
Non-Coding RNAs in COVID-19: Emerging Insights and Current Questions



Abstract
:1. SARS-CoV-2 and COVID-19
2. The Human Non-Coding Genome and COVID-19
3. ncRNAs in Cytokine Storm Syndrome, a Hallmark of Severe COVID-19
3.1. COVID-19 and Cytokine Storm Syndrome
3.2. COVID-19 Loss of miR-1275 and miR-766-3p may Promote an IL-6 Hyperinflammatory State
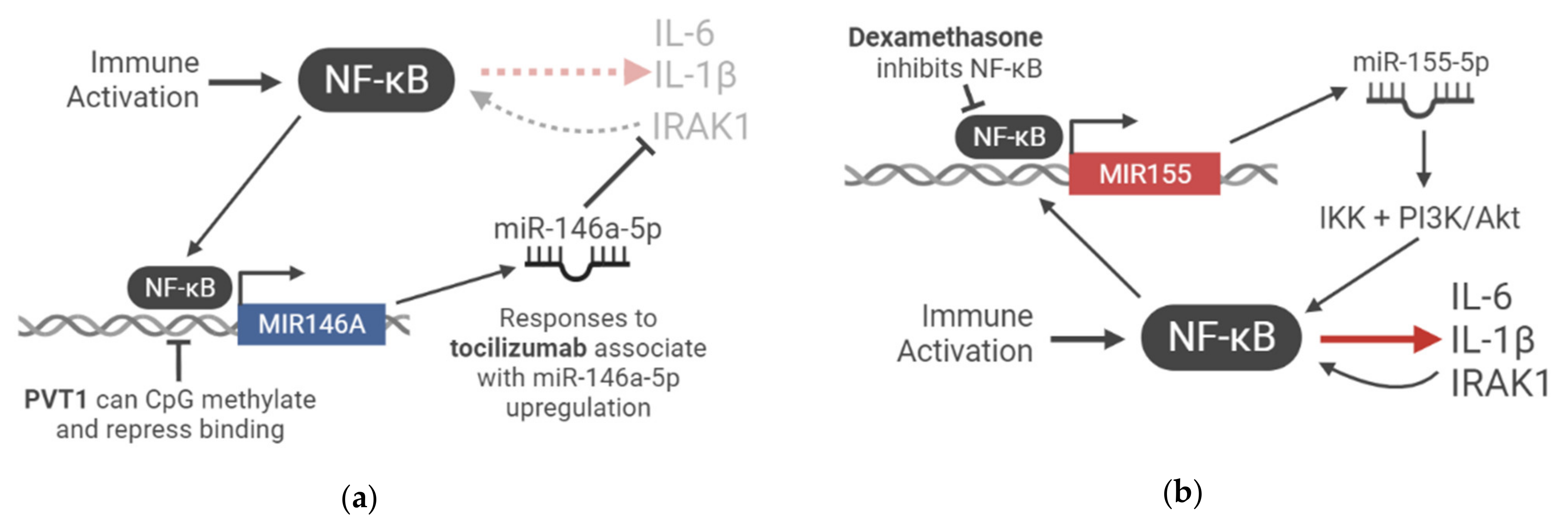
3.3. miR-146a-5p Expression Is a Determinant of Response to Therapeutic IL-6R Blockade
3.4. Upregulation of Inflammatory miR-155 May Be Combatted by Glucocorticoid Treatment
4. ncRNAs in Immune Cell Recruitment and Differentiation, a Precursor to Damage
4.1. The Infiltration of Immune Cells Is Associated with Worsened COVID-19
4.2. Differential miR-31-5p Expression May Impact Immune Cell Recruitment
4.3. MALAT1 Expression Is Altered in SARS-CoV-2 Infection with Inflammatory Implications
4.4. miRNA-Related Endothelial Dysfunction in the Lung Microenvironment
5. ncRNAs in COVID-19-Associated Haemostatic Dysregulation
5.1. Venous Thromboembolic Events in COVID-19
5.2. COVID-19 Patients with High Coagulability Have Deregulated Inflammatory miRNAs
5.3. A High D-Dimer Specific miRNA Predicts Thromboembolic Events
6. ncRNAs in COVID-19-Associated Vascular Dysregulation
6.1. miRNAs and Cardiovascular Disease in COVID-19
6.2. miRNAs and Cerebrovascular Disease in COVID-19
7. Interactions between Host and SARS-CoV-2-Derived ncRNAs
7.1. The SARS-CoV-2 RNA Genome as a Non-Coding Interactor with Host RNA
7.2. Differences in miRNA Targeting May Explain Divergent SARS-CoV-2 Virology
7.3. Host and Viral RNA-Binding Proteins and ncRNAs as Regulators of Infection
8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus Biology and Replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Hamming, I.; Timens, W.; Bulthuis, M.; Lely, A.; Navis, G.; van Goor, H. Tissue Distribution of ACE2 Protein, the Functional Receptor for SARS Coronavirus. A First Step in Understanding SARS Pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Chen, L.; Li, X.; Chen, M.; Feng, Y.; Xiong, C. The ACE2 Expression in Human Heart Indicates New Potential Mechanism of Heart Injury among Patients Infected with SARS-CoV-2. Cardiovasc. Res. 2020, 116, 1097–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmugam, C.; Mohammed, A.R.; Ravuri, S.; Luthra, V.; Rajagopal, N.; Karre, S. COVID-2019—A Comprehensive Pathology Insight. Pathol. Res. Pract. 2020, 216, 153222. [Google Scholar] [CrossRef] [PubMed]
- Petersen, E.; Koopmans, M.; Go, U.; Hamer, D.H.; Petrosillo, N.; Castelli, F.; Storgaard, M.; Al Khalili, S.; Simonsen, L. Comparing SARS-CoV-2 with SARS-CoV and Influenza Pandemics. Lancet Infect. Dis. 2020, 20, e238–e244. [Google Scholar] [CrossRef]
- Wu, J.T.; Leung, K.; Bushman, M.; Kishore, N.; Niehus, R.; de Salazar, P.M.; Cowling, B.J.; Lipsitch, M.; Leung, G.M. Estimating Clinical Severity of COVID-19 from the Transmission Dynamics in Wuhan, China. Nat. Med. 2020, 26, 506–510. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, N.; Laydon, D.; Nedjati Gilani, G.; Imai, N.; Ainslie, K.; Baguelin, M.; Bhatia, S.; Boonyasiri, A.; Cucunuba Perez, Z.; Cuomo-Dannenburg, G.; et al. Report 9: Impact of Non-Pharmaceutical Interventions (NPIs) to Reduce COVID19 Mortality and Healthcare Demand; Imperial College: London, UK, 2020. [Google Scholar]
- Holman, N.; Knighton, P.; Kar, P.; O’Keefe, J.; Curley, M.; Weaver, A.; Barron, E.; Bakhai, C.; Khunti, K.; Wareham, N.J.; et al. Risk Factors for COVID-19-Related Mortality in People with Type 1 and Type 2 Diabetes in England: A Population-Based Cohort Study. Lancet Diabetes Endocrinol. 2020, 8, 823–833. [Google Scholar] [CrossRef]
