
A Phylogenetic Approach to Structural Variation in Organization of Nuclear Group I Introns and Their Ribozymes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
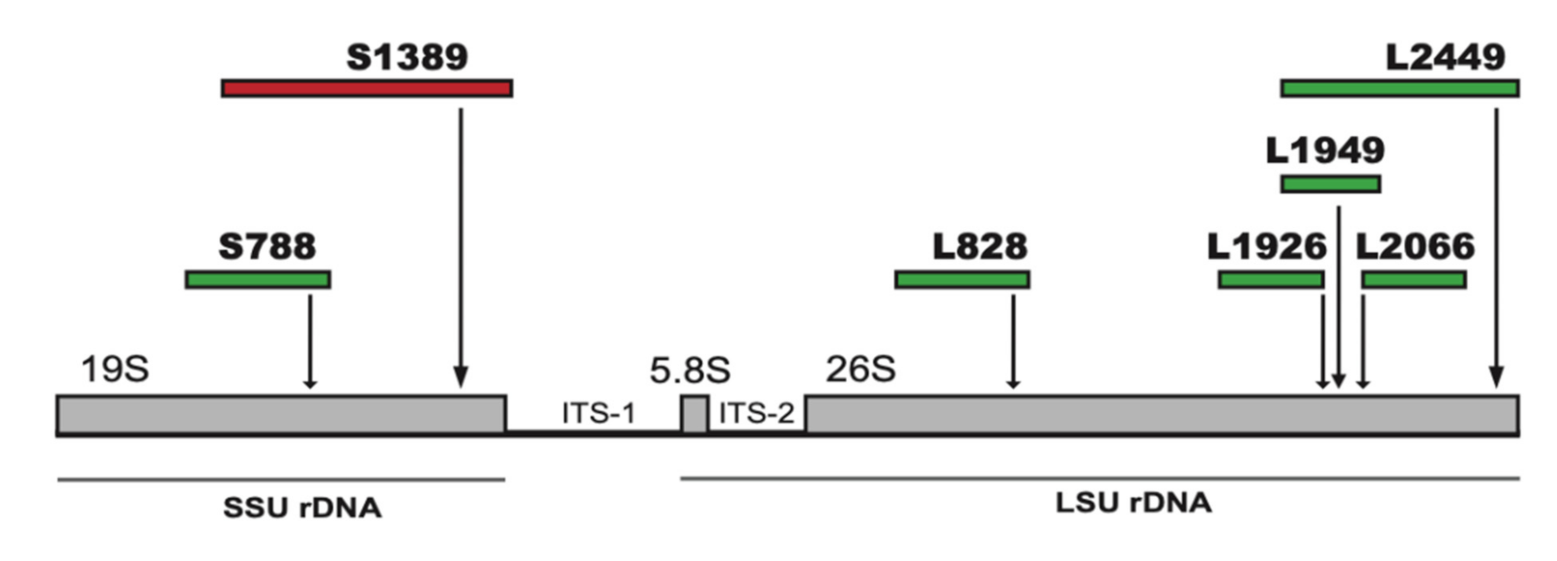
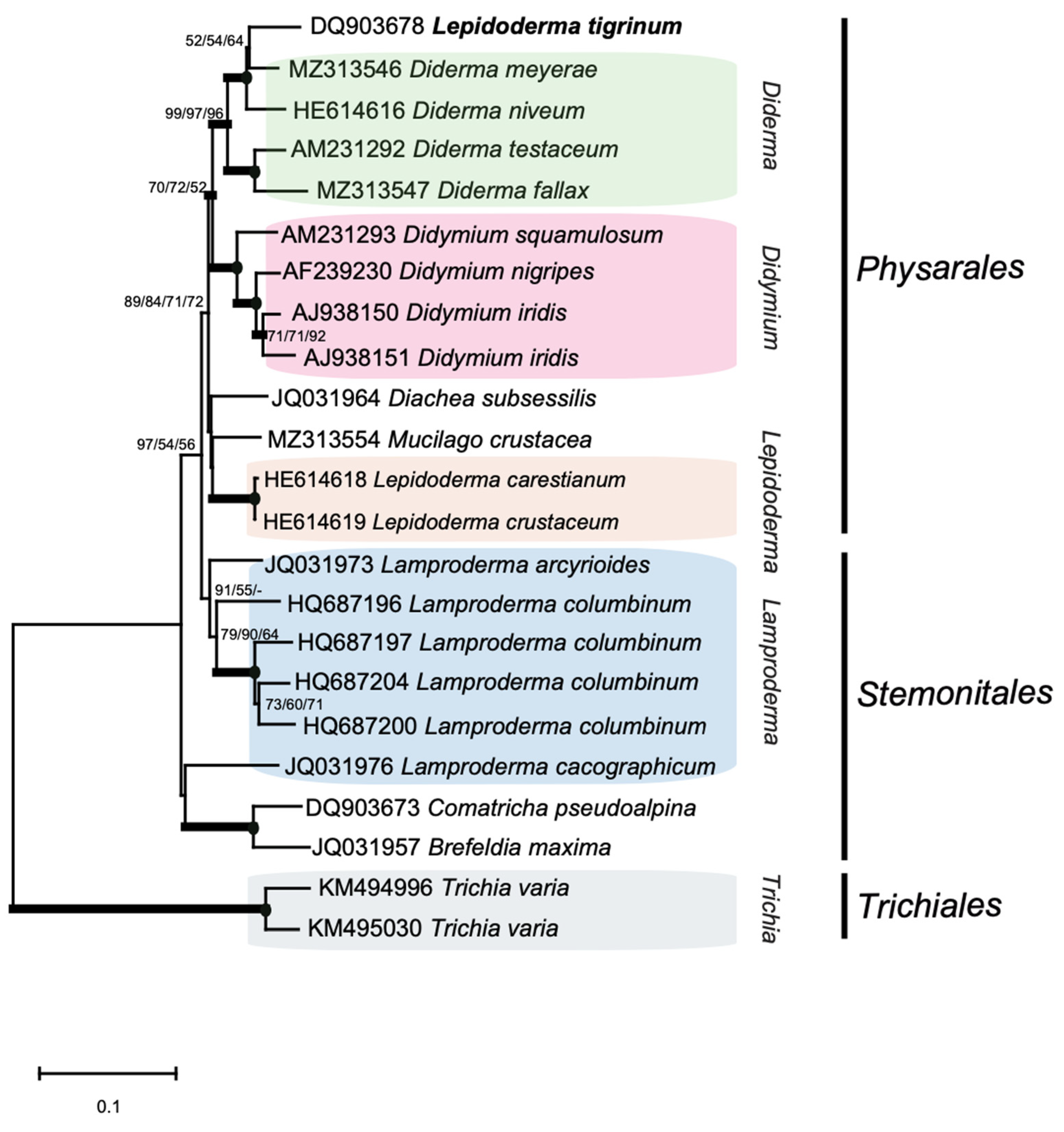
2.1. The Nuclear rDNA of Mucilago Crustacea Harbors Multiple Group I Intron Insertions
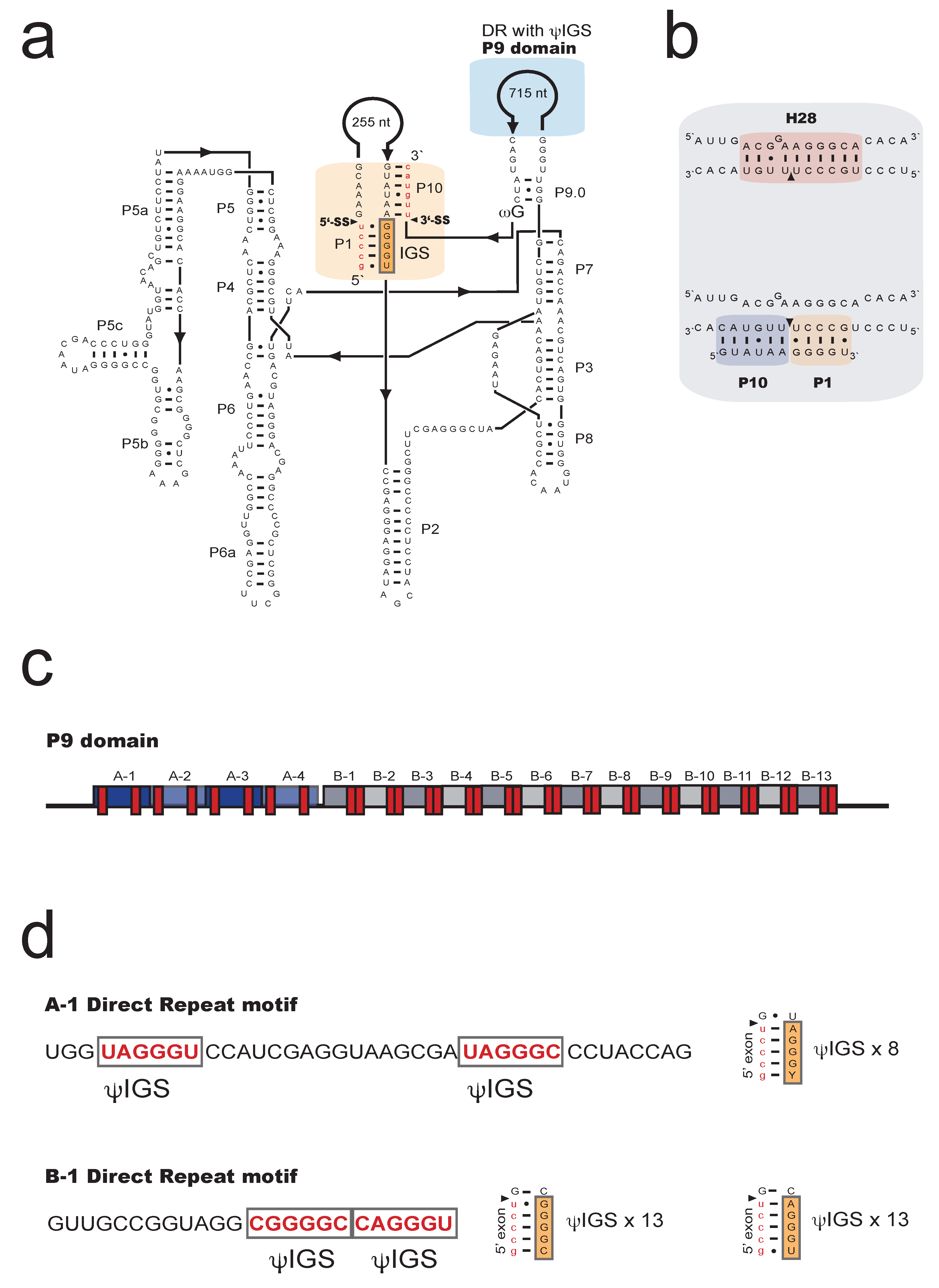
2.2. A Complex Group I Intron in M. Crustacea SSU rRNA Helix 28 at Insertion Site S1389
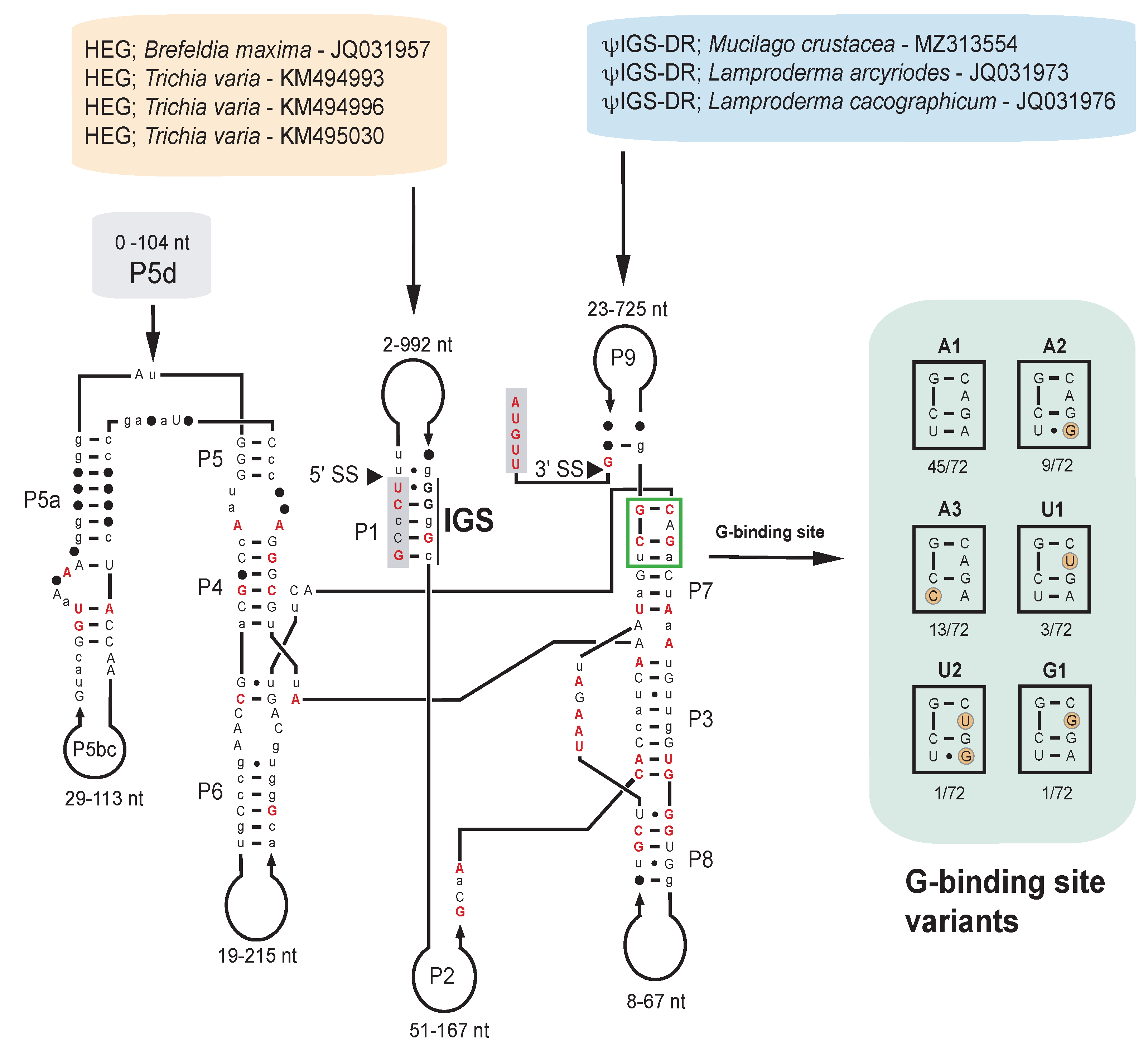
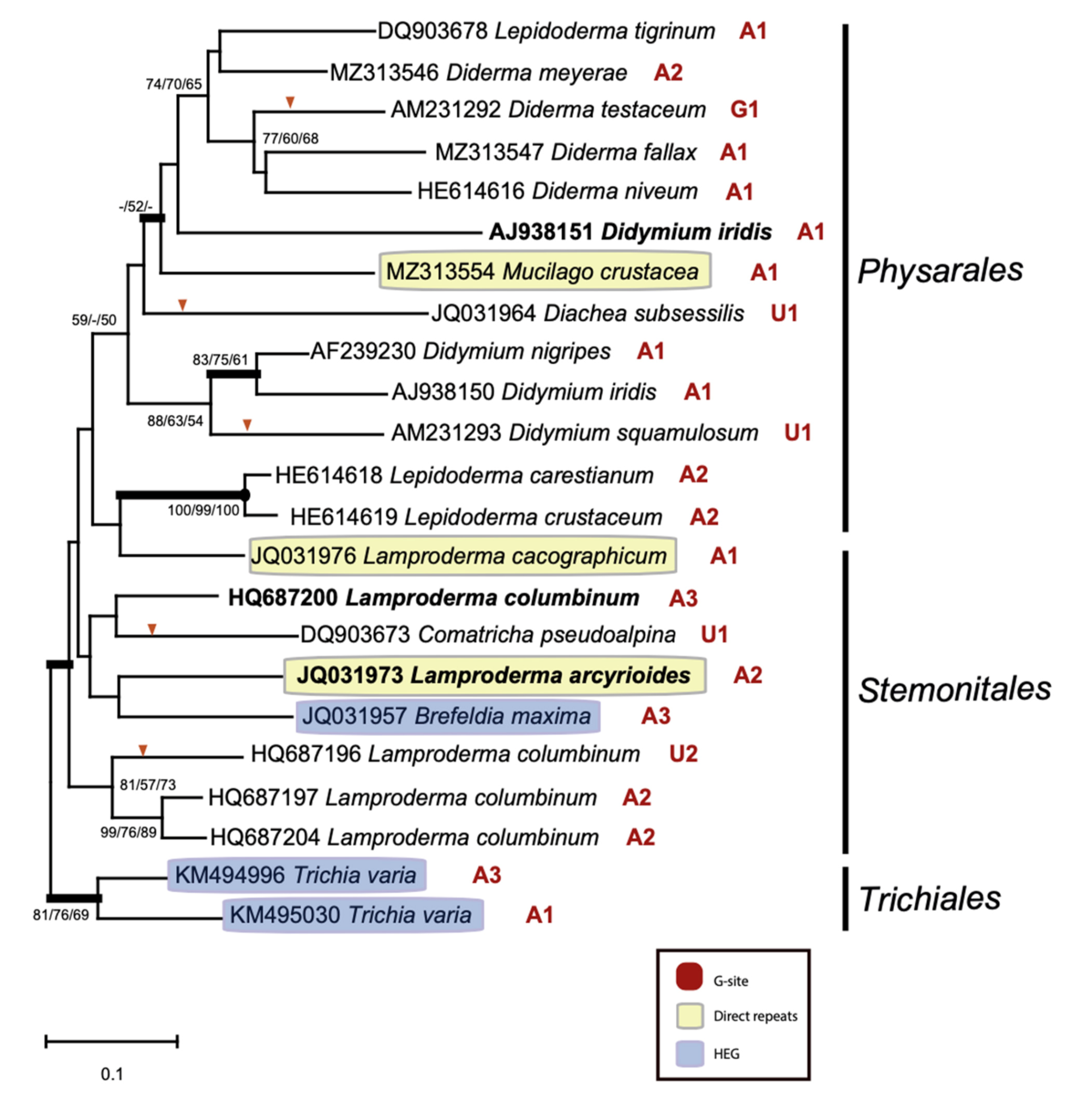
2.3. Structural Variation among the S1389 Introns
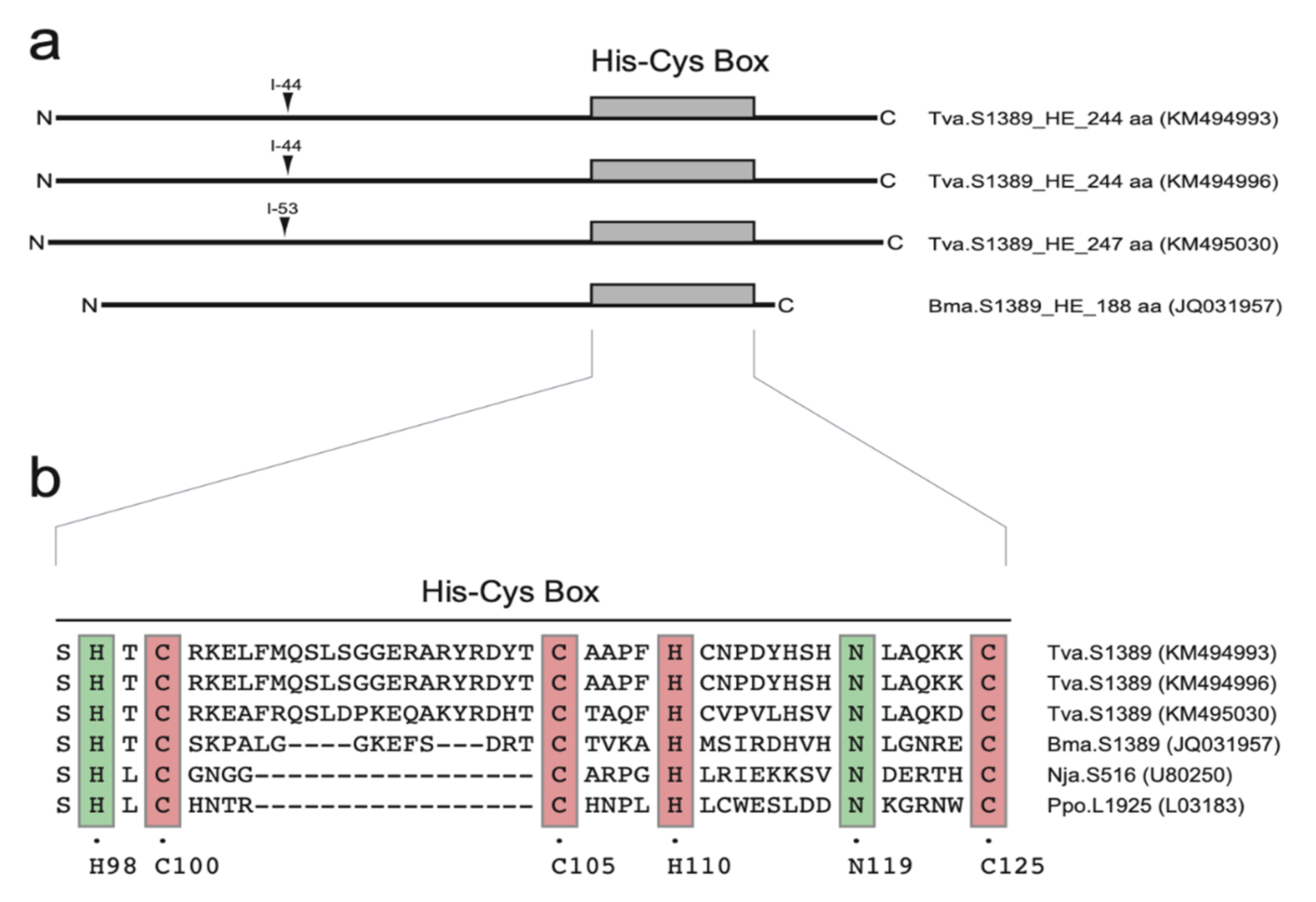
2.4. Spliceosomal Introns Interrupt S1389 Group I Intron HEGs
2.5. S1389 Group I Introns Appear Vertically and Horizontally Inherited
3. Discussion
4. Materials and Methods
4.1. Culturing, DNA Isolation, and DNA Sequencing
4.2. Sequence Alignments
4.3. Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cech, T.R. Self-splicing of group I introns. Ann. Rev. Biochem. 1990, 59, 543–568. [Google Scholar] [CrossRef]
- Haugen, P.; Simon, D.M.; Bhattacharya, D. The natural history of group I introns. Trends Genet. 2005, 21, 111–119. [Google Scholar] [CrossRef]
- Hedberg, A.; Johansen, S.D. Nuclear group I introns in self-splicing and beyond. Mob. DNA 2013, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Cech, T.R.; Damberger, S.H.; Gutell, R.R. Representation of the secondary and tertiary structure of group I introns. Nat. Struct. Biol. 1994, 1, 273–280. [Google Scholar] [CrossRef]
