Ion-Induced Volume Transition in Gels and Its Role in Biology



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Ion-Induced Volume Transition
2.1. Degree of Swelling
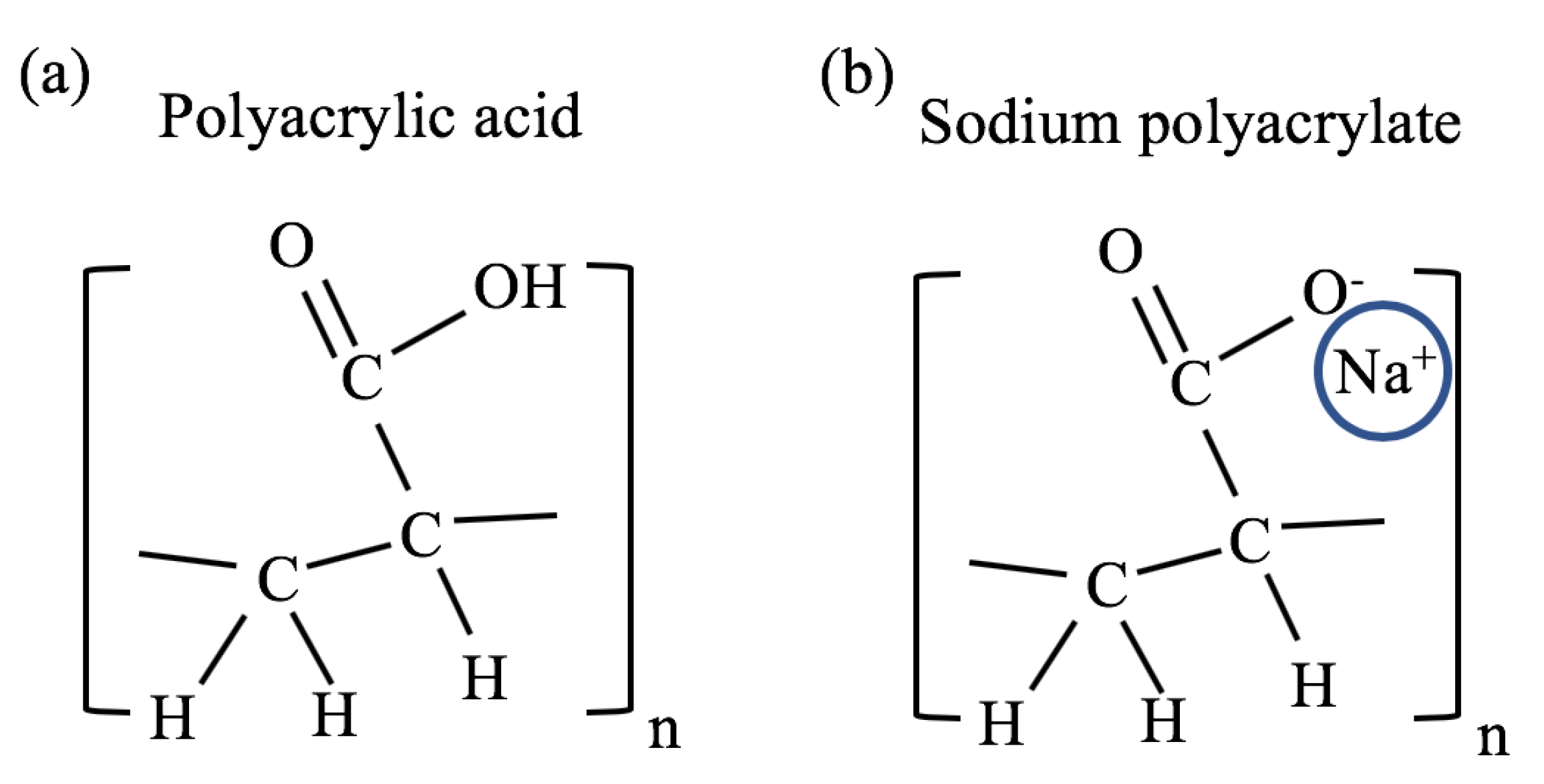
2.1.1. Effect of Monovalent Counterions on Gel Swelling
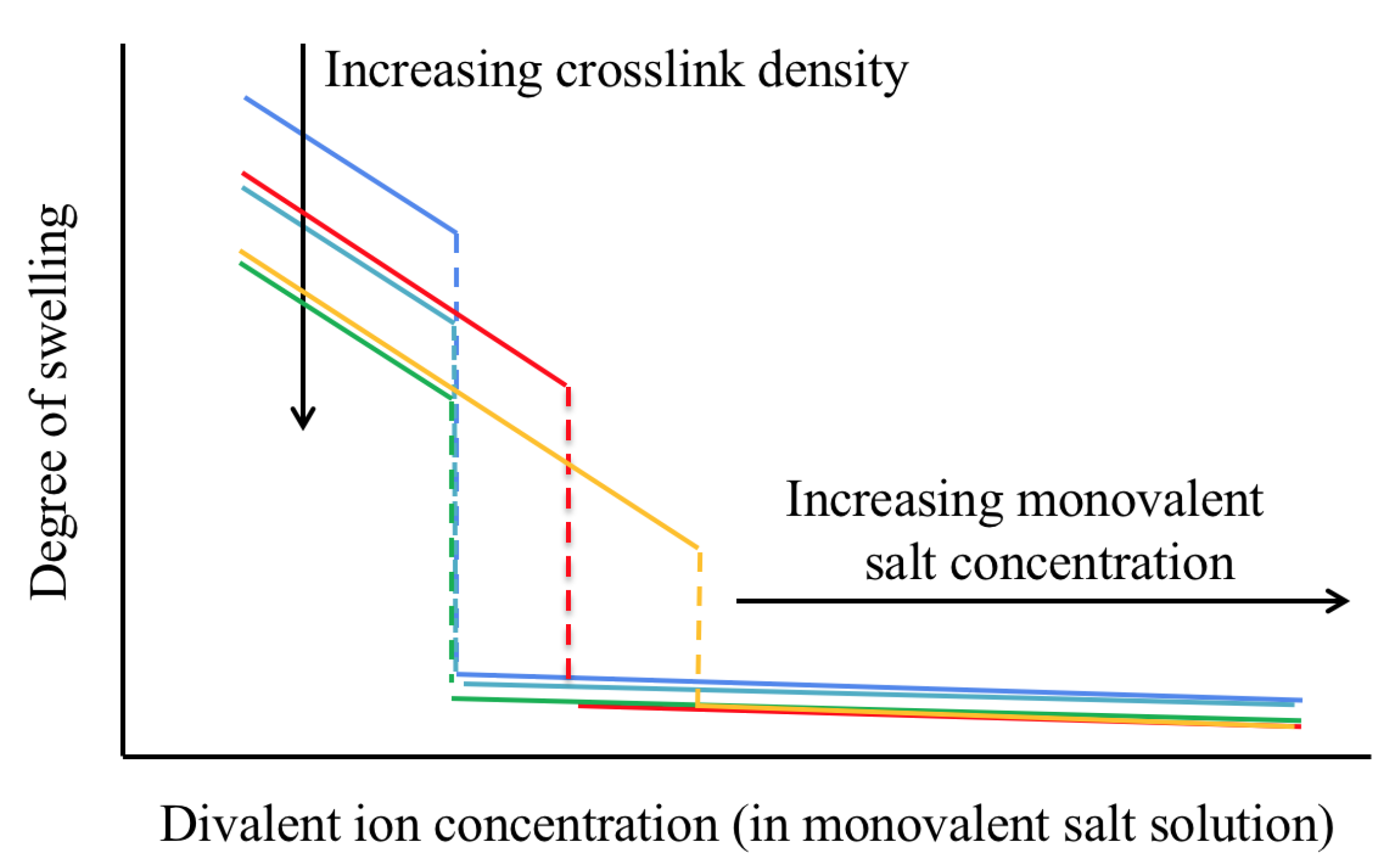
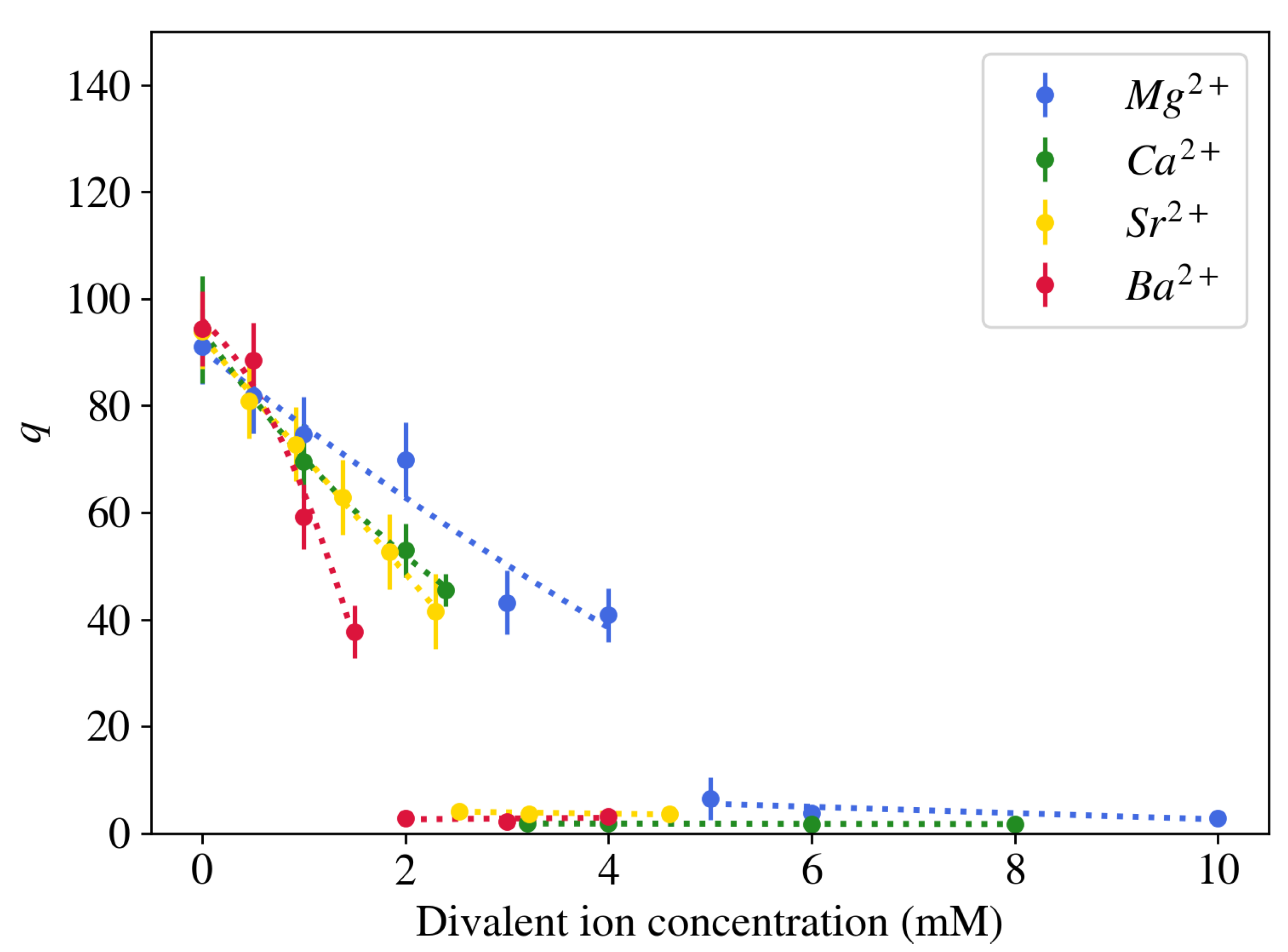
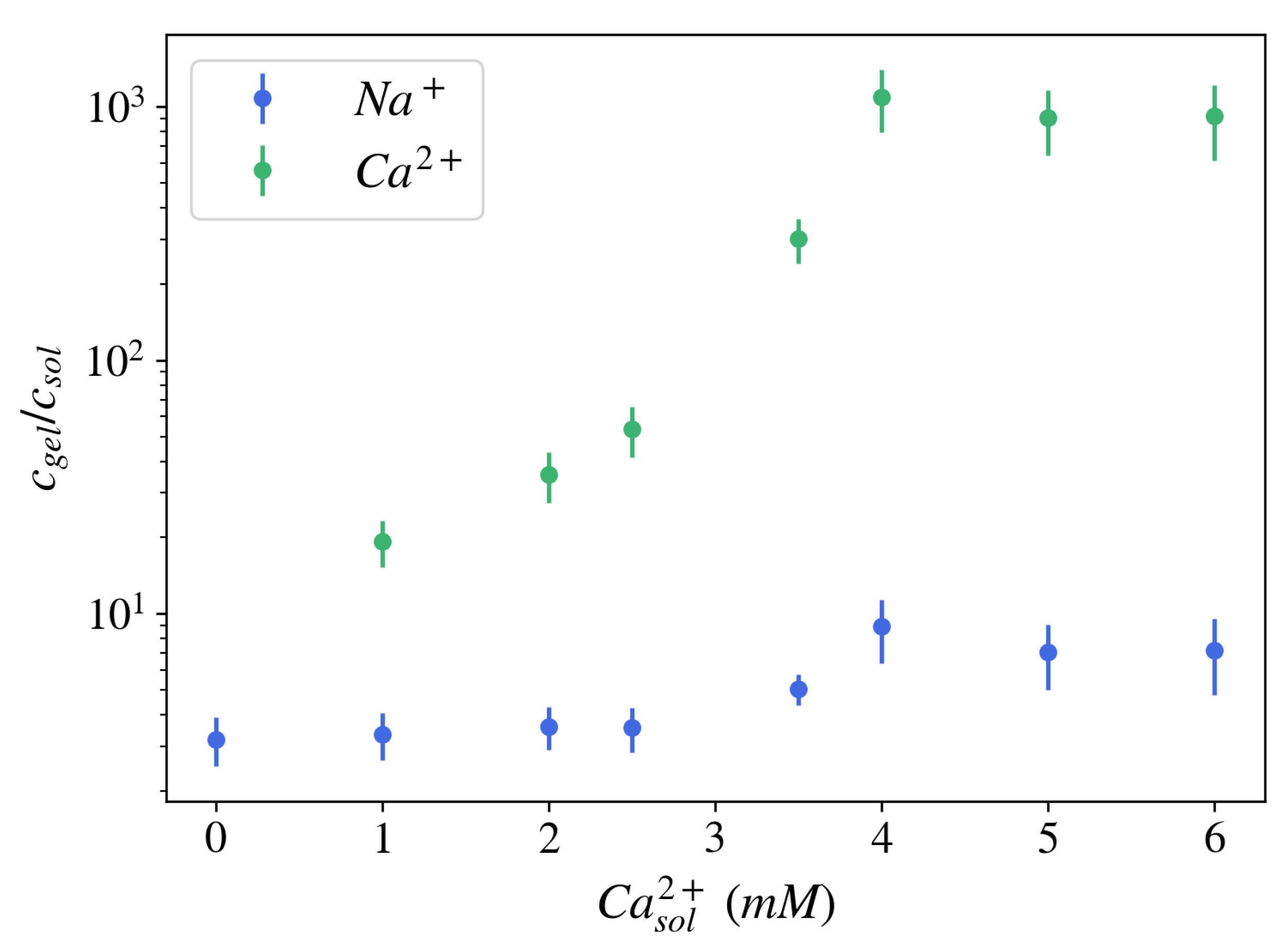
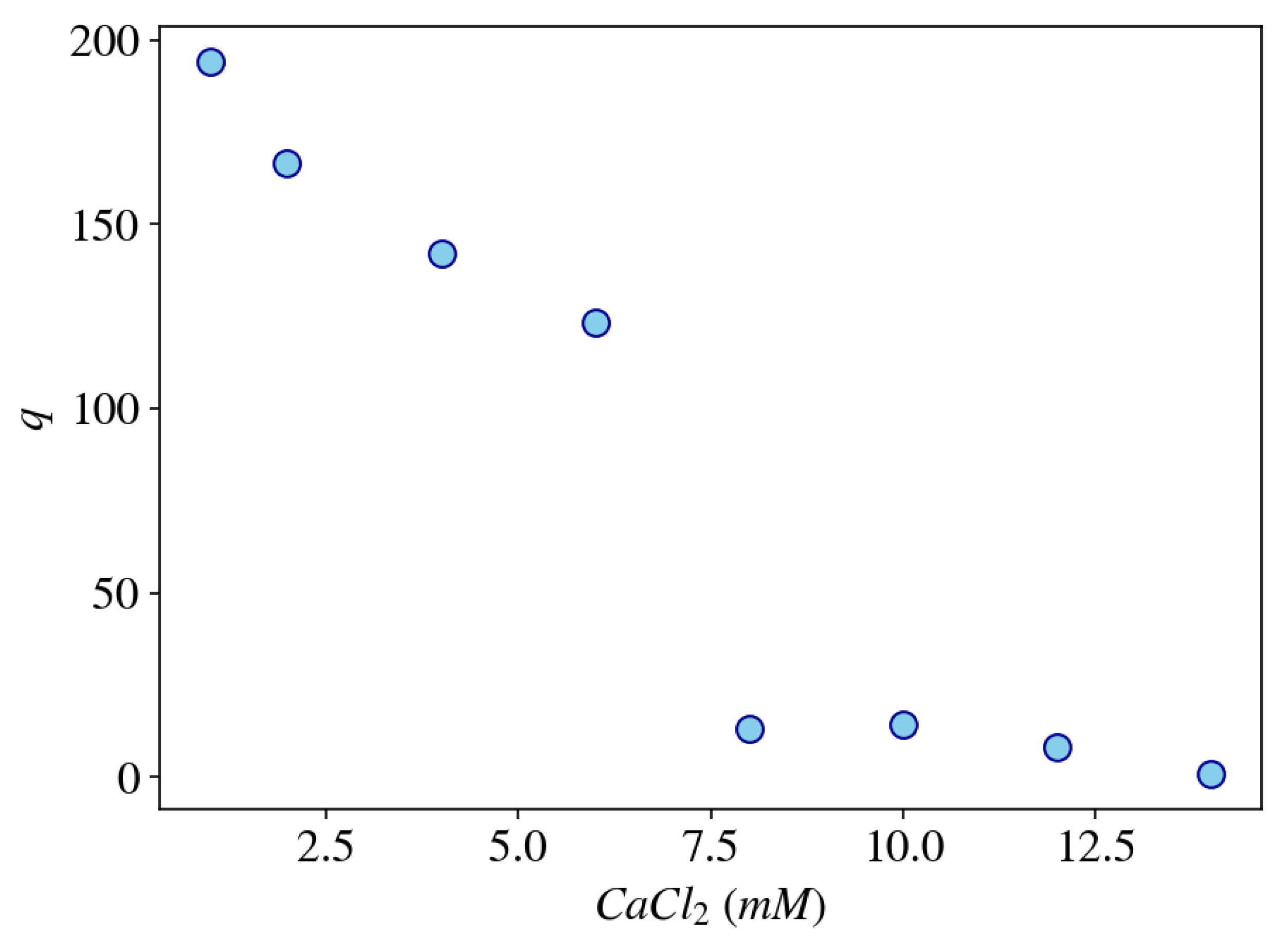
2.1.2. Effect of Multivalent Counterions on Gel Swelling
2.2. NMR Parameters
2.3. Electric Potential Difference and Ion Partitioning
3. Evidence of Ion-Induced Abrupt Transition in Biological Systems
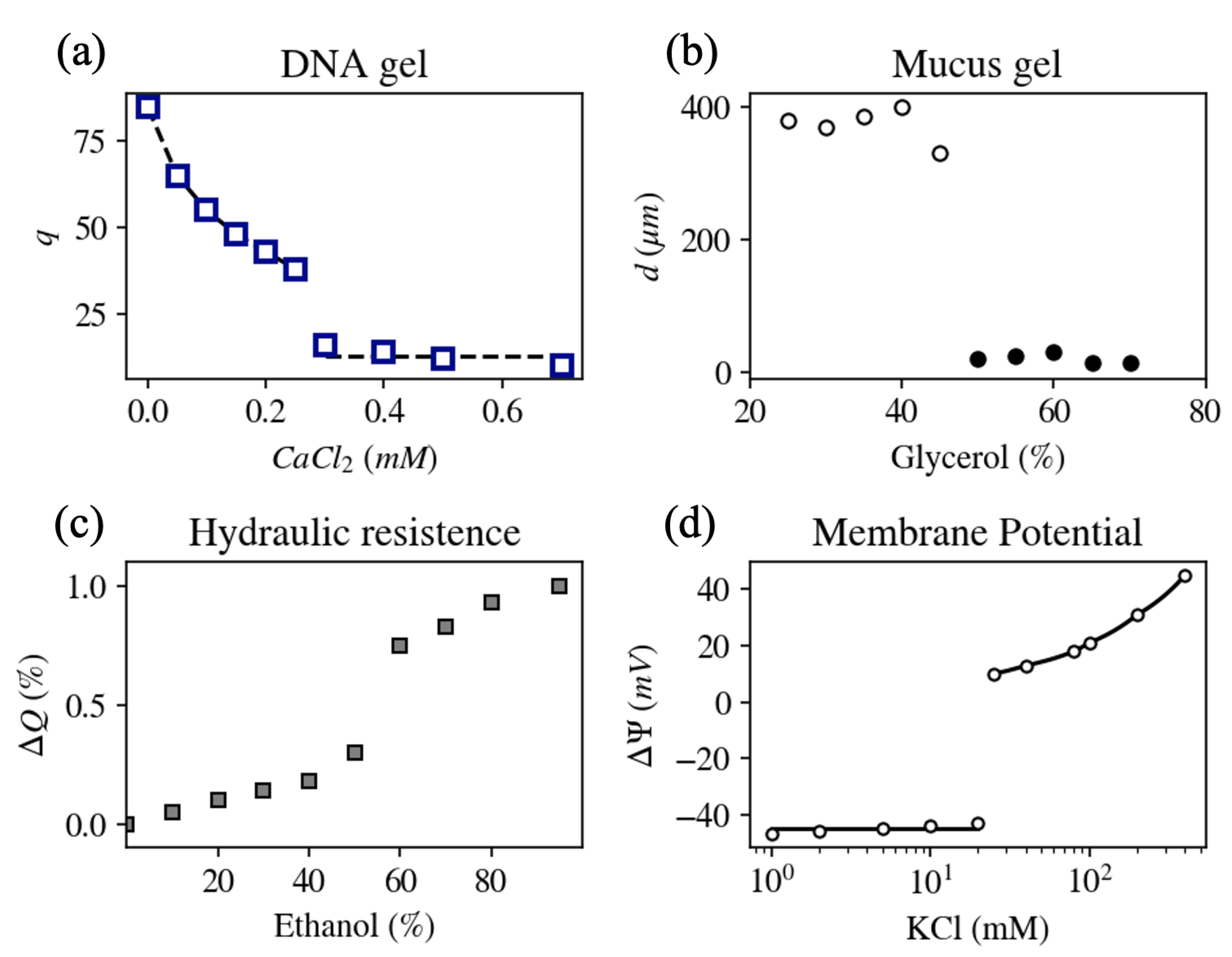
3.1. The Compaction Process of DNA Molecules
3.2. Storage and Release of Secretory Products
3.3. The Ability to Change the Hydraulic Resistance of Xylem in Plants
3.4. Abrupt Depolarization and the Cell Resting Potential
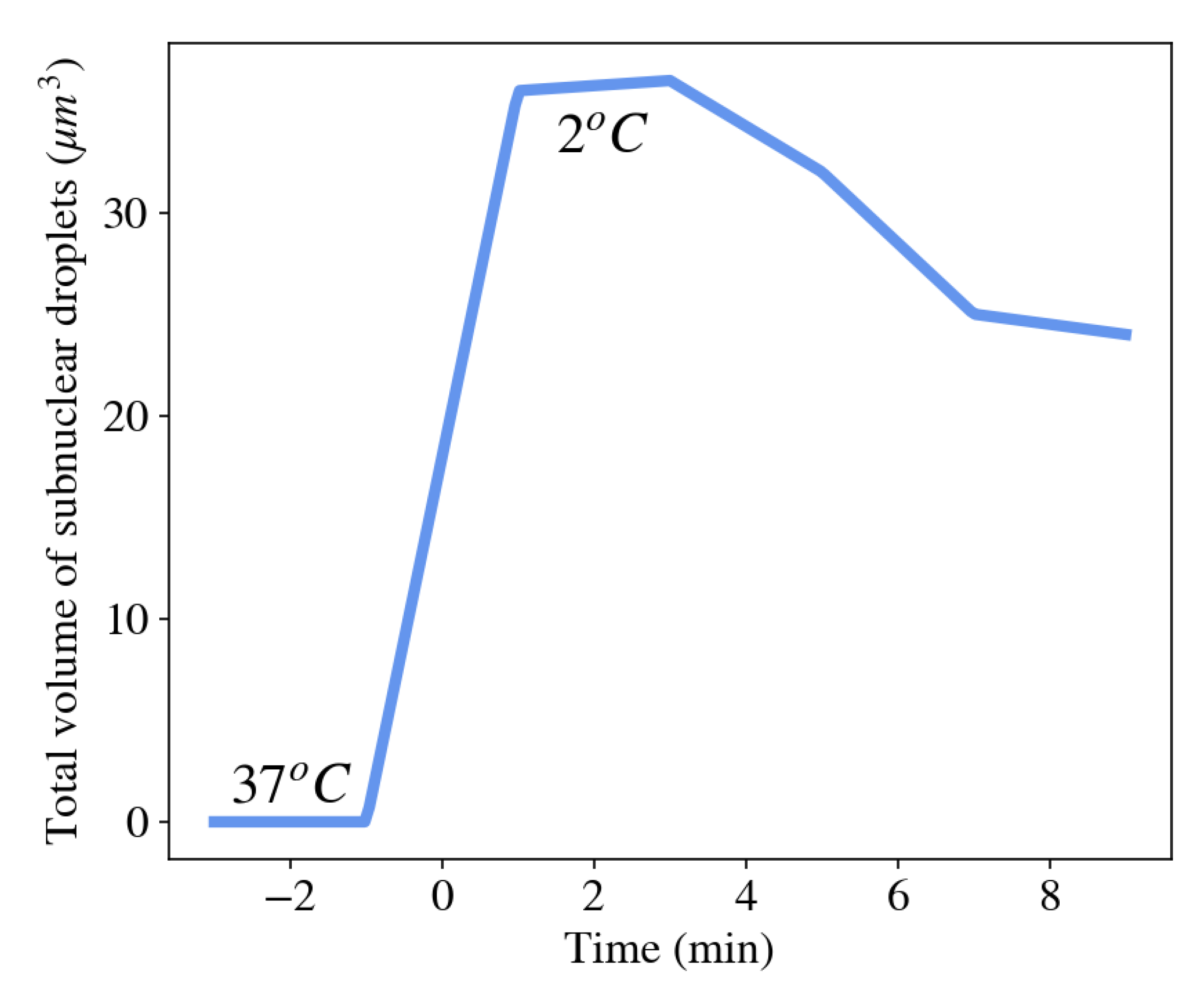
3.5. Formation of Membraneless Organelles
4. Theoretical Approaches to Quantify Gel-Related Biological Aspects
5. Conclusions and Outlook
Funding
Conflicts of Interest
Abbreviations
| NaPA | sodium polyacrylate |
| SANS | Small angle neutron scattering |
| SAXS | Small angle X-ray scattering |
| ADC | Apparent diffusion coefficient |
References
- Boal, D.H. Mechanics of the Cell; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Jansen, R.C. Studying complex biological systems using multifactorial perturbation. Nat. Rev. Genet. 2003, 4, 145. [Google Scholar] [CrossRef] [Green Version]
- Anderson, P.W. More is different. Science 1972, 177, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Goldenfeld, N.; Kadanoff, L.P. Simple lessons from complexity. Science 1999, 284, 87–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katchalsky, A. Polyelectrolytes and Their Biological Interactions. Biophys. J. 1964, 4, 9–41. [Google Scholar] [CrossRef] [Green Version]
