Here,
is the
y-coordinate at the flagellum base (the position at which the cell and flagellum connect),
is the characteristic length scale of the amplitude modulation,
denotes the wave number, and
the angular frequency. Lastly,
gives the lateral displacement of the flagellum along the
x-axis with respect to the longitudinal position from the flagellum base and a specific instant in time,
t, within a beat cycle. The characteristic parameters in Equation (
1) are given for the model organism
D. grandis in
Table 1.
2.1.1. Placement of Fixed Prey
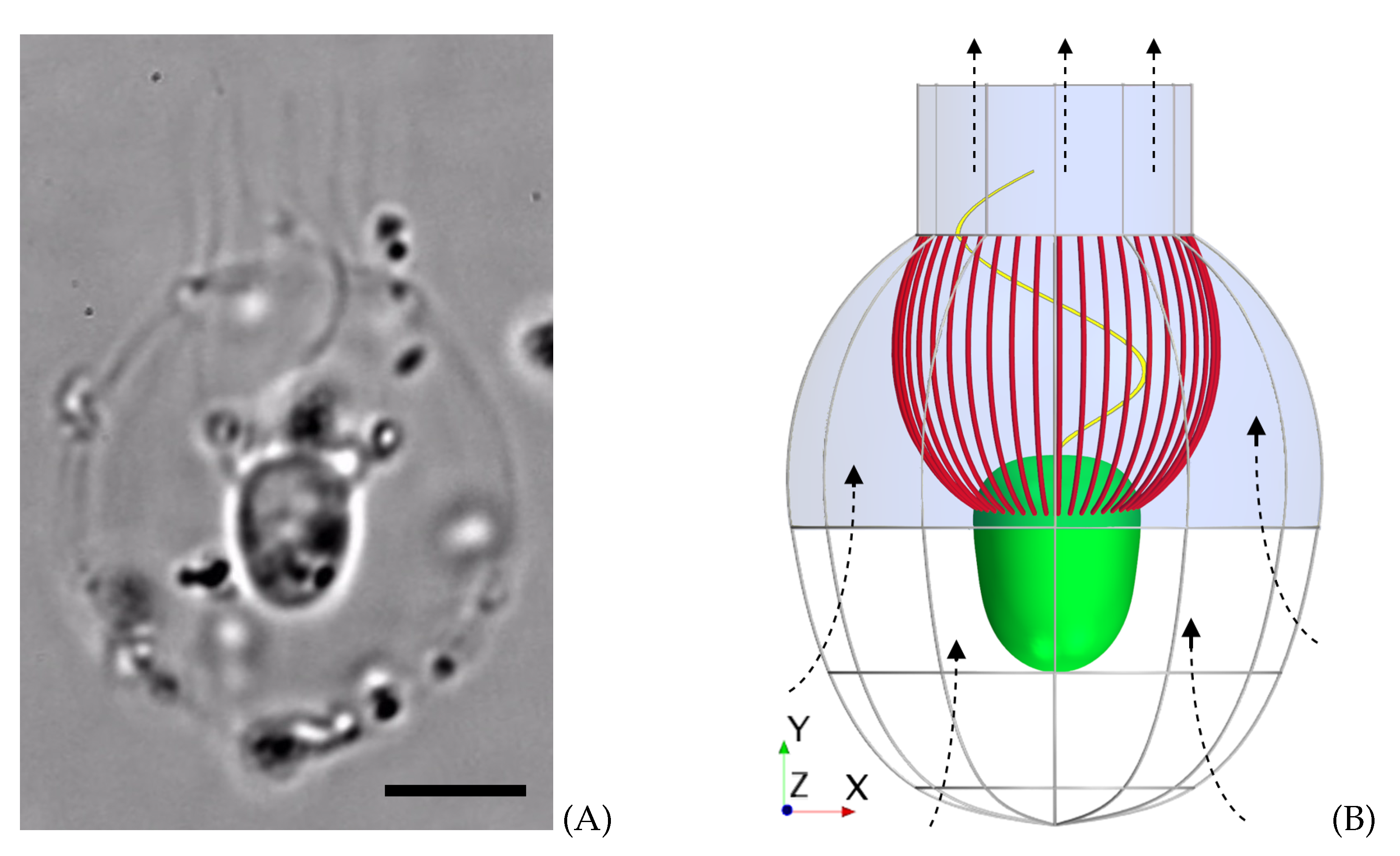
When constructing the model morphology of
D. grandis, the assumption of symmetry around the longitudinal center axis is utilized in shaping the lorica, filter, and cell. This allows for each part’s surface to be constructed as a simple curve, which can readily be revolved around the center axis of symmetry (
Figure 2A). The equations for the curves of revolutions were obtained by averaging the outline of the mid-plane view of six different instances of the species [
5,
9]. The first resultant curvature equation models both the lorica dome,
, and the cell,
, and the second curvature equation models the collar filter,
. Both curvature equations are given, respectively, as:
where
,
,
, and
are morphological parameters unique to the lorica and cell outline (
Supplementary Information S8.1, Table S1), and
and
are the polar angles at which the filter meets the cell and lorica, respectively. The polar angle
is defined relative to the longitudinal center axis, as seen in
Figure 2A.