- Vicens, Q.; Cech, T.R. Atomic level architecture of group I introns revealed. Trends Biochem. Sci. 2006, 31, 41–51. [Google Scholar] [CrossRef]
- Haugen, P.; Reeb, V.; Lutzoni, F.; Bhattacharya, D. The evolution of homing endonuclease genes and group I introns in nuclear rDNA. Mol. Biol. Evol. 2004, 21, 129–140. [Google Scholar] [CrossRef]
- Zhou, Y.; Lu, C.; Wu, Q.-J.; Wang, Y.; Sun, Z.-T.; Deng, J.-C.; Zhang, Y. GISSD: Group I intron sequence and structure database. Nucleic Acids Res. 2008, 36, D31–D37. [Google Scholar] [CrossRef]
- Guo, F.; Gooding, A.R.; Cech, T.R. Structure of the Tetrahymena ribozyme: Base triple sandwich and metal ion at the active site. Mol. Cell 2004, 16, 351–362. [Google Scholar] [CrossRef]
- Andersen, K.L.; Beckert, B.; Masquida, B.; Johansen, S.D.; Nielsen, H. Accumulation of stable full-length circular group I intron RNAs during heat-shock. Molecules 2016, 21, 1451. [Google Scholar] [CrossRef] [Green Version]
- Been, M.D.; Cech, T.R. Selection of circularization sites in a group I IVS RNA requires multiple alignments of an internal template-like sequence. Cell 1987, 50, 951–961. [Google Scholar] [CrossRef]
- Nielsen, H.; Fiskaa, T.; Birgisdottir, Á.B.; Haugen, P.; Einvik, C.; Johansen, S. The ability to form full-length intron RNA circles is a general property of nuclear group I introns. RNA 2003, 9, 1464–1475. [Google Scholar] [CrossRef] [Green Version]
- Johansen, S.; Embley, T.M.; Willassen, N.P. A family of nuclear homing endonucleases. Nucleic Acids Res. 1993, 21, 4405. [Google Scholar] [CrossRef] [PubMed]
- Stoddard, B.L. Homing endonuclease structure and function. Q. Rev. Biophys. 2005, 38, 49–95. [Google Scholar] [CrossRef]
