
MicroRNAs as Potential Biomarkers in Pituitary Adenomas

 ,
, 
Abstract
:1. Introduction
2. miRNAs in Non-Functioning PAs
3. miRNAs in Secreting PAs
3.1. miRNAs in Gonadotrophinomas
3.2. miRNAs in Prolactinomas
3.3. miRNAs in Somatotropinomas
3.4. miRNAs in Corticotropinomas
4. miRNAs in Invasive PAs (IPAs) and Non-Invasive PAs (NIPAs)
5. miRNAs Deregulated in a Variety of Pituitary Tumors
5.1. miRNA Expression Profiling in Human Samples from Patients with Pituitary Tumors
5.2. miRNA Expression Profiling in Mouse (AtT-20 and GT1.1) and Rat (GH3 and MMQ) Pituitary Adenoma Cell Lines
5.3. miRNA Expression Profiling in MEN1-Associated Pituitary Adenomas
6. Circulating miRNAs in PAs
7. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lake, M.G.; Krook, L.S.; Cruz, S.V. Pituitary Adenomas: An Overview. Am. Fam. Physician 2013, 88, 319–327. [Google Scholar]
- Wierinckx, A.; Roche, M.; Legras-Lachuer, C.; Trouillas, J.; Raverot, G.; Lachuer, J. MicroRNAs in Pituitary Tumors. Mol. Cell. Endocrinol. 2017, 456, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Gleiberman, A.S.; Rosenfeld, M.G. Molecular Physiology of Pituitary Development: Signaling and Transcriptional Networks. Physiol. Rev. 2007, 87, 933–963. [Google Scholar] [CrossRef] [Green Version]
- Aflorei, E.D.; Korbonits, M. Epidemiology and Etiopathogenesis of Pituitary Adenomas. J. Neurooncol. 2014, 117, 379–394. [Google Scholar] [CrossRef]
- Beylerli, O.; Beeraka, N.M.; Gareev, I.; Pavlov, V.; Yang, G.; Liang, Y.; Aliev, G. MiRNAs as Noninvasive Biomarkers and Therapeutic Agents of Pituitary Adenomas. Int. J. Mol. Sci. 2020, 21, 7287. [Google Scholar] [CrossRef]
- Feng, Y.; Mao, Z.-G.; Wang, X.; Du, Q.; Jian, M.; Zhu, D.; Xiao, Z.; Wang, H.-J.; Zhu, Y.-H. MicroRNAs and Target Genes in Pituitary Adenomas. Horm. Metab. Res. 2018, 50, 179–192. [Google Scholar] [CrossRef]
- Molitch, M.E. Diagnosis and Treatment of Pituitary Adenomas: A Review. JAMA 2017, 317, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Colao, A.; Grasso, L.F.S.; Giustina, A.; Melmed, S.; Chanson, P.; Pereira, A.M.; Pivonello, R. Acromegaly. Nat. Rev. Dis. Primers 2019, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Hurley, D.M.; Ho, K.K.Y. 9: Pituitary Disease in Adults. Med. J. Aust. 2004, 180, 419–425. [Google Scholar] [CrossRef]
- Loeffler, J.S.; Shih, H.A. Radiation Therapy in the Management of Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2011, 96, 1992–2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, D.; Starke, R.M.; Sheehan, J.P. Treatment Paradigms for Pituitary Adenomas: Defining the Roles of Radiosurgery and Radiation Therapy. J. Neurooncol. 2014, 117, 445–457. [Google Scholar] [CrossRef]
- Asa, S.L.; Ezzat, S. The Pathogenesis of Pituitary Tumors. Annu. Rev. Pathol. 2009, 4, 97–126. [Google Scholar] [CrossRef]
- Clayton, R.N.; Farrell, W.E. Pituitary Tumour Clonality Revisited. Mol. Pathol. Pituit. 2004, 32, 186–204. [Google Scholar] [CrossRef]
- Melmed, S. Pathogenesis of Pituitary Tumors. Nat. Rev. Endocrinol. 2011, 7, 257–266. [Google Scholar] [CrossRef]
- Wierinckx, A.; Roche, M.; Raverot, G.; Legras-Lachuer, C.; Croze, S.; Nazaret, N.; Rey, C.; Auger, C.; Jouanneau, E.; Chanson, P.; et al. Integrated Genomic Profiling Identifies Loss of Chromosome 11p Impacting Transcriptomic Activity in Aggressive Pituitary PRL Tumors. Brain Pathol. 2011, 21, 533–543. [Google Scholar] [CrossRef]
- Bottoni, A.; Piccin, D.; Tagliati, F.; Luchin, A.; Zatelli, M.C.; Degli Uberti, E.C. MiR-15a and MiR-16-1 down-Regulation in Pituitary Adenomas. J. Cell. Physiol. 2005, 204, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Malan-Müller, S.; Hemmings, S.M.J.; Seedat, S. Big Effects of Small RNAs: A Review of MicroRNAs in Anxiety. Mol. Neurobiol. 2013, 47, 726–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huntzinger, E.; Izaurralde, E. Gene Silencing by MicroRNAs: Contributions of Translational Repression and MRNA Decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Ipsaro, J.J.; Joshua-Tor, L. From Guide to Target: Molecular Insights into Eukaryotic RNA-Interference Machinery. Nat. Struct. Mol. Biol. 2015, 22, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Gossing, W.; Frohme, M.; Radke, L. Biomarkers for Liquid Biopsies of Pituitary Neuroendocrine Tumors. Biomedicines 2020, 8, 148. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating MicroRNAs as Stable Blood-Based Markers for Cancer Detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of MicroRNAs in Serum: A Novel Class of Biomarkers for Diagnosis of Cancer and Other Diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chim, S.S.C.; Shing, T.K.F.; Hung, E.C.W.; Leung, T.-Y.; Lau, T.-K.; Chiu, R.W.K.; Lo, Y.M.D. Detection and Characterization of Placental MicroRNAs in Maternal Plasma. Clin. Chem. 2008, 54, 482–490. [Google Scholar] [CrossRef] [Green Version]