Forty-eight microvilli tentacles are distributed equally along the collar surface. The lorica dome and chimney are modeled as an impermeable baffle, where the ribs in the inferior part of the lorica (pictured in
Figure 1B) are neglected for simplicity [
9]. The flagellar vane is modeled as a
wide sheet, which corresponds to the vane being slightly smaller than the diameter of the collar at the collar base. The traveling waveform of the beat of the flagellar vane is given in Equation (
1), along with additional morphological and kinematic parameters for
D. grandis (
Table 1). The resulting CFD model morphology is pictured in
Figure 2C. All surfaces (i.e., surfaces of the filter, the cell, the flagellum, and the lorica) are subject to the no-slip boundary condition.
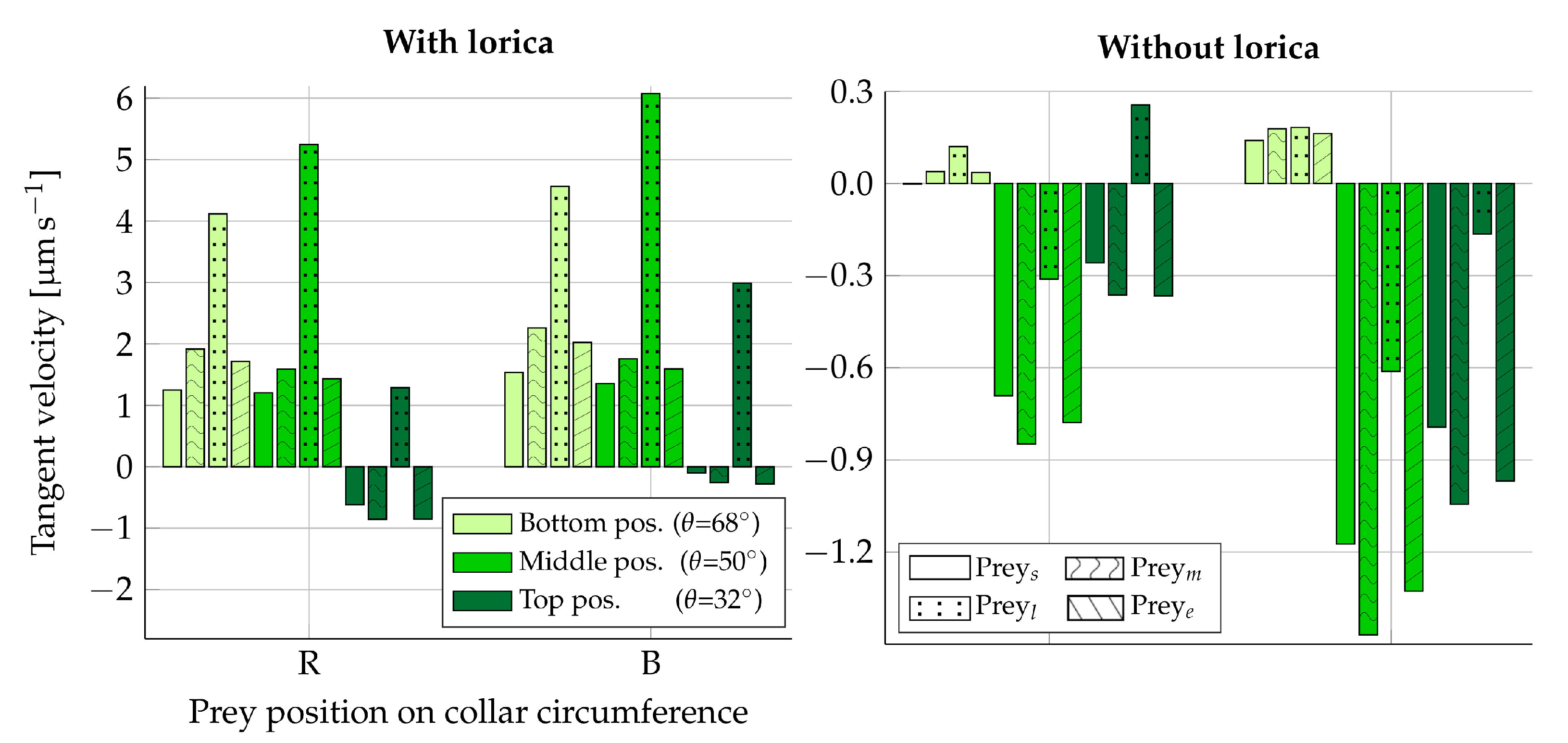
The analysis will consider four different prey types: three spherical preys and one elongated prey (
Table 2). The medium-sized prey,
, will serve as the reference prey. The surface forces on each prey are recorded with respect to its local coordinate system. The local coordinate system is placed such that the
x-axis is parallel to the normal vector at the point of prey–collar contact, and equivalently, the
y-axis is parallel to the tangent vector. Using Equation (
3), the gradients of the tangent,
, and the normal vector,
, are given by:
Furthermore, the local
x-axis is directed towards the collar, and the local
y-axis is directed away from the cell, as seen in
Figure 2B.
