
Hsa-miR-183-5p Modulates Cell Adhesion by Repression of ITGB1 Expression in Prostate Cancer

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Expression of MiR-183 Is Increased in Tumor Tissue and Associates with Worse Clinical Features in PrCa
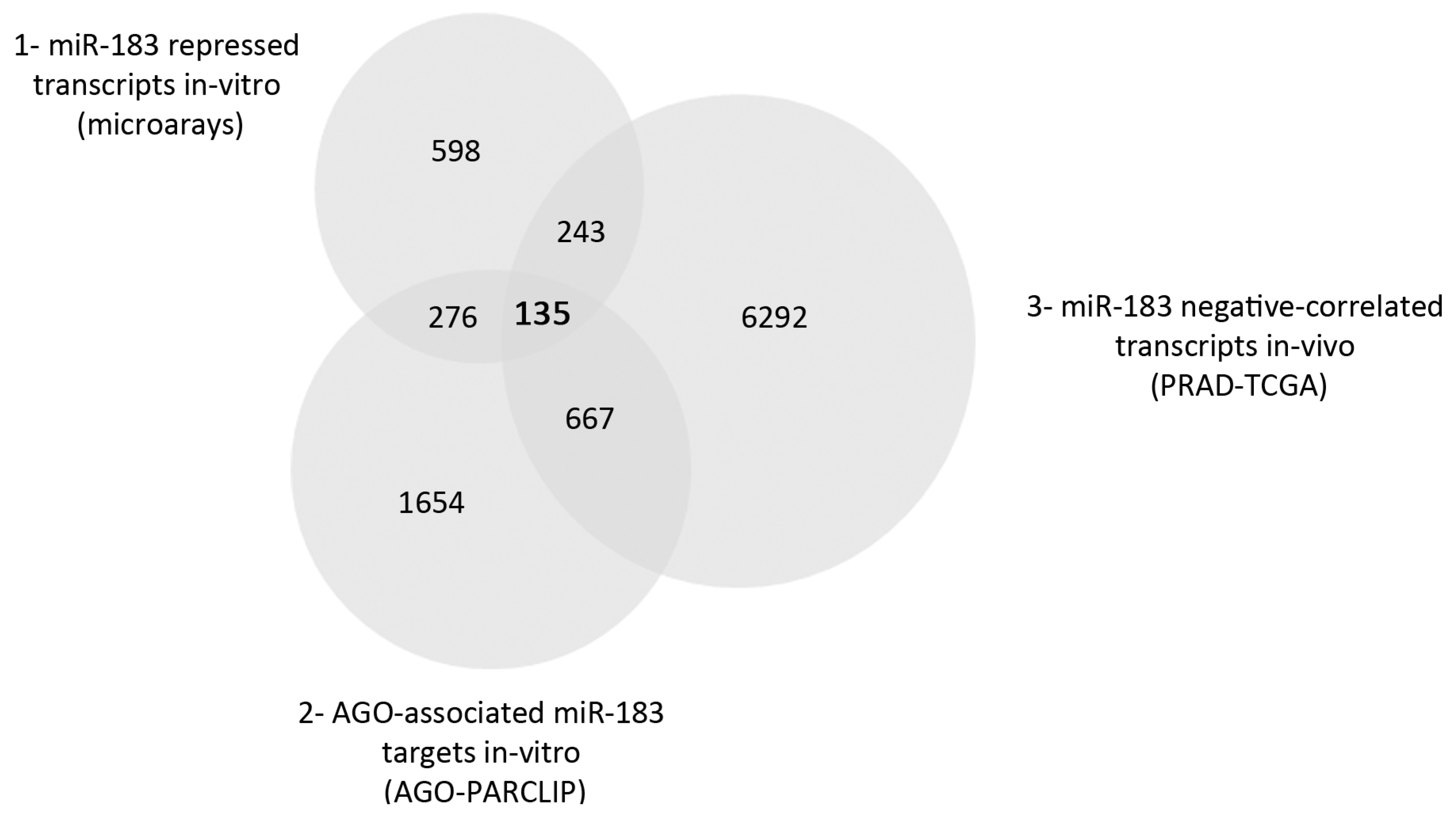
2.2. Direct Candidate Target Genes of MiR-183 Are Related to Cell Adhesion
2.3. Overexpression of MiR-183 in Prostate Cells Causes a Decrease in Cell Adhesion In Vitro
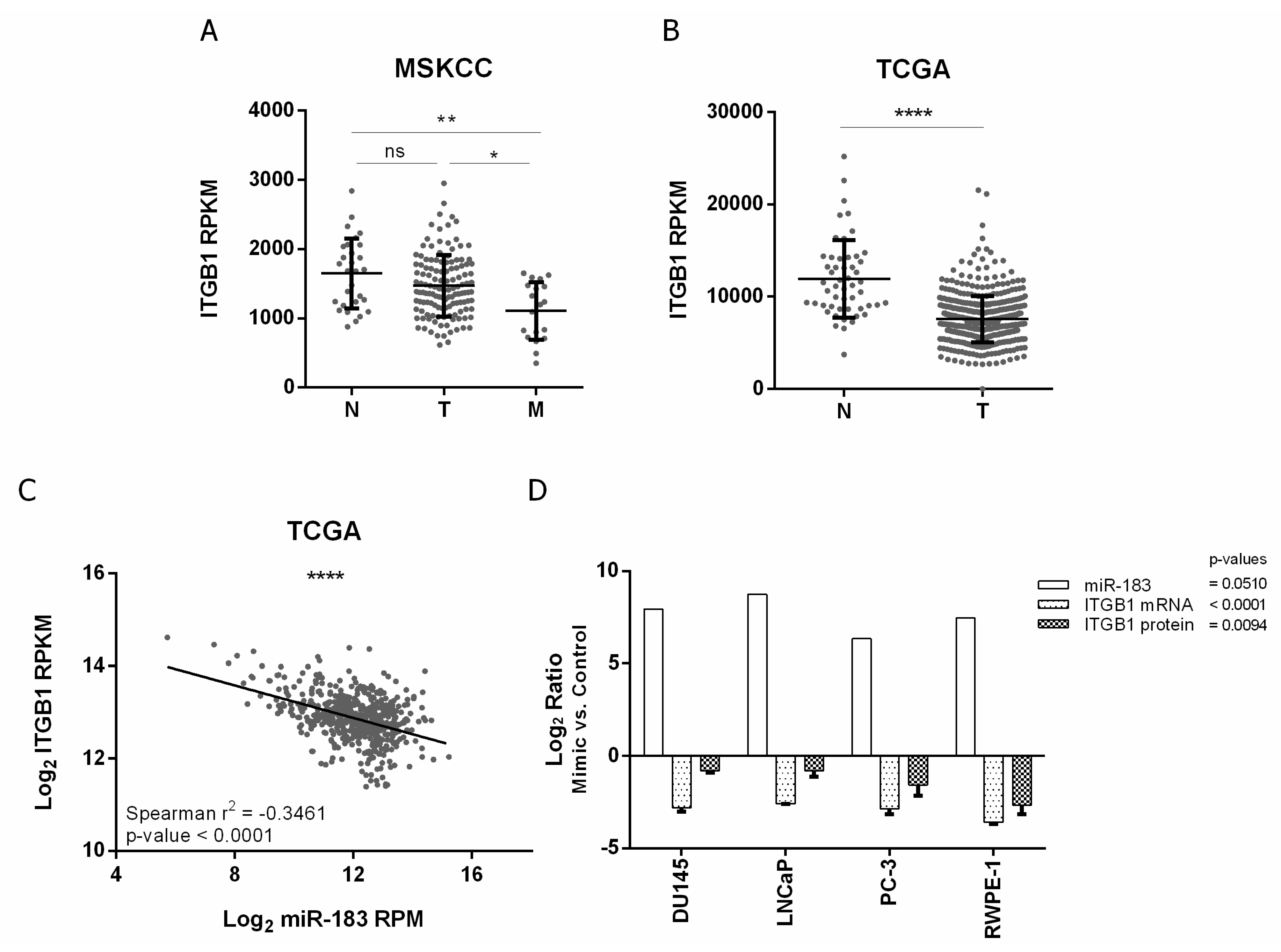
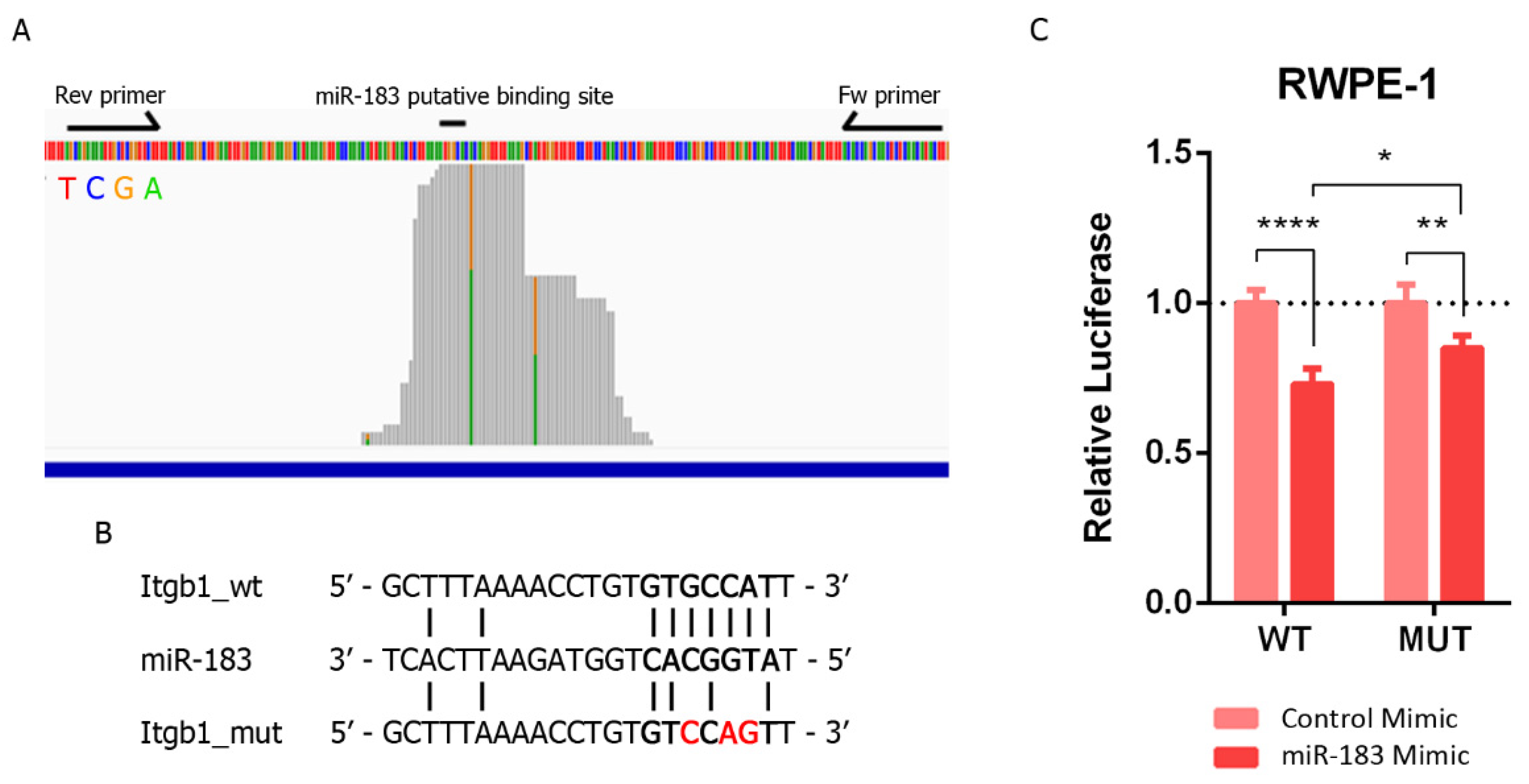
2.4. ITGB1 Regulation by MiR-183 May by Mediated by a Direct Interaction with the ITGB1 3′UTR in Prostate Cancer
2.5. The Inhibition of Cell Adhesion Provoked by miR-183 May Be Due to a Reduction of Focal Adhesions Mediated by ITGB1 Downregulation
3. Discussion
4. Materials and Methods
4.1. Human Specimens
4.2. Cell Lines
4.3. Cell Ttransfection
4.3.1. Microarrays Experiments
4.3.2. Functional Analysis
4.3.3. Reporter Gene Assays
4.4. Gene Expression Microarrays
4.5. Dataset Analysis
4.6. Biological Term Enrichment Analysis of the Direct Candidate Targets Genes of MiR-183
4.7. Cell Adhesion Assays
4.7.1. Quantitative Assessment
4.7.2. Qualitative Assessment
4.8. RNA Extraction, Reverse Transcription and Quantitative Real-Time PCR
4.9. Flow Cytometry for ITGB1 Quantification
4.10. Luciferase Reporter Gene Assay
4.11. ITGB1 Blockade
4.12. Focal Adhesion Quantification
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian MicroRNAs Predominantly Act to Decrease Target MRNA Levels. Nature 2010, 466, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Larsson, O.; Nadon, R. Re-Analysis of Genome Wide Data on Mammalian MicroRNA-Mediated Suppression of Gene Expression. Translation 2013, 1, e24557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambs, S.; Prueitt, R.L.; Yi, M.; Hudson, R.S.; Howe, T.M.; Petrocca, F.; Wallace, T.A.; Liu, C.G.; Volinia, S.; Calin, G.A.; et al. Genomic Profiling of MicroRNA and Messenger RNA Reveals Deregulated MicroRNA Expression in Prostate Cancer. Cancer Res. 2008, 68, 6162–6170. [Google Scholar] [CrossRef] [Green Version]
- Porkka, K.P.; Pfeiffer, M.J.; Waltering, K.K.; Vessella, R.L.; Tammela, T.L.J.; Visakorpi, T. MicroRNA Expression Profiling in Prostate Cancer. Cancer Res. 2007, 67, 6130–6135. [Google Scholar] [CrossRef] [Green Version]