- Jordan, R.E.; Adab, P.; Cheng, K.K. Covid-19: Risk Factors for Severe Disease and Death. BMJ 2020, 368, m1198. [Google Scholar] [CrossRef] [Green Version]
- Mattick, J.S. RNA Regulation: A New Genetics? Nat. Rev. Genet. 2004, 5, 316–323. [Google Scholar] [CrossRef]
- The FANTOM Consortium. The Transcriptional Landscape of the Mammalian Genome. Science 2005, 309, 1559–1563. [Google Scholar] [CrossRef] [Green Version]
- Brockdorff, N.; Ashworth, A.; Kay, G.F.; McCabe, V.M.; Norris, D.P.; Cooper, P.J.; Swift, S.; Rastan, S. The Product of the Mouse Xist Gene Is a 15 Kb Inactive X-Specific Transcript Containing No Conserved ORF and Located in the Nucleus. Cell 1992, 71, 515–526. [Google Scholar] [CrossRef]
- Ridings-Figueroa, R.; Stewart, E.R.; Nesterova, T.B.; Coker, H.; Pintacuda, G.; Godwin, J.; Wilson, R.; Haslam, A.; Lilley, F.; Ruigrok, R.; et al. The Nuclear Matrix Protein CIZ1 Facilitates Localization of Xist RNA to the Inactive X-Chromosome Territory. Genes Dev. 2017, 31, 876–888. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Arun, G.; Mao, Y.S.; Lazar, Z.; Hung, G.; Bhattacharjee, G.; Xiao, X.; Booth, C.J.; Wu, J.; Zhang, C.; et al. The LncRNA Malat1 Is Dispensable for Mouse Development but Its Transcription Plays a Cis-Regulatory Role in the Adult. Cell Rep. 2012, 2, 111–123. [Google Scholar] [CrossRef] [Green Version]
- Hewitson, J.P.; West, K.A.; James, K.R.; Rani, G.F.; Dey, N.; Romano, A.; Brown, N.; Teichmann, S.A.; Kaye, P.M.; Lagos, D. Malat1 Suppresses Immunity to Infection through Promoting Expression of Maf and IL-10 in Th Cells. J. Immunol. 2020, 204, 2949–2960. [Google Scholar] [CrossRef] [Green Version]
- Shimakami, T.; Yamane, D.; Jangra, R.K.; Kempf, B.J.; Spaniel, C.; Barton, D.J.; Lemon, S.M. Stabilization of Hepatitis C Virus RNA by an Ago2-MiR-122 Complex. Proc. Natl. Acad. Sci. USA 2012, 109, 941–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farr, R.J.; Rootes, C.L.; Rowntree, L.C.; Nguyen, T.H.O.; Hensen, L.; Kedzierski, L.; Cheng, A.C.; Kedzierska, K.; Au, G.G.; Marsh, G.A.; et al. Altered MicroRNA Expression in COVID-19 Patients Enables Identification of SARS-CoV-2 Infection. PLoS Pathog. 2021, 17, e1009759. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, A. Tocilizumab in Rheumatoid Arthritis: Efficacy, Safety and Its Place in Therapy. Ther. Adv. Chronic Dis. 2013, 4, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayakawa, K.; Kawasaki, M.; Hirai, T.; Yoshida, Y.; Tsushima, H.; Fujishiro, M.; Ikeda, K.; Morimoto, S.; Takamori, K.; Sekigawa, I. MicroRNA-766-3p Contributes to Anti-Inflammatory Responses through the Indirect Inhibition of NF-ΚB Signaling. IJMS 2019, 20, 809. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Meisgen, F.; Butler, L.M.; Han, G.; Wang, X.-J.; Söderberg-Nauclér, C.; Ståhle, M.; Pivarcsi, A.; Sonkoly, E. MicroRNA-31 is Overexpressed in Psoriasis and Modulates Inflammatory Cytokine and Chemokine Production in Keratinocytes via Targeting Serine/Threonine Kinase 40. J. Immunol. 2013, 190, 678–688. [Google Scholar] [CrossRef] [Green Version]
- Mi, B.; Li, Q.; Li, T.; Liu, G.; Sai, J. High MiR-31-5p Expression Promotes Colon Adenocarcinoma Progression by Targeting TNS1. Aging 2020, 12, 7480–7490. [Google Scholar] [CrossRef]
- Tian, Y.; Xu, J.; Li, Y.; Zhao, R.; Du, S.; Lv, C.; Wu, W.; Liu, R.; Sheng, X.; Song, Y.; et al. MicroRNA-31 Reduces Inflammatory Signaling and Promotes Regeneration in Colon Epithelium, and Delivery of Mimics in Microspheres Reduces Colitis in Mice. Gastroenterology 2019, 156, 2281–2296.e6. [Google Scholar] [CrossRef]
