
Targeting MicroRNAs with Small Molecules



Abstract
:1. Introduction
2. Identification of Small Molecules Interacting with miRs
2.1. Bioinformatics Tools
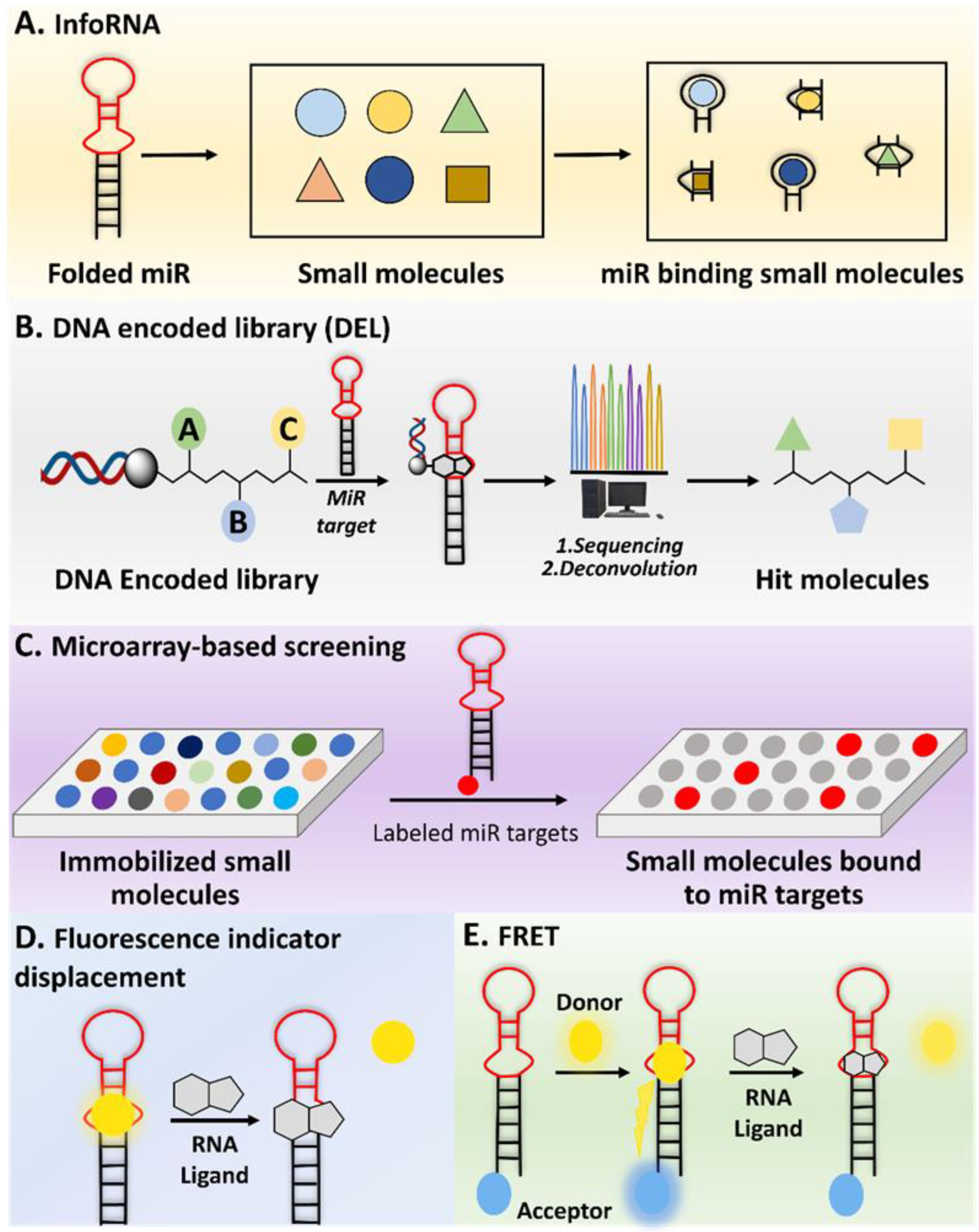
2.1.1. InfoRNA
2.1.2. Docking
2.1.3. AI
2.2. DNA Encoded Library (DEL)
2.3. Microarray Screening
2.4. Fluorescence Reporter
2.4.1. Fluorescence Indicator Displacement (FID)
2.4.2. Fluorescence Resonance Energy Transfer (FRET)
3. Inhibition of miR Processing
3.1. Inhibition of the Drosha Site
3.2. Inhibition of Dicer Site
4. Degradation of miR
4.1. Ribonuclease-Targeting Chimera (RIBOTAC)
4.2. Direct miR Degradation
4.2.1. Bleomycin
4.2.2. Direct Degradation via Alternative Cleaving Agents
5. Prospects in Targeting miR with Small Molecules
5.1. Comparison of ASOs vs. Small Molecules
5.2. Future Approaches to Degradation of miRs
5.3. Potential Methods for Indirectly Increasing Depleted miRs
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Lee, R.C.; Feinbaum, R.L.; Ambrost, V. The C. Elegans Heterochronic Gene Lin-4 Encodes Small RNAs with Antisense Complementarity to Lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of Novel Genes Coding for Small Expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-Nucleotide Let-7 RNA Developmental timing in Caenorhabditis elegans. Lett. Nat. 2000, 403, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Hausser, J.; Zavolan, M. Identification and Consequences of MiRNA-Target Interactions-beyond Repression of Gene Expression. Nat. Rev. Genet. 2014, 15, 599–612. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Kong, Y.W.; Cannell, I.G.; De Moor, C.H.; Hill, K.; Garside, P.G.; Hamilton, T.L.; Meijer, H.A.; Dobbyn, H.C.; Stoneley, M.; Spriggs, K.A.; et al. The Mechanism of Micro-RNA-Mediated Translation Repression Is Determined by the Promoter of the Target Gene. Proc. Natl. Acad. Sci. USA 2008, 105, 8866–8871. [Google Scholar] [CrossRef]
- Petersen, C.P.; Bordeleau, M.E.; Pelletier, J.; Sharp, P.A. Short RNAs Repress Translation after Initiation in Mammalian Cells. Mol. Cell 2006, 21, 533–542. [Google Scholar] [CrossRef]
- Willingham, A.T.; Orth, A.P.; Batalov, S.; Peters, E.C.; Wen, B.G.; Aza-Blanc, P.; Hogenesch, J.B.; Schultz, P.G. Molecular Biology: A Strategy for Probing the Function of Noncoding RNAs Finds a Repressor of NFAT. Science 2005, 309, 1570–1573. [Google Scholar] [CrossRef]
- Barbarotto, E.; Schmittgen, T.D.; Calin, G.A. MicroRNAs and Cancer: Profile, Profile, Profile. Int. J. Cancer 2008, 122, 969–977. [Google Scholar] [CrossRef]
- Weir, J.T.; Schluter, D. The Latitudinal Gradient in Recent Speciation and Extinction Rates of Birds and Mammals. Science 2007, 315, 1574–1576. [Google Scholar] [CrossRef] [PubMed]
- Colpaert, R.M.W.; Calore, M. MicroRNAs in Cardiac Diseases. Cells 2019, 8, 737. [Google Scholar] [CrossRef]
- Nelson, P.T.; Wang, W.X.; Rajeev, B.W. MicroRNAs (MiRNAs) in neurodegenerative diseases. Brain Pathol. 2008, 18, 130–138. [Google Scholar] [CrossRef]
- Wang, W.X.; Rajeev, B.W.; Stromberg, A.J.; Ren, N.; Tang, G.; Huang, Q.; Rigoutsos, I.; Nelson, P.T. The Expression of MicroRNA MiR-107 Decreases Early in Alzheimer’s Disease and May Accelerate Disease Progression through Regulation of β-Site Amyloid Precursor Protein-Cleaving Enzyme 1. J. Neurosci. 2008, 28, 1213–1223. [Google Scholar] [CrossRef]
- Hansen, T.; Olsen, L.; Lindow, M.; Jakobsen, K.D.; Ullum, H.; Jonsson, E.; Andreassen, O.A.; Djurovic, S.; Melle, I.; Agartz, I.; et al. Brain Expressed MicroRNAs Implicated in Schizophrenia Etiology. PLoS ONE 2007, 2, e873. [Google Scholar] [CrossRef]
- Garzon, R.; Fabbri, M.; Cimmino, A.; Calin, G.A.; Croce, C.M. MicroRNA Expression and Function in Cancer. Trends Mol. Med. 2006, 12, 580–587. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA Genes Are Transcribed by RNA Polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Borchert, G.M.; Lanier, W.; Davidson, B.L. RNA Polymerase III Transcribes Human MicroRNAs. Nat. Struct. Mol. Biol. 2006, 13, 1097–1101. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lee, Y.; Yeom, K.H.; Nam, J.W.; Heo, I.; Rhee, J.K.; Sohn, S.Y.; Cho, Y.; Zhang, B.T.; Kim, V.N. Molecular Basis for the Recognition of Primary MicroRNAs by the Drosha-DGCR8 Complex. Cell 2006, 125, 887–901. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Cullen, B.R. Sequence Requirements for Micro RNA Processing and Function in Human Cells. RNA 2003, 9, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lee, Y.; Yeom, K.H.; Kim, Y.K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 Complex in Primary MicroRNA Processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Rå Dmark, O.; Kim, S.; et al. The Nuclear RNase III Drosha Initiates MicroRNA Processing. Lett. Nat. 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Shechner, D.M.; Grant, R.A.; Bagby, S.C.; Koldobskaya, Y.; Piccirilli, J.A.; Bartel, D.P. Crystal Structure of the Catalytic Core of an RNA-Polymerase Ribozyme. Science 2009, 326, 1271–1275. [Google Scholar] [CrossRef]
- Zeng, Y.; Cullen, B.R. Structural Requirements for Pre-MicroRNA Binding and Nuclear Export by Exportin 5. Nucleic Acids Res. 2004, 32, 4776–4785. [Google Scholar] [CrossRef] [PubMed]