- Ozen, M.; Creighton, C.J.; Ozdemir, M.; Ittmann, M. Widespread Deregulation of MicroRNA Expression in Human Prostate Cancer. Oncogene 2008, 27, 1788–1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA Expression Profiles Classify Human Cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent Deletions and Down-Regulation of Micro-RNA Genes MiR15 and MiR16 at 13q14 in Chronic Lymphocytic Leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.; Peruzzi, P.P.; Lawler, S. MicroRNAs in Cancer: Biomarkers, Functions and Therapy. Trends Mol. Med. 2014, 20, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Witmer, P.D.; Lumayag, S.; Kovacs, B.; Valle, D. MicroRNA (MiRNA) Transcriptome of Mouse Retina and Identification of a Sensory Organ-Specific MiRNA Cluster. J. Biol. Chem. 2007, 282, 25053–25066. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.; Di, M.; Liang, J.; Shi, S.; Tan, Q.; Wang, Z. MicroRNA-183 in Cancer Progression. J. Cancer 2020, 11, 1315–1324. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Hao, X.; Song, Y. Identification of the Key MicroRNAs and the MiRNA-MRNA Regulatory Pathways in Prostate Cancer by Bioinformatics Methods. Biomed Res. Int. 2018, 2018, 6204128. [Google Scholar] [CrossRef]
- Ueno, K.; Hirata, H.; Shahryari, V.; Deng, G.; Tanaka, Y.; Tabatabai, Z.L.; Hinoda, Y.; Dahiya, R. MicroRNA-183 Is an Oncogene Targeting Dkk-3 and SMAD4 in Prostate Cancer. Br. J. Cancer 2013, 108, 1659–1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, A.; Jung, M.; Mollenkopf, H.-J.; Wagner, I.; Stephan, C.; Jentzmik, F.; Miller, K.; Lein, M.; Kristiansen, G.; Jung, K. Diagnostic and Prognostic Implications of MicroRNA Profiling in Prostate Carcinoma. Int. J. Cancer 2009, 126, 1166–1176. [Google Scholar] [CrossRef]
- Waseem, M.; Ahmad, M.K.; Serajuddin, M.; Bhaskar, V.; Sankhwar, S.N.; Mahdi, A.A. MicroRNA-183-5p: A New Potential Marker for Prostate Cancer. Indian J. Clin. Biochem. 2019, 34, 207–212. [Google Scholar] [CrossRef]
- Martens-Uzunova, E.S.; Jalava, S.E.; Dits, N.F.; Van Leenders, G.J.L.H.; Møller, S.; Trapman, J.; Bangma, C.H.; Litman, T.; Visakorpi, T.; Jenster, G. Diagnostic and Prognostic Signatures from the Small Non-Coding RNA Transcriptome in Prostate Cancer. Oncogene 2012, 31, 978–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, B.; Reis, S.T.; Viana, N.I.; Morais, D.R.; Moura, C.M.; Dip, N.; Silva, I.A.; Iscaife, A.; Srougi, M.; Leite, K.R.M. Comprehensive Study of Gene and MicroRNA Expression Related to Epithelial-Mesenchymal Transition in Prostate Cancer. PLoS ONE 2014, 9, e113700. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Lin, S.; Cheng, C.; Zhu, A.; Hu, Y.; Shi, Z.; Zhang, X.; Hong, Z. Long Non-Coding RNA CASC2 Regulates Sprouty2 via Functioning as a Competing Endogenous RNA for MiR-183 to Modulate the Sensitivity of Prostate Cancer Cells to Docetaxel. Arch. Biochem. Biophys. 2019, 665, 69–78. [Google Scholar] [CrossRef]
