
Inhibition of the lncRNA Coded within Transglutaminase 2 Gene Impacts Several Relevant Networks in MCF-7 Breast Cancer Cells

 ,
,  , , , ,
, , , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Structure and Expression of lncRNA of Transglutaminase Type 2 Gene
1.2. TGM2 Expression in Breast Cancer
2. Results
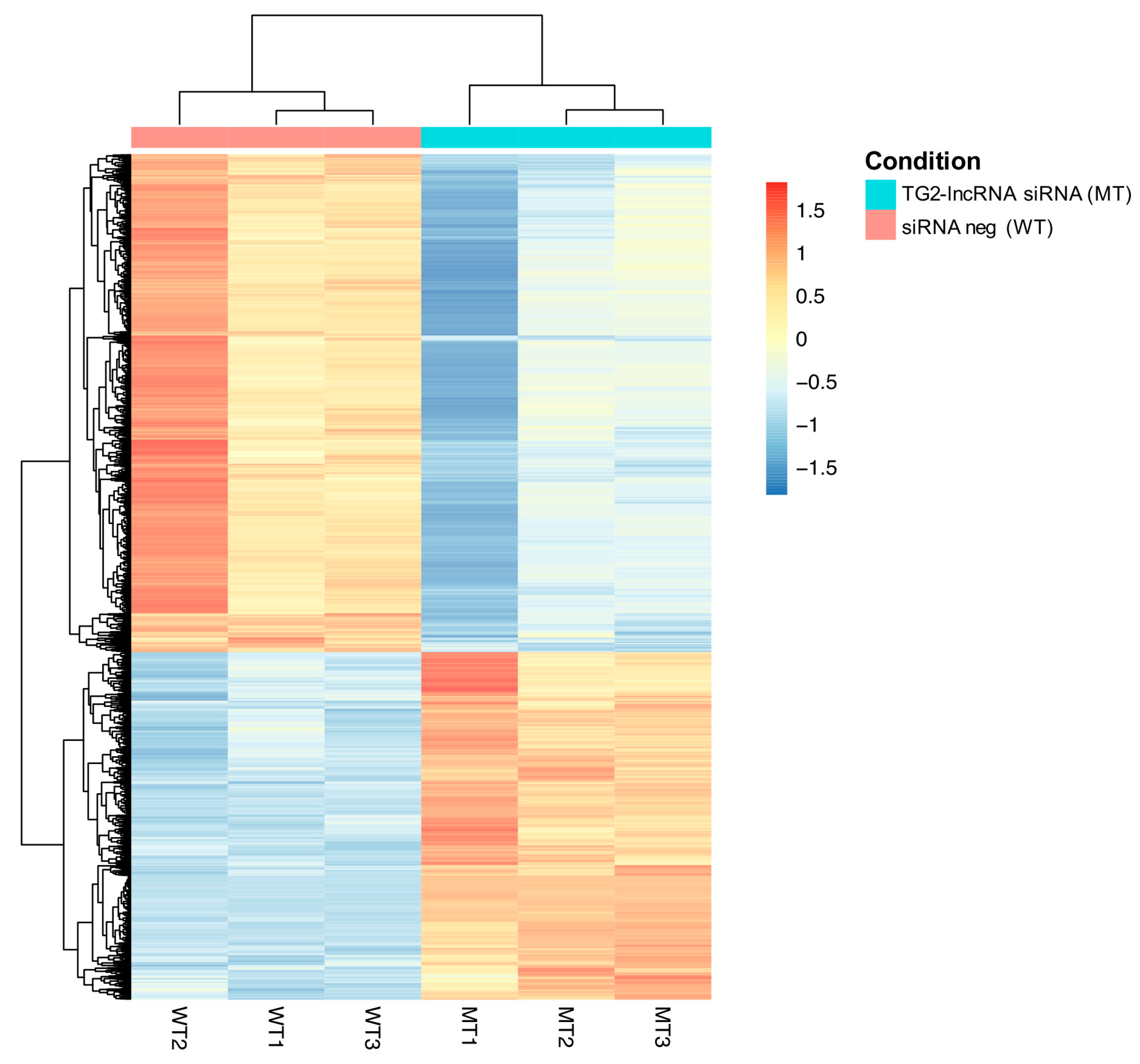
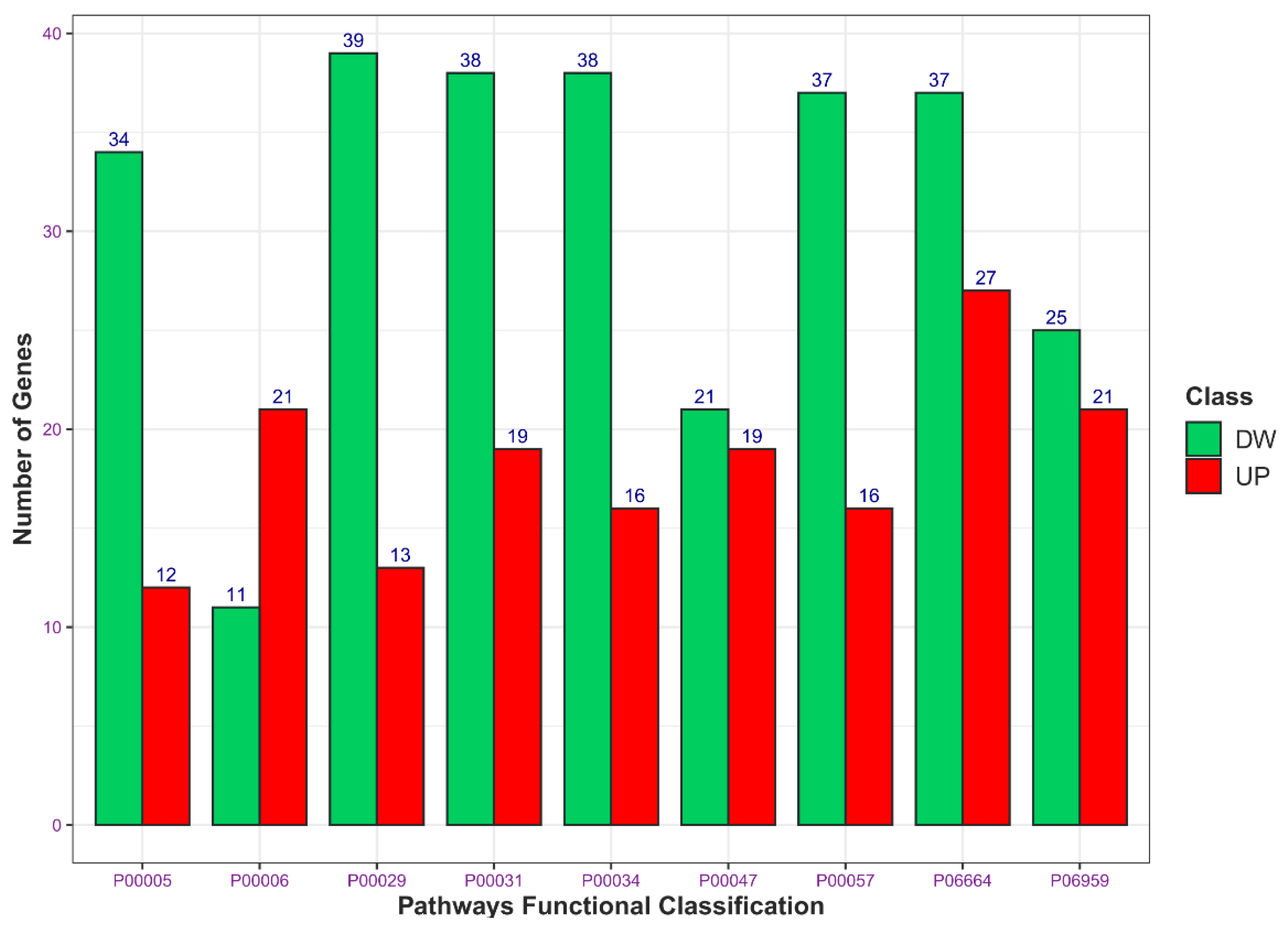
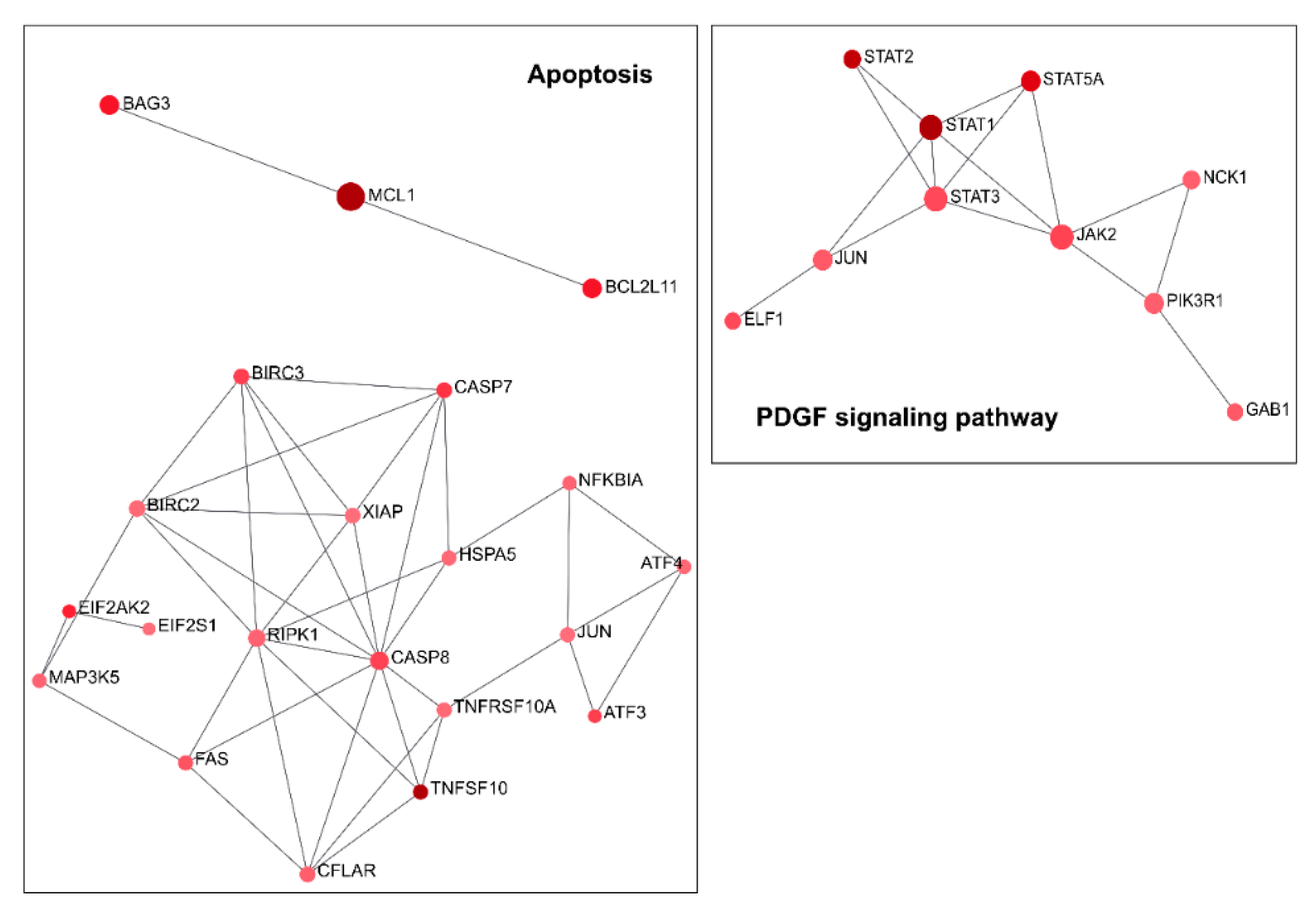
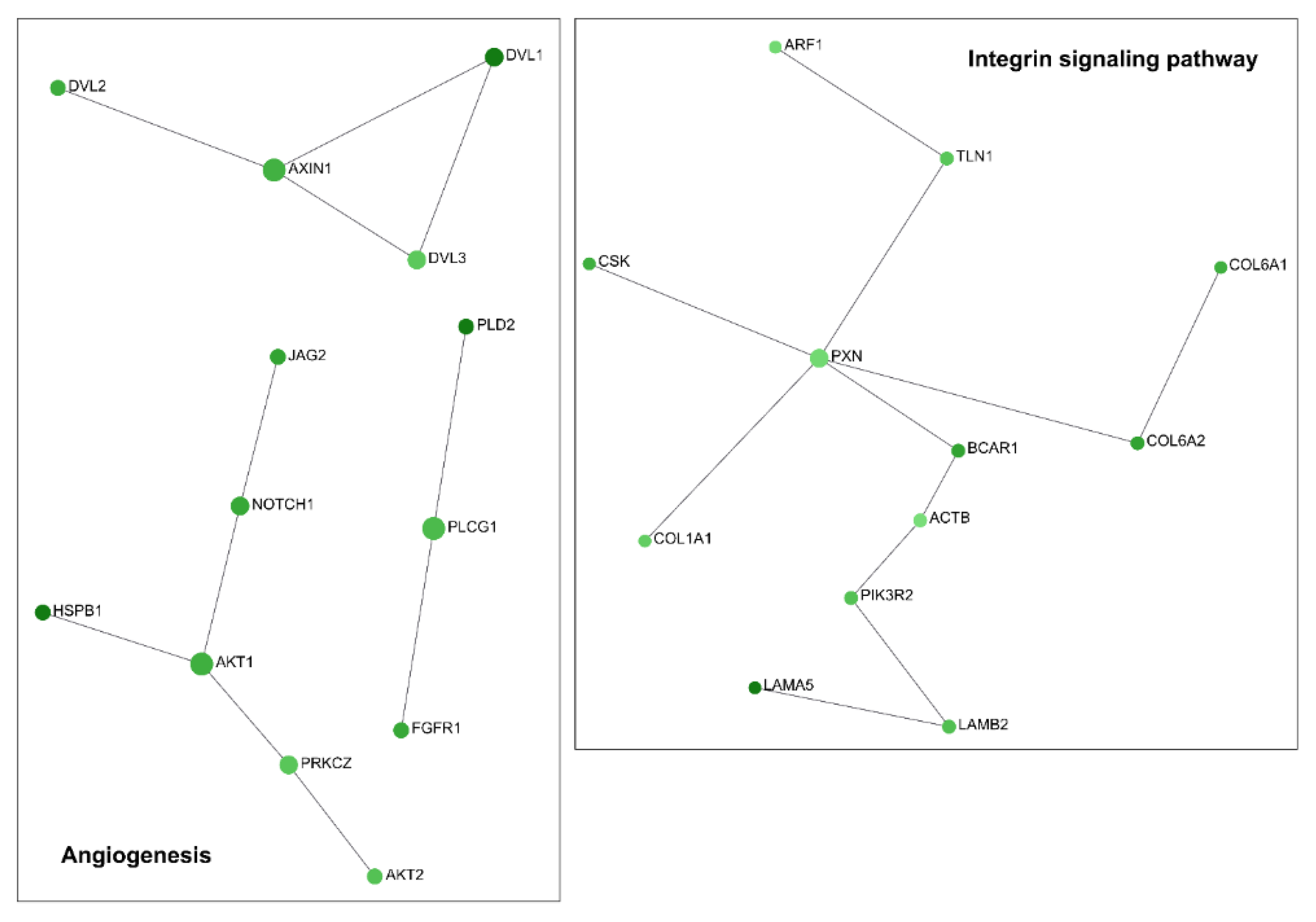
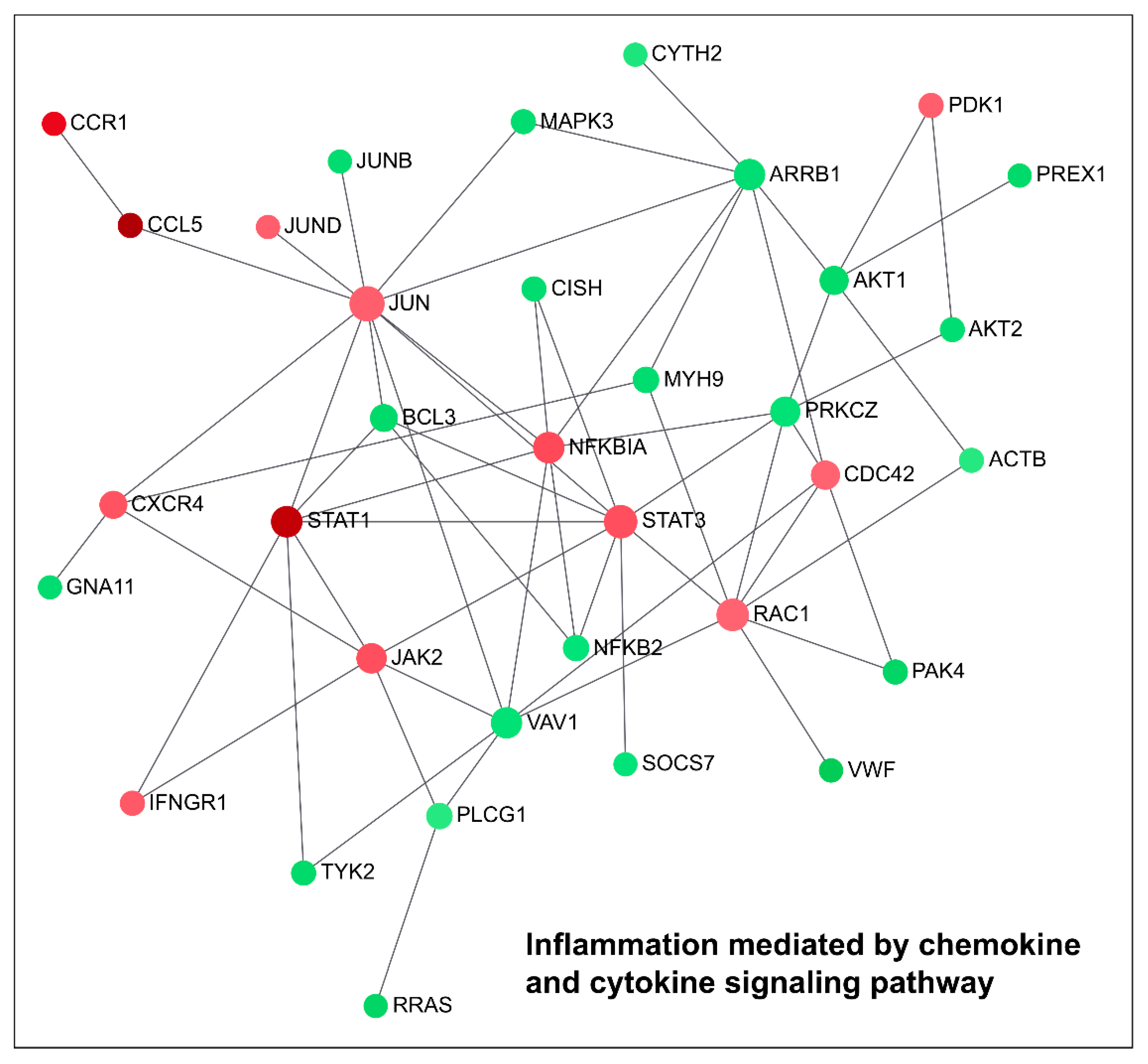
2.1. Downregulation of TG2-lncRNA Using a siRNA Molecule Changes Specific Pathways in MCF-7 Cancer Cells
2.2. Consensus Sequences Shared between TG2-lncRNA and Promoters of TG2-lncRNA Targeted Genes
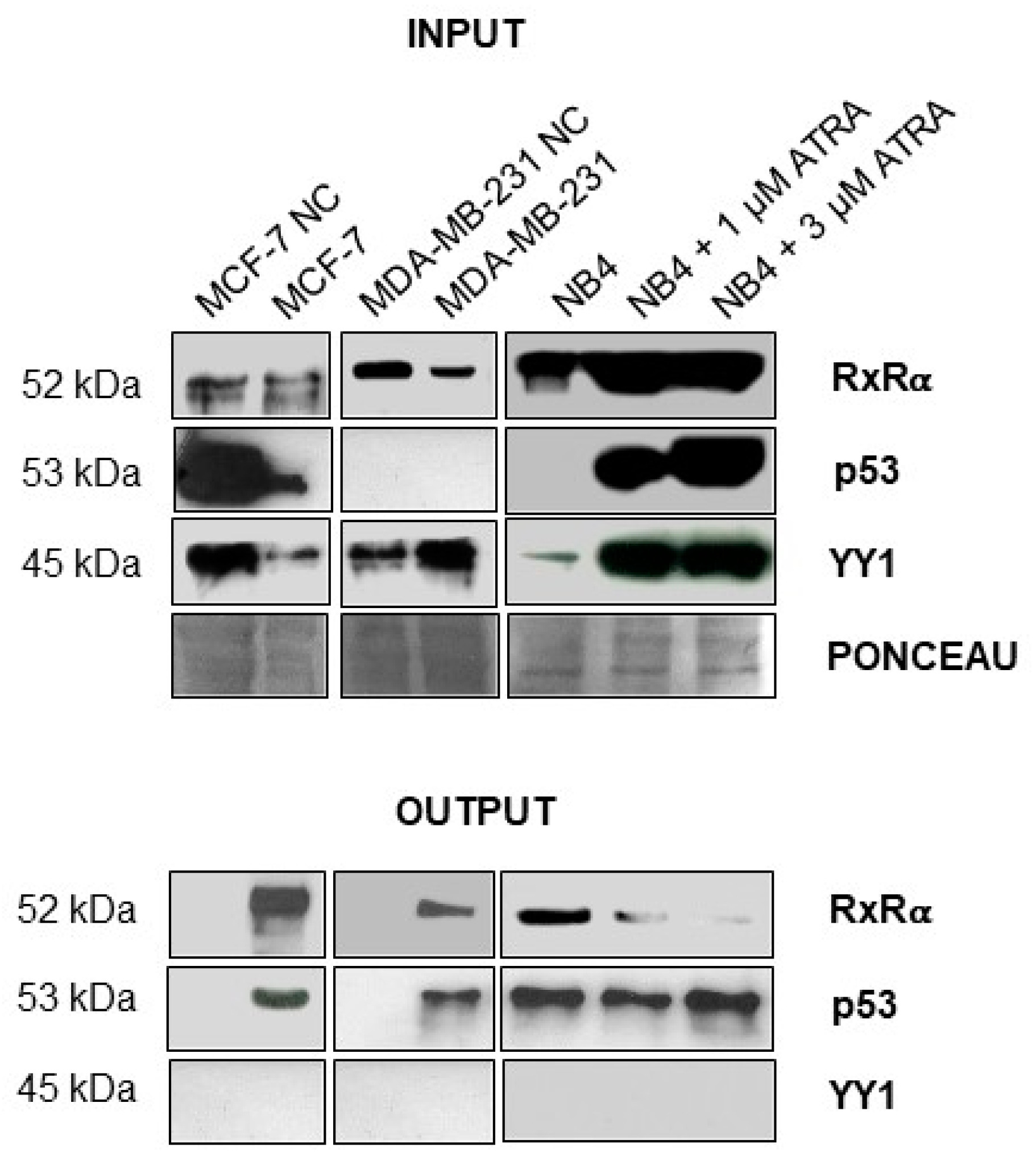
2.3. A Biotinylated Single Strand DNA Mimic of TG2-lncRNA Binds RXRα and TP53 in Nuclear Extracts of MCF-7 Cells
3. Discussion
4. Material and Methods
4.1. Culture and Transfection of MCF-7 Cells Using siRNA Molecules
4.2. RNA Extraction, Reverse Transcription and Quantitative PCR
4.3. RNA-Sequencing and Bioinformatic Analysis
4.4. In Vitro Assay for Binding of Nuclear Extracts by Biotinylated Single Stranded DNA Mimic of TG2-lncRNA
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATRA | All-trans retinoic acid |
| ACTB | Beta-actin |
| BrCa | Breast cancer |
| CCKR | Cholecystokinin receptor |
| DEGs | Differential Gene Expression |
| ECM | Extracellular matrix |
| EMT | Epithelial–mesenchymal transition |
| FBS | foetal bovine serum |
| FPKM | number of Fragments Per Kilobase of transcript per Million mapped reads |
| GO | Gene ontology |
| HPRT1 | Hypoxanthine Phosphoribosyltransferase 1 |
| JUN | Transcription factor AP-1 |
| lncRNAs | Long non-coding RNAs |
| Log2FC | Log2 Fold Change |
| NFĸBIA | NF-ĸB inhibitor alpha |
| nt | nucleotides |
| PCA | Principal Component Analysis |
| PDGF | Platelet-derived growth factor |
| PDK1 | 3-phosphoinositide-dependent protein kinase 1 |
| PXN | Paxillin |
| RIP | RNA immunoprecipitation |
| RNA-seq | RNA sequencing |
| RT-qPCR | Reverse Transcription and quantitative Polymerase Chain Reaction |
| RXRα | Retinoid X receptor alpha |
| siRNA | short interfering RNA |
| siRNA neg | siRNA negative control |
| TG2 | Transglutaminase type 2 |
| TG2-lncRNA | Long non-coding RNA of Transglutaminase type 2 gene |
| TGM2 | Transglutaminase type 2 gene |
| YY1 | Ying-Yang 1 |
References