- Bohnsack, M.T.; Czaplinski, K.; Görlich, D. Exportin 5 Is a RanGTP-Dependent DsRNA-Binding Protein That Mediates Nuclear Export of Pre-MiRNAs. RNA 2004, 10, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Haase, A.D.; Jaskiewicz, L.; Zhang, H.; Lainé, S.; Sack, R.; Gatignol, A.; Filipowicz, W. TRBP, a Regulator of Cellular PKR and HIV-1 Virus Expression, Interacts with Dicer and Functions in RNA Silencing. EMBO Rep. 2005, 6, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kolb, F.A.; Brondani, V.; Billy, E.; Filipowicz, W. Human Dicer Preferentially Cleaves DsRNAs at Their Termini without a Requirement for ATP. EMBO J. 2002, 21, 5875–5885. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.R.; Aderounmu, A.M.; Donelick, H.M.; Bass, B.L. Dicer’s Helicase Domain: A Meeting Place for Regulatory Proteins. Cold Spring Harb. Symp. Quant. Biol. 2019, 84, 185–193. [Google Scholar] [CrossRef] [PubMed]
- MacRae, I.J.; Ma, E.; Zhou, M.; Robinson, C.V.; Doudna, J.A. In Vitro Reconstitution of the Human-Loading Complex. Proc. Natl. Acad. Sci. USA 2008, 105, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. Review MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Maniataki, E.; Mourelatos, Z. A Human, ATP-Independent, RISC Assembly Machine Fueled by Pre-MiRNA. Genes Dev. 2005, 19, 2979–2990. [Google Scholar] [CrossRef]
- Rand, T.A.; Petersen, S.; Du, F.; Wang, X. Argonaute2 Cleaves the Anti-Guide Strand of SiRNA during RISC Activation. Cell 2005, 123, 621–629. [Google Scholar] [CrossRef]
- Robb, G.B.; Rana, T.M. RNA Helicase a Interacts with RISC in Human Cells and Functions in RISC Loading. Mol. Cell 2007, 26, 523–537. [Google Scholar] [CrossRef]
- Wu, W.; Sun, M.; Zou, G.M.; Chen, J. MicroRNA and Cancer: Current Status and Prospective. Int. J. Cancer 2007, 120, 953–960. [Google Scholar] [CrossRef]
- Syeda, Z.A.; Langden, S.S.S.; Munkhzul, C.; Lee, M.; Song, S.J. Regulatory Mechanism of Microrna Expression in Cancer. Int. J. Mol. Sci. 2020, 21, 1723. [Google Scholar] [CrossRef] [PubMed]
- Smolarz, B.; Durczyński, A.; Romanowicz, H.; Szyłło, K.; Hogendorf, P. MiRNAs in Cancer (Review of Literature). Int. J. Mol. Sci. 2022, 23, 2805. [Google Scholar] [CrossRef] [PubMed]
- Aguet, F.; Brown, A.A.; Castel, S.E.; Davis, J.R.; He, Y.; Jo, B.; Mohammadi, P.; Park, Y.S.; Parsana, P.; Segrè, A.V.; et al. Genetic Effects on Gene Expression across Human Tissues. Nature 2017, 550, 204–213. [Google Scholar] [CrossRef]
- Chi, L.H.; Cross, R.S.N.; Redvers, R.P.; Davis, M.; Hediyeh-zadeh, S.; Mathivanan, S.; Samuel, M.; Lucas, E.C.; Mouchemore, K.; Gregory, P.A.; et al. MicroRNA-21 Is Immunosuppressive and pro-Metastatic via Separate Mechanisms. Oncogenesis 2022, 11, 38. [Google Scholar] [CrossRef]
- Bautista-Sánchez, D.; Arriaga-Canon, C.; Pedroza-Torres, A.; De La Rosa-Velázquez, I.A.; González-Barrios, R.; Contreras-Espinosa, L.; Montiel-Manríquez, R.; Castro-Hernández, C.; Fragoso-Ontiveros, V.; Álvarez-Gómez, R.M.; et al. The Promising Role of MiR-21 as a Cancer Biomarker and Its Importance in RNA-Based Therapeutics. Mol. Ther. Nucleic Acids 2020, 20, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Gabriely, G.; Wurdinger, T.; Kesari, S.; Esau, C.C.; Burchard, J.; Linsley, P.S.; Krichevsky, A.M. MicroRNA 21 Promotes Glioma Invasion by Targeting Matrix Metalloproteinase Regulators. Mol. Cell Biol. 2008, 28, 5369–5380. [Google Scholar] [CrossRef]
- Greither, T.; Grochola, L.F.; Udelnow, A.; Lautenschläger, C.; Würl, P.; Taubert, H. Elevated Expression of MicroRNAs 155, 203, 210 and 222 in Pancreatic Tumors Is Associated with Poorer Survival. Int. J. Cancer 2010, 126, 73–80. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Q.; Guan, Y.; Sun, Y.; Wang, X.; Lively, K.; Wang, Y.; Luo, M.; Kim, J.A.; Murphy, E.A.; et al. Breast Cancer Cell–Derived MicroRNA-155 Suppresses Tumor Progression via Enhancing Immune Cell Recruitment and Antitumor Function. J. Clin. Investig. 2022, 132, e157248. [Google Scholar] [CrossRef]
- Stanczyk, J.; Leslie Pedrioli, D.M.; Brentano, F.; Sanchez-Pernaute, O.; Kolling, C.; Gay, R.E.; Detmar, M.; Gay, S.; Kyburz, D. Altered Expression of MicroRNA in Synovial Fibroblasts and Synovial Tissue in Rheumatoid Arthritis. Arthritis Rheum. 2008, 58, 1001–1009. [Google Scholar] [CrossRef]
- Porkka, K.P.; Pfeiffer, M.J.; Waltering, K.K.; Vessella, R.L.; Tammela, T.L.J.; Visakorpi, T. MicroRNA Expression Profiling in Prostate Cancer. Cancer Res. 2007, 67, 6130–6135. [Google Scholar] [CrossRef]
- Du, C.; Liu, C.; Kang, J.; Zhao, G.; Ye, Z.; Huang, S.; Li, Z.; Wu, Z.; Pei, G. MicroRNA MiR-326 Regulates TH-17 Differentiation and Is Associated with the Pathogenesis of Multiple Sclerosis. Nat. Immunol. 2009, 10, 1252–1259. [Google Scholar] [CrossRef]
- Otmani, K.; Lewalle, P. Tumor Suppressor MiRNA in Cancer Cells and the Tumor Microenvironment: Mechanism of Deregulation and Clinical Implications. Front. Oncol. 2021, 11, 708765. [Google Scholar] [CrossRef]
- Gramantieri, L.; Ferracin, M.; Fornari, F.; Veronese, A.; Sabbioni, S.; Liu, C.G.; Calin, G.A.; Giovannini, C.; Ferrazzi, E.; Grazi, G.L.; et al. Cyclin G1 Is a Target of MiR-122a, a MicroRNA Frequently Down-Regulated in Human Hepatocellular Carcinoma. Cancer Res. 2007, 67, 6092–6099. [Google Scholar] [CrossRef] [PubMed]
- Adrian Calin, G.; Dan Dumitru, C.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent Deletions and Down-Regulation of Micro-RNA Genes MiR15 and MiR16 at 13q14 in Chronic Lymphocytic Leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef]