- Lawrie, C.H.; Gal, S.; Dunlop, H.M.; Pushkaran, B.; Liggins, A.P.; Pulford, K.; Banham, A.H.; Pezzella, F.; Boultwood, J.; Wainscoat, J.S.; et al. Detection of Elevated Levels of Tumour-Associated MicroRNAs in Serum of Patients with Diffuse Large B-Cell Lymphoma. Br. J. Haematol. 2008, 141, 672–675. [Google Scholar] [CrossRef]
- Gallo, A.; Tandon, M.; Alevizos, I.; Illei, G.G. The Majority of MicroRNAs Detectable in Serum and Saliva Is Concentrated in Exosomes. PLoS ONE 2012, 7, e30679. [Google Scholar] [CrossRef] [Green Version]
- Cogswell, J.P.; Ward, J.; Taylor, I.A.; Waters, M.; Shi, Y.; Cannon, B.; Kelnar, K.; Kemppainen, J.; Brown, D.; Chen, C.; et al. Identification of MiRNA Changes in Alzheimer’s Disease Brain and CSF Yields Putative Biomarkers and Insights into Disease Pathways. J. Alzheimer’s Dis. 2008, 14, 27–41. [Google Scholar] [CrossRef]
- Da Silveira, J.C.; Veeramachaneni, D.N.R.; Winger, Q.A.; Carnevale, E.M.; Bouma, G.J. Cell-Secreted Vesicles in Equine Ovarian Follicular Fluid Contain MiRNAs and Proteins: A Possible New Form of Cell Communication within the Ovarian Follicle. Biol. Reprod. 2012, 86, 71. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The MicroRNA Spectrum in 12 Body Fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Li, M.; Wang, X.; Li, Q.; Wang, T.; Zhu, Q.; Zhou, X.; Wang, X.; Gao, X.; Li, X. Immune-Related MicroRNAs Are Abundant in Breast Milk Exosomes. Int. J. Biol. Sci. 2011, 8, 118–123. [Google Scholar] [CrossRef]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs Are Transported in Plasma and Delivered to Recipient Cells by High-Density Lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchinovich, A.; Weiz, L.; Burwinkel, B. Extracellular MiRNAs: The Mystery of Their Origin and Function. Trends Biochem. Sci. 2012, 37, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Tabet, F.; Vickers, K.C.; Cuesta Torres, L.F.; Wiese, C.B.; Shoucri, B.M.; Lambert, G.; Catherinet, C.; Prado-Lourenco, L.; Levin, M.G.; Thacker, S.; et al. HDL-Transferred MicroRNA-223 Regulates ICAM-1 Expression in Endothelial Cells. Nat. Commun. 2014, 5, 3292. [Google Scholar] [CrossRef] [Green Version]
- Iftikhar, H.; Carney, G.E. Evidence and Potential in Vivo Functions for Biofluid MiRNAs: From Expression Profiling to Functional Testing: Potential Roles of Extracellular MiRNAs as Indicators of Physiological Change and as Agents of Intercellular Information Exchange. Bioessays 2016, 38, 367–378. [Google Scholar] [CrossRef]
- Materozzi, M.; Merlotti, D.; Gennari, L.; Bianciardi, S. The Potential Role of MiRNAs as New Biomarkers for Osteoporosis. Int. J. Endocrinol. 2018, 2018, 2342860. [Google Scholar] [CrossRef] [PubMed]
- Mercado, M.; Melgar, V.; Salame, L.; Cuenca, D. Clinically Non-Functioning Pituitary Adenomas: Pathogenic, Diagnostic and Therapeutic Aspects. Endocrinol. Diabetes Nutr. 2017, 64, 384–395. [Google Scholar] [CrossRef]
- Tabaee, A.; Anand, V.K.; Barrón, Y.; Hiltzik, D.H.; Brown, S.M.; Kacker, A.; Mazumdar, M.; Schwartz, T.H. Endoscopic Pituitary Surgery: A Systematic Review and Meta-Analysis. J. Neurosurg. 2009, 111, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Di Ieva, A.; Rotondo, F.; Syro, L.V.; Cusimano, M.D.; Kovacs, K. Aggressive Pituitary Adenomas-Diagnosis and Emerging Treatments. Nat. Rev. Endocrinol. 2014, 10, 423–435. [Google Scholar] [CrossRef]
- Fernandez-Miranda, J.C.; Zwagerman, N.T.; Abhinav, K.; Lieber, S.; Wang, E.W.; Snyderman, C.H.; Gardner, P.A. Cavernous Sinus Compartments from the Endoscopic Endonasal Approach: Anatomical Considerations and Surgical Relevance to Adenoma Surgery. J. Neurosurg. 2018, 129, 430–441. [Google Scholar] [CrossRef]
- Boresowicz, J.; Kober, P.; Rusetska, N.; Maksymowicz, M.; Paziewska, A.; Dąbrowska, M.; Zeber-Lubecka, N.; Kunicki, J.; Bonicki, W.; Ostrowski, J.; et al. The Search of MiRNA Related to Invasive Growth of Nonfunctioning Gonadotropic Pituitary Tumors. Int. J. Endocrinol. 2020, 2020, 3730657. [Google Scholar] [CrossRef] [PubMed]
- Elston, M.S.; Gill, A.J.; Conaglen, J.V.; Clarkson, A.; Shaw, J.M.; Law, A.J.J.; Cook, R.J.; Little, N.S.; Clifton-Bligh, R.J.; Robinson, B.G.; et al. Wnt Pathway Inhibitors Are Strongly Down-Regulated in Pituitary Tumors. Endocrinology 2008, 149, 1235–1242. [Google Scholar] [CrossRef] [Green Version]
- Hong, L.; Wu, Y.; Feng, J.; Yu, S.; Li, C.; Wu, Y.; Li, Z.; Cao, L.; Wang, F.; Zhang, Y. Overexpression of the Cell Adhesion Molecule Claudin-9 Is Associated with Invasion in Pituitary Oncocytomas. Hum. Pathol. 2014, 45, 2423–2429. [Google Scholar] [CrossRef]