- Phatak, V.M.; Croft, S.M.; Rameshaiah Setty, S.G.; Scarpellini, A.; Hughes, D.C.; Rees, R.; McArdle, S.; Verderio, E.A. Expression of transglutaminase-2 isoforms in normal human tissues and cancer cell lines: Dysregulation of alternative splicing in cancer. Amino Acids 2013, 44, 33–44. [Google Scholar] [CrossRef]
- Bianchi, N.; Beninati, S.; Bergamini, C.M. Spotlight on the transglutaminase 2 gene: A focus on genomic and transcriptional aspects. Biochem. J. 2018, 475, 1643–1667. [Google Scholar] [CrossRef]
- Eckert, R.L.; Kaartinen, M.T.; Nurminskaya, M.; Belkin, A.M.; Colak, G.; Johnson, G.V.W.; Mehta, K. Transglutaminase regulation of cell function. Physiol. Rev. 2013, 94, 383–417. [Google Scholar] [CrossRef] [Green Version]
- Griffin, M.; Casadio, R.; Bergamini, C.M. Transglutaminases: Nature’s biological glues. Biochem. J. 2002, 368, 377–396. [Google Scholar] [CrossRef] [Green Version]
- Franzese, O.; Minotti, L.; Aguiari, G.; Corrà, F.; Cervellati, C.; Ferrari, C.; Volinia, S.; Bergamini, C.M.; Bianchi, N. Involvement of non-coding RNAs and transcription factors in the induction of Transglutaminase isoforms by ATRA. Amino Acids 2019, 51, 1273–1288. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Geng, Y.; Feng, R.; Zhu, Q.; Miao, B.; Cao, J.; Fei, S. The Human RNA Surveillance Factor UPF1 modulates gastric cancer progression by targeting long non-coding RNA MALAT1. Cell. Physiol. Biochem. 2017, 42, 2194–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minotti, L.; Baldassari, F.; Galasso, M.; Volinia, S.; Bergamini, C.M.; Bianchi, N. A long non-coding RNA inside the type 2 transglutaminase gene tightly correlates with the expression of its transcriptional variants. Amino Acids 2018, 50, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Huynh, T.T.; Sultan, M.; Vidovic, D.; Dean, C.A.; Cruickshank, B.M.; Lee, K.; Loung, C.Y.; Holloway, R.W.; Hoskin, D.W.; Waisman, D.M.; et al. Retinoic acid and arsenic trioxide induce lasting differentiation and demethylation of target genes in APL cells. Sci. Rep. 2019, 9, 9414–9427. [Google Scholar] [CrossRef]
- Hettasch, J.M.; Bandarenko, N.; Burchette, J.L.; Lai, T.S.; Marks, J.R.; Haroon, Z.A.; Peters, K.; Dewhirst, M.W.; Iglehart, J.D.; Greenberg, C.S. Tissue transglutaminase expression in human breast cancer. Lab. Investig. 1996, 75, 637–645. [Google Scholar] [PubMed]