- Tasaki, I. Physiology and Electrochemistry of Nerve Fibers; Academic Press: New York, NY, USA, 1982. [Google Scholar]
- Verdugo, P. Polymer gel phase transition in condensation-decondensation of secretory products. Adv. Polym. Sci. 1993, 110, 145–156. [Google Scholar]
- Marguet, M.; Bonduelle, C.; Lecommandoux, S. Multicompartmentalized polymeric systems: Towards biomimetic cellular structure and function. Chem. Soc. Rev. 2013, 42, 512–529. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, M.; Colby, R.H. Polymer Physics; Oxford University Press: New York, NY, USA, 2003; Volume 23. [Google Scholar]
- Israelachvili, J.N. Intermolecular and Surface Forces; Academic Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Tongu, C.; Kenmotsu, T.; Yoshikawa, Y.; Zinchenko, A.; Chen, N.; Yoshikawa, K. Divalent cation shrinks DNA but inhibits its compaction with trivalent cation. J. Chem. Phys. 2016, 144, 205101. [Google Scholar] [CrossRef] [Green Version]
- Zwieniecki, M.A.; Melcher, P.J.; Holbrook, N.M. Hydrogel control of xylem hydraulic resistance in plants. Science 2001, 291, 1059–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brangwynne, C.P. Phase transitions and size scaling of membrane-less organelles. J. Cell Biol. 2013, 203, 875–881. [Google Scholar] [CrossRef] [Green Version]
- Amiya, T.; Tanaka, T. Phase Transitions in Cross-Linked Gels of Natural Polymers. Macromolecules 1987, 20, 1162–1164. [Google Scholar] [CrossRef]
- Horkay, F.; Tasaki, I.; Basser, P.J. Osmotic Swelling of Polyacrylate Hydrogels in Physiological Salt Solutions. Biomacromolecules 2000, 1, 84–90. [Google Scholar] [CrossRef]