- Song, W.; Qian, L.; Jing, G.; Jie, F.; Xiaosong, S.; Chunhui, L.; Yangfang, L.; Guilin, L.; Gao, H.; Yazhuo, Z. Aberrant Expression of the SFRP and WIF1 Genes in Invasive Non-Functioning Pituitary Adenomas. Mol. Cell. Endocrinol. 2018, 474, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, F.; Bajetto, A.; Stumm, R.; Pattarozzi, A.; Porcile, C.; Zona, G.; Dorcaratto, A.; Ravetti, J.-L.; Minuto, F.; Spaziante, R.; et al. Overexpression of Stromal Cell–Derived Factor 1 and Its Receptor CXCR4 Induces Autocrine/Paracrine Cell Proliferation in Human Pituitary Adenomas. Clin. Cancer Res. 2008, 14, 5022–5032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, F.; Dai, C.; Chen, S.; Wu, Q.; Liu, X.; Hong, Y.; Wang, Z.; Li, L.; Yan, W.; Wang, R.; et al. CXCL12-Regulated MiR-370-3p Functions as a Tumor Suppressor Gene by Targeting HMGA2 in Nonfunctional Pituitary Adenomas. Mol. Cell. Endocrinol. 2019, 488, 25–35. [Google Scholar] [CrossRef]
- Du, Q.; Hu, B.; Feng, Y.; Wang, Z.; Wang, X.; Zhu, D.; Zhu, Y.; Jiang, X.; Wang, H. CircOMA1-Mediated MiR-145-5p Suppresses Tumor Growth of Nonfunctioning Pituitary Adenomas by Targeting TPT1. J. Clin. Endocrinol. Metab. 2019, 104, 2419–2434. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Yang, J.; Wang, G.; Wang, C.; Zhang, H. Bioinformatic Analysis of Gene Expression Profiles of Pituitary Gonadotroph Adenomas. Oncol. Lett. 2018, 15, 1655–1663. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, D.; Mussnich, P.; Sepe, R.; Raia, M.; Del Vecchio, L.; Cappabianca, P.; Pellecchia, S.; Petrosino, S.; Saggio, S.; Solari, D.; et al. RPSAP52 LncRNA Is Overexpressed in Pituitary Tumors and Promotes Cell Proliferation by Acting as MiRNA Sponge for HMGA Proteins. J. Mol. Med. 2019, 97, 1019–1032. [Google Scholar] [CrossRef]
- D’Angelo, D.; Arra, C.; Fusco, A. RPSAP52 LncRNA Inhibits P21Waf1/CIP Expression by Interacting With the RNA Binding Protein HuR. Oncol. Res. 2020, 28, 191–201. [Google Scholar] [CrossRef]
- LaPierre, M.P.; Godbersen, S.; Torres Esteban, M.; Schad, A.N.; Treier, M.; Ghoshdastider, U.; Stoffel, M. MicroRNA-7a2 Regulates Prolactin in Developing Lactotrophs and Prolactinoma Cells. Endocrinology 2021, 162, bqaa220. [Google Scholar] [CrossRef]
- Wu, Z.; Cai, L.; Lu, J.; Wang, C.D.; Guan, J.; Chen, X.; Wu, J.; Zheng, W.; Wu, Z.; Li, Q.; et al. MicroRNA-93 Mediates Cabergoline-Resistance by Targeting ATG7 in Prolactinoma. J. Endocrinol. 2018, 240, 1–13. [Google Scholar] [CrossRef]
- Wu, Z.; Zheng, Y.; Xie, W.; Li, Q.; Zhang, Y.; Ren, B.; Cai, L.; Cheng, Y.; Tang, H.; Su, Z.; et al. The Long Noncoding RNA-H19/MiRNA-93a/ATG7 Axis Regulates the Sensitivity of Pituitary Adenomas to Dopamine Agonists. Mol. Cell. Endocrinol. 2020, 518, 111033. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Mao, Z.; Du, Q.; Jiang, X.; Wang, Z.; Xiao, Z.; Zhu, D.; Wang, X.; Zhu, Y.; Wang, H. MiR-93-5p Targets Smad7 to Regulate the Transforming Growth Factor-Β1/Smad3 Pathway and Mediate Fibrosis in Drug-Resistant Prolactinoma. Brain Res. Bull. 2019, 149, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Wang, Z.; Hu, B.; Mao, Z.; Zhu, D.; Feng, Y.; Zhu, Y. MiR-1299 Promotes the Synthesis and Secretion of Prolactin by Inhibiting FOXO1 Expression in Drug-Resistant Prolactinomas. Biochem. Biophys. Res. Commun. 2019, 520, 79–85. [Google Scholar] [CrossRef]
- Jian, M.; Du, Q.; Zhu, D.; Mao, Z.; Wang, X.; Feng, Y.; Xiao, Z.; Wang, H.; Zhu, Y. Tumor Suppressor MiR-145-5p Sensitizes Prolactinoma to Bromocriptine by Downregulating TPT1. J. Endocrinol. Investig. 2019, 42, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, T.; Xiong, J.; Hu, B.; Luo, J.; Xi, Q.; Jiang, Q.; Sun, J.; Zhang, Y. MiR-130a-3p Inhibits PRL Expression and Is Associated With Heat Stress-Induced PRL Reduction. Front. Endocrinol. 2020, 11, 92. [Google Scholar] [CrossRef] [Green Version]
- Lei, C.; Jing, G.; Jichao, W.; Xiaohui, L.; Fang, Q.; Hua, G.; Yazhou, M.; Zhang, Y. MiR-137′s Tumor Suppression on Prolactinomas by Targeting MITF and Modulating Wnt Signaling Pathway. J. Clin. Endocrinol. Metab. 2019, 104, 6391–6402. [Google Scholar] [CrossRef]
- Xiong, Y.; Tang, Y.; Fan, F.; Zeng, Y.; Li, C.; Zhou, G.; Hu, Z.; Zhang, L.; Liu, Z. Exosomal Hsa-MiR-21-5p Derived from Growth Hormone-Secreting Pituitary Adenoma Promotes Abnormal Bone Formation in Acromegaly. Transl. Res. 2020, 215, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Daly, A.F.; Tichomirowa, M.A.; Petrossians, P.; Heliövaara, E.; Jaffrain-Rea, M.-L.; Barlier, A.; Naves, L.A.; Ebeling, T.; Karhu, A.; Raappana, A.; et al. Clinical Characteristics and Therapeutic Responses in Patients with Germ-Line AIP Mutations and Pituitary Adenomas: An International Collaborative Study. J. Clin. Endocrinol. Metab. 2010, 95, E373–E383. [Google Scholar] [CrossRef] [Green Version]
- Bogner, E.-M.; Daly, A.F.; Gulde, S.; Karhu, A.; Irmler, M.; Beckers, J.; Mohr, H.; Beckers, A.; Pellegata, N.S. MiR-34a Is Upregulated in AIP-Mutated Somatotropinomas and Promotes Octreotide Resistance. Int. J. Cancer 2020, 147, 3523–3538. [Google Scholar] [CrossRef]