- Assi, J.; Srivastava, G.; Matta, A.; Chang, M.C.; Walfish, P.G.; Ralhan, R. Transglutaminase 2 overexpression in tumor stroma identifies invasive ductal carcinomas of breast at high risk of recurrence. PLoS ONE 2013, 8, e74437. [Google Scholar] [CrossRef]
- Ai, L.; Kim, W.J.; Demircan, B.; Dyer, L.M.; Bray, K.J.; Skehan, R.R.; Massoll, N.A.; Brown, K.D. The transglutaminase 2 gene (TGM2), a potential molecular marker for chemotherapeutic drug sensitivity, is epigenetically silenced in breast cancer. Carcinogenesis 2008, 29, 510–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Gao, H.; Xu, J.; Reuben, J.; Yu, D.; Mehta, K. Evidence that aberrant expression of tissue transglutaminase promotes stem cell characteristics in mammary epithelial cells. PLoS ONE 2011, 6, e20701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Huang, W. Compensatory increase of Transglutaminase 2 is responsible for resistance to mTOR inhibitor treatment. PLoS ONE 2016, 11, e0149388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, C.; Szmacinski, H.; Fisher, M.L.; Nance, B.; Lakowicz, J.R.; Akbar, A.; Keillor, J.W.; Lok Wong, T.; Godoy-Ruiz, R.; Toth, E.A.; et al. Transamidase site-targeted agents alter the conformation of the transglutaminase cancer stem cell survival protein to reduce GTP binding activity and cancer stem cell survival. Oncogene 2017, 36, 2981–2990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckert, R.L. Transglutaminase 2 takes center stage as a cancer cell survival factor and therapy target. Mol. Carcinog. 2019, 58, 837–853. [Google Scholar] [CrossRef] [PubMed]
- Taccioli, C.; Wan, S.G.; Liu, C.G.; Alder, H.; Volinia, S.; Farber, J.L.; Croce, C.M.; Fong, L.Y.Y. Zinc replenishment reverses overexpression of the proinflammatory mediator S100A8 and esophageal preneoplasia in the rat. Gastroenterology 2009, 136, 953–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanoa, J.K.; Sethi, R.S.; Verma, R.; Arora, J.S.; Mukhopadhyay, C.S. Long non-coding RNA: Its evolutionary relics and biological implications in mammals: A review. J. Anim. Sci. Technol. 2018, 60, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Sharma, S.; Watabe, K. Roles of lncRNA in breast cancer. Front. Biosci. (Sch. Ed.) 2015, 7, 94–108. [Google Scholar]
- Tomar, D.; Yadav, A.S.; Kumar, D.; Bhadauriya, G.; Kundu, G.C. Non-coding RNAs as potential therapeutic targets in breast cancer. Biochim. Biophys. Acta Gene Regul. Mech. 2020, 1863, 194378–194399. [Google Scholar] [CrossRef]
