
Context-Dependent Regulation of Gene Expression by Non-Canonical Small RNAs


Abstract
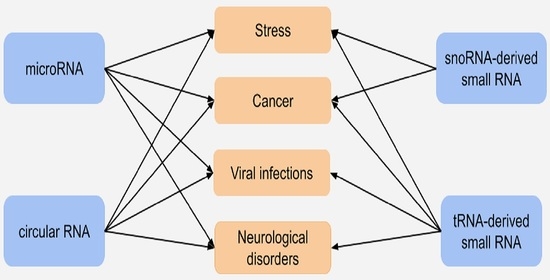
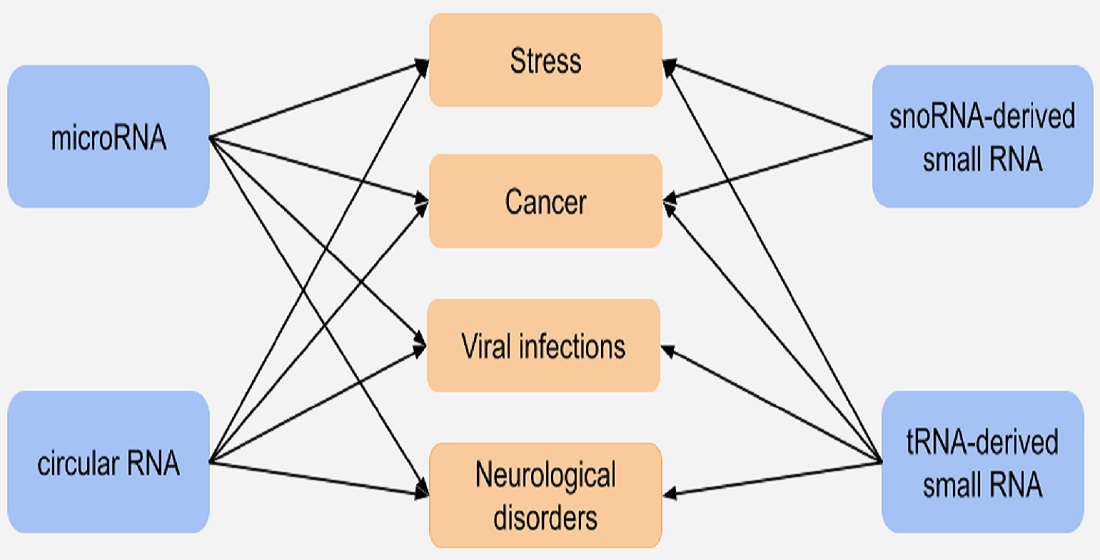
:
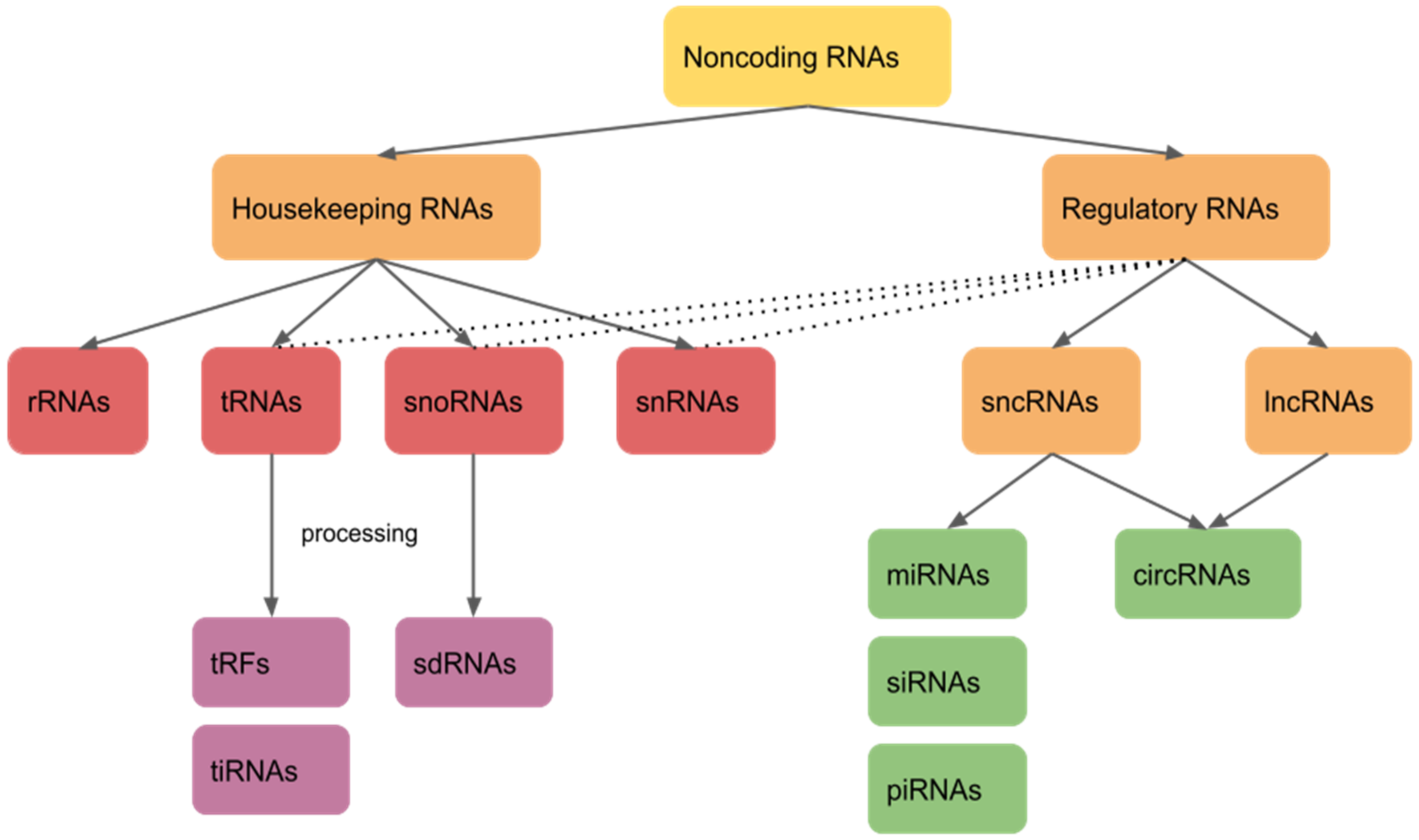
1. Introduction
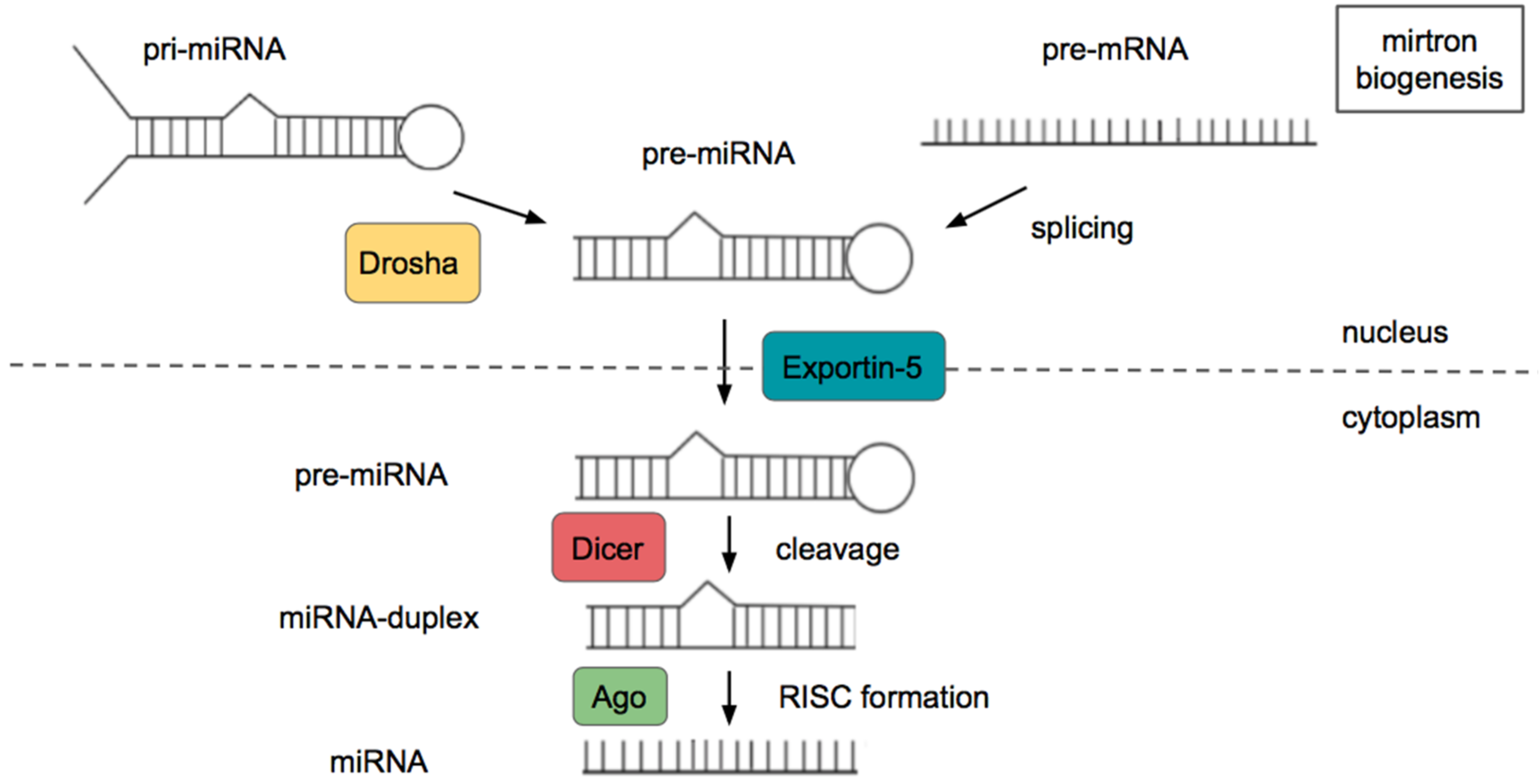
2. microRNAs
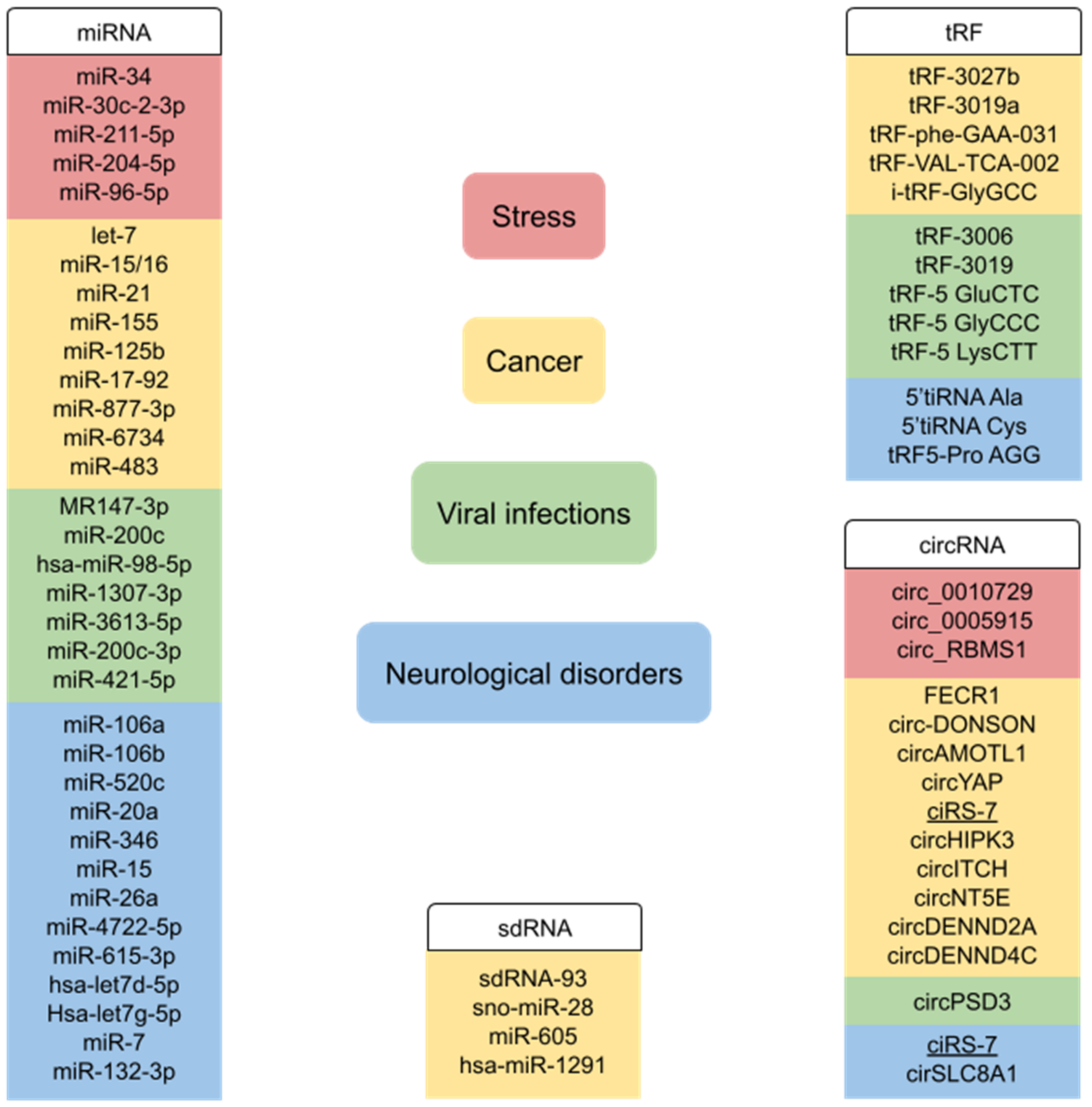
2.1. miRNAs in Stress Conditions
2.2. miRNAs in Cancer
2.3. miRNAs in Viral Infections
2.4. miRNAs in Neurological Disorders
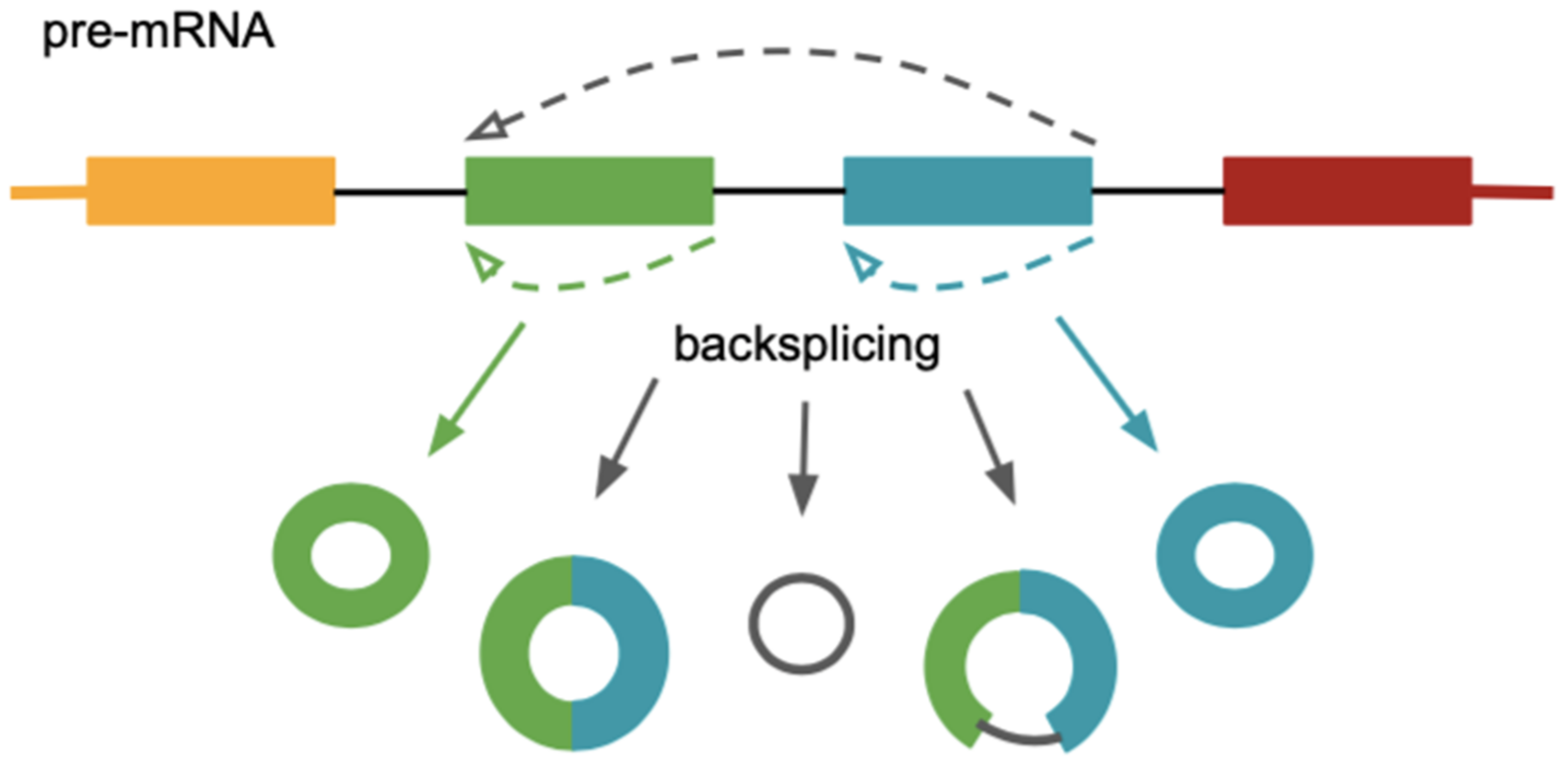
3. Circular RNAs
3.1. circRNAs in Stress Conditions
3.2. circRNAs in Cancer
3.3. circRNAs in Viral Infections
3.4. circRNAs in Neurological Disorders
4. snoRNA-Derived Small RNAs
4.1. sdRNAs in Stress Conditions
4.2. sdRNAs in Cancer
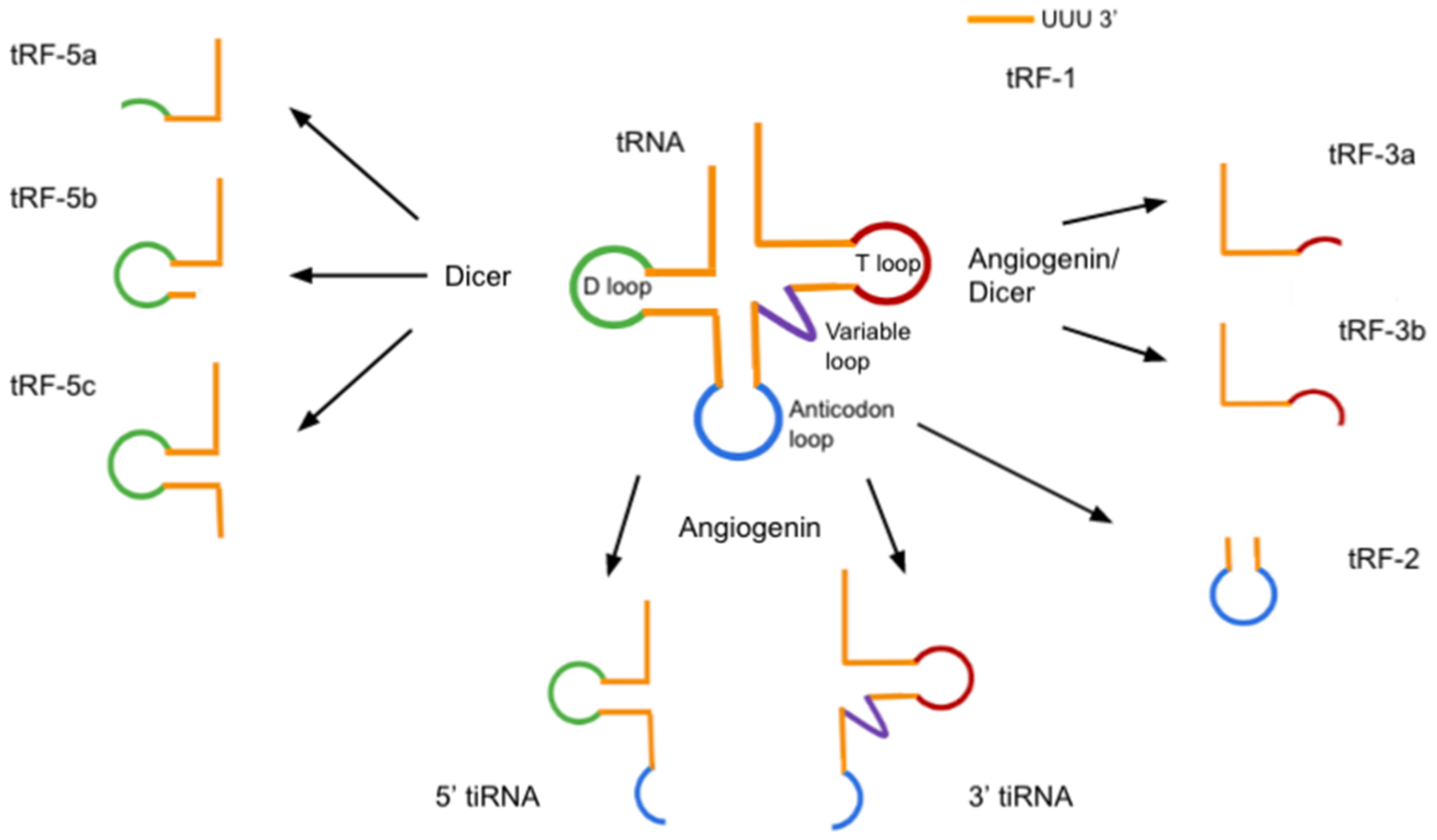
5. tRNA-Derived Small RNAs
5.1. tsRNAs in Stress Conditions
5.2. tsRNAs in Cancer
5.3. tsRNAs in Viral Infections
5.4. tsRNAs in Neurological Disorders
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ponting, C.P.; Belgard, T.G. Transcribed Dark Matter: Meaning or Myth? Hum. Mol. Genet. 2010, 19, R162–R168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulveling, D.; Francastel, C.; Hubé, F. When One Is Better than Two: RNA with Dual Functions. Biochimie 2011, 93, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Gonzàlez-Porta, M.; Frankish, A.; Rung, J.; Harrow, J.; Brazma, A. Transcriptome Analysis of Human Tissues and Cell Lines Reveals One Dominant Transcript per Gene. Genome Biol. 2013, 14, R70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beermann, J.; Piccoli, M.T.; Viereck, J.; Thum, T. Non-Coding Rnas in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [Green Version]
- Światowy, W.; Jagodzińśki, P.P. Molecules Derived from TRNA and SnoRNA: Entering the Degradome Pool. Biomed. Pharmacother. 2018, 108, 36–42. [Google Scholar] [CrossRef]
- Dai, X.; Kaushik, A.C.; Zhang, J. The Emerging Role of Major Regulatory RNAs in Cancer Control. Front. Oncol. 2019, 9, 920. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and Their Integrated Networks. J. Integr. Bioinform. 2019, 16, 20190027. [Google Scholar] [CrossRef]
- Romano, G.; Veneziano, D.; Acunzo, M.; Croce, C.M. Small Non-Coding RNA and Cancer. Carcinogenesis 2017, 38, 485–491. [Google Scholar] [CrossRef] [Green Version]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Rani, V.; Sengar, R.S. Biogenesis and Mechanisms of MicroRNA-Mediated Gene Regulation. Biotechnol. Bioeng. 2022, 119, 685–692. [Google Scholar] [CrossRef]