- Sabbatinelli, J.; Giuliani, A.; Matacchione, G.; Latini, S.; Laprovitera, N.; Pomponio, G.; Ferrarini, A.; Svegliati Baroni, S.; Pavani, M.; Moretti, M.; et al. Decreased Serum Levels of the Inflammaging Marker MiR-146a Are Associated with Clinical Non-Response to Tocilizumab in COVID-19 Patients. Mech. Ageing Dev. 2021, 193, 111413. [Google Scholar] [CrossRef] [PubMed]
- Ye, E.-A.; Steinle, J.J. MiR-146a Suppresses STAT3/VEGF Pathways and Reduces Apoptosis through IL-6 Signaling in Primary Human Retinal Microvascular Endothelial Cells in High Glucose Conditions. Vis. Res. 2017, 139, 15–22. [Google Scholar] [CrossRef]
- Jiang, S.; Hu, Y.; Deng, S.; Deng, J.; Yu, X.; Huang, G.; Kawai, T.; Han, X. MiR-146a Regulates Inflammatory Cytokine Production in Porphyromonas Gingivalis Lipopolysaccharide-Stimulated B Cells by Targeting IRAK1 but Not TRAF6. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2018, 1864, 925–933. [Google Scholar] [CrossRef]
- Moazzam-Jazi, M.; Lanjanian, H.; Maleknia, S.; Hedayati, M.; Daneshpour, M.S. Interplay between SARS-CoV-2 and Human Long Non-coding RNAs. J. Cell Mol. Med. 2021, 25, 5823–5827. [Google Scholar] [CrossRef]
- Iacona, J.R.; Lutz, C.S. MiR-146a-5p: Expression, Regulation, and Functions in Cancer. WIREs RNA 2019, e1533. [Google Scholar] [CrossRef]
- Miao, C.; Bai, L.; Yang, Y.; Huang, J. Dysregulation of LncRNAs in Rheumatoid Arthritis: Biomarkers, Pathogenesis and Potential Therapeutic Targets. Front. Pharmacol. 2021, 12, 652751. [Google Scholar] [CrossRef]
- Vishnubalaji, R.; Shaath, H.; Alajez, N.M. Protein Coding and Long Noncoding RNA (LncRNA) Transcriptional Landscape in SARS-CoV-2 Infected Bronchial Epithelial Cells Highlight a Role for Interferon and Inflammatory Response. Genes 2020, 11, 760. [Google Scholar] [CrossRef]
- Wei, L.; Li, J.; Han, Z.; Chen, Z.; Zhang, Q. Silencing of LncRNA MALAT1 Prevents Inflammatory Injury after Lung Transplant Ischemia-Reperfusion by Downregulation of IL-8 via P300. Mol. Ther. Nucleic Acids 2019, 18, 285–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.; Wang, C.; Vagts, C.; Raguveer, V.; Finn, P.W.; Perkins, D.L. Long Non-Coding RNAs (LncRNAs) NEAT1 and MALAT1 Are Differentially Expressed in Severe COVID-19 Patients: An Integrated Single Cell Analysis. medRxiv 2021. [Google Scholar] [CrossRef]
- Cui, H.; Banerjee, S.; Guo, S.; Xie, N.; Ge, J.; Jiang, D.; Zörnig, M.; Thannickal, V.J.; Liu, G. Long Noncoding RNA Malat1 Regulates Differential Activation of Macrophages and Response to Lung Injury. JCI Insight 2019, 4, e124522. [Google Scholar] [CrossRef] [Green Version]
- Masoumi, F.; Ghorbani, S.; Talebi, F.; Branton, W.G.; Rajaei, S.; Power, C.; Noorbakhsh, F. Malat1 Long Noncoding RNA Regulates Inflammation and Leukocyte Differentiation in Experimental Autoimmune Encephalomyelitis. J. Neuroimmunol. 2019, 328, 50–59. [Google Scholar] [CrossRef]
- Zhang, P.; Cao, L.; Zhou, R.; Yang, X.; Wu, M. The LncRNA Neat1 Promotes Activation of Inflammasomes in Macrophages. Nat. Commun. 2019, 10, 1495. [Google Scholar] [CrossRef] [Green Version]
- Centa, A.; Fonseca, A.S.; da Silva Ferreira, S.G.; Azevedo, M.L.V.; de Paula, C.B.V.; Nagashima, S.; Machado-Souza, C.; dos Santos Miggiolaro, A.F.R.; Pellegrino Baena, C.; de Noronha, L.; et al. Deregulated MiRNA Expression Is Associated with Endothelial Dysfunction in Post-Mortem Lung Biopsies of COVID-19 Patients. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2021, 320, L405–L412. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, T.; Fei, Z. MiR-26a-5p Alleviates Lipopolysaccharide-induced Acute Lung Injury by Targeting the Connective Tissue Growth Factor. Mol. Med. Rep. 2020, 23. [Google Scholar] [CrossRef]
- Yue, X.-G.; Li, S.-J.; Zhang, Y.; Yang, Z.-G.; Li, H. Regulation of Diabetes-Induced Inflammation in Macrophages by MicroRNA-26a-5p. Int. J. Clin. Exp. Pathol. 2016, 9, 10074–10079. [Google Scholar]