- Muscarella, D.E.; Vogt, V.M. A mobile group I intron in the nuclear rDNA of Physarum polycephalum. Cell 1989, 56, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Johansen, S.; Elde, M.; Vader, A.; Haugen, P.; Haugli, K.; Haugli, F. In vivo mobility of a group I twintron in nuclear ribosomal DNA of the myxomycete Didymium iridis. Mol. Microbiol. 1997, 24, 737–745. [Google Scholar] [CrossRef]
- Edgell, D.R.; Chalamcharla, V.R.; Belfort, M. Learning to live together: Mutualism between self-splicing introns and their hosts. BMC Biol. 2011, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Roman, J.; Rubin, M.N.; Woodson, S.A. Sequence specificity of in vivo reverse splicing of the Tetrahymena group I intron. RNA 1999, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birgisdottir, Á.B.; Johansen, S. Site-specific reverse splicing of a HEG-containing group I intron in ribosomal RNA. Nucleic Acids Res. 2005, 33, 2042–2051. [Google Scholar] [CrossRef] [Green Version]
- Margulis, L.; Schwartz, K. Five Kingdoms: An Illustrated Guide to the Phyla of Life in Earth, 3rd ed.; Freeman: New York, NY, USA, 1997; ISBN 9780716730279. [Google Scholar]
- Nielsen, H.; Johansen, S.D. Group I introns: Moving in new directions. RNA Biol. 2009, 6, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Neubert, H.; Nowotny, W.; Baumann, K. Die Myxomyceten; Karlheinz Baumann Verlag: Gomaringen, Germany, 1995; Volume 2, ISBN 3-929822-01-6. [Google Scholar]