- Park, S.M.; Gaur, A.B.; Lengyel, E.; Peter, M.E. The MiR-200 Family Determines the Epithelial Phenotype of Cancer Cells by Targeting the E-Cadherin Repressors ZEB1 and ZEB2. Genes Dev. 2008, 22, 894–907. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dong, X.; Wang, Z.; Wu, J. MicroRNA-1 in Cardiac Diseases and Cancers. Korean J. Physiol. Pharmacol. 2014, 18, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Gao, R.; Yan, B. Potential Roles of MicroRNA-1 and MicroRNA-133 in cardiovascular disease. Rev. Cardiovasc. Med. 2020, 21, 57–64. [Google Scholar] [CrossRef]
- Van Rooij, E.; Sutherland, L.B.; Qi, X.; Richardson, J.A.; Hill, J.; Olson, E.N. Control of Stress-Dependent Cardiac Growth and Gene Expression by a MicroRNA. Science 2007, 316, 575–579. [Google Scholar] [CrossRef]
- Pereira, P.A.; Tomás, J.F.; Queiroz, J.A.; Figueiras, A.R.; Sousa, F. Recombinant Pre-MiR-29b for Alzheimer’s Disease Therapeutics. Sci. Rep. 2016, 6, 19946. [Google Scholar] [CrossRef]
- Poy, M.N.; Eliasson, L.; Krutzfeldt, J.; Kuwajima, S.; Ma, X.; Macdonald, P.E.; Bastien Pfeffer, S.; Tuschl, T.; Rajewsky, N.; Rorsman, P.; et al. A Pancreatic Islet-Specific MicroRNA Regulates Insulin Secretion. Lett. Nat. 2004, 432, 226–230. [Google Scholar] [CrossRef]
- Xu, J.; Wan, X.; Chen, X.; Fang, Y.; Cheng, X.; Xie, X.; Lu, W. MiR-2861 Acts as a Tumor Suppressor via Targeting EGFR/AKT2/CCND1 Pathway in Cervical Cancer Induced by Human Papillomavirus Virus 16 E6. Sci. Rep. 2016, 6, 28968. [Google Scholar] [CrossRef] [PubMed]
- Jopling, C.L.; Yi, M.; Lancaster, A.M.; Lemon, S.M.; Sarnow, P. Modulation of Hepatitis C Virus RNA Abundance by a Liver-Specific MicroRNA. Science 2005, 309, 1577–1581. [Google Scholar] [CrossRef]
- Qi, Y.; Hu, H.; Guo, H.; Xu, P.; Shi, Z.; Huan, X.; Zhu, Z.; Zhou, M.; Cui, L. MicroRNA Profiling in Plasma of HIV-1 Infected Patients: Potential Markers of Infection and Immune Status. J. Public Health Emerg. 2017, 1, 65. [Google Scholar] [CrossRef]
- Meyer, S.M.; Williams, C.C.; Akahori, Y.; Tanaka, T.; Aikawa, H.; Tong, Y.; Childs-Disney, J.L.; Disney, M.D. Small Molecule Recognition of Disease-Relevant RNA Structures. Chem. Soc. Rev. 2020, 49, 7167–7199. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Bouhss, A.; Clément, M.-J.; Bauvais, C.; Taylor, J.P.; Bollot, G.; Pastré, D. Using the Structural Diversity of RNA: Protein Interfaces to Selectively Target RNA with Small Molecules in Cells: Methods and Perspectives. Front. Mol. Biosci. 2023, 10, 1298441. [Google Scholar] [CrossRef]
- Santos, R.; Ursu, O.; Gaulton, A.; Bento, A.P.; Donadi, R.S.; Bologa, C.G.; Karlsson, A.; Al-Lazikani, B.; Hersey, A.; Oprea, T.I.; et al. A Comprehensive Map of Molecular Drug Targets. Nat. Rev. Drug Discov. 2016, 16, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Costales, M.G.; Childs-Disney, J.L.; Haniff, H.S.; Disney, M.D. How We Think about Targeting RNA with Small Molecules. J. Med. Chem. 2020, 63, 8880–8900. [Google Scholar] [CrossRef]
- Su, H.; Peng, Z.; Yang, J. Recognition of Small Molecule-RNA Binding Sites Using RNA Sequence and Structure. Bioinformatics 2021, 37, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Umuhire Juru, A.; Hargrove, A.E. Frameworks for Targeting RNA with Small Molecules. J. Biol. Chem. 2021, 296, 100191. [Google Scholar] [CrossRef] [PubMed]
- Staedel, C.; Tran, T.P.A.; Giraud, J.; Darfeuille, F.; Di Giorgio, A.; Tourasse, N.J.; Salin, F.; Uriac, P.; Duca, M. Modulation of Oncogenic MiRNA Biogenesis Using Functionalized Polyamines. Sci. Rep. 2018, 8, 1667. [Google Scholar] [CrossRef] [PubMed]
- Warner, K.D.; Hajdin, C.E.; Weeks, K.M. Principles for Targeting RNA with Drug-like Small Molecules. Nat. Rev. Drug Discov. 2018, 17, 547–558. [Google Scholar] [CrossRef]
- Vo, D.D.; Tran, T.P.A.; Staedel, C.; Benhida, R.; Darfeuille, F.; Digiorgio, A.; Duca, M. Oncogenic MicroRNAs Biogenesis as a Drug Target: Structure-Activity Relationship Studies on New Aminoglycoside Conjugates. Chem. Eur. J. 2016, 22, 5350–5362. [Google Scholar] [CrossRef] [PubMed]
- Garner, A.L.; Lorenz, D.A.; Sandoval, J.; Gallagher, E.E.; Kerk, S.A.; Kaur, T.; Menon, A. Tetracyclines as Inhibitors of Pre-MicroRNA Maturation: A Disconnection between RNA Binding and Inhibition. ACS Med. Chem. Lett. 2019, 10, 816–821. [Google Scholar] [CrossRef]
- Pomplun, S.; Gates, Z.P.; Zhang, G.; Quartararo, A.J.; Pentelute, B.L. Discovery of Nucleic Acid Binding Molecules from Combinatorial Biohybrid Nucleobase Peptide Libraries. J. Am. Chem. Soc. 2020, 142, 19642–19651. [Google Scholar] [CrossRef]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA Therapeutics—Challenges and Potential Solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef]
- Das, S.; Shah, R.; Dimmeler, S.; Freedman, J.E.; Holley, C.; Lee, J.M.; Moore, K.; Musunuru, K.; Wang, D.Z.; Xiao, J.; et al. Noncoding RNAs in Cardiovascular Disease: Current Knowledge, Tools and Technologies for Investigation, and Future Directions: A Scientific Statement from the American Heart Association. Circ. Genom. Precis. Med. 2020, 13, E000062. [Google Scholar] [CrossRef]
