Li-Ping Tang 1541 (HKAS 70002). Wenshan Prefecture, Qiubei County, Jinping Town, Xiangqi Village, in forests with Fagaceae and Pinus yunnanensis, alt. 1570 m, 10 August 2014, Pan-Meng Wang 141 (HKAS 95222).
Notes—Turbinellus tomentosipes is similar to T. solidus, but the latter possesses a smaller basidioma, a glabrous stipe and filamentous hyphae in stipitipellis. Geographically, T. tomentosipes occurs in northern parts of Yunnan, while T. solidus fruits in its southern parts.
MycoBank: MB 843966
Etymology—From the Latin adjective, verrucosus, referring to the pileus densely covered with verrucose protuberances of the new species.
Holotypus—CHINA. YUNNAN: Nujiang Prefecture, Lushui City, Laowo Township, Chongren Village, mainly in forests with Pinus yunnanensis, supplemented by Fagaceae and Ericaceae, alt. 1700–1800 m, 7 August 2011, Gang Wu 546 (HKAS 74860; Genbank OQ858538).
Diagnosis—Turbinellus verrucosus is similar to T. solidus in having small basidioma, orange pileus and small basidiospores. However, the former has a pileus with densely verrucous protuberances, a hollow stipe and little inflated hyphae in stipitipellis. The latter has a pileus with sparsely appressed scales, a solid stipe and plenty of inflated hyphae in stipitipellis.
Description—Basidioma 4–6.5 cm tall; unipileate; tapering downward. Pileus 2.2–4.5 cm in diam.; funnel-shaped; depressed at center; surface orange (5A5–8) to dark red (7C6); rough and densely covered with verrucous protuberances; protuberances more or less radially arranged; pileal margin in-rolled and nearly entire. Hymenium surface off-white (1A1) to light yellow (2A4) or creamy orange (5A2); strongly wrinkled; longitudinally reticulate and extending to the stipe base; dense; thick to 0.2–0.4 cm; not reaching creamy yellow (4A3) to orange (5A6); margin upwards. Stipe about 1.5–3.5 cm long; 0.5–0.8 cm in diam.; approximately cylindrical; occasionally dilating obconically into the pileus; hollow; white (1A1) to creamy (2A3) or creamy orange (5A3); glabrous above the base; context pliable; thin to 0.1–0.2 cm; white (1A1); unchanging on exposure; basal mycelium white (1A1).
Basidiospores [61/3/3] 13–15.5 (–16) × 6–8 (–8.5) µm; Q = (1.69–) 1.86–2.25 (–2.33); Qm = 2.01 ± 0.14; oblong to cylindrical and inequilateral in side view with distinct suprahilar depression; ellipsoid in ventral view; non-amyloid, light yellowish in KOH; ornamentation moderately verrucose with coarse warts. Basidia 70–95 × 11–16 µm; clavate; generally 4-spored; sterigmata 5–10 µm long. Hymenial cystidia absent. Pileipellis composed of profusely inflated hyphae (7–19 μm in diam.) mostly with ampulliform swellings and little uninflated hyphae (3–6 μm in diam.); simple branched; constricted at septa; repent; more or less parallel arranged toward the pileal context; gloeoplerous hyphae abundant; yellow in KOH. Stipitipellis composed of some basidia-like cells; plenty of uninflated hyphae (2–6 μm in diam.) and little inflated hyphae (7–10 μm in diam.); gloeoplerous hyphae abundant; yellow in KOH. Pileus and stipe context composed of simple branched hyphae; approximately parallel; hyaline to yellow in KOH. Hymenial trama composed of subparallel to interwoven, undifferentiated and narrow hyphae (3–6 μm in diam.). Clamp connections absent in all parts of basidioma.
Habitat and distribution—Scattered in subtropical forests with Pinus; fruiting from June to August in southwestern China (Yunnan Province) between 1250 and 2100 m altitude.