- Belaya, Z.; Grebennikova, T.; Melnichenko, G.; Nikitin, A.; Solodovnikov, A.; Brovkina, O.; Grigoriev, A.; Rozhinskaya, L.; Lutsenko, A.; Dedov, I. Effects of Active Acromegaly on Bone MRNA and MicroRNA Expression Patterns. Eur. J. Endocrinol. 2018, 178, 353–364. [Google Scholar] [CrossRef]
- Armagan, D.M.; Akdemir, A.S.; Ozkaya, H.M.; Korkmaz, O.P.; Gazioglu, N.; Kadioglu, P.; Tanriover, N.; Dagistanli, K.-F.; Dirican, A.; Ozturk, M. SNPs of MiR-23b, MiR-107 and HMGA2 and Their Relations with the Response to Medical Treatment in Acromegaly Patients. Exp. Clin. Endocrinol. Diabetes 2021, 129, 1–8. [Google Scholar] [CrossRef] [PubMed]
- DeVore, S.B.; Young, C.H.; Li, G.; Sundararajan, A.; Ramaraj, T.; Mudge, J.; Schilkey, F.; Muth, A.; Thompson, P.R.; Cherrington, B.D. Histone Citrullination Represses MicroRNA Expression, Resulting in Increased Oncogene MRNAs in Somatolactotrope Cells. Mol. Cell. Biol. 2018, 38, e00084-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Zhang, T.; Wang, Q.; Gao, H. Overexpression of MicroRNA-34a Attenuates Proliferation and Induces Apoptosis in Pituitary Adenoma Cells via SOX7. Mol. Ther. Oncolytics 2018, 10, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Pivonello, R.; De Martino, M.C.; De Leo, M.; Lombardi, G.; Colao, A. Cushing’s Syndrome. Endocrinol. Metab. Clin. 2008, 37, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Gu, C.; Yang, Y.; Xue, J.; Sun, Y.; Jian, F.; Chen, D.; Bian, L.; Sun, Q. TSP-1 Is Downregulated and Inversely Correlates with MiR-449c Expression in Cushing’s Disease. J. Cell. Mol. Med. 2019, 23, 4097–4110. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Guo, J.; Shen, Y.; Dong, W.; Gao, H.; Miao, Y.; Li, C.; Zhang, Y. Functions and Mechanisms of Tumor Necrosis Factor-α and Noncoding RNAs in Bone-Invasive Pituitary Adenomas. Clin. Cancer Res. 2018, 24, 5757–5766. [Google Scholar] [CrossRef] [Green Version]
- Shen, D.-W.; Li, Y.-L.; Hou, Y.-J.; Xu, Z.-D.; Li, Y.-Z.; Chang, J.-Y. MicroRNA-543 Promotes Cell Invasion and Impedes Apoptosis in Pituitary Adenoma via Activating the Wnt/β-Catenin Pathway by Negative Regulation of Smad7. Biosci. Biotechnol. Biochem. 2019, 83, 1035–1044. [Google Scholar] [CrossRef]
- Zhang, C.; Qian, Y.; Qiao, Y.; Li, Y.; Wang, W.; Li, J.; Deng, X. Analysis of Whole Genome-Wide MicroRNA Transcriptome Profiling in Invasive Pituitary Adenomas and Non-Invasive Pituitary Adenomas. Chin. Neurosurg. J. 2019, 5, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grzywa, T.M.; Klicka, K.; Rak, B.; Mehlich, D.; Garbicz, F.; Zieliński, G.; Maksymowicz, M.; Sajjad, E.; Włodarski, P.K. Lineage-Dependent Role of MiR-410-3p as OncomiR in Gonadotroph and Corticotroph Pituitary Adenomas or Tumor Suppressor MiR in Somatotroph Adenomas via MAPK, PTEN/AKT, and STAT3 Signaling Pathways. Endocrine 2019, 65, 646–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Xu, T.; Qiu, P.; Xu, G. Caveolin-1 Promotes Pituitary Adenoma Cells Migration and Invasion by Regulating the Interaction between EGR1 and KLF5. Exp. Cell Res. 2018, 367, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, G.; Gao, Y.; Zhao, C.; Li, X.; Zhang, F.; Jiang, C.; Wu, B. Lnc-SNHG1 Activates the TGFBR2/SMAD3 and RAB11A/Wnt/β-Catenin Pathway by Sponging MiR-302/372/373/520 in Invasive Pituitary Tumors. Cell. Physiol. Biochem. 2018, 48, 1291–1303. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Xiao, Z.; Wang, Z.; Hu, B.; Duan, C.; Zhu, Z.; Gao, N.; Zhu, Y.; Wang, H. MEG3/MIR-376B-3P/HMGA2 Axis Is Involved in Pituitary Tumor Invasiveness. J. Neurosurg. 2020, 1–13. [Google Scholar] [CrossRef]
- Zhao, P.; Cheng, J.; Li, B.; Nie, D.; Li, C.; Gui, S.; Wang, H.; Zhang, Y. Up-Regulation of the Expressions of MiR-149-5p and MiR-99a-3p in Exosome Inhibits the Progress of Pituitary Adenomas. Cell Biol. Toxicol. 2021, 37, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Lu, G.; Li, Y.; Zhou, S.; Zhou, D.; Tao, H. MiR-137 Functions as a Tumor Suppressor Gene in Pituitary Adenoma by Targeting AKT2. Int. J. Clin. Exp. Pathol. 2019, 12, 1557–1564. [Google Scholar]
- Zhao, P.; Cheng, J.; Li, B.; Nie, D.; Wang, H.; Li, C.; Gui, S.; Zhang, Y. LncRNA PCAT6 Regulates the Progression of Pituitary Adenomas by Regulating the MiR-139-3p/BRD4 Axis. Cancer Cell Int. 2021, 21, 14. [Google Scholar] [CrossRef]
- Zhou, K.; Li, S.; Du, G.; Fan, Y.; Wu, P.; Sun, H.; Zhang, T. LncRNA XIST Depletion Prevents Cancer Progression in Invasive Pituitary Neuroendocrine Tumor by Inhibiting BFGF via Upregulation of MicroRNA-424-5p. OncoTargets Ther. 2019, 12, 7095–7109. [Google Scholar] [CrossRef] [Green Version]
- Emmrich, S.; Pützer, B.M. Checks and Balances: E2F-MicroRNA Crosstalk in Cancer Control. Cell Cycle 2010, 9, 2555–2567. [Google Scholar] [CrossRef] [PubMed]