- Kumar, A.; Xu, J.; Sung, B.; Kumar, S.; Yu, D.; Aggarwal, B.B.; Mehta, K. Evidence that GTP-binding domain but not catalytic domain of transglutaminase 2 is essential for epithelial-to-mesenchymal transition in mammary epithelial cells. Breast Cancer Res. 2012, 14, R4. [Google Scholar] [CrossRef] [Green Version]
- Karicheva, O.; Rodriguez-Vargas, J.M.; Wadier, N.; Martin-Hernandez, K.; Vauchelles, R.; Magroun, N.; Tissier, A.; Schreiber, V.; Dantzer, F. PARP3 controls TGFβ and ROS driven epithelial-to-mesenchymal transition and stemness by stimulating a TG2-Snail-E-cadherin axis. Oncotarget 2016, 7, 64109–64123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, R.; Khosla, C. Substrates, inhibitors, and probes of mammalian transglutaminase 2. Anal. Biochem. 2020, 591, 113560–113565. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Xu, J.; Brady, S.; Gao, H.; Yu, D.; Reuben, J.; Mehta, K. Tissue transglutaminase promotes drug resistance and invasion by inducing mesenchymal transition in mammary epithelial cells. PLoS ONE 2010, 5, e13390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lennox, K.A.; Behlke, M.A. Cellular localization of long non-coding RNAs affects silencing by RNAi more than by antisense oligonucleotides. Nucleic Acids Res. 2016, 44, 863–877. [Google Scholar] [CrossRef] [Green Version]
- Stojic, L.; Niemczyk, M.; Orjalo, A.; Ito, Y.; Ruijter, A.E.; Uribe-Lewis, S.; Joseph, N.; Weston, S.; Menon, S.; Odom, D.T.; et al. Transcriptional silencing of long noncoding RNA GNG12-AS1 uncouples its transcriptional and product-related functions. Nat. Commun. 2016, 7, 10406–10420. [Google Scholar] [CrossRef]
- Kim, D.S.; Park, S.S.; Nam, B.H.; Kim, I.H.; Kim, S.Y. Reversal of drug resistance in breast cancer cells by transglutaminase 2 inhibition and nuclear factor-kappaB inactivation. Cancer Res. 2006, 66, 10936–10943. [Google Scholar] [CrossRef] [Green Version]
- Devarajan, E.; Song, Y.H.; Krishnappa, S.; Alt, E. Epithelial-mesenchymal transition in breast cancer lines is mediated through PDGF-D released by tissue-resident stem cells. Int. J. Cancer 2012, 131, 1023–1031. [Google Scholar] [CrossRef]
- Guo, W.; Giancotti, F.G. Integrin signalling during tumour progression. Nat. Rev. Mol. Cell Biol. 2004, 5, 816–826. [Google Scholar] [CrossRef]