- Childs-Disney, J.L.; Yang, X.; Gibaut, Q.M.R.; Tong, Y.; Batey, R.T.; Disney, M.D. Targeting RNA Structures with Small Molecules. Nat. Rev. Drug Discov. 2022, 21, 736–762. [Google Scholar] [CrossRef]
- Lightfoot, H.L.; Smith, G.F. Targeting RNA with Small Molecules—A Safety Perspective. Br. J. Pharmacol. 2023, 1–20. [Google Scholar] [CrossRef]
- Zafferani, M.; Hargrove, A.E. Small Molecule Targeting of Biologically Relevant RNA Tertiary and Quaternary Structures. Cell Chem. Biol. 2021, 28, 594–609. [Google Scholar] [CrossRef] [PubMed]
- Baisden, J.T.; Childs-Disney, J.L.; Ryan, L.S.; Disney, M.D. Affecting RNA Biology Genome-Wide by Binding Small Molecules and Chemically Induced Proximity. Curr. Opin. Chem. Biol. 2021, 62, 119–129. [Google Scholar] [CrossRef]
- McGovern-Gooch, K.R.; Baird, N.J. Fluorescence-Based Investigations of RNA-Small Molecule Interactions. Methods 2019, 167, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Heikkinen, L.; Wang, C.; Yang, Y.; Sun, H.; Wong, G. Trends in the Development of MiRNA Bioinformatics Tools. Brief Bioinform. 2019, 20, 1836–1852. [Google Scholar] [CrossRef] [PubMed]
- Diener, C.; Keller, A.; Meese, E. Emerging Concepts of MiRNA Therapeutics: From Cells to Clinic. Trends Genet. 2022, 38, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, Z.; Talkhabi, M.; Taleahmad, S. Identification of Potential MicroRNA Diagnostic Panels and Uncovering Regulatory Mechanisms in Breast Cancer Pathogenesis. Sci. Rep. 2022, 12, 20135. [Google Scholar] [CrossRef]
- Akhtar, M.M.; Micolucci, L.; Islam, M.S.; Olivieri, F.; Procopio, A.D. Bioinformatic Tools for MicroRNA Dissection. Nucleic Acids Res. 2016, 44, 24–44. [Google Scholar] [CrossRef] [PubMed]
- Velagapudi, S.P.; Gallo, S.M.; Disney, M.D. Sequence-Based Design of Bioactive Small Molecules That Target Precursor MicroRNAs. Nat. Chem. Biol. 2014, 10, 291–297. [Google Scholar] [CrossRef]
- Baldassarre, A.; Felli, C.; Prantera, G.; Masotti, A. Circulating MicroRNAs and Bioinformatics Tools to Discover Novel Diagnostic Biomarkers of Pediatric Diseases. Genes 2017, 8, 234. [Google Scholar] [CrossRef]
- Kalaigar, S.S.; Rajashekar, R.B.; Nataraj, S.M.; Vishwanath, P.; Prashant, A. Bioinformatic Tools for the Identification of MicroRNAs Regulating the Transcription Factors in Patients with β-Thalassemia. Bioinform. Biol. Insights 2022, 16, 11779322221115536. [Google Scholar] [CrossRef] [PubMed]
- Gironda-Martínez, A.; Donckele, E.J.; Samain, F.; Neri, D. DNA-Encoded Chemical Libraries: A Comprehensive Review with Succesful Stories and Future Challenges. ACS Pharmacol. Transl. Sci. 2021, 4, 1265–1279. [Google Scholar] [CrossRef]
- Panei, F.P.; Torchet, R.; Menager, H.; Gkeka, P.; Bonomi, M. HARIBOSS: A Curated Database of RNA-Small Molecules Structures to Aid Rational Drug Design. Bioinformatics 2022, 38, 4185–4193. [Google Scholar] [CrossRef]
- Disney, M.D.; Winkelsas, A.M.; Velagapudi, S.P.; Southern, M.; Fallahi, M.; Childs-Disney, J.L. Inforna 2.0: A Platform for the Sequence-Based Design of Small Molecules Targeting Structured RNAs. ACS Chem. Biol. 2016, 11, 1720–1728. [Google Scholar] [CrossRef]
- Ursu, A.; Childs-Disney, J.L.; Andrews, R.J.; O’Leary, C.A.; Meyer, S.M.; Angelbello, A.J.; Moss, W.N.; Disney, M.D. Design of Small Molecules Targeting RNA Structure from Sequence. Chem. Soc. Rev. 2020, 49, 7252–7270. [Google Scholar] [CrossRef]
- Sun, S.; Yang, J.; Zhang, Z. RNALigands: A Database and Web Server for RNA-Ligand Interactions. RNA 2022, 28, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Childs-Disney, J.L.; Disney, M.D. Small Molecule Targeting of a MicroRNA Associated with Hepatocellular Carcinoma. ACS Chem. Biol. 2016, 11, 375–380. [Google Scholar] [CrossRef]
- Suresh, B.M.; Tong, Y.; Abegg, D.; Adibekian, A.; Childs-Disney, J.L.; Disney, M.D. Altering the Cleaving Effector in Chimeric Molecules That Target RNA Enhances Cellular Selectivity. ACS Chem. Biol. 2023, 18, 2385–2393. [Google Scholar] [CrossRef]
- Childs-Disney, J.L.; Tran, T.; Vummidi, B.R.; Velagapudi, S.P.; Haniff, H.S.; Matsumoto, Y.; Crynen, G.; Southern, M.R.; Biswas, A.; Wang, Z.F.; et al. A Massively Parallel Selection of Small Molecule-RNA Motif Binding Partners Informs Design of an Antiviral from Sequence. Chem 2018, 4, 2384–2404. [Google Scholar] [CrossRef]
- Costales, M.G.; Hoch, D.G.; Abegg, D.; Childs-Disney, J.L.; Velagapudi, S.P.; Adibekian, A.; Disney, M.D. A Designed Small Molecule Inhibitor of a Non-Coding RNA Sensitizes HER2 Negative Cancers to Herceptin. J. Am. Chem. Soc. 2019, 141, 2960–2974. [Google Scholar] [CrossRef]
- Benhamou, R.I.; Choudhary, S.; Lekah, E.; Tong, Y.; Disney, M.D. Bioinformatic Searching for Optimal RNA Targets of Dimeric Compounds Informs Design of a MicroRNA-27a Inhibitor. ACS Chem. Biol. 2022, 17, 5–10. [Google Scholar] [CrossRef]
- Velagapudi, S.P.; Cameron, M.D.; Haga, C.L.; Rosenberg, L.H.; Lafitte, M.; Duckett, D.R.; Phinney, D.G.; Disney, M.D. Design of a Small Molecule against an Oncogenic Noncoding RNA. Proc. Natl. Acad. Sci. USA 2016, 113, 5898–5903. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, X.; Abegg, D.; Tanaka, T.; Tong, Y.; Benhamou, R.I.; Baisden, J.; Crynen, G.; Meyer, S.M.; Cameron, M.D.; et al. Reprogramming of Protein-Targeted Small-Molecule Medicines to RNA by Ribonuclease Recruitment. J. Am. Chem. Soc. 2021, 143, 13044–13055. [Google Scholar] [CrossRef]