- Dong, P.; Maddali, M.V.; Srimani, J.K.; Thélot, F.; Nevins, J.R.; Mathey-Prevot, B.; You, L. Division of Labour between Myc and G1 Cyclins in Cell Cycle Commitment and Pace Control. Nat. Commun. 2014, 5, 4750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Martínez, A.; López-Muñoz, B.; Fajardo, C.; Cámara, R.; Lamas, C.; Silva-Ortega, S.; Aranda, I.; Picó, A. Increased E2F1 MRNA and MiR-17-5p Expression Is Correlated to Invasiveness and Proliferation of Pituitary Neuroendocrine Tumours. Diagnostics 2020, 10, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.; Chen, L.; Hu, X.; Tang, J.; He, L.; Hu, J.; Fei, F.; Wang, Q. Next-Generation Sequencing of MicroRNAs Reveals a Unique Expression Pattern in Different Types of Pituitary Adenomas. Endocr. J. 2019, 66, 709–722. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Yang, J.; Ding, J.; Li, S.; Wu, H.; Xiong, Y.; Zhou, F.; Jiang, Y.; Teng, L.; Yang, J. Downregulation of Glucose-6-phosphate Dehydrogenase by MicroRNA-1 Inhibits the Growth of Pituitary Tumor Cells. Oncol. Rep. 2018, 40, 3533–3542. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yuan, Y.; Li, J.; Ren, H.; Cai, Q.; Chen, X.; Liang, H.; Shan, H.; Fu, Z.D.; Gao, X.; et al. MicroRNA-1 Aggravates Cardiac Oxidative Stress by Post-Transcriptional Modification of the Antioxidant Network. Cell Stress Chaperones 2015, 20, 411–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Li, S. MiR-205-5p Inhibits Cell Migration and Invasion in Prostatic Carcinoma by Targeting ZEB1. Oncol. Lett. 2018, 16, 1715–1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, Y.; Nishikawa, R.; Kato, M.; Okato, A.; Arai, T.; Kojima, S.; Yamazaki, K.; Naya, Y.; Ichikawa, T.; Seki, N. Regulation of HMGB3 by Antitumor MiR-205-5p Inhibits Cancer Cell Aggressiveness and Is Involved in Prostate Cancer Pathogenesis. J. Hum. Genet. 2018, 63, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Nagai, H.; Hasegawa, S.; Uchida, F.; Terabe, T.; Ishibashi Kanno, N.; Kato, K.; Yamagata, K.; Sakai, S.; Kawashiri, S.; Sato, H.; et al. MicroRNA-205-5p Suppresses the Invasiveness of Oral Squamous Cell Carcinoma by Inhibiting TIMP-2 Expression. Int. J. Oncol. 2018, 52, 841–850. [Google Scholar] [CrossRef]
- Hu, A.; Zhang, Y.; Zhao, X.; Li, J.; Ying, Y. CBX1 Is a Direct Target of MiR-205-5p and Contributes to the Progression of Pituitary Tumor. Pharmazie 2019, 74, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Hou, B.; Ye, Z.; Ling, C.; Guo, Y. Knockdown of Long Non-Coding RNA AFAP1-AS1 Inhibits Growth and Promotes Apoptosis in Pituitary Adenomas. Int. J. Clin. Exp. Pathol. 2018, 11, 1238–1246. [Google Scholar]
- Tang, H.; Zhu, D.; Zhang, G.; Luo, X.; Xie, W. AFAP1-AS1 Promotes Proliferation of Pituitary Adenoma Cells through MiR-103a-3p to Activate PI3K/AKT Signaling Pathway. World Neurosurg. 2019, 130, e888–e898. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, J.; Zhang, C.; Wang, P.; Huang, C.; Peng, H. MiR-219a-2-3p Suppresses Cell Proliferation and Promotes Apoptosis by Targeting MDM2/P53 in Pituitary Adenomas Cells. Biosci. Biotechnol. Biochem. 2020, 84, 911–918. [Google Scholar] [CrossRef]
- Donati, S.; Ciuffi, S.; Marini, F.; Palmini, G.; Miglietta, F.; Aurilia, C.; Brandi, M.L. Multiple Endocrine Neoplasia Type 1: The Potential Role of MicroRNAs in the Management of the Syndrome. Int. J. Mol. Sci. 2020, 21, 7592. [Google Scholar] [CrossRef]
- Lines, K.E.; Newey, P.J.; Yates, C.J.; Stevenson, M.; Dyar, R.; Walls, G.V.; Bowl, M.R.; Thakker, R.V. MiR-15a/MiR-16-1 Expression Inversely Correlates with Cyclin D1 Levels in Men1 Pituitary NETs. J. Endocrinol. 2018, 240, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Bottoni, A.; Zatelli, M.C.; Ferracin, M.; Tagliati, F.; Piccin, D.; Vignali, C.; Calin, G.A.; Negrini, M.; Croce, C.M.; Degli Uberti, E.C. Identification of Differentially Expressed MicroRNAs by Microarray: A Possible Role for MicroRNA Genes in Pituitary Adenomas. J. Cell. Physiol. 2007, 210, 370–377. [Google Scholar] [CrossRef]
- Amaral, F.C.; Torres, N.; Saggioro, F.; Neder, L.; Machado, H.R.; Silva, W.A.; Moreira, A.C.; Castro, M. MicroRNAs Differentially Expressed in ACTH-Secreting Pituitary Tumors. J. Clin. Endocrinol. Metab. 2009, 94, 320–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, Z.R.; Asa, S.L.; Siomi, H.; Siomi, M.C.; Yoshimoto, K.; Yamada, S.; Wang, E.L.; Rahman, M.M.; Inoue, H.; Itakura, M.; et al. Overexpression of HMGA2 Relates to Reduction of the Let-7 and Its Relationship to Clinicopathological Features in Pituitary Adenomas. Mod. Pathol. 2009, 22, 431–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Angelo, D.; Palmieri, D.; Mussnich, P.; Roche, M.; Wierinckx, A.; Raverot, G.; Fedele, M.; Croce, C.M.; Trouillas, J.; Fusco, A. Altered MicroRNA Expression Profile in Human Pituitary GH Adenomas: Down-Regulation of MiRNA Targeting HMGA1, HMGA2, and E2F1. J. Clin. Endocrinol. Metab. 2012, 97, E1128–E1138. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, D.; D’Angelo, D.; Valentino, T.; De Martino, I.; Ferraro, A.; Wierinckx, A.; Fedele, M.; Trouillas, J.; Fusco, A. Downregulation of HMGA -Targeting MicroRNAs Has a Critical Role in Human Pituitary Tumorigenesis. Oncogene 2012, 31, 3857–3865. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.-K.; Zhao, G.-Y.; Tian, L.-Y.; Liu, L.; Yan, K.; Ma, Y.-L.; Ji, Z.-W.; Li, X.-X.; Han, K.; Gao, J.; et al. MiR-15a and MiR-16-1 Downregulate CCND1 and Induce Apoptosis and Cell Cycle Arrest in Osteosarcoma. Oncol. Rep. 2012, 28, 1764–1770. [Google Scholar] [CrossRef] [Green Version]