- Hua, X.; Liu, Z.; Zhou, M.; Tian, Y.; Zhao, P.P.; Pan, W.H.; Li, C.X.; Huang, X.X.; Liao, Z.X.; Xian, Q.; et al. LSAMP-AS1 Binds to MicroRNA-183–5p to Suppress the Progression of Prostate Cancer by up-Regulating the Tumor Suppressor DCN. EBioMedicine 2019, 50, 178–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihelich, B.L.; Khramtsova, E.A.; Arva, N.; Vaishnav, A.; Johnson, D.N.; Giangreco, A.A.; Martens-Uzunova, E.; Bagasra, O.; Kajdacsy-Balla, A.; Nonn, L. MiR-183-96-182 Cluster Is Overexpressed in Prostate Tissue and Regulates Zinc Homeostasis in Prostate Cells. J. Biol. Chem. 2011, 286, 44503–44511. [Google Scholar] [CrossRef] [Green Version]
- Paziewska, A.; Mikula, M.; Dabrowska, M.; Kulecka, M.; Goryca, K.; Antoniewicz, A.; Dobruch, J.; Borowka, A.; Rutkowski, P.; Ostrowski, J. Candidate Diagnostic MiRNAs That Can Detect Cancer in Prostate Biopsy. Prostate 2018, 78, 178–185. [Google Scholar] [CrossRef]
- Song, C.J.; Chen, H.; Chen, L.Z.; Ru, G.M.; Guo, J.J.; Ding, Q.N. The Potential of MicroRNAs as Human Prostate Cancer Biomarkers: A Meta-Analysis of Related Studies. J. Cell. Biochem. 2018, 119, 2763–2786. [Google Scholar] [CrossRef] [Green Version]
- Larne, O.; Martens-Uzunova, E.; Hagman, Z.; Edsjö, A.; Lippolis, G.; Den Berg, M.S.V.V.; Bjartell, A.; Jenster, G.; Ceder, Y. MiQ—A Novel MicroRNA Based Diagnostic and Prognostic Tool for Prostate Cancer. Int. J. Cancer 2013, 132, 2867–2875. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Gao, X. Inhibition of Cancer Cell-Derived Exosomal MicroRNA-183 Suppresses Cell Growth and Metastasis in Prostate Cancer by Upregulating TPM1. Cancer Cell Int. 2021, 21. [Google Scholar] [CrossRef]
- Dambal, S.; Baumann, B.; McCray, T.; Williams, L.T.; Richards, Z.; Deaton, R.; Prins, G.S.; Nonn, L. The MiR-183 Family Cluster Alters Zinc Homeostasis in Benign Prostate Cells, Organoids and Prostate Cancer Xenografts. Sci. Rep. 2017, 7, 7704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larne, O.; Östling, P.; Haflidadóttir, B.S.; Hagman, Z.; Aakula, A.; Kohonen, P.; Kallioniemi, O.; Edsjö, A.; Bjartell, A.; Lilja, H.; et al. MIR-183 in Prostate Cancer Cells Positively Regulates Synthesis and Serum Levels of Prostate-Specific Antigen. Eur. Urol. 2015, 68, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Taylor, B.S.; Schultz, N.; Hieronymus, H.; Gopalan, A.; Xiao, Y.; Carver, B.S.; Arora, V.K.; Kaushik, P.; Cerami, E.; Reva, B.; et al. Integrative Genomic Profiling of Human Prostate Cancer. Cancer Cell 2010, 18, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Wong, N.W.; Chen, Y.; Chen, S.; Wang, X. OncomiR: An Online Resource for Exploring Pan-Cancer MicroRNA Dysregulation. Bioinformatics 2018, 34, 713–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, D.W.; Bracken, C.P.; Szubert, J.M.; Goodall, G.J. On Measuring MiRNAs after Transient Transfection of Mimics or Antisense Inhibitors. PLoS ONE 2013, 8, 55214. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, M.P.; Rajapakshe, K.I.; Bader, D.A.; Cerne, J.Z.; Smith, E.A.; Coarfa, C.; Hartig, S.M.; McGuire, S.E. The Landscape of MicroRNA Targeting in Prostate Cancer Defined by AGO-PAR-CLIP. Neoplasia 2016, 18, 356–370. [Google Scholar] [CrossRef] [Green Version]