- Mussel, M.; Basser, P.J.; Horkay, F. Effects of mono- and divalent cations on the structure and thermodynamic properties of polyelectrolyte gels. Soft Matter 2019, 15, 4153–4161. [Google Scholar] [CrossRef]
- Mussel, M.; Horkay, F. Experimental Evidence for Universal Behavior of Ion-Induced Volume Phase Transition in Sodium Polyacrylate Gels. J. Phys. Chem. Lett. 2019, 10, 7831–7835. [Google Scholar] [CrossRef]
- Borkovec, M.; Koper, G.J.M.; Piguet, C. Ion binding to polyelectrolytes. Curr. Opin. Colloid Interface Sci. 2006, 11, 280–289. [Google Scholar] [CrossRef]
- Joesten, M.D.; Schaad, L.J. Hydrogen Bonding; Marcel Dekker Inc.: New York, NY, USA, 1974. [Google Scholar]
- Elliott, J.E.; Macdonald, M.; Nie, J.; Bowman, C.N. Structure and swelling of poly (acrylic acid) hydrogels: Effect of pH, ionic strength, and dilution on the crosslinked polymer structure. Polymer 2004, 45, 1503–1510. [Google Scholar] [CrossRef]
- Meister, E.C.; Willeke, M.; Angst, W.; Togni, A.; Walde, P. Confusing Quantitative Descriptions of Brønsted-Lowry Acid-Base Equilibria in Chemistry Textbooks—A Critical Review and Clarifications for Chemical Educators. Helv. Chim. Acta 2014, 97, 1–31. [Google Scholar] [CrossRef]
- Hornback, J.M. Organic Chemistry; Brooks: Seattle, WA, USA, 1998. [Google Scholar]
- Horkay, F.; Hecht, A.M.; Rochas, C.; Basser, P.J.; Geissler, E. Anomalous small angle x-ray scattering determination of ion distribution around a polyelectrolyte biopolymer in salt solution. J. Chem. Phys. 2006, 125, 234904. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.W.; Horkay, F.; Douglas, J.F.; Pablo, J.J.D. Molecular simulation of the swelling of polyelectrolyte gels by monovalent and divalent counterions. J. Chem. Phys. 2008, 129, 154902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrat, J.-L.; Joanny, J.-F.; Pincus, P. On the scattering properties of polyelectrolyte gels. J. Phys. II 1992, 2, 1531–1544. [Google Scholar] [CrossRef]