- Gazioglu, N.M.; Erensoy, N.; Kadioglu, P.; Sayitoglu, M.A.; Ersoy, I.H.; Hatirnaz, O.; Kisacik, B.; Oz, B.; Sar, M.; Ozbek, U.; et al. Altered Cyclin D1 Genotype Distribution in Human Sporadic Pituitary Adenomas. Med. Sci. Monit. 2007, 13, CR457–CR463. [Google Scholar] [PubMed]
- Salerno, E.; Scaglione, B.J.; Coffman, F.D.; Brown, B.D.; Baccarini, A.; Fernandes, H.; Marti, G.; Raveche, E.S. Correcting MiR-15a/16 Genetic Defect in New Zealand Black Mouse Model of CLL Enhances Drug Sensitivity. Mol. Cancer Ther. 2009, 8, 2684–2692. [Google Scholar] [CrossRef] [Green Version]
- Belaya, Z.; Khandaeva, P.; Nonn, L.; Nikitin, A.; Solodovnikov, A.; Sitkin, I.; Grigoriev, A.; Pikunov, M.; Lapshina, A.; Rozhinskaya, L.; et al. Circulating Plasma MicroRNA to Differentiate Cushing’s Disease From Ectopic ACTH Syndrome. Front. Endocrinol. 2020, 11, 331. [Google Scholar] [CrossRef]
- Németh, K.; Darvasi, O.; Likó, I.; Szücs, N.; Czirják, S.; Reiniger, L.; Szabó, B.; Krokker, L.; Pállinger, É.; Igaz, P.; et al. Comprehensive Analysis of Circulating MicroRNAs in Plasma of Patients with Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2019, 104, 4151–4168. [Google Scholar] [CrossRef]
- Liu, H.-Y.; Gu, W.-J.; Wang, C.-Z.; Ji, X.-J.; Mu, Y.-M. Matrix Metalloproteinase-9 and -2 and Tissue Inhibitor of Matrix Metalloproteinase-2 in Invasive Pituitary Adenomas. Medicine 2016, 95, e3904. [Google Scholar] [CrossRef]
- Ortiz, L.D.; Syro, L.V.; Scheithauer, B.W.; Ersen, A.; Uribe, H.; Fadul, C.E.; Rotondo, F.; Horvath, E.; Kovacs, K. Anti-VEGF Therapy in Pituitary Carcinoma. Pituitary 2012, 15, 445–449. [Google Scholar] [CrossRef]
- Lu, B.; Liu, G.-L.; Yu, F.; Li, W.-J.; Xiang, X.-X.; Xiao, H.-Z. MicroRNA-16/VEGFR2/P38/NF-κB Signaling Pathway Regulates Cell Growth of Human Pituitary Neoplasms. Oncol. Rep. 2018, 39, 1235–1244. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Li, J.; Feng, J.; Li, Z.; Liu, Q.; Lv, P.; Wang, F.; Gao, H.; Zhang, Y. Identification of Serum MiRNA-423-5p Expression Signature in Somatotroph Adenomas. Int. J. Endocrinol. 2019, 2019, 8516858. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, A.; Karavitaki, N.; Wass, J.A.H. Prevalence of Pituitary Adenomas: A Community-Based, Cross-Sectional Study in Banbury (Oxfordshire, UK). Clin. Endocrinol. 2010, 72, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Scheithauer, B.W.; Gaffey, T.A.; Lloyd, R.V.; Sebo, T.J.; Kovacs, K.T.; Horvath, E.; Yapicier, O.; Young, W.F.; Meyer, F.B.; Kuroki, T.; et al. Pathobiology of Pituitary Adenomas and Carcinomas. Neurosurgery 2006, 59, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Ezzat, S.; Cheng, S.; Asa, S.L. Epigenetics of Pituitary Tumors: Pathogenetic and Therapeutic Implications. Mol. Cell. Endocrinol. 2018, 469, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Marini, F.; Cianferotti, L.; Brandi, M.L. Epigenetic Mechanisms in Bone Biology and Osteoporosis: Can They Drive Therapeutic Choices? Int. J. Mol. Sci. 2016, 17, 1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Su, Z.; Sanai, N.; Xue, X.; Lu, L.; Chen, Y.; Wu, J.; Zheng, W.; Zhuge, Q.; Wu, Z.B. MicroRNA Expression Profile and Differentially-Expressed Genes in Prolactinomas Following Bromocriptine Treatment. Oncol. Rep. 2012, 27, 1312–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.B.; Li, W.Q.; Lin, S.J.; Wang, C.D.; Cai, L.; Lu, J.L.; Chen, Y.X.; Su, Z.P.; Shang, H.B.; Yang, W.L.; et al. MicroRNA Expression Profile of Bromocriptine-Resistant Prolactinomas. Mol. Cell. Endocrinol. 2014, 395, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Mao, Z.; He, D.; Liao, C.; Jiang, X.; Lei, N.; Hu, B.; Wang, X.; Li, Z.; Lin, Y.; et al. Expression of Somatostatin Receptor Subtype 2 in Growth Hormone-Secreting Pituitary Adenoma and the Regulation of MiR-185. J. Endocrinol. Investig. 2015, 38, 1117–1128. [Google Scholar] [CrossRef]
- Krützfeldt, J. Strategies to Use MicroRNAs as Therapeutic Targets. Best Pract. Res. Clin. Endocrinol. Metab. 2016, 30, 551–561. [Google Scholar] [CrossRef]