- Wikmark, O.G.; Haugen, P.; Lundblad, E.W.; Haugli, K.; Johansen, S.D. The molecular evolution and structural organization of group I introns at position 1389 in nuclear small subunit rDNA of myxomycetes. J. Euk. Microbiol. 2007, 54, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Johansen, S.; Haugen, P. A new nomenclature of group I introns in ribosomal DNA. RNA 2001, 7, 935–936. [Google Scholar] [CrossRef] [Green Version]
- Mohan, S.; Donohue, J.P.; Noller, H.F. Molecular mechanics of 30S subunit head rotation. Proc. Natl Acad. Sci. USA 2014, 111, 13325–13330. [Google Scholar] [CrossRef] [Green Version]
- Elde, M.; Haugen, P.; Willassen, N.P.; Johansen, S. I-NjaI, a nuclear intron-encoded homing endonuclease from Naegleria, generates a pentanucleotide 3′ cleavage-overhang within a 19 base-pair partially symmetric DNA recognition site. Eur. J. Biochem. 1999, 259, 281–288. [Google Scholar] [CrossRef]
- Flick, K.E.; Jurica, M.S.; Monnat, R.J.J.; Stoddard, B.L. DNA binding and cleavage by the nuclear intron-encoded homing endonuclease I-PpoI. Nature 1998, 394, 96–101. [Google Scholar] [CrossRef]
- Fiore-Donno, A.M.; Mayer, M.; Baldauf, S.L.; Pawlowski, J. Evolution of dark-spored myxomycetes (slime-molds): Molecules versus morphology. Mol. Phylogenet. Evol. 2008, 46, 878–889. [Google Scholar] [CrossRef]
- Fiore-Donno, A.M.; Kamono, A.; Meyer, M.; Schnittler, M.; Fukui, M.; Cavalier-Smith, T. 18S rDNA phylogeny of lamproderma and allied genera (Stemonirales, Myxomycetes, Amoebozoa). PLoS ONE 2012, 7, e35359. [Google Scholar] [CrossRef] [Green Version]