- McGeary, S.E.; Lin, K.S.; Shi, C.Y.; Pham, T.M.; Bisaria, N.; Kelley, G.M.; Bartel, D.P. The Biochemical Basis of MicroRNA Targeting Efficacy. Science 2019, 366, eaav1741. [Google Scholar] [CrossRef]
- Witwer, K.W.; Halushka, M.K. Toward the Promise of MicroRNAs—Enhancing Reproducibility and Rigor in MicroRNA Research. RNA Biol. 2016, 13, 1103–1116. [Google Scholar] [CrossRef] [PubMed]
- La Rocca, G.; Olejniczak, S.H.; González, A.J.; Briskin, D.; Vidigal, J.A.; Spraggon, L.; DeMatteo, R.G.; Radler, M.R.; Lindsten, T.; Ventura, A.; et al. In Vivo, Argonaute-Bound MicroRNAs Exist Predominantly in a Reservoir of Low Molecular Weight Complexes Not Associated with MRNA. Proc. Natl. Acad. Sci. USA 2015, 112, 767–772. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.Y.; Gonzalez-Martin, A.; Miletic, A.V.; Lai, M.; Knight, S.; Sabouri-Ghomi, M.; Head, S.R.; Macauley, M.S.; Rickert, R.C.; Xiao, C. Transfection of MicroRNA Mimics Should Be Used with Caution. Front. Genet. 2015, 6, 340. [Google Scholar] [CrossRef] [Green Version]
- Andl, T.; Ganapathy, K.; Bossan, A.; Chakrabarti, R. MicroRNAs as Guardians of the Prostate: Those Who Stand before Cancer. What Do We Really Know about the Role of MicroRNAs in Prostate Biology? Int. J. Mol. Sci. 2020, 21, 4796. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and Collaborative HTML5 Gene List Enrichment Analysis Tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moro, L.; Dolce, L.; Cabodi, S.; Bergatto, E.; Erba, E.B.; Smeriglio, M.; Turco, E.; Retta, S.F.; Giuffrida, M.G.; Venturino, M.; et al. Integrin-Induced Epidermal Growth Factor (EGF) Receptor Activation Requires c-Src and P130Cas and Leads to Phosphorylation of Specific EGF Receptor Tyrosines. J. Biol. Chem. 2002, 277, 9405–9414. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting Effective MicroRNA Target Sites in Mammalian MRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Hafner, M.; Landthaler, M.; Burger, L.; Khorshid, M.; Hausser, J.; Berninger, P.; Rothballer, A.; Ascano, M.; Jungkamp, A.C.; Munschauer, M.; et al. Transcriptome-Wide Identification of RNA-Binding Protein and MicroRNA Target Sites by PAR-CLIP. Cell 2010, 141, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative Genomics Viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnapp, L.M.; Hatch, N.; Ramos, D.M.; Klimanskaya, I.V.; Sheppard, D.; Pytela, R. The Human Integrin A8β1 Functions as a Receptor for Tenascin, Fibronectin, and Vitronectin. J. Biol. Chem. 1995, 270, 23196–23202. [Google Scholar] [CrossRef] [Green Version]