- Creemers, E.E.; Tijsen, A.J.; Pinto, Y.M. Circulating MicroRNAs: Novel Biomarkers and Extracellular Communicators in Cardiovascular Disease? Circ. Res. 2012, 110, 483–495. [Google Scholar] [CrossRef]
- Zhao, C.; Sun, X.; Li, L. Biogenesis and Function of Extracellular MiRNAs. ExRNA 2019, 1, 38. [Google Scholar] [CrossRef] [Green Version]
- Allegra, A.; Alonci, A.; Campo, S.; Penna, G.; Petrungaro, A.; Gerace, D.; Musolino, C. Circulating MicroRNAs: New Biomarkers in Diagnosis, Prognosis and Treatment of Cancer (Review). Int. J. Oncol. 2012, 41, 1897–1912. [Google Scholar] [CrossRef] [Green Version]
- Michael, M.Z.; O’ Connor, S.M.; van Holst Pellekaan, N.G.; Young, G.P.; James, R.J. Reduced Accumulation of Specific MicroRNAs in Colorectal Neoplasia. Mol. Cancer Res. 2003, 1, 882–891. [Google Scholar]
- Ntali, G.; Wass, J.A. Epidemiology, Clinical Presentation and Diagnosis of Non-Functioning Pituitary Adenomas. Pituitary 2018, 21, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Shen, J.; Medico, L.; Wang, D.; Ambrosone, C.B.; Liu, S. A Pilot Study of Circulating MiRNAs as Potential Biomarkers of Early Stage Breast Cancer. PLoS ONE 2010, 5, e13735. [Google Scholar] [CrossRef]
- Duttagupta, R.; Jiang, R.; Gollub, J.; Getts, R.C.; Jones, K.W. Impact of Cellular MiRNAs on Circulating MiRNA Biomarker Signatures. PLoS ONE 2011, 6, e20769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-T.; Tsai, P.-C.; Liao, Y.-C.; Hsu, C.-Y.; Juo, S.-H.H. Circulating MicroRNAs Have a Sex-Specific Association with Metabolic Syndrome. J. Biomed. Sci. 2013, 20, 72. [Google Scholar] [CrossRef] [Green Version]
- Badrnya, S.; Baumgartner, R.; Assinger, A. Smoking Alters Circulating Plasma Microvesicle Pattern and MicroRNA Signatures. Thromb. Haemost. 2014, 112, 128–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoi, W.; Sakuma, K. Does Regulation of Skeletal Muscle Function Involve Circulating MicroRNAs? Front. Physiol. 2014, 5, 39. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.A.; Davis, C.D. MicroRNA, Nutrition, and Cancer Prevention1. Adv. Nutr. 2011, 2, 472–485. [Google Scholar] [CrossRef]
- Witwer, K.W. XenomiRs and MiRNA Homeostasis in Health and Disease. RNA Biol. 2012, 9, 1147–1154. [Google Scholar] [CrossRef]
- De Boer, H.C.; van Solingen, C.; Prins, J.; Duijs, J.M.G.J.; Huisman, M.V.; Rabelink, T.J.; van Zonneveld, A.J. Aspirin Treatment Hampers the Use of Plasma MicroRNA-126 as a Biomarker for the Progression of Vascular Disease. Eur. Heart J. 2013, 34, 3451–3457. [Google Scholar] [CrossRef] [PubMed]
- Donati, S.; Ciuffi, S.; Brandi, M.L. Human Circulating MiRNAs Real-Time QRT-PCR-Based Analysis: An Overview of Endogenous Reference Genes Used for Data Normalization. Int. J. Mol. Sci. 2019, 20, 4353. [Google Scholar] [CrossRef] [PubMed] [Green Version]


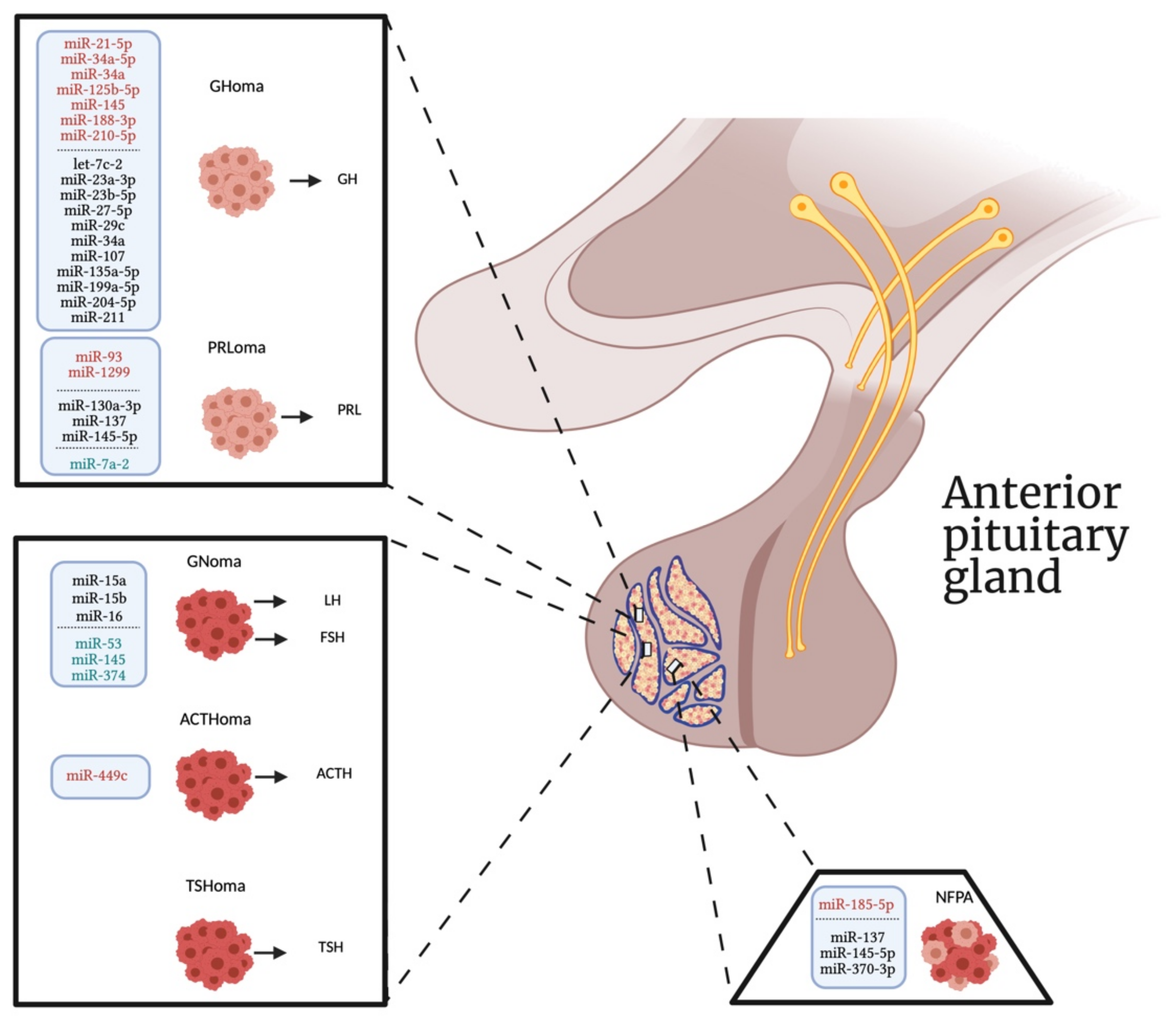{kind=link}
| miRNA | Expression Levels | miRNA Profiling Platform | Biological Function | Target Genes | Reference |
|---|---|---|---|---|---|
| miR-185-5p | ↑ | NGS, qPCR | Status of invasive growth not confirmed by ROC analysis | / 1 | [40] |
| miR-137 | ↓ | qPCR | Regulates Wnt signaling pathway | WIF1 | [43] |
| miR-370-3p | ↓ | RT-PCR | Increases cell proliferation and invasiveness | HMGA2 | [45] |
| miR-145-5p | ↓ | qPCR | Increases cell proliferation and invasiveness | TPT1 | [46] |
| miRNA | Expression Levels | miRNA Profiling Platform | Biological Function | Target Genes | Reference |
|---|---|---|---|---|---|
| miR-145, miR-53, and miR-374 | / 1 | RNA-sequencing | Contribute to tumor development | DEGs 2 mainly enriched in cell cycle and neuroactive ligand-receptor interaction | [47] |
| miR-15a, miR-15b, and miR-16 | ↓ | qPCR | Contributes to pituitary tumorigenesis | HMGA1 and HMGA2 | [49] |
| miRNA | Expression Levels | miRNA Profiling Platform | Biological Function | Target Genes | Reference |
|---|---|---|---|---|---|
| miR-7a2 | / 1 | qPCR | Regulates prolactin production | Raf1 | [50] |
| miR-93 | ↑ | qPCR | Mediates CAB resistance | ATG7 | [51,52] |
| miR-93-5p | ↑ | RNA-sequencing, PCR | Regulates TGF-β1/Smad3 signaling-mediated fibrosis | Smad7 | [53] |
| miR-1299 | ↑ | RNA-sequencing, qPCR | Regulates PRL gene transcription participating in the drug resistance | FOXO1 | [54] |
| miR-145-5p | ↓ | qPCR | Regulates drug resistance | TPT1 | [55] |
| miR-130a-3p | ↓ | Microarray, qPCR | Role in prolactin regulation | ERα | [56] |
| miR-137 | ↓ | Microarray, qPCR | Regulates Wnt-β-catenin signaling pathway | MITF | [57] |
| miRNA | Expression Levels | miRNA Profiling Platform | Biological Function | Target Genes | Reference |
|---|---|---|---|---|---|
| Exosomal miR-21-5p | ↑ | qPCR | Regulates osteoblast proliferation, collagen I and osteocalcin synthesis, and bone formation | / 1 | [58] |
| miR-34a and miR-145 | ↑ | Microarray, qPCR | Regulates cell proliferation and GH secretion in vitro, but also mediates resistance to the antiproliferative and hormonal properties of octreotide | Gnai2 | [60] |