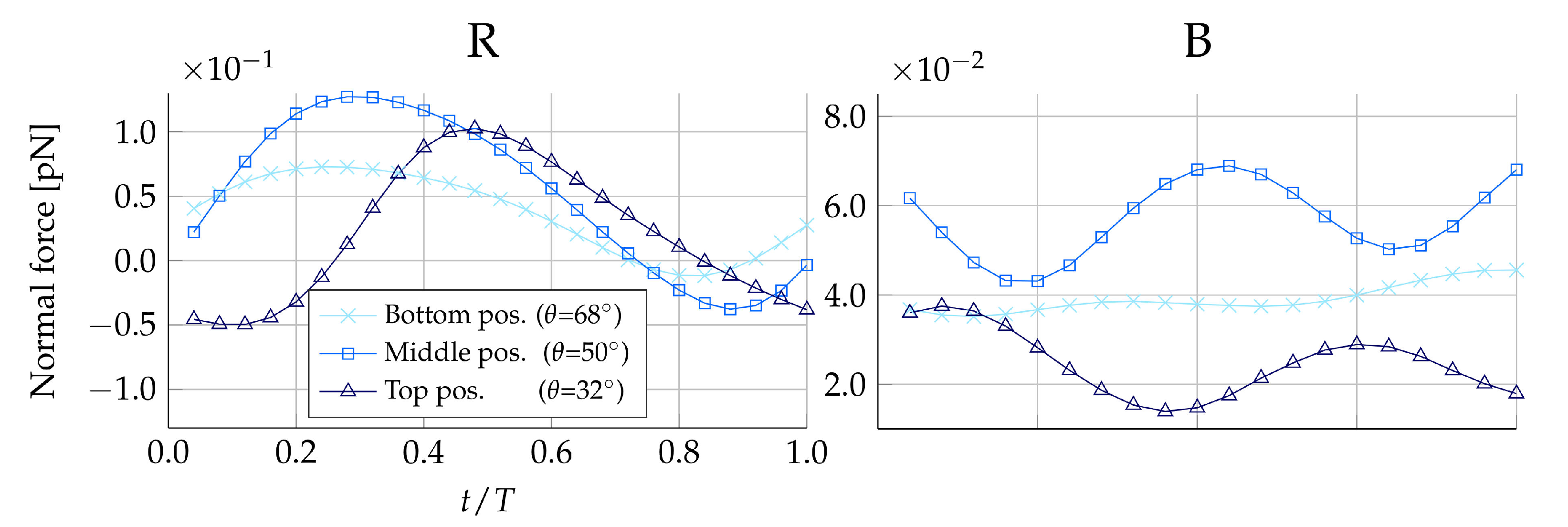
The prey will be placed in two vertical perpendicular planes, each of which contains the longitudinal symmetry axis. In the first plane, called the beat plane (
-plane in
Figure 1), the prey will be placed on the right-hand side and left-hand side of the collar, denoted
R and
L, respectively. In the second plane, called the vane plane (
-plane in
Figure 1), the prey will similarly be placed on the back and front of the collar with respect to the view pictured in
Figure 1. These two positions are denoted
B and
F, respectively. In each of the four positions,
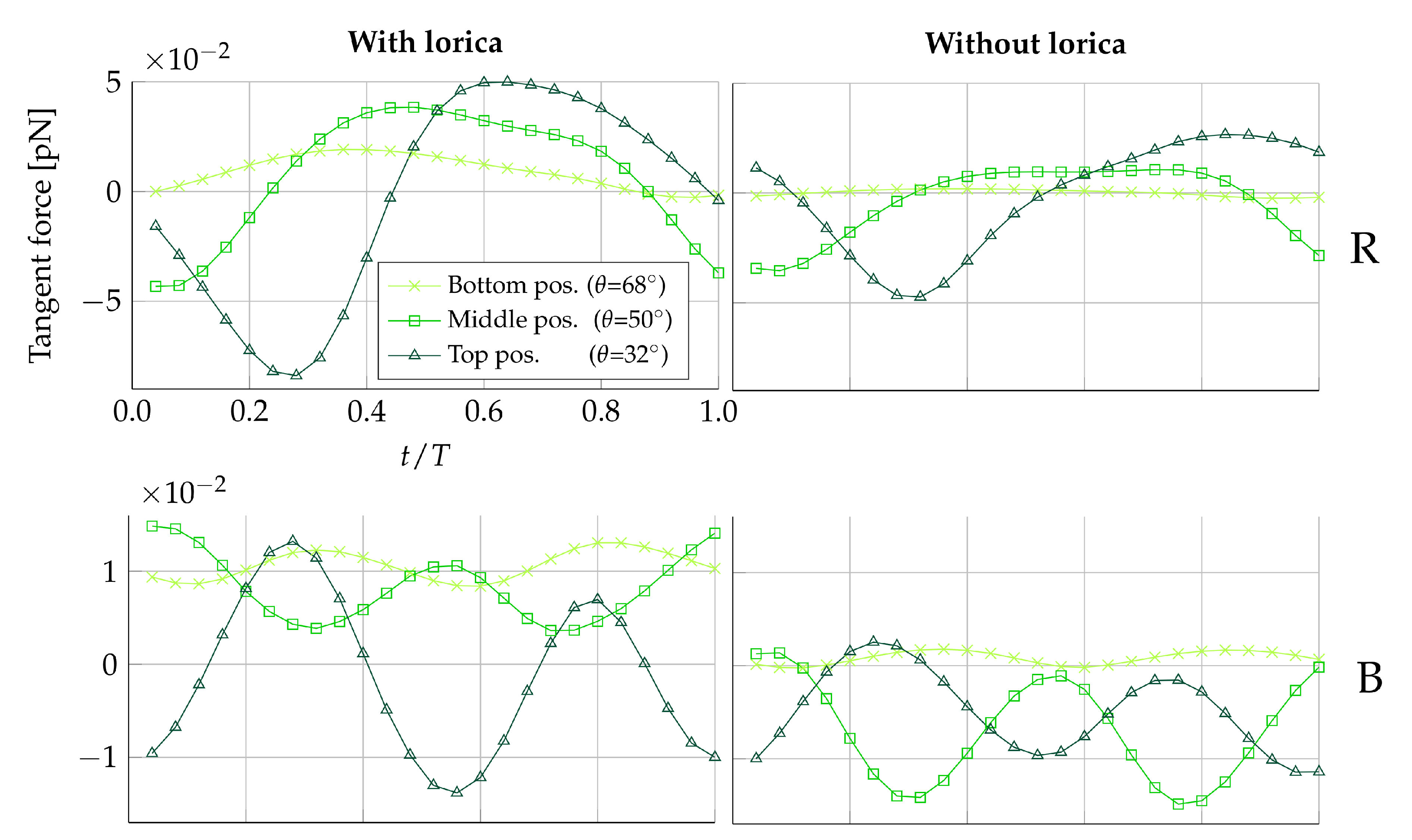
and
the vertical position on the collar will be varied in three ways: at the bottom (
), the middle (
), and the top (
), summing to a total of 12 positions. As an example, a medium-sized prey placed at the middle and on the left-hand side of the collar is referred to as:
.
2.1.2. Governing Equations and Hydrodynamic Effects
For the prey particles captured by
D. grandis, the Reynolds number (
) is in the order of
. Here,
is the fluid density,
is the fluid dynamic viscosity,
D is the characteristic length scale given by the diameter of the prey, and
is the characteristic velocity of the flagellum, where
and
f denote the wavelength and frequency of the beating flagellum (cf.