- Berrier, A.L.; Yamada, K.M. Cell–Matrix Adhesion. J. Cell. Physiol. 2007, 213, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Kuo, J.-C. Focal Adhesions Function as a Mechanosensor. Mechanotransduction 2014, 126, 55–73. [Google Scholar] [CrossRef]
- Zhong, X.; Rescorla, F.J. Cell Surface Adhesion Molecules and Adhesion-Initiated Signaling: Understanding of Anoikis Resistance Mechanisms and Therapeutic Opportunities. Cell. Signal. 2012, 24, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Wu, J.; Gao, Z.; Yu, S.; Cui, Y. Screening the Key MicroRNAs and Transcription Factors in Prostate Cancer Based on MicroRNA Functional Synergistic Relationships. Medicine 2017, 96, e5679. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Sun, H.M.; Zheng, R.Z.; Li, Y.C.; Zhang, Q.; Cheng, P.; Tang, Z.H.; Huang, F. Meta-Analysis of MicroRNA-183 Family Expression in Human Cancer Studies Comparing Cancer Tissues with Noncancerous Tissues. Gene 2013, 527, 26–32. [Google Scholar] [CrossRef]
- Gorlov, I.P.; Byun, J.; Gorlova, O.Y.; Aparicio, A.M.; Efstathiou, E.; Logothetis, C.J. Candidate pathways and genes for prostate cancer: A meta-analysis of gene expression data. BMC Med. Genom. 2009, 2, 48. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Li, S.-L.; Wang, S.-Y. Construction and analysis of mRNA, miRNA, lncRNA, and TF regulatory networks reveal the key genes associated with prostate cancer. PLoS ONE 2018, 13, e0198055. [Google Scholar] [CrossRef] [Green Version]
- Bianconi, D.; Unseld, M.; Prager, G.W. Integrins in the Spotlight of Cancer. Int. J. Mol. Sci. 2016, 17, 2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedl, P. Prespecification and Plasticity: Shifting Mechanisms of Cell Migration. Curr. Opin. Cell Biol. 2004, 16, 14–23. [Google Scholar] [CrossRef]
- Fort, R.S.; Mathó, C.; Oliveira-Rizzo, C.; Garat, B.; Sotelo-Silveira, J.R.; Duhagon, M.A. An Integrated View of the Role of MiR-130b/301b MiRNA Cluster in Prostate Cancer. Exp. Hematol. Oncol. 2018, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Papatheodorou, I.; Moreno, P.; Manning, J.; Fuentes, A.M.P.; George, N.; Fexova, S.; Fonseca, N.A.; Füllgrabe, A.; Green, M.; Huang, N.; et al. Expression Atlas Update: From Tissues to Single Cells. Nucleic Acids Res. 2020, 48, D77–D83. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhang, M.; Liu, L.; Jin, D.; Wang, P.; Hu, J. MicroRNA-183-5p Inhibits Aggressiveness of Cervical Cancer Cells by Targeting Integrin Subunit Beta 1 (ITGB1). Med. Sci. Monit. 2018, 24, 7137–7145. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Gu, L.; Ni, J.; Hu, P.; Hu, K.; Shi, Y.L. MIR-183 Regulates ITGB1P Expression and Promotes Invasion of Endometrial Stromal Cells. Biomed Res. Int. 2015, 2015, 340218. [Google Scholar] [CrossRef]