- Salim, U.; Kumar, A.; Kulshreshtha, R.; Vivekanandan, P. Biogenesis, Characterization, and Functions of Mirtrons. Wiley Interdiscip. Rev. RNA 2022, 13, e1680. [Google Scholar] [CrossRef]
- Kilikevicius, A.; Meister, G.; Corey, D.R. Reexamining Assumptions about MiRNA-Guided Gene Silencing. Nucleic Acids Res. 2022, 50, 617–634. [Google Scholar] [CrossRef]
- Wahid, F.; Shehzad, A.; Khan, T.; Kim, Y.Y. MicroRNAs: Synthesis, Mechanism, Function, and Recent Clinical Trials. Biochim. Biophys. Acta 2010, 1803, 1231–1243. [Google Scholar] [CrossRef] [Green Version]
- Naeli, P.; Winter, T.; Hackett, A.P.; Alboushi, L.; Jafarnejad, S.M. The Intricate Balance between MicroRNA-Induced MRNA Decay and Translational Repression. FEBS J. 2022. [Google Scholar] [CrossRef]
- Fukao, A.; Mishima, Y.; Takizawa, N.; Oka, S.; Imataka, H.; Pelletier, J.; Sonenberg, N.; Thoma, C.; Fujiwara, T. MicroRNAs Trigger Dissociation of EIF4AI and EIF4AII from Target MRNAs in Humans. Mol. Cell 2014, 56, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Vasudevan, S.; Steitz, J.A. AU-Rich-Element-Mediated Upregulation of Translation by FXR1 and Argonaute 2. Cell 2007, 128, 1105–1118. [Google Scholar] [CrossRef] [Green Version]
- Bukhari, S.I.A.; Truesdell, S.S.; Lee, S.; Kollu, S.; Classon, A.; Boukhali, M.; Jain, E.; Mortensen, R.D.; Yanagiya, A.; Sadreyev, R.I.; et al. A Specialized Mechanism of Translation Mediated by FXR1a-Associated MicroRNP in Cellular Quiescence. Mol. Cell 2016, 61, 760–773. [Google Scholar] [CrossRef] [Green Version]
- Ørom, U.A.; Nielsen, F.C.; Lund, A.H. MicroRNA-10a Binds the 5′UTR of Ribosomal Protein MRNAs and Enhances Their Translation. Mol. Cell 2008, 30, 460–471. [Google Scholar] [CrossRef]
- Liao, J.-Y.; Ma, L.-M.; Guo, Y.-H.; Zhang, Y.-C.; Zhou, H.; Shao, P.; Chen, Y.-Q.; Qu, L.-H. Deep Sequencing of Human Nuclear and Cytoplasmic Small RNAs Reveals an Unexpectedly Complex Subcellular Distribution of MiRNAs and TRNA 3′ Trailers. PLoS ONE 2010, 5, e10563. [Google Scholar] [CrossRef] [Green Version]