- Arens, L.; Barther, D.; Landsgesell, J.; Holm, C.; Wilhelm, M. Poly(sodium acrylate) hydrogels: Synthesis of various network architectures, local molecular dynamics, salt partitioning, desalination and simulation. Soft Matter 2019, 15, 9949–9964. [Google Scholar] [CrossRef] [Green Version]
- Morfin, I.; Horkay, F.; Basser, P.J.; Bley, F.; Hecht, A.M.; Rochas, C.; Geissler, E. Adsorption of divalent cations on DNA. Biophys. J. 2004, 87, 2897–2904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horkay, F.; Tasaki, I.; Basser, P.J. Effect of Monovalent-Divalent Cation Exchange on the Swelling of Polyacrylate Hydrogels in Physiological Salt Solutions. Biomacromolecules 2001, 2, 195–199. [Google Scholar] [CrossRef]
- Sircar, S.; Roberts, A.J. Ion mediated crosslink driven mucous swelling kinetics. Discret. Contin. Dyn. Syst. Ser. B 2015, 21, 1937–1951. [Google Scholar] [CrossRef]
- Horkay, F.; Basser, P.J. Ionic and pH effects on the osmotic properties and structure of polyelectrolyte gels. J. Polym. Sci. Part B Polym. Phys. 2008, 46, 2803–2810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sircar, S.; Keener, J.P.; Fogelson, A.L. The effect of divalent vs. monovalent ions on the swelling of Mucin-like polyelectrolyte gels: Governing equations and equilibrium analysis. J. Chem. Phys. 2013, 138, 14901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horkay, F.; Hecht, A.; Grillo, I.; Basser, P.J.; Geissler, E. Experimental evidence for two thermodynamic length scales in neutralized polyacrylate gels. J. Chem. Phys. 2002, 117, 9103–9106. [Google Scholar] [CrossRef]