- Kumbrink, J.; Kirsch, K.H. Regulation of p130Cas/BCAR1 Expression in tamoxifen-sensitive and tamoxifen-resistant breast cancer cells by EGR1 and NAB2. Neoplasia 2012, 14, 108–120. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Srivastava, S.K. Inhibition of HER2-integrin signaling by Cucurbitacin B leads to in vitro and in vivo breast tumor growth suppression. Oncotarget 2014, 5, 1812–1828. [Google Scholar] [CrossRef] [Green Version]
- Ashaie, M.A.; Islam, R.A.; Kamaruzman, N.I.; Ibnat, N.; Tha, K.K.; Chowdhury, E.H. Targeting cell adhesion molecules via carbonate apatite-mediated delivery of specific siRNAs to breast cancer cells in vitro and in vivo. Pharmaceutics 2019, 11, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khurana, S.; Chakraborty, S.; Cheng, X.; Su, Y.T.; Kao, H.Y. The actin-binding protein, actinin alpha 4 (ACTN4), is a nuclear receptor coactivator that promotes proliferation of MCF-7 breast cancer cells. J. Biol. Chem. 2011, 286, 1850–1859. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Ge, N.; Wang, N.; Sun, W.; Mao, R.; Bu, W.; Creighton, C.J.; Zheng, P.; Vasudevan, S.; An, L.; et al. Amplification and over-expression of MAP3K3 gene in human breast cancer promotes formation and survival of breast cancer cells. J. Pathol. 2014, 232, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Vadlamudi, R.; Adam, L.; Tseng, B.; Costa, L.; Kumar, R. Transcriptional up-regulation of Paxillin expression by heregulin in human breast cancer cells. Cancer Res. 1999, 59, 2843–2846. [Google Scholar]
- Pardo, O.E.; Castellano, L.; Munro, C.E.; Hu, Y.; Mauri, F.; Krell, J.; Lara, R.; Pinho, F.G.; Choudhury, T.; Frampton, A.E.; et al. MiR-515-5p controls cancer cell migration through MARK4 regulation. EMBO Rep. 2016, 17, 570–584. [Google Scholar] [CrossRef] [Green Version]
- Fesus, L.; Piacentini, M. Transglutaminase 2: An enigmatic enzyme with diverse functions. Trends Biochem. Sci. 2002, 27, 534–539. [Google Scholar] [CrossRef]
- Karam, M.; Legay, C.; Auclair, C.; Ricort, J.M. Protein kinase D1 stimulates proliferation and enhances tumorigenesis of MCF-7 human breast cancer cells through a MEK/ERK-dependent signaling pathway. Exp. Cell Res. 2012, 318, 558–569. [Google Scholar] [CrossRef]
- Cayado-Gutiérrez, N.; Moncalero, V.L.; Rosales, E.M.; Berón, W.; Salvatierra, E.E.; Alvarez-Olmedo, D.; Radrizzani, M.; Ciocca, D.R. Downregulation of Hsp27 (HSPB1) in MCF-7 human breast cancer cells induces upregulation of PTEN. Cell Stress Chaperon. 2013, 18, 243–249. [Google Scholar] [CrossRef]