- Sun, Y.; Cheng, H.; Wang, G.; Yu, G.; Zhang, D.; Wang, Y.; Fan, W.; Yang, W. Deregulation of MiR-183 Promotes Melanoma Development via LncRNA MALAT1 Regulation and ITGB1 Signal Activation. Oncotarget 2017, 8, 3509–3518. [Google Scholar] [CrossRef] [Green Version]
- Cao, C.; Han, S.; Yuan, Y.; Wu, Y.; Lian, W.; Zhang, X.; Pan, L.; Li, M. Downregulated Circular RNA Hsa_circ_0000291 Suppresses Migration and Proliferation of Gastric Cancer via Targeting the MiR-183/ITGB1 Axis. Cancer Manag. Res. 2019, 11, 9675–9683. [Google Scholar] [CrossRef] [Green Version]
- Kurozumi, A.; Goto, Y.; Matsushita, R.; Fukumoto, I.; Kato, M.; Nishikawa, R.; Sakamoto, S.; Enokida, H.; Nakagawa, M.; Ichikawa, T.; et al. Tumor-Suppressive MicroRNA-223 Inhibits Cancer Cell Migration and Invasion by Targeting ITGA3/ITGB1 Signaling in Prostate Cancer. Cancer Sci. 2016, 107, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Pellinen, T.; Blom, S.; Sánchez, S.; Välimäki, K.; Mpindi, J.P.; Azegrouz, H.; Strippoli, R.; Nieto, R.; Vitón, M.; Palacios, I.; et al. ITGB1-Dependent Upregulation of Caveolin-1 Switches TGFβ Signalling from Tumour-Suppressive to Oncogenic in Prostate Cancer. Sci. Rep. 2018, 8, 2338. [Google Scholar] [CrossRef]
- Broustas, C.G.; Lieberman, H.B. RAD9 Enhances Radioresistance of Human Prostate Cancer Cells through Regulation of ITGB1 Protein Levels. Prostate 2014, 74, 1359–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, N.E.; Zhang, Z.; Madamanchi, A.; Boyd, K.L.; O’Rear, L.D.; Nashabi, A.; Li, Z.; Dupont, W.D.; Zijlstra, A.; Zutter, M.M. The A₂β₁ Integrin Is a Metastasis Suppressor in Mouse Models and Human Cancer. J. Clin. Investig. 2011, 121, 226–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varzavand, A.; Hacker, W.; Ma, D.; Gibson-Corley, K.; Hawayek, M.; Tayh, O.J.; Brown, J.A.; Henry, M.D.; Stipp, C.S. A3β1 Integrin Suppresses Prostate Cancer Metastasis via Regulation of the Hippo Pathway. Cancer Res. 2016, 76, 6577–6587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallio, M.A.; Tuimala, J.T.; Hupponen, T.; Klemelä, P.; Gentile, M.; Scheinin, I.; Koski, M.; Käki, J.; Korpelainen, E.I. Chipster: User-Friendly Analysis Software for Microarray and Other High-Throughput Data. BMC Genom. 2011, 12, 507. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI Gene Expression and Hybridization Array Data Repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodama, Y.; Shumway, M.; Leinonen, R. The Sequence Read Archive: Explosive Growth of Sequencing Data. Nucleic Acids Res. 2012, 40, D54–D56. [Google Scholar] [CrossRef] [Green Version]