- Roberts, T.C. The MicroRNA Biology of the Mammalian Nucleus. Mol. Ther.-Nucleic Acids 2014, 3, e188. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lei, C.; He, Q.; Pan, Z.; Xiao, D.; Tao, Y. Nuclear Functions of Mammalian MicroRNAs in Gene Regulation, Immunity and Cancer. Mol. Cancer. 2018, 17, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benhamed, M.; Herbig, U.; Ye, T.; Dejean, A.; Bischof, O. Senescence Is an Endogenous Trigger for MicroRNA-Directed Transcriptional Gene Silencing in Human Cells. Nat. Cell Biol. 2012, 14, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Yao, H.; Li, C.; Pu, M.; Yao, X.; Yang, H.; Qi, X.; Ren, J.; Wang, Y. A Dual Inhibition: MicroRNA-552 Suppresses Both Transcription and Translation of Cytochrome P450 2E1. Biochim. Biophys. Acta 2016, 1859, 650–662. [Google Scholar] [CrossRef]
- Zardo, G.; Ciolfi, A.; Vian, L.; Starnes, L.M.; Billi, M.; Racanicchi, S.; Maresca, C.; Fazi, F.; Travaglini, L.; Noguera, N.; et al. Polycombs and MicroRNA-223 Regulate Human Granulopoiesis by Transcriptional Control of Target Gene Expression. Blood 2012, 119, 4034–4046. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Fan, M.; Zhang, X.; Huang, F.; Wu, K.; Zhang, J.; Liu, J.; Huang, Z.; Luo, H.; Tao, L.; et al. Cellular MicroRNAs Up-Regulate Transcription via Interaction with Promoter TATA-Box Motifs. RNA 2014, 20, 1878–1889. [Google Scholar] [CrossRef] [Green Version]
- Place, R.F.; Li, L.-C.; Pookot, D.; Noonan, E.J.; Dahiya, R. MicroRNA-373 Induces Expression of Genes with Complementary Promoter Sequences. Proc. Natl. Acad. Sci. USA 2008, 105, 1608. [Google Scholar] [CrossRef] [Green Version]
- Xiao, M.; Li, J.; Li, W.; Wang, Y.; Wu, F.; Xi, Y.; Zhang, L.; Ding, C.; Luo, H.; Li, Y.; et al. MicroRNAs Activate Gene Transcription Epigenetically as an Enhancer Trigger. RNA Biol. 2017, 14, 1326–1334. [Google Scholar] [CrossRef]
- Boudreau, R.L.; Jiang, P.; Gilmore, B.L.; Spengler, R.M.; Tirabassi, R.; Nelson, J.A.; Ross, C.A.; Xing, Y.; Davidson, B.L. Transcriptome-Wide Discovery of MicroRNA Binding Sites in Human Brain. Neuron 2014, 81, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Spengler, R.M.; Zhang, X.; Cheng, C.; McLendon, J.M.; Skeie, J.M.; Johnson, F.L.; Davidson, B.L.; Boudreau, R.L. Elucidation of Transcriptome-Wide MicroRNA Binding Sites in Human Cardiac Tissues by Ago2 HITS-CLIP. Nucleic Acids Res. 2016, 44, 7120–7131. [Google Scholar] [CrossRef]