- Costales, M.G.; Aikawa, H.; Li, Y.; Childs-Disney, J.L.; Abegg, D.; Hoch, D.G.; Pradeep Velagapudi, S.; Nakai, Y.; Khan, T.; Wang, K.W.; et al. Small-Molecule Targeted Recruitment of a Nuclease to Cleave an Oncogenic RNA in a Mouse Model of Metastatic Cancer. Proc. Natl. Acad. Sci. USA 2020, 117, 2406–2411. [Google Scholar] [CrossRef]
- Agu, P.C.; Afiukwa, C.A.; Orji, O.U.; Ezeh, E.M.; Ofoke, I.H.; Ogbu, C.O.; Ugwuja, E.I.; Aja, P.M. Molecular Docking as a Tool for the Discovery of Molecular Targets of Nutraceuticals in Diseases Management. Sci. Rep. 2023, 13, 13398. [Google Scholar] [CrossRef]
- Mcconkey, B.J.; Sobolev, V.; Edelman, M. The Performance of Current Methods in Ligand-Protein Docking. Curr. Sci. 2002, 83, 845–856. [Google Scholar]
- Fan, R.; Xiao, C.; Wan, X.; Cha, W.; Miao, Y.; Zhou, Y.; Qin, C.; Cui, T.; Su, F.; Shan, X. Small Molecules with Big Roles in MicroRNA Chemical Biology and MicroRNA-Targeted Therapeutics. RNA Biol. 2019, 16, 707–718. [Google Scholar] [CrossRef]
- Yu, F.; Li, B.; Sun, J.; Qi, J.; De Wilde, R.L.; Torres-de la Roche, L.A.; Li, C.; Ahmad, S.; Shi, W.; Li, X.; et al. PSRR: A Web Server for Predicting the Regulation of MiRNAs Expression by Small Molecules. Front. Mol. Biosci. 2022, 9, 817294. [Google Scholar] [CrossRef]
- Tran, T.P.A.; Poulet, S.; Pernak, M.; Rayar, A.; Azoulay, S.; Di Giorgio, A.; Duca, M. Development of 2-Deoxystreptamine-Nucleobase Conjugates for the Inhibition of Oncogenic MiRNA Production. RSC Med. Chem. 2021, 13, 311–319. [Google Scholar] [CrossRef]
- Becquart, C.; Le Roch, M.; Azoulay, S.; Uriac, P.; Di Giorgio, A.; Duca, M. Exploring Heterocycle-Spermine Conjugates as Modulators of Oncogenic MicroRNAs Biogenesis. ACS Omega 2018, 3, 16500–16508. [Google Scholar] [CrossRef]
- Bose, D.; Jayaraj, G.; Suryawanshi, H.; Agarwala, P.; Pore, S.K.; Banerjee, R.; Maiti, S. The Tuberculosis Drug Streptomycin as a Potential Cancer Therapeutic: Inhibition of Mir-21 Function by Directly Targeting Its Precursor. Angew. Chem. Int. Ed. 2012, 51, 1019–1023. [Google Scholar] [CrossRef]
- Dallaire, P.; Tan, H.; Szulwach, K.; Ma, C.; Jin, P.; Major, F. Structural Dynamics Control the MicroRNA Maturation Pathway. Nucleic Acids Res. 2016, 44, 9956–9964. [Google Scholar] [CrossRef]
- Lv, Y.; Wang, S.; Meng, F.; Yang, L.; Wang, Z.; Wang, J.; Chen, X.; Jiang, W.; Li, Y.; Li, X. Identifying Novel Associations between Small Molecules and MiRNAs Based on Integrated Molecular Networks. Bioinformatics 2015, 31, 3638–3644. [Google Scholar] [CrossRef]
- Li, J.; Lin, H.; Wang, Y.; Li, Z.; Wu, B. Prediction of Potential Small Molecule−miRNA Associations Based on Heterogeneous Network Representation Learning. Front. Genet. 2022, 13, 1079053. [Google Scholar] [CrossRef]
- Ottl, J.; Leder, L.; Schaefer, J.V.; Dumelin, C.E. Encoded Library Technologies as Integrated Lead Finding Platforms for Drug Discovery. Molecules 2019, 24, 1629. [Google Scholar] [CrossRef] [PubMed]
- Szymański, P.; Markowicz, M.; Mikiciuk-Olasik, E. Adaptation of High-Throughput Screening in Drug Discovery-Toxicological Screening Tests. Int. J. Mol. Sci. 2012, 13, 427–452. [Google Scholar] [CrossRef] [PubMed]
- Benhamou, R.I.; Suresh, B.M.; Tong, Y.; Cochrane, W.G.; Cavett, V.; Vezina-Dawod, S.; Abegg, D.; Childs-Disney, J.L.; Adibekian, A.; Paegel, B.M.; et al. DNA-Encoded Library versus RNA-Encoded Library Selection Enables Design of an Oncogenic Noncoding RNA Inhibitor. Proc. Natl. Acad. Sci. USA 2022, 119, e2114971119. [Google Scholar] [CrossRef]
- William, L. Jorgensen the Many Roles of Computation in Drug Discovery. Science 2004, 303, 1813–1818. [Google Scholar] [CrossRef]
- Velagapudi, S.P.; Disney, M.D. Two-Dimensional Combinatorial Screening Enables the Bottom-up Design of a MicroRNA-10b Inhibitor. Chem. Commun. 2014, 50, 3027–3029. [Google Scholar] [CrossRef]
- Shcheholeva, I.; Fernández-Remacha, D.; Estrada-Tejedor, R.; Duca, M.; Michelet, V. De-Novo Design of Pre-MiR-21 Maturation Inhibitors: Synthesis and Activity Assessment. Chem. Eur. J. 2023, 29, e202300825. [Google Scholar] [CrossRef]
- Peng, B.; Thorsell, A.; Karlberg, T.; Schüler, H.; Yao, S.Q. Small Molecule Microarray Based Discovery of PARP14 Inhibitors. Angew. Chem. 2017, 129, 254–259. [Google Scholar] [CrossRef]
- Disney, M.D.; Labuda, L.P.; Paul, D.J.; Poplawski, S.G.; Pushechnikov, A.; Tran, T.; Velagapudi, S.P.; Wu, M.; Childs-Disney, J.L. Two-Dimensional Combinatorial Screening Identifies Specific Aminoglycoside-RNA Internal Loop Partners. J. Am. Chem. Soc. 2008, 130, 11185–11194. [Google Scholar] [CrossRef]
- Uttamchandani, M.; Yao Editors, S.Q. The Expanding World of Small Molecule Microarrays. In Small Molecule Microarrays: Methods in Molecular Biology, 2nd ed.; 2017; Volume 1518, pp. 1–257. [Google Scholar] [CrossRef]
- Sztuba-Solinska, J.; Shenoy, S.R.; Gareiss, P.; Krumpe, L.R.H.; Le Grice, S.F.J.; O’Keefe, B.R.; Schneekloth, J.S. Identification of Biologically Active, HIV TAR RNA-Binding Small Molecules Using Small Molecule Microarrays. J. Am. Chem. Soc. 2014, 136, 8402–8410. [Google Scholar] [CrossRef]
- Connelly, C.M.; Boer, R.E.; Moon, M.H.; Gareiss, P.; Schneekloth, J.S. Discovery of Inhibitors of MicroRNA-21 Processing Using Small Molecule Microarrays. ACS Chem. Biol. 2017, 12, 435–443. [Google Scholar] [CrossRef]