Additional specimens examined: CHINA. YUNNAN: Baoshan City, Longyang District, Lujiang Township, Pumanshao, in the mixed coniferous broad-leaved forests, alt. 2033 m, 23 March 2014, Xiao-Bin Liu 708 (HKAS 87257). Baoshan City, Changning County, Wojiaodi Village, alt. 1784 m, 24 July 2009, Li-Ping Tang 931 (HKAS 56888). Dali Prefecture, Heqing County, Xiyi Town, Beiya Village, 26 August 2018, Jian-Wei Liu 1180 (HKAS 122949). Puer City, Lancang County, Donghui Township, in forests with Pinus kesiya, alt. 1260 m, 29 August 2017, Zhu-Liang Yang 5996 (HKAS 101185). Puer City, Lancang County, Menglang Township, Zhazhai Village, 21 August 2016, Jian-Wei Liu 50 (HKAS 97554). Puer City, Jiangcheng County, 29 July 2008, Li-Ping Tang 511 (HKAS 54742). Sipsongpanna, Jinghong City, alt. 1400 m, 1 August 2008, Xi-Hui Du 19 (HKAS 55476).
Notes—
Turbinellus verrucosus is distinguished by the combination characters of the small basidioma, orange to orange-red pileus densely covered with verrucous protuberances and hollow stipe. This species can be easily confused with
T. floccosus, native to North America, due to their similar appearance of basidiomata. However,
T. floccosus possesses a significantly larger basidioma (5–20 cm), a larger pileus (3–15 cm) and a thicker stipe (3–10 × 1–3 cm) [
61]. Moreover, our molecular data resolved
T. floccosus as a lineage independent of those
T. floccosus-like fungi in China (
Figure 1).
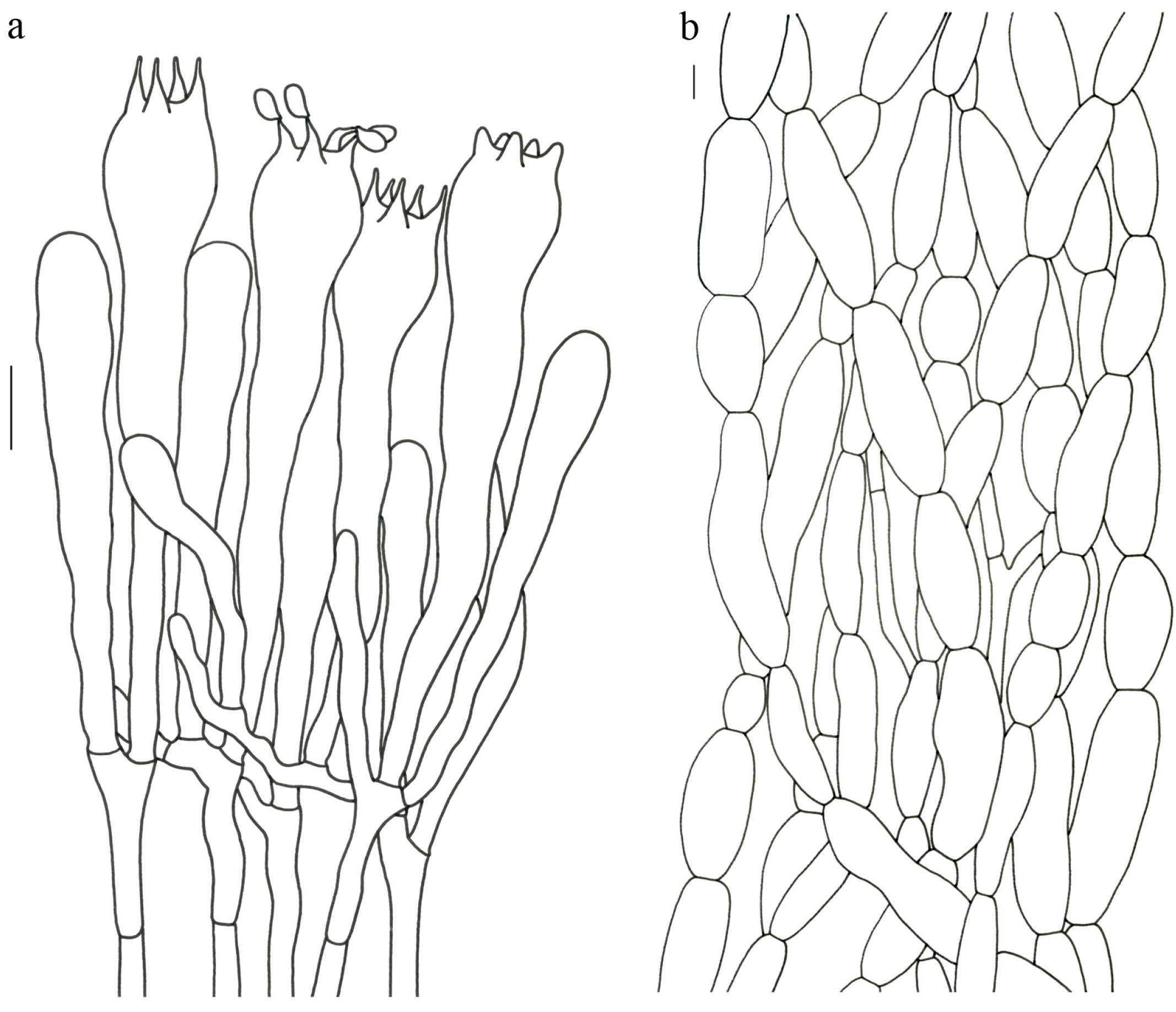
Turbinellus yunnanensis (R.H. Petersen & M. Zang) Xue-Ping Fan & Zhu L. Yang,
comb. nov. (
Figure 4 and
Figure 5).