- Sadakierska-Chudy, A. Micrornas: Diverse Mechanisms of Action and Their Potential Applications as Cancer Epi-Therapeutics. Biomolecules 2020, 10, 1285. [Google Scholar] [CrossRef]
- Byrd, A.E.; Brewer, J.W. Micro(RNA)Managing Endoplasmic Reticulum Stress. IUBMB Life 2013, 65, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Olejniczak, M.; Kotowska-Zimmer, A.; Krzyzosiak, W. Stress-Induced Changes in MiRNA Biogenesis and Functioning. Cell. Mol. Life Sci. 2018, 75, 177–191. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liao, Y.; Tang, L. MicroRNA-34 Family: A Potential Tumor Suppressor and Therapeutic Candidate in Cancer. J. Exp. Clin. Cancer Res. 2019, 38, 53. [Google Scholar] [CrossRef] [Green Version]
- Le, M.T.N.; Teh, C.; Shyh-Chang, N.; Xie, H.; Zhou, B.; Korzh, V.; Lodish, H.F.; Lim, B. MicroRNA-125b Is a Novel Negative Regulator of P53. Genes Dev. 2009, 23, 862–876. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.I.; Yamagata, K.; Sugimoto, K.; Iwamoto, T.; Kato, S.; Miyazono, K. Modulation of MicroRNA Processing by P53. Nature 2009, 460, 529–533. [Google Scholar] [CrossRef]
- Navarro, F.; Lieberman, J. MiR-34 and P53: New Insights into a Complex Functional Relationship. PLoS ONE 2015, 10, e0132767. [Google Scholar] [CrossRef]
- Tabas, I.; Ron, D. Integrating the Mechanisms of Apoptosis Induced by Endoplasmic Reticulum Stress. Nat. Cell Biol. 2011, 13, 184–190. [Google Scholar] [CrossRef]
- Byrd, A.E.; Aragon, I.V.; Brewer, J.W. MicroRNA-30c-2* Limits Expression of Proadaptive Factor XBP1 in the Unfolded Protein Response. J. Cell Biol. 2012, 196, 689. [Google Scholar] [CrossRef] [Green Version]
- Grieco, F.A.; Schiavo, A.A.; Brozzi, F.; Juan-Mateu, J.; Bugliani, M.; Marchetti, P.; Eizirik, D.L. The MicroRNAs MiR-211-5p and MiR-204-5p Modulate ER Stress in Human Beta Cells. J. Mol. Endocrinol. 2019, 63, 139. [Google Scholar] [CrossRef]
- Li, G.; Luna, C.; Qiu, J.; Epstein, D.L.; Gonzalez, P. Role of MiR-204 in the Regulation of Apoptosis, Endoplasmic Reticulum Stress Response, and Inflammation in Human Trabecular Meshwork Cells. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2999. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Yang, Y.; Shi, M.H.; Wang, J.F.; Ran, X.Q. MiR-96-5p Attenuates Malathion-Induced Apoptosis of Human Kidney Cells by Targeting the ER Stress Marker DDIT3. J. Environ. Sci. Health Part B 2020, 55, 1080–1086. [Google Scholar] [CrossRef]