- Hall, M.D.; Yasgar, A.; Peryea, T.; Braisted, J.C.; Jadhav, A.; Simeonov, A.; Coussens, N.P. Fluorescence Polarization Assays in High-Throughput Screening and Drug Discovery: A Review. Methods Appl. Fluoresc. 2016, 4, 022001. [Google Scholar] [CrossRef] [PubMed]
- Sedgwick, A.C.; Brewster, J.T.; Wu, T.; Feng, X.; Bull, S.D.; Qian, X.; Sessler, J.L.; James, T.D.; Anslyn, E.V.; Sun, X. Indicator Displacement Assays (IDAs): The Past, Present and Future. Chem. Soc. Rev. 2021, 50, 9–38. [Google Scholar] [CrossRef] [PubMed]
- Wicks, S.L.; Hargrove, A.E. Fluorescent Indicator Displacement Assays to Identify and Characterize Small Molecule Interactions with RNA. Methods 2019, 167, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Murata, A.; Harada, Y.; Fukuzumi, T.; Nakatani, K. Fluorescent Indicator Displacement Assay of Ligands Targeting 10 MicroRNA Precursors. Bioorg. Med. Chem. 2013, 21, 7101–7106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Umemoto, S.; Nakatani, K. Fluorescent Indicator Displacement Assay for Ligand-RNA Interactions. J. Am. Chem. Soc. 2010, 132, 3660–3661. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Bhattarai, U.; Guo, Z.F.; Liang, F. Sen Regulating MiRNA-21 Biogenesis by Bifunctional Small Molecules. J. Am. Chem. Soc. 2017, 139, 4987–4990. [Google Scholar] [CrossRef]
- Shrestha, D.; Jenei, A.; Nagy, P.; Vereb, G.; Szöllősi, J. Understanding FRET as a Research Tool for Cellular Studies. Int. J. Mol. Sci. 2015, 16, 6718–6756. [Google Scholar] [CrossRef]
- Sekar, R.B.; Periasamy, A. Fluorescence Resonance Energy Transfer (FRET) Microscopy Imaging of Live Cell Protein Localizations. J. Cell Biol. 2003, 160, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Broussard, J.A.; Rappaz, B.; Webb, D.J.; Brown, C.M. Fluorescence Resonance Energy Transfer Microscopy as Demonstrated by Measuring the Activation of the Serine/Threonine Kinase Akt. Nat. Protoc. 2013, 8, 265–281. [Google Scholar] [CrossRef]
- Watson, B.S.; Hazlett, T.L.; Eccleston, J.F.; Davis, C.; Jameson, D.M.; Johnson, A.E. Macromolecular Arrangement in the Aminoacyl-TRNA-Elongation Factor Tu*GTP Ternary Complex. A Fluorescence Energy Transfer Study. Biochemistry 1995, 34, 7904–7912. [Google Scholar] [CrossRef]
- Broussard, J.A.; Green, K.J. Research Techniques Made Simple: Methodology and Applications of Förster Resonance Energy Transfer (FRET) Microscopy. J. Investig. Dermatol. 2017, 137, e185–e191. [Google Scholar] [CrossRef]
- Nguyen, T.A.; Sarkar, P.; Veetil, J.V.; Koushik, S.V.; Vogel, S.S. Fluorescence Polarization and Fluctuation Analysis Monitors Subunit Proximity, Stoichiometry, and Protein Complex Hydrodynamics. PLoS ONE 2012, 7, e38209. [Google Scholar] [CrossRef]
- Davies, B.P.; Arenz, C. A Homogenous Assay for Micro RNA Maturation. Angew. Chem. Int. Ed. 2006, 45, 5550–5552. [Google Scholar] [CrossRef]
- Bose, D.; Jayaraj, G.G.; Kumar, S.; Maiti, S. A Molecular-Beacon-Based Screen for Small Molecule Inhibitors of MiRNA Maturation. ACS Chem. Biol. 2013, 8, 930–938. [Google Scholar] [CrossRef]
- Xie, Y.; Dix, A.V.; Tor, Y. FRET Enabled Real Time Detection of RNA-Small Molecule Binding. J. Am. Chem. Soc. 2009, 131, 17605–17614. [Google Scholar] [CrossRef]
- Vo, D.D.; Duca, M. Design of Multimodal Small Molecules Targeting MiRNAs Biogenesis: Synthesis and in Vitro Evaluation. Methods Mol. Biol. 2017, 1517, 137–154. [Google Scholar] [CrossRef]
- Maucort, C.; Vo, D.D.; Aouad, S.; Charrat, C.; Azoulay, S.; Di Giorgio, A.; Duca, M. Design and Implementation of Synthetic RNA Binders for the Inhibition of MiR-21 Biogenesis. ACS Med. Chem. Lett. 2021, 12, 899–906. [Google Scholar] [CrossRef]
- Yan, H.; Liang, F. Sen MiRNA Inhibition by Proximity-Enabled Dicer Inactivation. Methods 2019, 167, 117–123. [Google Scholar] [CrossRef]
- Zeng, Y.; Cullen, B.R. Efficient Processing of Primary MicroRNA Hairpins by Drosha Requires Flanking Nonstructured RNA Sequences. J. Biol. Chem. 2005, 280, 27595–27603. [Google Scholar] [CrossRef]
- Monroig, P.d.C.; Chen, L.; Zhang, S.; Calin, G.A. Small Molecule Compounds Targeting MiRNAs for Cancer Therapy. Adv. Drug Deliv. Rev. 2015, 81, 104–116. [Google Scholar] [CrossRef]
- Davis, B.N.; Hata, A. Regulation of MicroRNA Biogenesis: A MiRiad of Mechanisms. Cell Commun. Signal. 2009, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Sall, A.; Yang, D. MicroRNA: An Emerging Therapeutic Target and Intervention Tool. Int. J. Mol. Sci. 2008, 9, 978–999. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Li, W.; She, H.; Dou, J.; Duong, D.M.; Du, Y.; Yang, S.H.; Seyfried, N.T.; Fu, H.; Gao, G.; et al. Stress Induces P38 MAPK-Mediated Phosphorylation and Inhibition of Drosha-Dependent Cell Survival. Mol. Cell 2015, 57, 721–734. [Google Scholar] [CrossRef] [PubMed]
- Gromak, N.; Dienstbier, M.; Macias, S.; Plass, M.; Eyras, E.; Cáceres, J.F.; Proudfoot, N.J. Drosha Regulates Gene Expression Independently of RNA Cleavage Function. Cell Rep. 2013, 5, 1499–1510. [Google Scholar] [CrossRef] [PubMed]
- Kuehbacher, A.; Urbich, C.; Zeiher, A.M.; Dimmeler, S. Role of Dicer and Drosha for Endothelial MicroRNA Expression and Angiogenesis. Circ. Res. 2007, 101, 59–68. [Google Scholar] [CrossRef]