- Li, Z.; Yi, N.; Chen, R.; Meng, Y.; Wang, Y.; Liu, H.; Cao, W.; Hu, Y.; Gu, Y.; Tong, C.; et al. MiR-29b-3p Protects Cardiomyocytes against Endotoxin-Induced Apoptosis and Inflammatory Response through Targeting FOXO3A. Cell. Signal. 2020, 74, 109716. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, M.; Ye, L.; Chen, C.; She, J.; Song, Y. MiR-29b-3p Promotes Particulate Matter-Induced Inflammatory Responses by Regulating the C1QTNF6/AMPK Pathway. Aging 2020, 12, 1141–1158. [Google Scholar] [CrossRef]
- Gambardella, J.; Sardu, C.; Morelli, M.B.; Messina, V.; Marfella, R.; Maggi, P.; Paolisso, G.; Wang, X.; Santulli, G. Abstract 221: Exosomal MicroRNAs Drive Tromboembolism in Covid-19. Circulation 2020, 142, A221. [Google Scholar] [CrossRef]
- Sun, S.; Chai, S.; Zhang, F.; Lu, L. Overexpressed MicroRNA-103a-3p Inhibits Acute Lower-extremity Deep Venous Thrombosis via Inhibition of CXCL12. IUBMB Life 2020, 72, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-L.; Hung, J.-Y.; Chang, W.-A.; Jian, S.-F.; Lin, Y.-S.; Pan, Y.-C.; Wu, C.-Y.; Kuo, P.-L. Hypoxic Lung-Cancer-Derived Extracellular Vesicle MicroRNA-103a Increases the Oncogenic Effects of Macrophages by Targeting PTEN. Mol. Ther. 2018, 26, 568–581. [Google Scholar] [CrossRef] [Green Version]
- Sahu, A.; Jha, P.K.; Prabhakar, A.; Singh, H.D.; Gupta, N.; Chatterjee, T.; Tyagi, T.; Sharma, S.; Kumari, B.; Singh, S.; et al. MicroRNA-145 Impedes Thrombus Formation via Targeting Tissue Factor in Venous Thrombosis. EBioMedicine 2017, 26, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Grover, S.P.; Mackman, N. Tissue Factor: An Essential Mediator of Hemostasis and Trigger of Thrombosis. ATVB 2018, 38, 709–725. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Guo, J.; Gu, J.; Wang, Z.; Wang, G.; Li, H.; Wang, J. Identifying the Key Genes and MicroRNAs in Colorectal Cancer Liver Metastasis by Bioinformatics Analysis and In vitro Experiments. Oncol. Rep. 2018, 41, 279–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delrue, M.; Siguret, V.; Neuwirth, M.; Joly, B.; Beranger, N.; Sène, D.; Chousterman, B.G.; Voicu, S.; Bonnin, P.; Mégarbane, B.; et al. Von Willebrand Factor/ADAMTS13 Axis and Venous Thromboembolism in Moderate-to-severe COVID-19 Patients. Br. J. Haematol. 2021, 192, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sundquist, K.; Elf, J.L.; Strandberg, K.; Svensson, P.J.; Hedelius, A.; Palmér, K.; Memon, A.A.; Sundquist, J.; Zöller, B. Diagnostic Potential of Plasma MicroRNA Signatures in Patients with Deep-Vein Thrombosis. Thromb. Haemost. 2016, 116, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Starikova, I.; Jamaly, S.; Sorrentino, A.; Blondal, T.; Latysheva, N.; Sovershaev, M.; Hansen, J.-B. Differential Expression of Plasma MiRNAs in Patients with Unprovoked Venous Thromboembolism and Healthy Control Individuals. Thromb. Res. 2015, 136, 566–572. [Google Scholar] [CrossRef]
- Forrest, A.R.R.; Kanamori-Katayama, M.; Tomaru, Y.; Lassmann, T.; Ninomiya, N.; Takahashi, Y.; de Hoon, M.J.L.; Kubosaki, A.; Kaiho, A.; Suzuki, M.; et al. Induction of MicroRNAs, Mir-155, Mir-222, Mir-424 and Mir-503, Promotes Monocytic Differentiation through Combinatorial Regulation. Leukemia 2010, 24, 460–466. [Google Scholar] [CrossRef] [Green Version]
- Garg, A.; Seeliger, B.; Derda, A.A.; Xiao, K.; Gietz, A.; Scherf, K.; Sonnenschein, K.; Pink, I.; Hoeper, M.M.; Welte, T.; et al. Circulating Cardiovascular microRNAs in Critically Ill COVID-19 Patients. Eur. J. Heart Fail. 2021, 23, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Ji, R.; Yue, J.; Yang, J.; Liu, X.; Chen, H.; Dean, D.B.; Zhang, C. MicroRNAs Are Aberrantly Expressed in Hypertrophic Heart. Am. J. Pathol. 2007, 170, 1831–1840. [Google Scholar] [CrossRef] [Green Version]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. MicroRNA-21 Contributes to Myocardial Disease by Stimulating MAP Kinase Signalling in Fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef]