- Zenga, B.; Liab, Y.; Fengab, Y.; Lua, M.; Yuana, H.; Yiab, Z.; Wua, Y.; Xianga, T.; Liab, H.; Renab, G. Downregulated miR-1247-5p associates with poor prognosis and facilitates tumor cell growth via DVL1/Wnt/β-catenin signaling in breast cancer. Biochem. Biophys. Res. Commun. 2018, 505, 302–308. [Google Scholar] [CrossRef]
- Engel, N.; Lisec, J.; Piechulla, B.; Nebe, B. Metabolic profiling reveals sphingosine-1-phosphate kinase 2 and lyase as key targets of (phyto-) estrogen action in the breast cancer cell line MCF-7 and not in MCF-12A. PLoS ONE 2012, 7, e47833. [Google Scholar] [CrossRef]
- Petanidis, S.; Hadzopoulou-Cladaras, M.; Salifoglou, A. Cadmium modulates H-ras expression and caspase-3 apoptotic cell death in breast cancer epithelial MCF-7 cells. J. Inorg. Biochem. 2013, 121, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Kiesel, V.A.; Stan, S.D. Diallyl trisulfide, a chemopreventive agent from Allium vegetables. inhibits alpha-secretases in breast cancer cells. Biochem. Biophys. Res. Commun. 2017, 84, 833–838. [Google Scholar] [CrossRef]
- Yun, J.; Espinoza, I.; Pannuti, A.; Romero, A.; Martinez, L.; Caskey, M.; Stanculescu, A.; Bocchetta, M.; Rizzo, P.; Band, V.; et al. TP53 Modulates Notch signaling in MCF-7 breast cancer cells by associating with the Notch transcriptional complex Via MAML1. J. Cell. Physiol. 2015, 230, 3115–3127. [Google Scholar] [CrossRef] [Green Version]
- Beaver, J.A.; Gustin, J.P.; Yi, K.H.; Rajpurohit, A.; Thomas, M.; Gilbert, S.F.; Rosen, D.M.; Park, B.H.; Lauring, J. PIK3CA and AKT1 mutations have distinct effects on sensitivity to targeted pathway inhibitors in an isogenic luminal breast cancer model system. Clin. Cancer Res. 2013, 19, 5413–5422. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Shen, T.; Liu, J.; Zheng, J.; Zhang, Y.; Xu, R.; Sun, C.; Du, J.; Chen, Y.; Gu, L. Rab35 is required for Wnt5a/Dvl2-induced Rac1 activation and cell migration in MCF-7 breast cancer cells. Cell. Signal. 2013, 25, 1075–1085. [Google Scholar] [CrossRef]
- Ganesan, R.; Mallets, E.; Gomez-Cambronero, J. The transcription factors Slug (SNAI2) and Snail (SNAI1) regulate phospholipase D (PLD) promoter in opposite ways towards cancer cell invasion. Mol. Oncol. 2016, 10, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Bai, T.; Luoh, S.W. GRB-7 facilitates HER-2/Neu-mediated signal transduction and tumor formation. Carcinogenesis 2008, 29, 473–479. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhao, Y.; Li, L.; Shen, Y.; Cai, X.; Zhang, X.; Ye, X. The oncoprotein HBXIP upregulates PDGFB via activating transcription factor Sp1 to promote the proliferation of breast cancer cells. Biochem. Biophys. Res. Commun. 2013, 434, 305–310. [Google Scholar] [CrossRef]