- Nandipati, S.C.; Haugli, K.; Coucheron, D.H.; Haskins, E.F.; Johansen, S.D. Polyphyletic origin of the genus Physarum (Physarales, Myxomycetes) revealed by nuclear rDNA mini-chromosome analysis and group I intron synapomorphy. BMC Evol. Biol. 2012, 12, 166. [Google Scholar] [CrossRef] [Green Version]
- Cainelli, R.; de Haan, M.; Meyer, M.; Bonkowski, M.; Fiore-Donno, A.M. Phylogeny of Physarida (Amoebozoa, Myxogastria) based on the small-subunit ribosomal RNA gene, redefinition of Physarum pusillum s. str. and reinstatement of P. gravidum Morgan. J. Eukaryot. Microbiol. 2020, 67, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Johansen, S.; Johansen, T.; Haugli, F. Structure and evolution of myxomycete nuclear group I introns: A model for horizontal transfer by intron homing. Curr. Genet. 1992, 22, 297–304. [Google Scholar] [CrossRef]
- Ruoff, B.; Johansen, S.; Vogt, V.M. Characterization of the self-splicing products of a mobile intron from the nuclear rDNA of Physarum polycephalum. Nucleic Acids Res. 1992, 20, 5899–5906. [Google Scholar] [CrossRef] [Green Version]
- Johansen, S.; Vogt, V.M. An intron in the nuclear ribosomal DNA of Didymium iridis codes for a group I ribozyme and a novel ribozyme that cooperate in self-splicing. Cell 1994, 76, 725–734. [Google Scholar] [CrossRef]
- Lundblad, E.W.; Einvik, C.; Rønning, S.; Haugli, K.; Johansen, S. Twelve group I introns in the same pre-rRNA transcript of the myxomycete Fuligo septica: RNA processing and evolution. Mol. Biol. Evol. 2004, 21, 1283–1293. [Google Scholar] [CrossRef] [Green Version]
- Wikmark, O.G.; Haugen, P.; Haugli, K.; Johansen, S.D. Obligatory group I introns with unusual features at positions 1949 and 2449 in nuclear LSU rDNA of Didymiaceae myxomycetes. Mol. Phylogenet. Evol. 2007, 43, 596–604. [Google Scholar] [CrossRef]