- Bloembergen, N.; Purcell, E.M.; Pound, R.V. Relaxation Effects in Nuclear Magnetic Resonance Absorption. Phys. Rev. 1948, 73, 679–712. [Google Scholar] [CrossRef]
- Tanner, J.E. Pulsed field gradients for NMR spin-echo diffusion measurements. Rev. Sci. Instruments 1965, 36, 1086–1087. [Google Scholar] [CrossRef]
- Spěváček, J. NMR Investigations of Phase Transition in Aqueous Polymer Solutions and Gels. Curr. Opin. Colloid Interface Sci. 2009, 14, 184–191. [Google Scholar] [CrossRef]
- Woessner, D.E.; Snowden, B.S. Pulsed Nmr Study of Water in Agar Gels. J. Colloid Interface Sci. 1970, 34, 290–299. [Google Scholar] [CrossRef]
- Mussel, M.; Wilczynski, E.; Eliav, U.; Gottesman, J.; Wilk, M.; Nevo, U. Dynamics of water and sodium in gels under salt-induced phase transition. J. Polym. Sci. Part B Polym. Phys. 2015, 53, 1620–1628. [Google Scholar] [CrossRef]
- Wallmersperger, T.; Kröplin, B.; Gülch, R.W. Coupled chemo-electro-mechanical formulation for ionic polymer gels—Numerical and experimental investigations. Mech. Mater. 2004, 36, 411–420. [Google Scholar] [CrossRef]
- Higa, M.; Tanioka, A.; Kira, A. A novel measurement method of Donnan potential at an interface between a charged membrane and mixed salt solution. J. Membr. Sci. 1998, 140, 213–220. [Google Scholar] [CrossRef]
- Guo, H.; Kurokawa, T.; Takahata, M.; Hong, W.; Katsuyama, Y.; Luo, F.; Ahmed, J.; Nakajima, T.; Nonoyama, T.; Gong, J.P. Quantitative observation of electric potential distribution of brittle polyelectrolyte hydrogels using microelectrode technique. Macromolecules 2016, 49, 3100–3108. [Google Scholar] [CrossRef]
- May, C.E.; Philipp, W.H. Ion Exchange Selectivity for Cross-Linked Polyacrylic Acid; NASA TM-83427; National Aeronautics and Space Administration: Washington, DC, USA, 1983.