- Ma, F.; Zhang, L.; Ma, L.; Zhang, Y.; Zhang, J.; Guo, B. MiR-361-5p inhibits glycolytic metabolism. proliferation and invasion of breast cancer by targeting FGFR1 and MMP-1. J. Exp. Clin. Cancer Res. 2017, 36, 158–170. [Google Scholar] [CrossRef] [Green Version]
- Drzewiecka, H.; Jagodzinski, P.P. Trichostatin A reduced phospholipase C gamma-1 transcript and protein contents in MCF-7 breast cancer cells. Biomed. Pharmacother. 2012, 66, 1–5. [Google Scholar] [CrossRef]
- Park, K.S.; Kim, D.S.; Jeong, K.C.; Kim, S.Y. Increase in Transglutaminase 2 expression is associated with NF-kappaB activation in breast cancer tissues. Front. Biosci. (Landmark Ed.) 2009, 14, 1945–1951. [Google Scholar] [CrossRef] [Green Version]
- Croci, S.; Nanni, P.; Palladini, A.; Nicoletti, G.; Grosso, V.; Benegiamo, G.; Landuzzi, L.; Lamolinara, A.; Ianzano, M.L.; Ranieri, D.; et al. Interleukin-15 is required for immunosurveillance and immunoprevention of HER2/neu-driven mammary carcinogenesis. Breast Cancer Res. 2015, 17, 70–83. [Google Scholar] [CrossRef] [Green Version]
- Oh, K.; Lee, O.Y.; Park, Y.; Seo, M.W.; Lee, D.S. IL-1β induces IL-6 production and increases invasiveness and estrogen-independent growth in a TG2-dependent manner in human breast cancer cells. BMC Cancer 2016, 16, 724–735. [Google Scholar] [CrossRef] [Green Version]
- Oh, K.; Ko, E.; Kim, H.S.; Park, A.K.; Moon, H.G.; Noh, D.Y.; Lee, D.S. Transglutaminase 2 facilitates the distant hematogenous metastasis of breast cancer by modulating interleukin-6 in cancer cells. Breast Cancer Res. 2011, 13, R96. [Google Scholar] [CrossRef] [Green Version]
- Zlatanova, J.; Caiafa, P. CTCF and its protein partners: Divide and rule? J. Cell Sci. 2009, 122, 1275–1284. [Google Scholar] [CrossRef] [Green Version]
- Beninati, S.; Piacentini, M.; Bergamini, C.M. Transglutaminase 2, a double face enzyme. Amino Acids 2017, 49, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Zhang, S.; Yazdanparast, A. The Expression Patterns of ER, PR, HER2, CK5/6, EGFR, Ki-67 and AR by Immunohistochemical Analysis in Breast Cancer Cell Lines. BMC Genom. 2016, 17, 525–546. [Google Scholar] [CrossRef] [Green Version]