- Vader, A.; Naess, J.; Haugli, K.; Haugli, F.; Johansen, S. Nucleolar introns from Physarum flavicomum contain insertion elements that may explain how mobile group I introns gained their open reading frames. Nucleic Acids Res. 1994, 22, 4553–4559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golden, B.L. Group I introns: Biochemical and crystallographic characterization of the active site structure. In Ribozymes and RNA Catalysis; Lilley, D.M.J., Eckstein, F., Eds.; RSC Press: Cambridge, UK, 2008; pp. 178–200. ISBN 978-0-85404-253-1. [Google Scholar]
- Knitt, D.S.; Narlikar, G.J.; Herschlag, D. Dissection of the role of the conserved G.U pair in group I RNA self-splicing. Biochemistry 1994, 33, 13864–13879. [Google Scholar] [CrossRef]
- Ikawa, Y.; Yoshimura, T.; Hara, H.; Shiraishi, H.; Inoue, T. Two conserved structural components, A-rich bulge and P4XJ6/7 base-triples, in activating the group I ribozymes. Genes Cells 2002, 7, 1205–1215. [Google Scholar] [CrossRef] [PubMed]
- Tanner, M.A.; Cech, T.R. Joining the two domains of a group I ribozyme to form the catalytic core. Science 1997, 275, 847–849. [Google Scholar] [CrossRef]
- Michel, F.; Westhof, E. Modelling of the three-dimensional architecture of group-I catalytic introns based on comparative sequence analysis. J. Mol. Biol. 1990, 216, 585–610. [Google Scholar] [CrossRef]
- Couture, S.; Ellington, A.D.; Gerber, A.S.; Cherry, J.M.; Doudna, J.A.; Green, R.; Hanna, M.; Pace, U.; Rajagopal, J.; Szostak, J.W. Mutational analysis of conserved nucleotides in self-splicing group I intron. J. Mol. Biol. 1990, 215, 345–358. [Google Scholar] [CrossRef]