- Sayed, D.; He, M.; Hong, C.; Gao, S.; Rane, S.; Yang, Z.; Abdellatif, M. MicroRNA-21 Is a Downstream Effector of AKT That Mediates Its Antiapoptotic Effects via Suppression of Fas Ligand. J. Biol. Chem. 2010, 285, 20281–20290. [Google Scholar] [CrossRef] [Green Version]
- Mahesh, G.; Biswas, R. MicroRNA-155: A Master Regulator of Inflammation. J. Interferon Cytokine Res. 2019, 39, 321–330. [Google Scholar] [CrossRef]
- Van Rooij, E.; Quiat, D.; Johnson, B.A.; Sutherland, L.B.; Qi, X.; Richardson, J.A.; Kelm, R.J.; Olson, E.N. A Family of MicroRNAs Encoded by Myosin Genes Governs Myosin Expression and Muscle Performance. Dev. Cell 2009, 17, 662–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rooij, E.; Sutherland, L.B.; Qi, X.; Richardson, J.A.; Hill, J.; Olson, E.N. Control of Stress-Dependent Cardiac Growth and Gene Expression by a MicroRNA. Science 2007, 316, 575–579. [Google Scholar] [CrossRef] [Green Version]
- Henderson, L.A.; Canna, S.W.; Schulert, G.S.; Volpi, S.; Lee, P.Y.; Kernan, K.F.; Caricchio, R.; Mahmud, S.; Hazen, M.M.; Halyabar, O.; et al. On the Alert for Cytokine Storm: Immunopathology in COVID-19. Arthritis Rheumatol. 2020, 72, 1059–1063. [Google Scholar] [CrossRef] [Green Version]
- Ragab, D.; Salah Eldin, H.; Taeimah, M.; Khattab, R.; Salem, R. The COVID-19 Cytokine Storm; What We Know So Far. Front. Immunol. 2020, 11, 1446. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Q.; Yang, K.; Wang, W.; Jiang, L.; Song, J. Clinical Predictors of Mortality Due to COVID-19 Based on an Analysis of Data of 150 Patients from Wuhan, China. Intensive Care Med. 2020, 46, 846–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campochiaro, C.; Dagna, L. The Conundrum of Interleukin-6 Blockade in COVID-19. Lancet Rheumatol. 2020, 2, e579–e580. [Google Scholar] [CrossRef]
- Thwaites, R.S.; Sanchez Sevilla Uruchurtu, A.; Siggins, M.K.; Liew, F.; Russell, C.D.; Moore, S.C.; Fairfield, C.; Carter, E.; Abrams, S.; Short, C.-E.; et al. Inflammatory Profiles across the Spectrum of Disease Reveal a Distinct Role for GM-CSF in Severe COVID-19. Sci. Immunol. 2021, 6, eabg9873. [Google Scholar] [CrossRef] [PubMed]
- Henter, J.-I.; Horne, A.; Aricó, M.; Egeler, R.M.; Filipovich, A.H.; Imashuku, S.; Ladisch, S.; McClain, K.; Webb, D.; Winiarski, J.; et al. HLH-2004: Diagnostic and Therapeutic Guidelines for Hemophagocytic Lymphohistiocytosis. Pediatr. Blood Cancer 2007, 48, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Dufranc, E.; Del Bello, A.; Belliere, J.; Kamar, N.; Faguer, S.; On Behalf of the TAIDI (Toulouse Acquired Immune Deficiency and Infection) Study Group. IL6-R Blocking with Tocilizumab in Critically Ill Patients with Hemophagocytic Syndrome. Crit. Care 2020, 24, 166. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.-F.; Fu, Z.-Y.; Chen, X.-H.; Cui, Y.; Ji, C.-B.; Guo, X.-R. Tumor Necrosis Factor-α and Interleukin-6 Suppress MicroRNA-1275 Transcription in Human Adipocytes through Nuclear Factor-ΚB. Mol. Med. Rep. 2017, 16, 5965–5971. [Google Scholar] [CrossRef] [Green Version]
- The RECOVERY Collaborative Group. Dexamethasone in Hospitalized Patients with COVID-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar] [CrossRef]
- Abani, O.; Abbas, A.; Abbas, F.; Abbas, M.; Abbasi, S.; Abbass, H.; Abbott, A.; Abdallah, N.; Abdelaziz, A.; Abdelfattah, M.; et al. Tocilizumab in Patients Admitted to Hospital with COVID-19 (RECOVERY): A Randomised, Controlled, Open-Label, Platform Trial. Lancet 2021, 397, 1637–1645. [Google Scholar] [CrossRef]