| miR-125b-5p, miR-34a-5p, miR-188-3p, miR-210-5p, miR-27-5p miR-135a-5p, miR-199a-5p miR-211, miR-23a-3p, and miR-204-5p | ↑ ↑ ↑ ↑ ↓ ↓ ↓ ↓ ↓ ↓ | qPCR | Involvement in mesenchymal stem cell commitment | / 1 | [61] |
| miR-23b and miR-107 | ↓ | qPCR | / 1 | HMGA2 and AIP | [62] |
| let-7c-2, miR-23b, and miR-29c | ↓ | qPCR | Promotes cell proliferation | HMGA, IGF-1, and N-MYC | [63] |
| miR-34a | ↓ | qPCR | Promotes cell proliferation and inhibits cell apoptosis | SOX7 | [64] |
| miRNA | Expression Levels | miRNA Profiling Platform | Biological Function | Target Genes | Reference |
|---|---|---|---|---|---|
| miR-449c | ↑ | qPCR | Regulates POMC transcription, ACTH synthesis, cells proliferation, migration, and invasion | TSP-1 | [66] |
| miRNA | Expression Levels | miRNA Profiling Platform | Biological Function | Target Genes | Reference |
|---|---|---|---|---|---|
| miR-181c-5p, and miR-454-3p | / 1 | Microarray, qPCR | Regulate TNFα signaling pathway | / ¹ | [67] |
| miR-543 | ↑ | qPCR | Regulates cell proliferation, migration, invasion, apoptosis, and Wnt/β-catenin signaling pathway | Smad7 | [68] |
| 55 miRNAs | 31 ↑ and 24 ↓ | RNA sequencing | Regulate the invasive behavior | DEGs 2 mainly enriched in cell proliferation and cell cycle pathway | [69] |
| miR-410-3p | ↑ or ↓ | qPCR | Regulates MAPK, PTEN/AKT, and STAT3 signaling pathways | / ¹ | [70] |
| miR-145, miR-124 and miR-183 | ↓ | Microarray, qPCR | Regulate the migration and invasion | FSCN1, PTTG1IP, and EZR | [71] |
| miR-302/372/373/520 | ↓ | qPCR | Regulate cell proliferation, apoptosis, migration, invasion, EMT, and tumor growth | TGFBR2 and RAB11A | [72] |
| miR-376B-3P | ↓ | qPCR | Regulates tumor invasiveness | HMGA2 | [73] |
| Exosomal miR-99a-3p and miR-149-5p | ↓ | qPCR | Regulate growth and metastasis of pituitary adenoma cells | NOVA1, DTL and RAB27B | [74] |
| miR-137 | ↓ | qPCR | Promotes cell proliferation and invasion | AKT2 | [75] |
| miR-139-3p | ↓ | qPCR | Promotes cell viability, proliferation, migration, invasion and inhibits apoptosis | BRD4 | [76] |
| miR-424-5p | ↓ | qPCR | Promotes cell proliferation, migration, invasion and inhibits apoptosis | bFGF | [77] |
| PitNET Types | miRNA | Expression Levels | miRNA Profiling Platform | Biological Function | Target Genes | Reference |
|---|---|---|---|---|---|---|
| GT 2, CT 3, sCT 4, ST 5 | miR-17-5p | ↑ | qPCR | Involvement in tumor growth and invasiveness | / 1 | [80] |
| GH 6, NF 7, PRL 8 | miR-184 miR-34c-3p miR-34b-5p, miR-378, miR-338-5p, miR-124-3p | ↑ ↓ ↓ ↓ ↓ ↓ | NGS, qPCR | / ¹ | / ¹ | [81] |
| / 1 | miR-1 | ↓ | qPCR | Promotes cell proliferation, inhibits cell apoptosis, and glucose metabolism of cancer cells | G6PD | [82] |
| GH 6, PRL 8 | miR-205-5p | ↓ | qPCR | Contributes cell proliferation and migration | CBX1 | [87] |
| GH 6, PRL 8 | miR-103a-3p | ↓ | qPCR | Promotes rat pituitary adenoma cell proliferation and PI3K/AKT signaling pathway, inhibits cell apoptosis and PRL and GH secretion | / ¹ | [89] |
| GT 2, ACTH 9 | miR-219a-2-3p | ↓ | qPCR | Promotes cell proliferation and inhibits cell apoptosis | MDM2 | [90] |
| MEN1-related 10 | miR-15a, miR-16, and let-7a | ↓ | qPCR | Regulate CCND1 expression | / ¹ | [92] |
| Biological Fluid | miRNA | Expression Levels | miRNA Profiling Platform | Potential Use as Biomarker | Reference |
|---|---|---|---|---|---|
| Plasma | miR-16-5p, mir-145-5p, and miR-7g-5p | ↑ | qPCR | Distinguishing between CD 1 and EAS 2 patients | [101] |
| Plasma | miR-143-3p | ↓ | NGS, qPCR | Patient FSH/LH+ follow up | [102] |
| Serum | miR-16 | ↓ | qPCR | Correlation with longer OS 3 and DFS 4 in pituitary tumor patients | [105] |
| Serum | miR-423-5p | ↓ | Microarray, NGS, qPCR | Clinical treatment of somatotroph adenomas | [106] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donati, S.; Aurilia, C.; Palmini, G.; Miglietta, F.; Falsetti, I.; Iantomasi, T.; Brandi, M.L. MicroRNAs as Potential Biomarkers in Pituitary Adenomas. Non-Coding RNA 2021, 7, 55. https://doi.org/10.3390/ncrna7030055
Donati S, Aurilia C, Palmini G, Miglietta F, Falsetti I, Iantomasi T, Brandi ML. MicroRNAs as Potential Biomarkers in Pituitary Adenomas. Non-Coding RNA. 2021; 7(3):55. https://doi.org/10.3390/ncrna7030055
Chicago/Turabian StyleDonati, Simone, Cinzia Aurilia, Gaia Palmini, Francesca Miglietta, Irene Falsetti, Teresa Iantomasi, and Maria Luisa Brandi. 2021. "MicroRNAs as Potential Biomarkers in Pituitary Adenomas" Non-Coding RNA 7, no. 3: 55. https://doi.org/10.3390/ncrna7030055
APA StyleDonati, S., Aurilia, C., Palmini, G., Miglietta, F., Falsetti, I., Iantomasi, T., & Brandi, M. L. (2021). MicroRNAs as Potential Biomarkers in Pituitary Adenomas. Non-Coding RNA, 7(3), 55. https://doi.org/10.3390/ncrna7030055