MycoBank: MB 843944
Basionym—Gomphus yunnanensis R.H. Petersen and M. Zang, in Zang, Li and Xi, Fungi of the Hengduan Mountains: 181 (1996).
Description—Basidioma up to 12 cm tall; unipileate; tapering downward. Pileus up to 7 cm in diam.; funnel-shaped; depressed at center; surface orange to orange-red; sparsely covered with minute appressed scales; pileal margin undulate to entire. Hymenium surface orange-red to reddish; wrinkled; irregularly reticulate and extending to the upper stipe; sparse. Stipe 1–4 cm long; 1–1.7 cm in diam.; subcylindrical; hollow; orange-red to reddish; glabrous above the base; context white.
Basidiospores 13–17 × 6.5–7.5 µm; oblong and inequilateral in side view with distinct suprahilar depression; oblong in ventral view; non-amyloid; ornamentation of very densely coarse ridges or warts. Hymenial cystidia absent. Gloeoplerous hyphae abundant in pileipellis, stipitipellis and context. Clamp connections absent in all parts of basidioma.
Habitat and locality—Gregarious in forests of Abies and Picea; fruiting from summer to autumn in southwestern China between 2800 and 3200 m altitude.
Specimens examined—CHINA. YUNNAN: Lijiang City, Yulong County, Yulong Snow Mountain, in forests of Abies and Picea, alt. 3000–3200 m, 3 September 1986, R.H. Petersen 56,349 (HKAS 18142, holotype of Gomphus yunnanensis; Genbank OQ858460/OQ858528/OQ847616). Lijiang City, Yulong County, Yulong Snow Mountain, in forests of Picea, alt. 3000 m, 6 September 1986, R.H. Petersen 56,818 (HKAS 18145). Diqing Prefecture, Xianggelila City, Haba Snow Mountain, alt. 2858 m, 1 October 2007, Bang Feng 195 (HKAS 52916). Lijiang City, Yulong County, Yulong Snow Mountain, in forests of Pinus densata, alt. 3048 m, 18 August 2013, Yang-Yang Cui 115 (HKAS 79785). Lijiang City, Yulong County, Alpine Garden, 26 September 2019, Jian-Wei Liu 2143 (HKAS 122962). Lijiang City, Yulong County, Lijiang Observatory, in forests of Picea, Pinus and Quercus, alt. 3200 m, 17 August 2014, Qi Zhao 2147 (HKAS 87913).
Notes—
Turbinellus yunnanensis was once placed in the genus
Gomphus [
59]. Our morphological and molecular data indicated that it is a species of
Turbinellus (
Figure 1,
Figure 4 and
Figure 5); thus, a new combination was made.
Turbinellus yunnanensis is characterized by its reddish basidioma, orangish pileus, hollow stipe and medium-sized basidiospores (13–17 × 6.5–7.5 µm).
Turbinellus yunnanensis is similar to
T. longistipes and
T. solidus in having orange-yellow to orange pileus with minute appressed scales. However,
T. yunnanensis differs from
T. longistipes in its shorter cylindrical stipe and smaller basidiospores from
T. solidus with its reddish basidioma and hollow stipe. Furthermore,
T. yunnanensis is distributed in forests of
Abies,
Picea,
Pinus densata and
Quercus at 2800–3200 m altitude, while
T. longistipes occurs in forests of
Abies,
Picea,
Pinus densata and
Quercus semicarpifolia at 3200–3600 m altitude in the northern parts of Yunnan.
T. solidus occurs in forests of Fagaceae and
Pinus at 1300–1600 m altitude in the area’s south.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}