- Sun, H.; Guo, P.; Zhang, L.; Wang, F. Serum Interleukin-6 Concentrations and the Severity of COVID-19 Pneumonia: A Retrospective Study at a Single Center in Bengbu City, Anhui Province, China, in January and February 2020. Med. Sci. Monit. 2020, 26. [Google Scholar] [CrossRef]
- Wang, D.; Xu, X. A Multicenter, Randomized Controlled Trial for the Efficacy and Safety of Tocilizumab in the Treatment of New Coronavirus Pneumonia (COVID-19); Chinese Clinical Trial Registry. 2020. Available online: www.chictr.org.cn/showprojen.aspx?proj=49409 (accessed on 9 August 2021).
- Salama, C.; Han, J.; Yau, L.; Reiss, W.G.; Kramer, B.; Neidhart, J.D.; Criner, G.J.; Kaplan-Lewis, E.; Baden, R.; Pandit, L.; et al. Tocilizumab in Patients Hospitalized with Covid-19 Pneumonia. N. Engl. J. Med. 2021, 384, 20–30. [Google Scholar] [CrossRef]
- Luly, F.R.; Lévêque, M.; Licursi, V.; Cimino, G.; Martin-Chouly, C.; Théret, N.; Negri, R.; Cavinato, L.; Ascenzioni, F.; Del Porto, P. MiR-146a Is over-Expressed and Controls IL-6 Production in Cystic Fibrosis Macrophages. Sci. Rep. 2019, 9, 16259. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Du, M.; Cao, Z.; Zhang, C.; Hao, Y.; Zhu, J.; He, H. MiR-146a-5p Attenuates IL-1β-induced IL-6 and IL-1β Expression in a Cementoblast-derived Cell Line. Oral Dis. 2020, 26, 1308–1317. [Google Scholar] [CrossRef]
- Boldin, M.P.; Taganov, K.D.; Rao, D.S.; Yang, L.; Zhao, J.L.; Kalwani, M.; Garcia-Flores, Y.; Luong, M.; Devrekanli, A.; Xu, J.; et al. MiR-146a Is a Significant Brake on Autoimmunity, Myeloproliferation, and Cancer in Mice. J. Exp. Med. 2011, 208, 1189–1201. [Google Scholar] [CrossRef]
- Liu, G.; Park, Y.-J.; Abraham, E. Interleukin-1 Receptor-associated Kinase (IRAK) -1- Mediated NF-κ Activation Requires Cytosolic and Nuclear Activity. FASEB J. 2008, 22, 2285–2296. [Google Scholar] [CrossRef]
- Saulle, I.; Garziano, M.; Fenizia, C.; Cappelletti, G.; Parisi, F.; Clerici, M.; Cetin, I.; Savasi, V.; Biasin, M. MiRNA Profiling in Plasma and Placenta of SARS-CoV-2-Infected Pregnant Women. Cells 2021, 10, 1788. [Google Scholar] [CrossRef]
- Mann, M.; Mehta, A.; Zhao, J.L.; Lee, K.; Marinov, G.K.; Garcia-Flores, Y.; Lu, L.-F.; Rudensky, A.Y.; Baltimore, D. An NF-ΚB-MicroRNA Regulatory Network Tunes Macrophage Inflammatory Responses. Nat. Commun. 2017, 8, 851. [Google Scholar] [CrossRef] [Green Version]
- Tomazini, B.M.; Maia, I.S.; Cavalcanti, A.B.; Berwanger, O.; Rosa, R.G.; Veiga, V.C.; Avezum, A.; Lopes, R.D.; Bueno, F.R.; Silva, M.V.A.O.; et al. Effect of Dexamethasone on Days Alive and Ventilator-Free in Patients with Moderate or Severe Acute Respiratory Distress Syndrome and COVID-19: The CoDEX Randomized Clinical Trial. JAMA 2020, 324, 1307. [Google Scholar] [CrossRef]
- Zheng, Y.; Xiong, S.; Jiang, P.; Liu, R.; Liu, X.; Qian, J.; Zheng, X.; Chu, Y. Glucocorticoids Inhibit Lipopolysaccharide-Mediated Inflammatory Response by Downregulating MicroRNA-155: A Novel Anti-Inflammation Mechanism. Free. Radic. Biol. Med. 2012, 52, 1307–1317. [Google Scholar] [CrossRef]
- Li, X.; Liu, C.; Mao, Z.; Xiao, M.; Wang, L.; Qi, S.; Zhou, F. Predictive Values of Neutrophil-to-Lymphocyte Ratio on Disease Severity and Mortality in COVID-19 Patients: A Systematic Review and Meta-Analysis. Crit. Care 2020, 24, 647. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Liu, Y.; Yuan, J.; Wen, Y.; Xu, G.; Zhao, J.; Cheng, L.; Li, J.; Wang, X.; Wang, F.; et al. Single-Cell Landscape of Bronchoalveolar Immune Cells in Patients with COVID-19. Nat. Med. 2020, 26, 842–844. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Q.; Yin, Y.; Zhang, Y.; Cao, Y.; Lin, X.; Huang, L.; Hoffmann, D.; Lu, M.; Qiu, Y. Excessive Neutrophils and Neutrophil Extracellular Traps in COVID-19. Front. Immunol. 2020, 11, 2063. [Google Scholar] [CrossRef] [PubMed]