- Haniff, H.S.; Liu, X.; Tong, Y.; Meyer, S.M.; Knerr, L.; Lemurell, M.; Abegg, D.; Aikawa, H.; Adibekian, A.; Disney, M.D. A Structure-Specific Small Molecule Inhibits a MiRNA-200 Family Member Precursor and Reverses a Type 2 Diabetes Phenotype. Cell Chem. Biol. 2022, 29, 300–311.e10. [Google Scholar] [CrossRef]
- Yang, S.; Maurin, T.; Robine, N.; Rasmussen, K.D.; Jeffrey, K.L.; Chandwani, R.; Papapetrou, E.P.; Sadelain, M.; O’Carroll, D.; Lai, E.C. Conserved Vertebrate Mir-451 Provides a Platform for Dicer-Independent, Ago2-Mediated MicroRNA Biogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 15163–15168. [Google Scholar] [CrossRef]
- Yoshida, T.; Asano, Y.; Ui-Tei, K. Modulation of Microrna Processing by Dicer via Its Associated Dsrna Binding Proteins. Noncoding RNA 2021, 7, 57. [Google Scholar] [CrossRef]
- Sczepanski, J.T.; Joyce, G.F. Specific Inhibition of MicroRNA Processing Using l -RNA Aptamers. J. Am. Chem. Soc. 2015, 137, 16032–16037. [Google Scholar] [CrossRef]
- Shortridge, M.D.; Chaubey, B.; Zhang, H.J.; Pavelitz, T.; Vidadala, V.; Tang, C.; Olsen, G.L.; Calin, G.A.; Varani, G. Drug-Like Small Molecules That Inhibit Expression of the Oncogenic MicroRNA-21. ACS Chem. Biol. 2023, 18, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Zhang, J.; Qian, X.; Han, L.; Zhang, K.; Chen, L.; Liu, J.; Ren, Y.; Yang, M.; Zhang, A.; et al. AC1MMYR2, an Inhibitor of Dicer-Mediated Biogenesis of Oncomir MiR-21, Reverses Epithelial-Mesenchymal Transition and Suppresses Tumor Growth and Progression. Cancer Res. 2013, 73, 5519–5531. [Google Scholar] [CrossRef] [PubMed]
- Naro, Y.; Thomas, M.; Stephens, M.D.; Connelly, C.M.; Deiters, A. Aryl Amide Small-Molecule Inhibitors of MicroRNA MiR-21 Function. Bioorg. Med. Chem. Lett. 2015, 25, 4793–4796. [Google Scholar] [CrossRef]
- Ankenbruck, N.; Kumbhare, R.; Naro, Y.; Thomas, M.; Gardner, L.; Emanuelson, C.; Deiters, A. Small Molecule Inhibition of MicroRNA-21 Expression Reduces Cell Viability and Microtumor Formation. Bioorg. Med. Chem. 2019, 27, 3735–3743. [Google Scholar] [CrossRef] [PubMed]
- Costales, M.G.; Haga, C.L.; Velagapudi, S.P.; Childs-Disney, J.L.; Phinney, D.G.; Disney, M.D. Small Molecule Inhibition of MicroRNA-210 Reprograms an Oncogenic Hypoxic Circuit. J. Am. Chem. Soc. 2017, 139, 3446–3455. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Haniff, H.S.; Suresh, B.M.; Yang, D.; Zhang, P.; Crynen, G.; Teijaro, C.N.; Yan, W.; Abegg, D.; Adibekian, A.; et al. Rational Approach to Identify RNA Targets of Natural Products Enables Identification of Nocathiacin as an Inhibitor of an Oncogenic RNA. ACS Chem. Biol. 2022, 17, 474–482. [Google Scholar] [CrossRef]
- Dey, S.K.; Jaffrey, S.R. RIBOTACs: Small Molecules Target RNA for Degradation. Cell Chem. Biol. 2019, 26, 1047–1049. [Google Scholar] [CrossRef]
- Tong, Y.; Lee, Y.; Liu, X.; Childs-Disney, J.L.; Suresh, B.M.; Benhamou, R.I.; Yang, C.; Li, W.; Costales, M.G.; Haniff, H.S.; et al. Programming Inactive RNA-Binding Small Molecules into Bioactive Degraders. Nature 2023, 618, 169–179. [Google Scholar] [CrossRef]
- Hecht, S.M. RNA Degradation by Bleomycin, a Naturally Occurring Bioconjugate. Bioconjugate Chem. 1994, 5, 513–526. [Google Scholar] [CrossRef]
- Haj-Yahia, S.; Nandi, A.; Benhamou, R.I. Targeted Degradation of Structured RNAs via Ribonuclease-Targeting Chimeras (RiboTacs). Expert Opin. Drug Discov. 2023, 18, 929–942. [Google Scholar] [CrossRef]
- Haniff, H.S.; Tong, Y.; Liu, X.; Chen, J.L.; Suresh, B.M.; Andrews, R.J.; Peterson, J.M.; O’Leary, C.A.; Benhamou, R.I.; Moss, W.N.; et al. Targeting the SARS-CoV-2 RNA Genome with Small Molecule Binders and Ribonuclease Targeting Chimera (RiboTAC) Degraders. ACS Cent. Sci. 2020, 6, 1713–1721. [Google Scholar] [CrossRef]
- Costales, M.G.; Suresh, B.; Vishnu, K.; Disney, M.D. Targeted Degradation of a Hypoxia-Associated Non-Coding RNA Enhances the Selectivity of a Small Molecule Interacting with RNA. Cell Chem. Biol. 2019, 26, 1180–1186. [Google Scholar] [CrossRef]
- Meyer, S.M.; Tanaka, T.; Zanon, P.R.A.; Baisden, J.T.; Abegg, D.; Yang, X.; Akahori, Y.; Alshakarchi, Z.; Cameron, M.D.; Adibekian, A.; et al. DNA-Encoded Library Screening to Inform Design of a Ribonuclease Targeting Chimera (RiboTAC). J. Am. Chem. Soc. 2022, 144, 21096–21102. [Google Scholar] [CrossRef] [PubMed]
- Costales, M.G.; Matsumoto, Y.; Velagapudi, S.P.; Disney, M.D. Small Molecule Targeted Recruitment of a Nuclease to RNA. J. Am. Chem. Soc. 2018, 140, 6741–6744. [Google Scholar] [CrossRef] [PubMed]
- Maucort, C.; Bonnet, M.; Ortuno, J.C.; Tucker, G.; Quissac, E.; Verreault, M.; Azoulay, S.; Di Giorgio, C.; Di Giorgio, A.; Duca, M. Synthesis of Bleomycin-Inspired RNA Ligands Targeting the Biogenesis of Oncogenic MiRNAs. J. Med. Chem. 2023, 66, 10639–10657. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Disney, M.D. Precise Small Molecule Degradation of a Noncoding RNA Identifies Cellular Binding Sites and Modulates an Oncogenic Phenotype. ACS Chem. Biol. 2018, 13, 3065–3071. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Haniff, H.S.; Childs-Disney, J.L.; Shuster, A.; Aikawa, H.; Adibekian, A.; Disney, M.D. Targeted Degradation of the Oncogenic MicroRNA 17-92 Cluster by Structure-Targeting Ligands. J. Am. Chem. Soc. 2020, 142, 6970–6982. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, J.; Wang, Y.; Feng, Y.; Li, J.; Zhang, Y. Bifunctional Chimera for Ligand-Directed Photo-Degradation of Oncogenic MicroRNA. Chem. Commun. 2023, 59, 7639–7642. [Google Scholar] [CrossRef] [PubMed]