Supplementary Information S8.1). We note that a flow at this Reynolds number is governed by the time-independent Stokes equations:
where
u and
p denote the flow velocity and pressure, respectively. The time independency allows for the flow to be resolved in a series of discrete time steps, as the response of the fluid to the motion of boundaries can be considered instantaneous [
13]. One flagellum beat cycle is discretized into 25 time steps. Doubling the number of time steps introduces a variation of less than 1% in the results [
5]. Although it suffices to solve the Stokes equations, the full Navier–Stokes equations are solved at each discrete point in time by the CFD software. In combination with using mesh-morphing techniques, this allows for the flagellum position to be updated in each time step in a computationally efficient way. Given the symmetry of prey placed in the same plane but on opposing sides of the collar, it is only necessary to model half of a flagellum beat cycle. However, to ensure that mesh morphing does not have a significant impact on the symmetry at hand, we still choose to simulate the entire beat cycle.
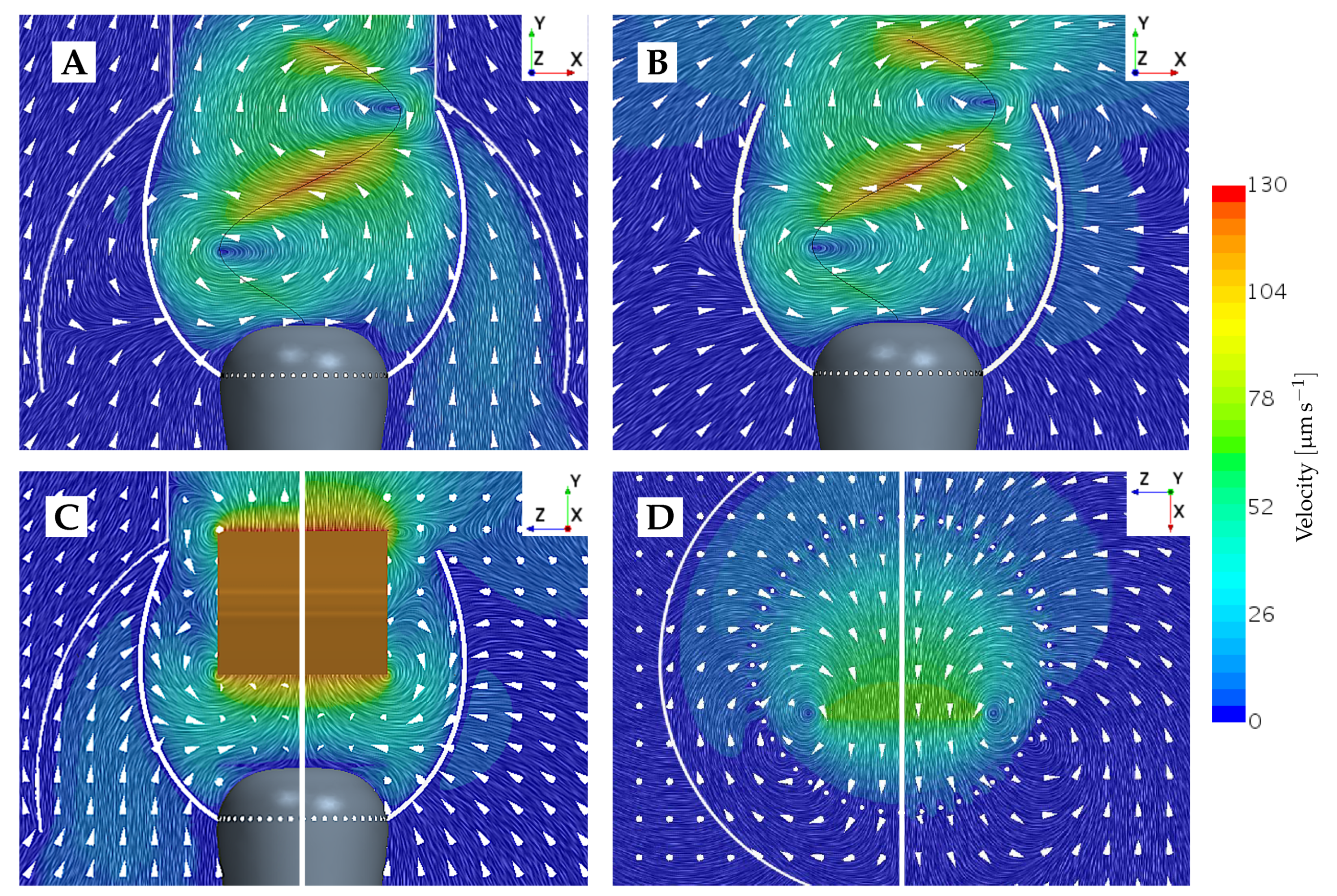
An overset mesh using linear interpolation between the overset and background meshes is added around all prey particles, which allows for the same mesh to be used in both the dynamic and static simulations. The drawback of using an overset mesh in the static setting is that it is not possible for the body within the overset to be in direct contact with another surface. Subsequently, we introduce a gap between the prey and the microvilli. The combination of introducing a gap of size ∼5% of the prey radius and opting for the overset mesh method has been shown to result in an accumulated variation of up to ∼5% in the calculated prey forces as compared to a mesh without overset. Similarly to the mesh-morphing techniques used for updating the flagellum position, an overset mesh greatly reduces the solver workload in the dynamic prey simulations, as the regeneration of the entire volume mesh in each time step becomes dispensable.