- Estevez-Torres, A.; Baigl, D. DNA compaction: Fundamentals and applications. Soft Matter 2011, 7, 6746–6756. [Google Scholar] [CrossRef]
- Todd, B.A.; Rau, D.C. Interplay of ion binding and attraction in DNA condensed by multivalent cations. Nucleic Acids Res. 2008, 36, 501–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cranford, S.W.; Buehler, M.J. Variation of Weak Polyelectrolyte Persistence Length through an Electrostatic Contour Length. Macromolecules 2012, 45, 67–8082. [Google Scholar] [CrossRef]
- Horkay, F.; Basser, P.J.; Hecht, A.; Geissler, E. Effect of calcium/sodium ion exchange on the osmotic properties and structure of polyelectrolyte gels. Proc. Inst. Mech. Eng. Part J. Eng. Med. 2015, 229, 895–904. [Google Scholar] [CrossRef]
- Horkay, F. Effect of the Ionic Environment on the Supramolecular Structure and Thermodynamics of DNA Gels. Macromol. Symp. 2019, 385, 1800199. [Google Scholar] [CrossRef]
- Horkay, F.; Basser, P.J.; Hecht, A.; Geissler, E. Counterion and pH-Mediated Structural Changes in Charged Biopolymer Gels. Macromol. Symp. 2010, 291–292, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Larson, A.G.; Elnatan, D.; Keenen, M.M.; Trnka, M.J.; Johnston, J.B.; Burlingame, A.L.; Agard, D.A.; Redding, S.; Narlikar, G.J. Liquid droplet formation by HP1a suggests a role for phase separation in heterochromatin. Nature 2017, 547, 236–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boija, A.; Klein, I.A.; Sabari, B.R.; Dall’Agnese, A.; Coffey, E.L.; Zamudio, A.V.; Li, C.H.; Shrinivas, K.; Manteiga, J.C.; Hannett, N.M.; et al. Transcription factors activate genes through the phase-separation capacity of their activation domains. Cell 2018, 175, 1842–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdugo, P.; Aitken, M.; Langley, L.; Villalon, M.J. Molecular mechanism of product storage and release in mucin secretion. II. The role of extracellular Ca++. Biorheology 1987, 24, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Uvnaes, B.; Aborg, C. On the Cation Exchanger Properties of Rat Mast Cell Granules and their Storage of Histamine. Acta Physiol. Scand. 1977, 100, 309–314. [Google Scholar] [CrossRef]
- Marszalek, P.E.; Farrell, B.; Verdugo, P.; Fernandez, J.M. Kinetics of release of serotonin from isolated secretory granules II. Ion exchange determines, the diffusivity of serotonin. Biophys. J. 1997, 73, 1169–1183. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, T.J.; Borges, R.; Finnegan, J.M.; Pihel, K.; Amatore, C.; Wightman, R.M. Temporally resolved, independent stages of individual exocytotic secretion events. Biophys. J. 1996, 70, 1061–1068. [Google Scholar] [CrossRef] [Green Version]
- Olsen, I.D.; Luchtel, D.L.; Martin, A.W. Components of mucus of terrestrial slugs (Gastropoda). Am. J. Physiol. Regul. Integr. Comp. Physiol. 1983, 245, R448–R452. [Google Scholar] [CrossRef]
- Turkewitz, A.P. Out with a Bang! Tetrahymena as a Model System to Study Secretory Granule Biogenesis. Traffic 2004, 5, 63–68. [Google Scholar] [CrossRef]
- Verdugo, P. Supramolecular dynamics of mucus. Cold Spring Harb. Perspect. Med. 2012, 2, a009597. [Google Scholar] [CrossRef] [Green Version]
- Uvnas, B.; Aborg, C.-H. Role of Ion Exchange in Release of Biogenic Amines. Physiology 1989, 4, 68–71. [Google Scholar] [CrossRef]
- Barasch, J.; Kiss, B.; Prince, A.; Saiman, L.; Gruenert, D.; Ai-Awqati, Q. Defective acidification of intracellular organelles in cystic fibrosis. Nature 1991, 352, 70–73. [Google Scholar] [CrossRef]
- Sheppard, M.N.; Nicholson, A.G. The pathology of cystic fibrosis. Curr. Diagn. Pathol. 2002, 8, 50–59. [Google Scholar] [CrossRef]
- Boyce, C.K.; Zwieniecki, M.A.; Cody, G.D.; Jacobsen, C.; Wirick, S.; Knoll, A.H.; Holbrook, N.M. Evolution of xylem lignification and hydrogel transport regulation. Proc. Natl. Acad. Sci. USA 2004, 101, 17555–17558. [Google Scholar] [CrossRef] [Green Version]
- Nardini, A.; Salleo, S.; Jansen, S. More than just a vulnerable pipeline: Xylem physiology in the light of ion-mediated regulation of plant water transport. J. Exp. Bot. 2011, 62, 4701–4718. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.H. Hydraulic architecture of some diffuse-porous trees. Can. J. Bot. 1978, 56, 2286–2295. [Google Scholar] [CrossRef] [Green Version]
- Ryden, P.; MacDougall, A.J.; Tibbits, C.W.; Ring, S.G. Hydration of pectic polysaccharides. Biopolymers 2000, 54, 398–405. [Google Scholar] [CrossRef]
- Yoshida, T.; Nin, F.; Murakami, S.; Ogata, G.; Uetsuka, S.; Choi, S.; Nakagawa, T.; Inohara, H.; Komune, S.; Kurachi, Y.; et al. The unique ion permeability profile of cochlear fibrocytes and its contribution to establishing their positive resting membrane potential. Pflugers Arch. Eur. J. Physiol. 2016, 468, 1609–1619. [Google Scholar] [CrossRef] [PubMed]
- Beilby, M.J. Action potential in charophytes. Int. Rev. Cytol. 2007, 257, 43–82. [Google Scholar] [PubMed]
- Sundelacruz, S.; Levin, M.; Kaplan, D.L. Role of membrane potential in the regulation of cell proliferation and differentiation. Stem Cell Rev. Rep. 2009, 5, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Curtis, H.J.; Cole, K.S. Membrane resting and action potentials from the squid giant axon. J. Cell. Comp. Physiol. 1942, 19, 135–144. [Google Scholar] [CrossRef]
- Hill, S.E.; Osterhout, W.J.V. Calculations of bioelectric potentials II. The Concentration Potential of KCl in Nitella. J. Gen. Physiol. 1938, 21, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Tasaki, I. Demonstration of two stable states of the nerve membrane in potassium-rich media. J. Physiol. 1959, 148, 306–331. [Google Scholar] [CrossRef] [Green Version]
- Tasaki, I.; Takenaka, T.; Yamagishi, S. Abrupt Depolarization and Bi-Ionic Action Potentials in Internally Perfused Squid Giant Axons. Am. J. Physiol. 1968, 215, 152–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tazawa, M.; Shimmen, T. Cell motility and ionic relations in characean cells as revealed by internal perfusion and cell models. Int. Rev. Cytol. 1987, 109, 259–312. [Google Scholar]