- Si, L.; Yang, Z.; Ding, L.; Zhang, D. Regulatory Effects of LncRNAs and MiRNAs on the Crosstalk between Autophagy and EMT in Cancer: A New Era for Cancer Treatment. J. Cancer Res. Clin. Oncol. 2022, 148, 547–564. [Google Scholar] [CrossRef]
- Dragomir, M.P.; Knutsen, E.; Calin, G.A. Classical and Noncanonical Functions of MiRNAs in Cancers. Trends Genet. 2022, 38, 379–394. [Google Scholar] [CrossRef]
- Bernstein, D.L.; Jiang, X.; Rom, S. Let-7 MicroRNAs: Their Role in Cerebral and Cardiovascular Diseases, Inflammation, Cancer, and Their Regulation. Biomedicines 2021, 9, 606. [Google Scholar] [CrossRef]
- Pekarsky, Y.; Balatti, V.; Croce, C.M. BCL2 and MiR-15/16: From Gene Discovery to Treatment. Cell Death Differ. 2017, 25, 21–26. [Google Scholar] [CrossRef]
- Dai, L.; Chen, F.; Zheng, Y.; Zhang, D.; Qian, B.; Ji, H.; Long, F.; Cretoiu, D. MiR-21 Regulates Growth and EMT in Lung Cancer Cells via PTEN/Akt/GSK3β Signaling. Front. Biosci. 2019, 24, 1426–1439. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Tan, Z.; Hu, H.; Liu, H.; Wu, T.; Zheng, C.; Wang, X.; Luo, Z.; Wang, J.; Liu, S.; et al. MicroRNA-21 Promotes Breast Cancer Proliferation and Metastasis by Targeting LZTFL1. BMC Cancer 2019, 19, 738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, F.; Yin, H.B.; Li, X.Y.; Zhu, G.M.; He, W.Y.; Gou, X. Bladder Cancer Cell-secreted Exosomal MiR-21 Activates the PI3K/AKT Pathway in Macrophages to Promote Cancer Progression. Int. J. Oncol. 2020, 56, 151–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wu, J. Role of MiR-155 in Breast Cancer. Front. Biosci. 2012, 17, 2350–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattiske, S.; Suetani, R.J.; Neilsen, P.M.; Callen, D.F. The Oncogenic Role of MiR-155 in Breast Cancer. Cancer Epidemiol. Biomark. Prev. 2012, 21, 1236–1243. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wei, Y.; Fan, X.; Zhang, P.; Wang, P.; Cheng, S.; Zhang, J. MicroRNA-125b as a Tumor Suppressor by Targeting MMP11 in Breast Cancer. Thorac. Cancer 2020, 11, 1613. [Google Scholar] [CrossRef] [Green Version]
- Peng, B.; Theng, P.Y.; Le, M.T.N. Essential Functions of MiR-125b in Cancer. Cell Prolif. 2021, 54, e12913. [Google Scholar] [CrossRef]