The computational domain is discretized using from 6 to 11 million computational cells, where the highest density of cells is found along the microvilli tentacles and prey surfaces. This cell density is kept approximately constant for all analyses. The computational cost for a full simulation containing 11 million cells is approximately 50 h when performed on seven 16-core nodes (Xeon E5-2650 running at 2.60 GHz). The extent of the prey overset mesh is increased when the prey size is increased; hence, to obtain a sufficient connectivity between the two grids, the extent of refined cells in the background mesh has to increase accordingly. The mesh is pictured in
Supplementary Information S8.3, Figure S3. In the loricate case using
, the discretization error was shown to cause a variation of less than 1% in the measured prey forces and the time-averaged clearance rate through the filter (
Supplementary S8.3). Here, a very fine mesh acted as an exact solution; thus, the discretization error was quantified based on a comparison with this solution. The geometry is contained within a spherical computational domain with a radius of
, where a pressure boundary condition (B.C.) is applied to the outer surface (note the drawbacks of the pressure B.C. in
Supplementary Information S8.2). Increasing the computational domain radius to
results in a variation of less than
in the results [
5].
The hydrodynamic forces,
F, and torque,
L, acting on the fixed prey are obtained by integrating the stress tensor,
, over the prey surface [
13],
S:
where the stress tensor is given by
(
is the identity tensor),
denotes the normal vector directed outwards from the prey surface, and
denotes the position vector on the prey surface. Hence, prior to constructing the stress tensor, the pressure,
p, and velocity,
, of the incompressible fluid must be computed.
For a fixed sphere of radius
r held in a uniform stream of velocity
U, the Stokes law states the drag force on a sphere [
14]:
The total drag force is made up of two force contributions: pressure and viscous shear stress. Here, viscous forces account for two-thirds of the total drag force, and pressure forces account for the remaining one-third [
15]. If the ratio of pressure to shear stress forces on the spherical prey surface is found to be approximately 1:2, then this could suggest that it may be reasonable to use the Stokes law to give a first-order estimate of the order of the prey velocity. This can be done by rearranging Equation (
10) and using the magnitude of the simulated prey force as an input. Similarly to Equation (
10), the drag on a prolate spheroid placed in a flow normal to its axis of revolution takes the approximate Stokes form [
16,
17,
18]:
Here,
a is the major and
b is the minor radius of the spheroid, and
is the specific drag coefficient for the described case. The limitation of Equations (
10) and (
11) is that they do not take into account the non-uniform velocity field in the vicinity of the filter or the influence of the finite size of the prey and its boundary condition—as it is either non-slip for the fixed prey or globally force and torque free for the freely moving prey. Hence, when using the Stokes law, the velocity estimate for the prey may be an overestimation. Evaluating the shape of the curvature of the velocity profile near the filter would allow for a second-order estimate of the prey velocity using Faxén relations [
14]. However, as we simply wish to evaluate the order of the prey velocity, the first-order Stokes estimate is considered sufficient. Consequently, we choose to use Stokes (Equations (
10) and (
11)) to approximate the prey velocity from the forces on the fixed prey.