- Tasaki, I.; Byrne, P.M. Discontinuous Volume Transitions in Ionic Gels and Their Possible Involvement in the Nerve Excitation Process. Biopolymers 1992, 32, 1019–1023. [Google Scholar] [CrossRef] [PubMed]
- Wnek, G.E. Perspective: Do macromolecules play a role in the mechanisms of nerve stimulation and nervous transmission? J. Polym. Sci. Part B Polym. Phys. 2016, 54, 7–14. [Google Scholar] [CrossRef]
- Kozawa, S.K.; Wnek, G.E. Macromolecules of the cell: A polymer science viewpoint. Polym. Int. 2020. [Google Scholar] [CrossRef]
- Metuzals, J.; Tasaki, I. Subaxolemmal filamentous network in the giant nerve fiber of the squid (Loligo Peali L.) and its possible role in excitability. J. Cell Biol. 1978, 78, 597–621. [Google Scholar] [CrossRef]
- Tasaki, I.; Takenaka, T. Effects of various potassium salts and proteases upon excitability of intracellularly perfused squid giant axons. Proc. Natl. Acad. Sci. USA 1964, 52, 804. [Google Scholar] [CrossRef] [Green Version]
- Brangwynne, C.P.; Eckmann, C.R.; Courson, D.S.; Rybarska, A.; Hoege, C.; Gharakhani, J.; Jülicher, F.; Hyman, A.A. Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science 2009, 324, 1729–1732. [Google Scholar] [CrossRef]
- Falahati, H.; Haji-Akbari, A. Thermodynamically driven assemblies and liquid-liquid phase separations in biology. Soft Matter 2019, 15, 1135–1154. [Google Scholar] [CrossRef] [Green Version]
- Clifford, P. Brangwynne, Peter Tompa, and Rohit V. Pappu. Polymer physics of intracellular phase transitions. Nat. Phys. 2015, 11, 899–904. [Google Scholar]
- Schmidt, H.B.; Goerlich, D. Transport selectivity of nuclear pores, phase separation, and membraneless organelles. Trends Biochem. Sci. 2016, 41, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Beltran, E.; Moschou, P.N.; Smertenko, A.P.; Bozhkov, P.V. Tudor staphylococcal nuclease links formation of stress granules and processing bodies with mRNA catabolism in arabidopsis. Plant Cell 2015, 27, 926–943. [Google Scholar] [CrossRef] [Green Version]
- Schuetz, S.; Nöldeke, E.R.; Sprangers, R. A synergistic network of interactions promotes the formation of in vitro processing bodies and protects mRNA against decapping. Nucleic Acids Res. 2017, 45, 6911–6922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, J.; Weber, S.C.; Vaidya, N.; Haataja, M.; Brangwynne, C.P.; Weitz, D.A. RNA transcription modulates phase transition-driven nuclear body assembly. Proc. Natl. Acad. Sci. USA 2015, 112, E5237–E5245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feric, M.; Vaidya, N.; Harmon, T.S.; Mitrea, D.M.; Zhu, L.; Richardson, T.M.; Kriwacki, R.W.; Pappu, R.V.; Brangwynne, C.P. Coexisting Liquid Phases Underlie Nucleolar Subcompartments. Cell 2016, 165, 1686–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gall, J.G. Cajal Bodies: The First 100 Years. Annu. Rev. Cell Dev. Biol. 2000, 16, 273–300. [Google Scholar] [CrossRef]
- Hennig, S.; Kong, G.; Mannen, T.; Sadowska, A.; Kobelke, S.; Blythe, A.; Knott, G.J.; Iyer, S.S.; Ho, D.; Newcombe, E.A.; et al. Prion-like domains in RNA binding proteins are essential for building subnuclear paraspeckles. J. Cell Biol. 2015, 210, 529–539. [Google Scholar] [CrossRef]
- Langdon, E.M.; Qiu, Y.; Niaki, A.G.; McLaughlin, G.A.; Weidmann, C.A.; Gerbich, T.M.; Smith, J.A.; Crutchley, J.M.; Termini, C.M.; Weeks, K.M.; et al. mRNA structure determines specificity of a polyQ-driven phase separation. Science 2018, 360, 922–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, P.; Kedersha, N. RNA granules. J. Cell Biol. 2006, 172, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, S. Regulation of protein synthesis during heat shock. Nature 1981, 293, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Nott, T.J.; Petsalaki, E.; Farber, P.; Jervis, D.; Fussner, E.; Plochowietz, A.; Craggs, T.D.; Bazett-Jones, D.P.; Pawson, T.; Forman-Kay, J.D.; et al. Phase transition of a disordered nuage protein generates environmentally responsive membraneless organelles. Mol. Cell 2015, 57, 936–947. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Banjade, S.; Cheng, H.; Kim, S.; Chen, B.; Guo, L.; Llaguno, M.; Hollingsworth, J.V.; King, D.S.; Banani, S.F.; et al. Phase transitions in the assembly of multivalent signalling proteins. Nature 2012, 483, 336–340. [Google Scholar] [CrossRef]
- Falahati, H.; Wieschaus, E. Independent active and thermodynamic processes govern the nucleolus assembly in vivo. Proc. Natl. Acad. Sci. USA 2017, 114, 1335–1340. [Google Scholar] [CrossRef] [Green Version]
- Walter, H.; Brooks, D.E. Phase separation in cytoplasm, due to macromolecular crowding, is the basis for microcompartmentation. FEBS Lett. 1995, 361, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-M.; Holehouse, A.S.; Pappu, R.V. Physical Principles Underlying the Complex Biology of Intracellular Phase Transitions. Annu. Rev. Biophys. 2020, 49, 107–133. [Google Scholar] [CrossRef] [Green Version]
- Nandi, S.K.; Heidenreich, M.; Levy, E.D.; Safran, S.A. Interacting multivalent molecules: Affinity and valence impact the extent and symmetry of phase separation. arXiv 2019, arXiv:1910.11193. [Google Scholar]
- Flory, P.J. Principles of Polymer Chemistry; Cornell University Press: Ithaca, NY, USA, 1953. [Google Scholar]
- De Gennes, P.-G. Scaling Concepts in Polymer Physics; Cornell University Press: Ithaca, NY, USA, 1979. [Google Scholar]
- Hua, J.; Mitra, M.K.; Muthukumar, M. Theory of Volume Transition in Polyelectrolyte Gels with Charge Regularization. J. Chem. Phys. 2012, 136, 134901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrynin, A.V.; Rubinstein, M. Theory of Polyelectrolytes in Solutions and at Surfaces. Prog. Polym. Sci. 2005, 30, 1049–1118. [Google Scholar] [CrossRef]
- Dyson, F. A Meeting with Enrico Fermi. Nature 2004, 427, 297. [Google Scholar] [CrossRef]
- Ou, X.; Han, Q.; Dai, H.; Wang, J. Molecular dynamic simulations of the water absorbency of hydrogels. J. Mol. Model. 2015, 21, 231. [Google Scholar] [CrossRef]