- Toraih, E.A.; Sedhom, J.A.; Dokunmu, T.M.; Hussein, M.H.; Ruiz, E.M.L.; Muthusamy, K.; Zerfaoui, M.; Kandil, E. Hidden in Plain Sight: The Effects of BCG Vaccination in the COVID-19 Pandemic. J. Med. Virol. 2021, 93, 1950–1966. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, T.; Deng, R.; Xia, X.; Li, B.; Wang, X. A Study of Differential CircRNA and LncRNA Expressions in COVID-19-Infected Peripheral Blood. Sci. Rep. 2021, 11, 7991. [Google Scholar] [CrossRef]
- Hyers, T.M. Venous Thromboembolism. Am. J. Respir. Crit. Care Med. 1999, 159, 1–14. [Google Scholar] [CrossRef]
- Tan, B.K.; Mainbourg, S.; Friggeri, A.; Bertoletti, L.; Douplat, M.; Dargaud, Y.; Grange, C.; Lobbes, H.; Provencher, S.; Lega, J.-C. Arterial and Venous Thromboembolism in COVID-19: A Study-Level Meta-Analysis. Thorax 2021. [Google Scholar] [CrossRef]
- Weitz, J.I.; Fredenburgh, J.C.; Eikelboom, J.W. A Test in Context: D-Dimer. J. Am. Coll. Cardiol. 2017, 70, 2411–2420. [Google Scholar] [CrossRef]
- Calabrò, P.; Gragnano, F.; Golia, E.; Grove, E. Von Willebrand Factor and Venous Thromboembolism: Pathogenic Link and Therapeutic Implications. Semin. Thromb. Hemost. 2018, 44, 249–260. [Google Scholar] [CrossRef]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus–Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061. [Google Scholar] [CrossRef]
- Ji, R.; Cheng, Y.; Yue, J.; Yang, J.; Liu, X.; Chen, H.; Dean, D.B.; Zhang, C. MicroRNA Expression Signature and Antisense-Mediated Depletion Reveal an Essential Role of MicroRNA in Vascular Neointimal Lesion Formation. Circ. Res. 2007, 100, 1579–1588. [Google Scholar] [CrossRef]
- Gupta, M.P. Factors Controlling Cardiac Myosin-Isoform Shift during Hypertrophy and Heart Failure. J. Mol. Cell. Cardiol. 2007, 43, 388–403. [Google Scholar] [CrossRef] [Green Version]
- Gambardella, J.; Coppola, A.; Izzo, R.; Fiorentino, G.; Trimarco, B.; Santulli, G. Role of Endothelial MiR-24 in COVID-19 Cerebrovascular Events. Crit. Care 2021, 25, 306. [Google Scholar] [CrossRef]
- Mone, P.; Gambardella, J.; Wang, X.; Jankauskas, S.S.; Matarese, A.; Santulli, G. MiR-24 Targets the Transmembrane Glycoprotein Neuropilin-1 in Human Brain Microvascular Endothelial Cells. Noncoding RNA 2021, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Maillard, P.V.; Ciaudo, C.; Marchais, A.; Li, Y.; Jay, F.; Ding, S.W.; Voinnet, O. Antiviral RNA Interference in Mammalian Cells. Science 2013, 342, 235–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, L.; Oliveros, J.C.; Fernandez-Delgado, R.; ten Oever, B.R.; Enjuanes, L.; Sola, I. SARS-CoV-Encoded Small RNAs Contribute to Infection-Associated Lung Pathology. Cell Host Microbe 2017, 21, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.; Sany, M.; Us, R.; Islam, M.; Islam, A.B.M.M. Epigenetic Regulator MiRNA Pattern Differences Among SARS-CoV, SARS-CoV-2, and SARS-CoV-2 World-Wide Isolates Delineated the Mystery Behind the Epic Pathogenicity and Distinct Clinical Characteristics of Pandemic COVID-19. Front. Genet. 2020, 11, 765. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, J.; Xu, Y.; Guo, M.; Mi, K.; Xu, R.; Pei, Y.; Zhang, Q.; Luan, X.; Hu, Z.; et al. Implications of the Virus-Encoded MiRNA and Host MiRNA in the Pathogenicity of SARS-CoV-2. arXiv 2020, arXiv:2004.04874. [Google Scholar]
- Yousefi, H.; Poursheikhani, A.; Bahmanpour, Z.; Vatanmakanian, M.; Taheri, M.; Mashouri, L.; Alahari, S.K. SARS-CoV Infection Crosstalk with Human Host Cell Noncoding-RNA Machinery: An in-Silico Approach. Biomed. Pharmacother. 2020, 130, 110548. [Google Scholar] [CrossRef]