- Andrés-León, E.; Núñez-Torres, R.; Rojas, A.M. MiARma-Seq: A Comprehensive Tool for MiRNA, MRNA and CircRNA Analysis. Sci. Rep. 2016, 6, 25749. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq-A Python Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- UCSC. Xena Browser. Available online: https://xenabrowser.net/ (accessed on 23 April 2020).
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental Algorithms for Scientific Computing in Python. Nat. Methods 2020, 17, 261. [Google Scholar] [CrossRef] [Green Version]
- Carbon, S.; Ireland, A.; Mungall, C.J.; Shu, S.; Marshall, B.; Lewis, S.; Lomax, J.; Mungall, C.; Hitz, B.; Balakrishnan, R.; et al. AmiGO: Online Access to Ontology and Annotation Data. Bioinformatics 2009, 25, 288–289. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bohmer, M.; Sharbati, J.; Zur Bruegge, J.; Einspanier, R.; Sharbati, S. Structural Analysis of MicroRNA-Target Interaction by Sequential Seed Mutagenesis and Stem-Loop 3′ RACE. PLoS ONE 2013, 8, e81427. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort | Clinical Parameter | Conditions Compared | Ratio between Conditions | p-Value |
|---|---|---|---|---|
| MSKCC | Tissue type | Primary tumor vs. Normal | 3.3 ± 2.4 | <0.0001 |
| Metastasis vs. Normal | 5.3 ± 4.1 | <0.0001 | ||
| PSA at diagnosis | High vs. Low | 1.5 ± 1.1 | 0.0182 | |
| Preoperative PSA | High vs. Low | 2.5 ± 1.9 | <0.0001 | |
| Time until recurrence (month) | Shorter vs. Longer | 1.3 ± 1.9 | 0.0195 | |
| PRAD-TCGA | Tissue type | Primary tumor vs. Normal | 4.3 ± 3.0 | <0.0001 |
| Biochemical recurrence | YES vs. NO | 1.3 ± 1.1 | 0.0404 | |
| Clinical T | T3-T4 vs. T1-T2 | 1.5 ± 1.3 | 0.0020 | |
| Gleason Score | 8-9-10 vs. 6-7 | 1.4 ± 1.0 | <0.0001 | |
| Pathologic N | N1 vs. N0 | 1.3 ± 0.9 | 0.0030 |
| Wikipathways | ||
|---|---|---|
| Term | Adjusted p-Value | Genes |
| EGF/EGFR Signaling Pathway WP437 | 3.59 × 10−5 | MAP3K2;MAPK7;GRB2;PTPN11;IQGAP1;CBL;CRK;SOS2;AP2M1;EGFR |
| Focal Adhesion WP306 | 4.92 × 10−5 | ITGB1;CCND1;PIK3CA;CTNNB1;GRB2;ARHGAP5;CRK;ACTB;RHOA;EGFR;VEGFA |
| RAC1/PAK1/p38/MMP2 Pathway WP3303 | 5.91 × 10−5 | ITGB1;PIK3CA;CTNNB1;PTPN11;GRB2;CRK;EGFR |
| VEGFA-VEGFR2 Signaling Pathway WP3888 | 7.15 × 10−5 | ITGB1;CCND1;PIK3CA;CTNNB1;GRB2;PTPN11;IQGAP1;CBL;CRK;RHOA;VEGFA |
| Regulation of Actin Cytoskeleton WP51 | 7.80 × 10−5 | PIK3CA;RDX;IQGAP1;CRK;SOS2;ACTB;RHOA;EGFR;SSH1 |
| Signaling of Hepatocyte Growth Factor Receptor WP313 | 1.82 × 10−4 | ITGB1;PIK3CA;PTPN11;GRB2;CRK |
| MET in type 1 papillary renal cell carcinoma WP4205 | 1.92 × 10−4 | PIK3CA;PTPN11;GRB2;CBL;SOS2;CRK |