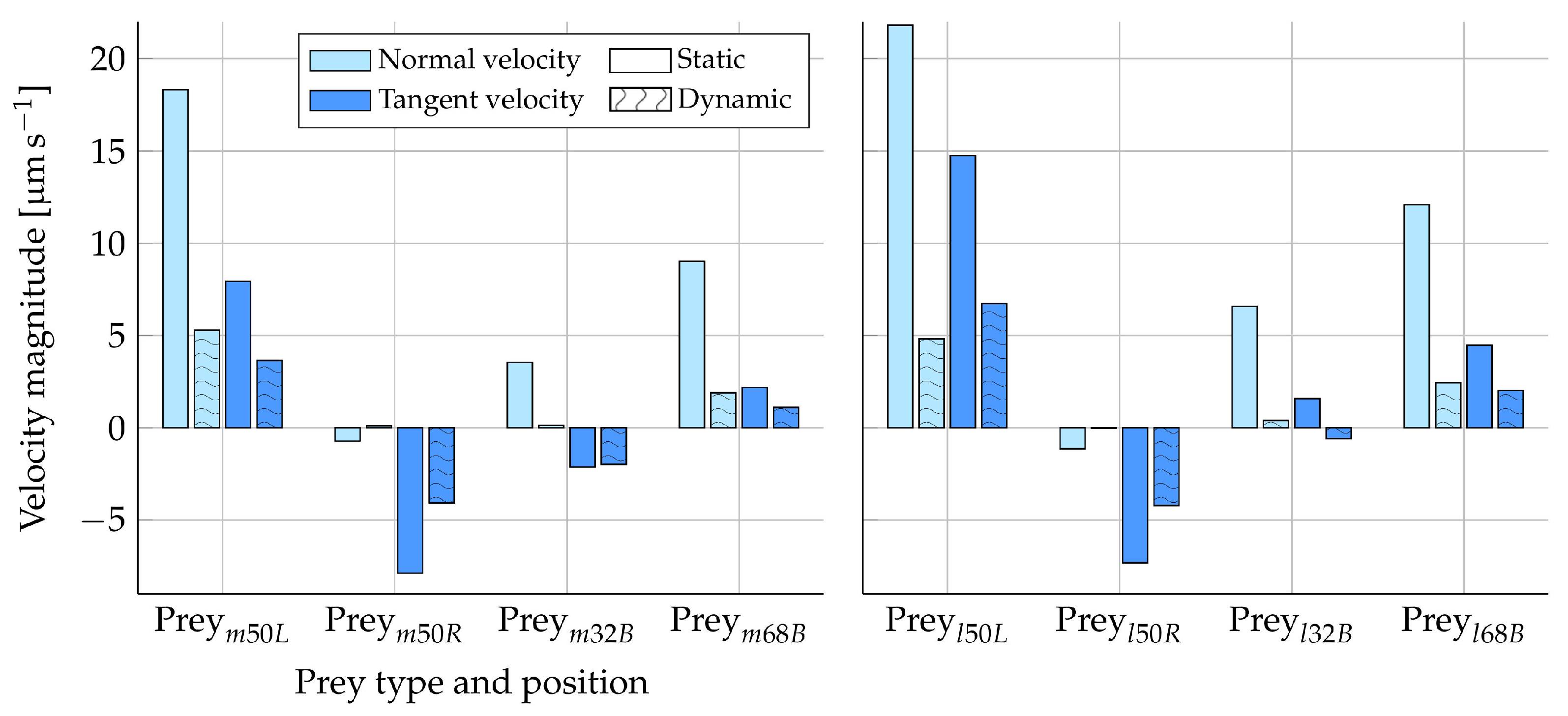
2.1.3. Solution Procedure to Model Drifting Prey
We define the prey velocity and rotation rate as
and
, respectively. The hydrodynamic forces,
, and torques,
, are evaluated as stated in Equations (
8) and (
9). The Stokes number, the ratio of the response time of a particle to the response time of the fluid in which it is submerged, is, in the case of a Stokes flow, given by:
. Here, employing the density of the prey
(where we have assumed the prey density to be equal to that of the fluid), the fluid velocity
, and the radius of the prey
r. For a Stokes number that is much smaller than unity, the prey will adhere nearly perfectly to the motion of the streamlines [
19]. Moreover, relative to the motion of the rest of the domain, the prey will catch up with the flow instantaneously in every discrete time step. Simulating that the prey catches up with the flow corresponds to assigning the prey a velocity and a rotation rate such that, consequently, its surface becomes force and torque free in each time step. Applying the same logic, we thus refrain from the approach of simply solving Newton’s
law of motion, since the concept of acceleration becomes elusive at such low Stokes numbers. Instead, we opt for an iterative method to ensure a force- and torque-free particle. Rather than focusing on dynamics, this method iterates to determine a rigid body motion such that the condition of a force- and torque-free surface is met. Here, we assume deformations of the prey to be negligible for simplicity.