- de Kok, J.B.; Roelofs, R.W.; Giesendorf, B.A.; Pennings, J.L.; Waas, E.T.; Feuth, T.; Swinkels, D.W.; Span, P.N. Normalization of gene expression measurements in tumor tissues: Comparison of 13 endogenous control genes. Lab. Investig. 2005, 85, 154–159. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Yalamanchili, H.K.; Wan, Y.W.; Liu, Z. Data analysis pipeline for RNA-seq experiments: From differential expression to cryptic splicing. Curr. Protoc. Bioinform. 2017, 59, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Institute: Cambridge, UK, 2010; Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 13 August 2021).
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.M.; Ruan, J.M.; Homer, N.; Marth, G.; Abecasis, G.; Durbi, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550–571. [Google Scholar] [CrossRef] [Green Version]
- Mi, H.; Thomas, P. Protein networks and pathway analysis. Methods Mol. Biol. 2009, 563, 123–140. [Google Scholar] [CrossRef]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A visual analytics platform for comprehensive gene expression profiling and meta-analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergamini, C.M.; Vischioni, C.; Aguiari, G.; Grandi, C.; Terrazzan, A.; Volinia, S.; Bianchi, N.; Taccioli, C. Inhibition of the lncRNA Coded within Transglutaminase 2 Gene Impacts Several Relevant Networks in MCF-7 Breast Cancer Cells. Non-Coding RNA 2021, 7, 49. https://doi.org/10.3390/ncrna7030049
Bergamini CM, Vischioni C, Aguiari G, Grandi C, Terrazzan A, Volinia S, Bianchi N, Taccioli C. Inhibition of the lncRNA Coded within Transglutaminase 2 Gene Impacts Several Relevant Networks in MCF-7 Breast Cancer Cells. Non-Coding RNA. 2021; 7(3):49. https://doi.org/10.3390/ncrna7030049
Chicago/Turabian StyleBergamini, Carlo M., Chiara Vischioni, Gianluca Aguiari, Carmen Grandi, Anna Terrazzan, Stefano Volinia, Nicoletta Bianchi, and Cristian Taccioli. 2021. "Inhibition of the lncRNA Coded within Transglutaminase 2 Gene Impacts Several Relevant Networks in MCF-7 Breast Cancer Cells" Non-Coding RNA 7, no. 3: 49. https://doi.org/10.3390/ncrna7030049
APA StyleBergamini, C. M., Vischioni, C., Aguiari, G., Grandi, C., Terrazzan, A., Volinia, S., Bianchi, N., & Taccioli, C. (2021). Inhibition of the lncRNA Coded within Transglutaminase 2 Gene Impacts Several Relevant Networks in MCF-7 Breast Cancer Cells. Non-Coding RNA, 7(3), 49. https://doi.org/10.3390/ncrna7030049