- Tang, L.; Yuan, Y.; Zhai, H.; Wang, J.; Zhang, D.; Liang, H.; Shi, Y.; Duan, L.; Jiang, X. MicroRNA-125b-5p Correlates with Prognosis and Lung Adenocarcinoma Progression. Front. Mol. Biosci. 2021, 8, 788690. [Google Scholar] [CrossRef]
- MacFarlane, L.-A.; Paul, R.M. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [Green Version]
- Gruszka, R.; Zakrzewski, K.; Liberski, P.P.; Zakrzewska, M. MRNA and MiRNA Expression Analyses of the MYC/E2F/MiR-17-92 Network in the Most Common Pediatric Brain Tumors. Int. J. Mol. Sci. 2021, 22, 543. [Google Scholar] [CrossRef]
- Huang, Q.; Shen, Y.J.; Hsueh, C.Y.; Guo, Y.; Zhang, Y.F.; Li, J.Y.; Zhou, L. MiR-17-5p Drives G2/M-Phase Accumulation by Directly Targeting CCNG2 and Is Related to Recurrence of Head and Neck Squamous Cell Carcinoma. BMC Cancer 2021, 21, 1074. [Google Scholar] [CrossRef]
- Li, J.; Lai, Y.; Ma, J.; Liu, Y.; Bi, J.; Zhang, L.; Chen, L.; Yao, C.; Lv, W.; Chang, G.; et al. MiR-17-5p Suppresses Cell Proliferation and Invasion by Targeting ETV1 in Triple-Negative Breast Cancer. BMC Cancer 2017, 17, 745. [Google Scholar] [CrossRef]
- Liu, D.L.; Lu, L.L.; Dong, L.L.; Liu, Y.; Bian, X.Y.; Lian, B.F.; Xie, L.; Wen, D.; Gao, D.M.; Ke, A.W.; et al. MiR-17-5p and MiR-20a-5p Suppress Postoperative Metastasis of Hepatocellular Carcinoma via Blocking HGF/ERBB3-NF-ΚB Positive Feedback Loop. Theranostics 2020, 10, 3668–3683. [Google Scholar] [CrossRef]
- Shell, S.; Park, S.-M.; Radjabi, A.R.; Schickel, R.; Kistner, E.O.; Jewell, D.A.; Feig, C.; Lengyel, E.; Peter, M.E. Let-7 Expression Defines Two Differentiation Stages of Cancer. Proc. Natl. Acad. Sci. USA 2007, 104, 11400–11405. [Google Scholar] [CrossRef] [Green Version]
- Lovat, F.; Fassan, M.; Sacchi, D.; Ranganathan, P.; Palamarchuk, A.; Bill, M.; Karunasiri, M.; Gasparini, P.; Nigita, G.; Distefano, R.; et al. Knockout of Both MiR-15/16 Loci Induces Acute Myeloid Leukemia. Proc. Natl. Acad. Sci. USA 2018, 115, 13069–13074. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Zhang, H.-W.; Lu, M.-H.; He, X.-H.; Li, Y.; Gu, H.; Liu, M.-F.; Wang, E.-D. MicroRNA-155 Functions as an OncomiR in Breast Cancer by Targeting the Suppressor of Cytokine Signaling 1 Gene. Cancer Res. 2010, 70, 3119–3127. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhu, Y.; Liang, Z.; Wang, X.; Meng, S.; Xu, X.; Xu, X.; Wu, J.; Ji, A.; Hu, Z.; et al. Up-Regulation of P16 by MiR-877-3p Inhibits Proliferation of Bladder Cancer. Oncotarget 2016, 7, 51773. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.R.; Park, K.H.; Yang, J.-O.; Lee, C.W.; Oh, S.J.; Yun, J.; Lee, M.Y.; Han, S.-B.; Kang, J.S. MiR-6734 Up-Regulates P21 Gene Expression and Induces Cell Cycle Arrest and Apoptosis in Colon Cancer Cells. PLoS ONE 2016, 11, e0160961. [Google Scholar] [CrossRef]
- Tang, S.; Chen, Y.; Feng, S.; Yi, T.; Liu, X.; Li, Q.; Liu, Z.; Zhu, C.; Hu, J.; Yu, X.; et al. MiR-483-5p Promotes IGF-II Transcription and Is Associated with Poor Prognosis of Hepatocellular Carcinoma. Oncotarget 2017, 8, 99871–99888. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Fernández-Pato, A.; Virseda-Berdices, A.; Resino, S.; Ryan, P.; Martínez-González, O.; Pérez-García, F.; Martin-Vicente, M.; Valle-Millares, D.; Brochado-Kith, O.; Blancas, R.; et al. Plasma MiRNA Profile at COVID-19 Onset Predicts Severity Status and Mortality. Emerg. Microbes Infect. 2022, 11, 676–688. [Google Scholar] [CrossRef]
- Li, C.; Hu, X.; Li, L.; Li, J. Differential MicroRNA Expression in the Peripheral Blood from Human Patients with COVID-19. J. Clin. Lab. Anal. 2020, 34, e23590. [Google Scholar] [CrossRef]
- Khan, M.A.A.K.; Sany, M.R.U.; Islam, M.S.; Islam, A.B.M.M.K. Epigenetic Regulator MiRNA Pattern Differences Among SARS-CoV, SARS-CoV-2, and SARS-CoV-2 World-Wide Isolates Delineated the Mystery Behind the Epic Pathogenicity and Distinct Clinical Characteristics of Pandemic COVID-19. Front. Genet. 2020, 11, 765. [Google Scholar] [CrossRef]
- Zhang, S.; Amahong, K.; Sun, X.; Lian, X.; Liu, J.; Sun, H.; Lou, Y.; Zhu, F.; Qiu, Y. The MiRNA: A Small but Powerful RNA for COVID-19. Brief Bioinform. 2021, 22, 1137–1149. [Google Scholar] [CrossRef]
- Satyam, R.; Bhardwaj, T.; Goel, S.; Jha, N.K.; Jha, S.K.; Nand, P.; Ruokolainen, J.; Kamal, M.A.; Kesari, K.K. MiRNAs in SARS-CoV 2: A Spoke in the Wheel of Pathogenesis. Curr. Pharm. Des. 2020, 27, 1628–1641. [Google Scholar] [CrossRef] [PubMed]
- Turnquist, C.; Ryan, B.M.; Horikawa, I.; Harris, B.T.; Harris, C.C. Cytokine Storms in Cancer and COVID-19. Cancer Cell 2020, 38, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Demongeot, J.; Seligmann, H. SARS-CoV-2 and MiRNA-like Inhibition Power. Med. Hypotheses 2020, 144, 110245. [Google Scholar] [CrossRef] [PubMed]