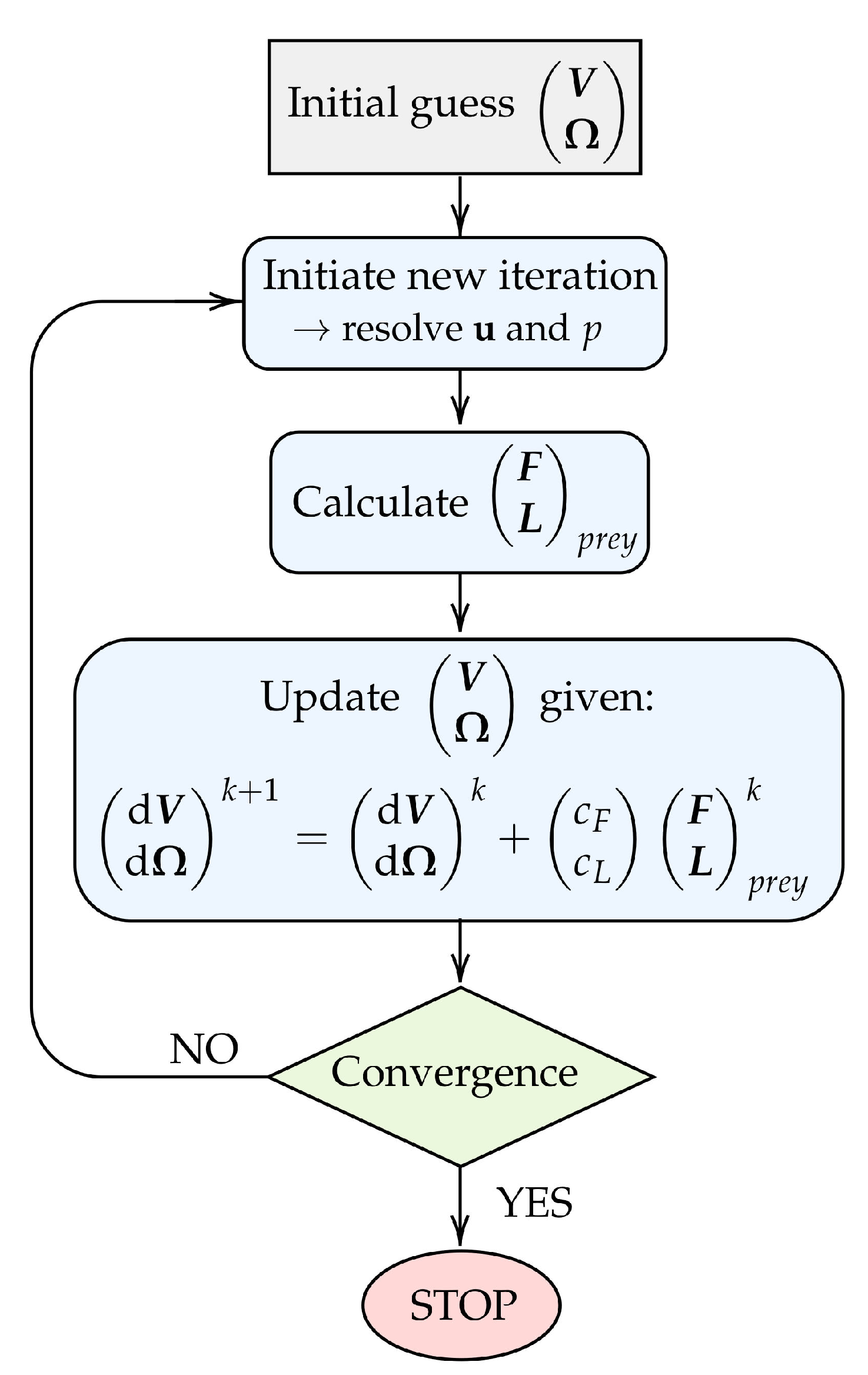
The procedure for obtaining the prey velocity and rotation rate in each discrete time step is illustrated in
Figure 3. Firstly, an arbitrary initial guess is assigned to the unknown
and
. This allows for the flow velocity and pressure fields to be computed using the SIMPLE algorithm [
20]. Subsequently, the forces and torques acting on the prey surface can be deduced. Given the prey forces and torques, we update the prey velocity and rotation rate using the relation:
where
k denotes an iteration counter and
and
are relaxation constants for the translational velocity and the rotation rate, respectively. The constants are arbitrarily chosen such that
and
are much smaller than
and
, respectively. The magnitude of
and
is found only to influence the rate of convergence, and we use
throughout.
The updated prey velocity and rotation rate serve as the input for a new iteration, in which the above procedure is repeated. The iteration process continues until the forces and torques on the prey are converged to a negligible level. At this point, the prey position is updated based on the converged prey velocity and rotation rate. The entire procedure is repeated for the subsequent time step.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}