| ErbB Signaling Pathway WP673 | 2.17 × 10−4 | CCND1;PIK3CA;GRB2;CBL;SOS2;CRK;EGFR |
| Endometrial cancer WP4155 | 2.20 × 10−4 | CCND1;PIK3CA;CTNNB1;GRB2;SOS2;EGFR |
| ESC Pluripotency Pathways WP3931 | 5.93 × 10−4 | MAPK7;LRP5;CTNNB1;GRB2;PTPN11;IL6ST;EGFR |
| KEGG | ||
| Proteoglycans in cancer | 2.75 × 10−8 | ITGB1;RDX;PTPN11;IQGAP1;CBL;ACTB;RHOA;EGFR;VEGFA;CCND1;PIK3CA;CTNNB1;GRB2;SOS2P |
| Focal adhesion | 1.65 × 10−6 | ITGB1;CCND1;PIK3CA;CTNNB1;GRB2;ARHGAP5;CRK;SOS2;ACTB;RHOA;EGFR;VEGFA |
| Human cytomegalovirus infection | 4.31 × 10−6 | AKAP13;CCND1;PIK3CA;GNA11;CREB3L2;CTNNB1;GRB2;CRK;SOS2;RHOA;EGFR;VEGFA |
| Chronic myeloid leukemia | 4.99 × 10−5 | CCND1;PIK3CA;PTPN11;GRB2;CBL;SOS2;CRK |
| Bacterial invasion of epithelial cells | 5.19 × 10−5 | ITGB1;PIK3CA;CTNNB1;CBL;CRK;ACTB;RHOA |
| Regulation of actin cytoskeleton | 8.63 × 10−5 | ITGB1;PIK3CA;RDX;IQGAP1;CRK;SOS2;ACTB;RHOA;EGFR;SSH1 |
| Colorectal cancer | 9.66 × 10−5 | CCND1;PIK3CA;CTNNB1;GRB2;SOS2;RHOA;EGFR |
| Endometrial cancer | 9.88 × 10−5 | CCND1;PIK3CA;CTNNB1;GRB2;SOS2;EGFR |
| Prostate cancer | 1.45 × 10−4 | CCND1;PIK3CA;CREB3L2;CTNNB1;GRB2;SOS2;EGFR |
| Renal cell carcinoma | 2.20 × 10−4 | PIK3CA;PTPN11;GRB2;SOS2;CRK;VEGFA |
| GO Cellular Component | ||
| Focal adhesion (GO:0005925) | 6.05 × 10−5 | NUP214;ITGB1;RDX;ADAM10;IQGAP1;LPP;ACTB;RHOA;EGFR;SENP1;MPRIP;ANXA6;CTNNB1;SNTB2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira-Rizzo, C.; Ottati, M.C.; Fort, R.S.; Chavez, S.; Trinidad, J.M.; DiPaolo, A.; Garat, B.; Sotelo-Silveira, J.R.; Duhagon, M.A. Hsa-miR-183-5p Modulates Cell Adhesion by Repression of ITGB1 Expression in Prostate Cancer. Non-Coding RNA 2022, 8, 11. https://doi.org/10.3390/ncrna8010011
Oliveira-Rizzo C, Ottati MC, Fort RS, Chavez S, Trinidad JM, DiPaolo A, Garat B, Sotelo-Silveira JR, Duhagon MA. Hsa-miR-183-5p Modulates Cell Adhesion by Repression of ITGB1 Expression in Prostate Cancer. Non-Coding RNA. 2022; 8(1):11. https://doi.org/10.3390/ncrna8010011
Chicago/Turabian StyleOliveira-Rizzo, Carolina, María Carolina Ottati, Rafael Sebastián Fort, Santiago Chavez, Juan Manuel Trinidad, Andrés DiPaolo, Beatriz Garat, José Roberto Sotelo-Silveira, and María Ana Duhagon. 2022. "Hsa-miR-183-5p Modulates Cell Adhesion by Repression of ITGB1 Expression in Prostate Cancer" Non-Coding RNA 8, no. 1: 11. https://doi.org/10.3390/ncrna8010011
APA StyleOliveira-Rizzo, C., Ottati, M. C., Fort, R. S., Chavez, S., Trinidad, J. M., DiPaolo, A., Garat, B., Sotelo-Silveira, J. R., & Duhagon, M. A. (2022). Hsa-miR-183-5p Modulates Cell Adhesion by Repression of ITGB1 Expression in Prostate Cancer. Non-Coding RNA, 8(1), 11. https://doi.org/10.3390/ncrna8010011