- Holmes, C.E.; Duff, R.J.; Van Der Marel, G.A.; Van Boom, J.; Hecht, S.M. On the Chemistry of RNA Degradation by Fe-Bleomycin. Bioorg. Med. Chem. 1997, 5, 1235–1248. [Google Scholar] [CrossRef] [PubMed]
- Katano, K.; An, H.; Aoyagi, Y.; Overhand, M.; Sucheck, S.J.; Stevens, W.C.; Hess, C.D.; Zhou, X.; Hecht, S.M. Total Synthesis of Bleomycin Group Antibiotics. Total Syntheses of Bleomycin Demethyl A2, Bleomycin A2, and Decarbamoyl Bleomycin Demethyl A2. J. Am. Chem. Soc. 1998, 120, 11285–11296. [Google Scholar] [CrossRef]
- Boyer, V.; Moustacchi, E.; Sage, E. Sequence Specificity in Photoreaction of Various Psoralen Derivatives with DNA: Role in Biological Activity. Biochemistry 1988, 27, 3011–3018. [Google Scholar] [CrossRef] [PubMed]
- Kane, S.A.; Hecht, S.M.; Sun, J.S.; Garestier, T.; Helene, C. Specific Cleavage of a DNA Triple Helix by FeII.Bleomycin. Nucleic Acids Res. 1995, 34, 4219–4224. [Google Scholar] [CrossRef]
- Xie, T.; Liang, J.; Guo, R.; Liu, N.; Noble, P.W.; Jiang, D. Comprehensive MicroRNA Analysis in Bleomycin-Induced Pulmonary Fibrosis Identifies Multiple Sites of Molecular Regulation. Physiol. Genom. 2011, 43, 479–487. [Google Scholar] [CrossRef]
- Gagliardi, M.; Ashizawa, A.T. The Challenges and Strategies of Antisense Oligonucleotide Drug Delivery. Biomedicines 2021, 9, 433. [Google Scholar] [CrossRef]
- Mikutis, S.; Rebelo, M.; Yankova, E.; Gu, M.; Tang, C.; Coelho, A.R.; Yang, M.; Hazemi, M.E.; Pires De Miranda, M.; Eleftheriou, M.; et al. Proximity-Induced Nucleic Acid Degrader (PINAD) Approach to Targeted RNA Degradation Using Small Molecules. ACS Cent. Sci. 2023, 9, 892–904. [Google Scholar] [CrossRef]
- Imamura, T.; Komatsu, S.; Ichikawa, D.; Miyamae, M.; Okajima, W.; Ohashi, T.; Kiuchi, J.; Nishibeppu, K.; Konishi, H.; Shiozaki, A.; et al. Depleted Tumor Suppressor MiR-107 in Plasma Relates to Tumor Progression and Is a Novel Therapeutic Target in Pancreatic Cancer. Sci. Rep. 2017, 7, e120. [Google Scholar] [CrossRef]
- Roos, M.; Pradère, U.; Ngondo, R.P.; Behera, A.; Allegrini, S.; Civenni, G.; Zagalak, J.A.; Marchand, J.R.; Menzi, M.; Towbin, H.; et al. A Small-Molecule Inhibitor of Lin28. ACS Chem. Biol. 2016, 11, 2773–2781. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.; Byun, W.G.; Koo, J.Y.; Park, H.; Park, S.B. Discovery of a Small-Molecule Inhibitor of Protein-MicroRNA Interaction Using Binding Assay with a Site-Specifically Labeled Lin28. J. Am. Chem. Soc. 2016, 138, 13630–13638. [Google Scholar] [CrossRef] [PubMed]
- Borgelt, L.; Hohnen, L.; Pallesen, J.S.; Hommen, P.; Goebel, G.L.; Bosica, F.; Liu, Y.; O’Mahony, G.; Wu, P. N-Biphenyl Pyrrolinones and Dibenzofurans as RNA-Binding Protein LIN28 Inhibitors Disrupting the LIN28-Let-7 Interaction. ACS Med. Chem. Lett. 2023, 14, 1707–1715. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pri/Pre-miRs | Small Molecules | Discovery Methods | Motifs | Ref. |
|---|---|---|---|---|
| Mir-27a |  | DEL |  | [95] |
| Mir-27a |  | InfoRNA |  | [91] |
| MiR-27a |  Homo-dimer | InfoRNA |  | [91] |
| MiR-10b |  | 2DCS |  | [96] |
| MiR-96 |  Hetero-dimer | InfoRNA Chem-CLIP |  | [92] |
| MiR-515 |  Homo-dimer | InfoRNA |  | [90] |
| MiR-525 |  | InfoRNA |  | [87] |
| Pri/Pr-miRs | Small Molecules | Discovery Methods | Motifs | Ref. |
|---|---|---|---|---|
| MiR-21 |  | Fluorescence Polarization-Based Screening |  | [121] |
| MiR-21 |  | NMR Assay |  | [136] |
| MiR-21 |  | Dual Luciferase Reporter Assay |  | [103] |
| MiR-21 |  | FRET |  | [132] |
| MiR-21 |  | High-Throughput Screening |  | [137] |
| MiR-21 |  | High-Throughput Screening | Not defined | [138] |
| MiR-21 |  | Docking |  | [110] |
| MiR-21 |  | High-Throughput Screening | Not defined | [139] |
| MiR-21 |  | MicroArray |  | [115] |
| MiR-210 |  | InfoRNA |  | [140] |
| MiR-372 |  | Molecular Docking |  | [101] |
| MiR-372 |  | FRET |  | [131] |
| MiR-372 |  | Molecular Docking |  | [102] |
| MiR-18a |  | 2DCS |  | [141] |
| MiR-200 |  Hetero-dimer | Chem Clip |  | [142] |
| Pri/Pre-miRs | Small Molecules | Discovery Methods | Motifs | Ref. |
|---|---|---|---|---|
| MiR-155 |  | 2DCS |  | [152] |
| MiR-21 |  | Binding assay + 2DCS |  | [93] |
| MiR-21 |  | Binding assay + 2DCS |  | [157] |
| MiR-21 |  | InfoRNA |  | [94] |
| MiR-210 |  | InfoRNA |  | [156] |
| MiR-96 |  | InfoRNA |  | [158] |
| MiR-372 |  | InfoRNA |  | [88] |
| MiR-21 |  | Molecular docking |  | [159] |
| MiR-96 |  | InfoRNA |  | [160] |
| MiR-17-92 |  | InfoRNA |  | [161] |
| MiR-210 |  | InfoRNA |  | [162] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tadesse, K.; Benhamou, R.I. Targeting MicroRNAs with Small Molecules. Non-Coding RNA 2024, 10, 17. https://doi.org/10.3390/ncrna10020017
Tadesse K, Benhamou RI. Targeting MicroRNAs with Small Molecules. Non-Coding RNA. 2024; 10(2):17. https://doi.org/10.3390/ncrna10020017
Chicago/Turabian StyleTadesse, Kisanet, and Raphael I. Benhamou. 2024. "Targeting MicroRNAs with Small Molecules" Non-Coding RNA 10, no. 2: 17. https://doi.org/10.3390/ncrna10020017
APA StyleTadesse, K., & Benhamou, R. I. (2024). Targeting MicroRNAs with Small Molecules. Non-Coding RNA, 10(2), 17. https://doi.org/10.3390/ncrna10020017