- Galindo-Murillo, R.; Robertson, J.C.; Zgarbova, M.; Sponer, J.; Otyepka, M.; Jurecka, P.; Cheatham, T.E., III. Assessing the current state of amber force field modifications for DNA. J. Chem. Theory Comput. 2016, 12, 4114–4127. [Google Scholar] [CrossRef]
- Kmiecik, S.; Gront, D.; Kolinski, M.; Wieteska, L.; Dawid, A.E.; Kolinski, A. Coarse-grained protein models and their applications. Chem. Rev. 2016, 116, 7898–7936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alegre-Requena, J.V.; Saldias, C.; Inostroza-Rivera, R.; Diaz, D.D. Understanding hydrogelation processes through molecular dynamics. J. Mater. Chem. B 2019, 7, 1652–1673. [Google Scholar] [CrossRef]
- Dai, L.; Mu, Y.; Nordenskiöld, L.; van der Maarel, J.R.C. Molecular dynamics simulation of multivalent-ion mediated attraction between DNA molecules. Phys. Rev. Lett. 2008, 100, 118301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Shi, X. Molecular dynamics simulation of diffusion of nanoparticles in mucus. Acta Mech. Solida Sin. 2017, 30, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Assifaoui, A.; Lerbret, A.; Uyen, H.T.D.; Neiers, F.; Chambin, O.; Loupiac, C.; Cousin, F. Structural behaviour differences in low methoxy pectin solutions in the presence of divalent cations (Ca2+ and Zn2+): A process driven by the binding mechanism of the cation with the galacturonate unit. Soft Matter 2015, 11, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Landau, L.D.; Lifshitz, E.M. Volume 6 of Course of Theoretical Physics. In Fluid Mechanics; Sykes, J.B., Reid, W.H., Eds.; Pergamon Press: Oxford, UK, 1987. [Google Scholar]
- Hariharan, D.; Peppas, N.A. Modelling of water transport in ionic hydrophilic polymers. J. Polym. Sci. Part B Polym. Phys. 1994, 32, 1093–1103. [Google Scholar] [CrossRef]
- Segalman, D.J.; Witkowski, W.R. Two-dimensional finite element analysis of a polymer gel drug delivery system. Mater. Sci. Eng. C 1995, 2, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Achilleos, E.C.; Prud’Homme, R.K.; Kevrekidis, I.G.; Christodoulou, K.N.; Gee, K.R. Quantifying deformation in gel swelling: Experiments and simulations. AIChE J. 2000, 46, 2128–2139. [Google Scholar] [CrossRef]
- Achilleos, E.C.; Christodoulou, K.N.; Kevrekidis, I.G. A transport model for swelling of polyelectrolyte gels in simple and complex geometries. Comput. Theor. Polym. Sci. 2001, 11, 63–80. [Google Scholar] [CrossRef]
- Feng, L.; Jia, Y.; Chen, X.; Li, X.; An, L. A multiphasic model for the volume change of polyelectrolyte hydrogels. J. Chem. Phys. 2010, 133, 114904. [Google Scholar] [CrossRef]
- Mori, Y.; Chen, H.; Micek, C.; Calderer, M. A dynamic model of polyelectrolyte gels. SIAM J. Appl. Math. 2013, 73, 104–133. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Calderer, M.; Mori, Y. Analysis and simulation of a model of polyelectrolyte gel in one spatial dimension. Nonlinearity 2014, 27, 1241. [Google Scholar] [CrossRef] [Green Version]
- Drozdov, A.D.; Christiansen, J.D. Modeling the effects of pH and ionic strength on swelling of polyelectrolyte gels. J. Chem. Phys. 2015, 142, 114904. [Google Scholar] [CrossRef]
- Lewis, O.L.; Keener, J.P.; Fogelson, A.L. Electrodiffusion-mediated swelling of a two-phase gel model of gastric mucus. Gels 2018, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Lewis, O.L.; Keener, J.P.; Fogelson, A.L. A physics-based model for maintenance of the pH gradient in the gastric mucus layer. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G599–G612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruinsma, R.; Grosberg, A.Y.; Rabin, Y.; Zidovska, A. Chromatin hydrodynamics. Biophys. J. 2014, 106, 1871–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, W.M.; Hou, J.S.; Mow, V.C. A triphasic theory for the swelling and deformation behaviors of articular cartilage. J. Biomech. Eng. 1991, 113, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Wolgemuth, C.W.; Mogilner, A.; Oster, G. The hydration dynamics of polyelectrolyte gels with applications to cell motility and drug delivery. Eur. Biophys. J. 2004, 33, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Cogan, N.G.; Keener, J.P. The role of the biofilm matrix in structural development. Math. Med. Biol. 2004, 21, 147–166. [Google Scholar] [CrossRef]
- Herant, M.; Marganski, W.A.; Dembo, M. The mechanics of neutrophils: Synthetic modeling of three experiments. Biophys. J. 2003, 84, 3389–3413. [Google Scholar] [CrossRef] [Green Version]
- Herant, M.; Heinrich, V.; Dembo, M. Mechanics of neutrophil phagocytosis: Experiments and quantitative models. J. Cell Sci. 2006, 119, 1903–1913. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.P.; King, J.R. Mathematical modelling of avascular-tumour growth. Math. Med. Biol. 1997, 14, 39–69. [Google Scholar] [CrossRef]
- Breward, C.J.W.; Byrne, H.M.; Lewis, C.E. The role of cell-cell interactions in a two-phase model for avascular tumour growth. J. Math. Biol. 2002, 45, 125–152. [Google Scholar] [CrossRef]
- Mao, S.; Kuldinow, D.; Haataja, M.P.; Košmrlj, A. Phase behavior and morphology of multicomponent liquid mixtures. Soft Matter 2019, 15, 1297–1311. [Google Scholar] [CrossRef] [Green Version]
- Gasior, K.; Forest, M.G.; Gladfelter, A.S.; Newby, J.M. Modeling the Mechanisms by Which Coexisting Biomolecular RNA–Protein Condensates Form. Bull. Math. Biol. 2020, 82, 1–16. [Google Scholar] [CrossRef]
- Calvert, P. Hydrogels for soft machines. Adv. Mater. 2009, 21, 743–756. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mussel, M.; Basser, P.J.; Horkay, F. Ion-Induced Volume Transition in Gels and Its Role in Biology. Gels 2021, 7, 20. https://doi.org/10.3390/gels7010020
Mussel M, Basser PJ, Horkay F. Ion-Induced Volume Transition in Gels and Its Role in Biology. Gels. 2021; 7(1):20. https://doi.org/10.3390/gels7010020
Chicago/Turabian StyleMussel, Matan, Peter J. Basser, and Ferenc Horkay. 2021. "Ion-Induced Volume Transition in Gels and Its Role in Biology" Gels 7, no. 1: 20. https://doi.org/10.3390/gels7010020
APA StyleMussel, M., Basser, P. J., & Horkay, F. (2021). Ion-Induced Volume Transition in Gels and Its Role in Biology. Gels, 7(1), 20. https://doi.org/10.3390/gels7010020