- Meng, F.; Siu, G.K.-H.; Mok, B.W.-Y.; Sun, J.; Fung, K.S.C.; Lam, J.Y.-W.; Wong, N.K.; Gedefaw, L.; Luo, S.; Lee, T.M.H.; et al. Viral MicroRNAs Encoded by Nucleocapsid Gene of SARS-CoV-2 Are Detected during Infection, and Targeting Metabolic Pathways in Host Cells. Cells 2021, 10, 1762. [Google Scholar] [CrossRef]
- Bartoszewski, R.; Dabrowski, M.; Jakiela, B.; Matalon, S.; Harrod, K.S.; Sanak, M.; Collawn, J.F. SARS-CoV-2 May Regulate Cellular Responses through Depletion of Specific Host MiRNAs. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2020, 319, L444–L455. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, Y.; Chen, S.; Li, W.; Chen, W.; Gu, W. LncRNA MACC1-AS1 Sponges Multiple MiRNAs and RNA-Binding Protein PTBP1. Oncogenesis 2019, 8, 73. [Google Scholar] [CrossRef]
- Kamel, W.; Noerenberg, M.; Cerikan, B.; Chen, H.; Järvelin, A.I.; Kammoun, M.; Lee, J.Y.; Shuai, N.; Garcia-Moreno, M.; Andrejeva, A.; et al. Global Analysis of Protein-RNA Interactions in SARS-CoV-2-Infected Cells Reveals Key Regulators of Infection. Mol. Cell 2021, 81, 2851–2867.e7. [Google Scholar] [CrossRef]
- Srivastava, R.; Daulatabad, S.V.; Srivastava, M.; Janga, S.C. Role of SARS-CoV-2 in Altering the RNA-Binding Protein and MiRNA-Directed Post-Transcriptional Regulatory Networks in Humans. IJMS 2020, 21, 7090. [Google Scholar] [CrossRef] [PubMed]
- Phillips, N. The Coronavirus Is Here to Stay—Here’s What That Means. Nature 2021, 590, 382–384. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Study | ncRNA of Interest | Effect on ncRNA from Infection | Context Investigated | Additional Findings |
|---|---|---|---|---|
| [19] | miR-766-3p | Downregulated | COVID-19 patients | |
| miR-1275 | Downregulated | COVID-19 patients |
| |
| miR-31-5p | Upregulated | COVID-19 patients |
| |
| [25] | miR-146a-5p | Downregulated | COVID-19 patients | |
| [28] | PVT1 | Upregulated | COVID-19 patients | |
| [31] | MALAT1 | Upregulated | SARS-CoV-2 infected NHBE cells (bronchial) |
|
| [33] | MALAT1 | Downregulated | Mild COVID-19 patients—monocytes and macrophages | |
| MALAT1 | Upregulated | Mild COVID-19 patients–CD4+ T cells | ||
| MALAT1 | Downregulated | Severe COVID-19 patients—CD4+ T cells | ||
| NEAT1 | Downregulated | Mild COVID-19 patients—BAL cells |
| |
| NEAT1 | Upregulated | Severe COVID-19 patients—BAL cells | ||
| [37] | miR-26a-5p | Downregulated | COVID-19 patients |
|
| miR-29-3p | Downregulated | COVID-19 patients |
| |
| [42] | miR-103a | Downregulated | Higher D-dimer COVID-19 patients | |
| miR-145 | Downregulated | Higher D-dimer COVID-19 patients | ||
| miR-885 | Downregulated | Higher D-dimer COVID-19 patients | ||
| miR-424 | Upregulated | Higher D-dimer COVID-19 patients |
| |
| [52] | miR-21 | Upregulated | COVID-19 patients |
|
| miR-155 | Upregulated | COVID-19 patients |
| |
| miR-208a | Upregulated | COVID-19 patients |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plowman, T.; Lagos, D. Non-Coding RNAs in COVID-19: Emerging Insights and Current Questions. Non-Coding RNA 2021, 7, 54. https://doi.org/10.3390/ncrna7030054
Plowman T, Lagos D. Non-Coding RNAs in COVID-19: Emerging Insights and Current Questions. Non-Coding RNA. 2021; 7(3):54. https://doi.org/10.3390/ncrna7030054
Chicago/Turabian StylePlowman, Tobias, and Dimitris Lagos. 2021. "Non-Coding RNAs in COVID-19: Emerging Insights and Current Questions" Non-Coding RNA 7, no. 3: 54. https://doi.org/10.3390/ncrna7030054
APA StylePlowman, T., & Lagos, D. (2021). Non-Coding RNAs in COVID-19: Emerging Insights and Current Questions. Non-Coding RNA, 7(3), 54. https://doi.org/10.3390/ncrna7030054