- Oe, Y.; Ikawa, Y.; Shiraishi, H.; Inoue, T. Analysis of the P7 region within the catalytic core of the Tetrahymena ribozyme by employing in vitro selection. Nucleic Acids Symp. Ser. 2000, 44, 197–198. [Google Scholar] [CrossRef]
- Palazzo, A.F.; Koonin, E.V. Functional long non-coding RNAs evolve from junk transcripts. Cell 2020, 183, 1151–1161. [Google Scholar] [CrossRef]
- Vader, A.; Nielsen, H.; Johansen, S. In vivo expression of the nucleolar group I intron-encoded I-DirI homing endonuclease involves the removal of a spliceosomal intron. EMBO J. 1999, 18, 1003–1013. [Google Scholar] [CrossRef] [Green Version]
- Johansen, S.D.; Vader, A.; Sjøttem, E.; Nielsen, H. In vivo expression of a group I intron HEG from the antisense strand of Didymium ribosomal DNA. RNA Biol. 2006, 3, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Johansen, S.D.; Haugen, P.; Nielsen, H. Expression of protein-coding genes embedded in ribosomal DNA. Biol. Chem. 2007, 388, 679–686. [Google Scholar] [CrossRef]
- Schlautmann, L.P.; Gehring, N.H. A day in the life of the exon junction complex. Biomolecules 2020, 10, 866. [Google Scholar] [CrossRef]
- Haugen, P.; Coucheron, D.H.; Rønning, S.B.; Haugli, K.; Johansen, S. The molecular evolution and structural organization of self-splicing group I introns at position 516 in nuclear SSU rDNA of myxomycetes. J. Eukaryot. Microbiol. 2003, 50, 283–292. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvement in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Johansen, T.; Johansen, S.; Haugli, F.B. Nucleotide sequence of the Physarum polycephalum small subunit ribosomal RNA as inferred from the gene sequence: Secondary structure and evolutionary implications. Curr. Genet. 1988, 14, 265–273. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F.; Nielsen, R.; Bollback, J.P. Bayesian inference of phylogeny and its impact on evolutionary biology. Science 2001, 294, 2310–2314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; ISBN 9780195135855. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furulund, B.M.N.; Karlsen, B.O.; Babiak, I.; Johansen, S.D. A Phylogenetic Approach to Structural Variation in Organization of Nuclear Group I Introns and Their Ribozymes. Non-Coding RNA 2021, 7, 43. https://doi.org/10.3390/ncrna7030043
Furulund BMN, Karlsen BO, Babiak I, Johansen SD. A Phylogenetic Approach to Structural Variation in Organization of Nuclear Group I Introns and Their Ribozymes. Non-Coding RNA. 2021; 7(3):43. https://doi.org/10.3390/ncrna7030043
Chicago/Turabian StyleFurulund, Betty M. N., Bård O. Karlsen, Igor Babiak, and Steinar D. Johansen. 2021. "A Phylogenetic Approach to Structural Variation in Organization of Nuclear Group I Introns and Their Ribozymes" Non-Coding RNA 7, no. 3: 43. https://doi.org/10.3390/ncrna7030043
APA StyleFurulund, B. M. N., Karlsen, B. O., Babiak, I., & Johansen, S. D. (2021). A Phylogenetic Approach to Structural Variation in Organization of Nuclear Group I Introns and Their Ribozymes. Non-Coding RNA, 7(3), 43. https://doi.org/10.3390/ncrna7030043