- Abedi, F.; Rezaee, R.; Hayes, A.W.; Nasiripour, S.; Karimi, G. MicroRNAs and SARS-CoV-2 Life Cycle, Pathogenesis, and Mutations: Biomarkers or Therapeutic Agents? Cell Cycle 2021, 20, 143–153. [Google Scholar] [CrossRef]
- Lu, D.; Chatterjee, S.; Xiao, K.; Riedel, I.; Wang, Y.; Foo, R.; Bär, C.; Thum, T. MicroRNAs Targeting the SARS-CoV-2 Entry Receptor ACE2 in Cardiomyocytes. J. Mol. Cell. Cardiol. 2020, 148, 46–49. [Google Scholar] [CrossRef]
- Matarese, A.; Gambardella, J.; Sardu, C.; Santulli, G. MiR-98 Regulates TMPRSS2 Expression in Human Endothelial Cells: Key Implications for COVID-19. Biomedicines 2020, 8, 462. [Google Scholar] [CrossRef]
- Rizkita, L.D.; Astuti, I. The Potential of MiRNA-Based Therapeutics in Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection: A Review. J. Pharm. Anal. 2021, 11, 265–271. [Google Scholar] [CrossRef]
- Chen, L.; Zhong, L. Genomics Functional Analysis and Drug Screening of SARS-CoV-2. Genes Dis. 2020, 7, 542–550. [Google Scholar] [CrossRef]
- Bruscella, P.; Bottini, S.; Baudesson, C.; Pawlotsky, J.-M.; Feray, C.; Trabucchi, M. Viruses and MiRNAs: More Friends than Foes. Front. Microbiol. 2017, 8, 824. [Google Scholar] [CrossRef] [Green Version]
- Saksena, N.; Bonam, S.R.; Miranda-Saksena, M. Epigenetic Lens to Visualize the Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2) Infection in COVID-19 Pandemic. Front. Genet. 2021, 12, 581726. [Google Scholar] [CrossRef]
- Abdolahi, S.; Hosseini, M.; Rezaei, R.; Mohebbi, S.R.; Rostami-Nejad, M.; Mojarad, E.N.; Mirjalali, H.; Yadegar, A.; Asadzadeh Aghdaei, H.; Zali, M.R.; et al. Evaluation of MiR-200c-3p and MiR-421-5p Levels during Immune Responses in the Admitted and Recovered COVID-19 Subjects. Infect. Genet. Evol. 2022, 98, 105207. [Google Scholar] [CrossRef]
- Khezri, M.R.; Yousefi, K.; Zolbanin, N.M.; Ghasemnejad-Berenji, M. MicroRNAs in the Pathophysiology of Alzheimer’s Disease and Parkinson’s Disease: An Overview. Mol. Neurobiol. 2022, 59, 1589–1603. [Google Scholar] [CrossRef]
- Rybak-Wolf, A.; Plass, M. RNA Dynamics in Alzheimer’s Disease. Molecules 2021, 26, 5113. [Google Scholar] [CrossRef]
- Patel, N.; Hoang, D.; Miller, N.; Ansaloni, S.; Huang, Q.; Rogers, J.T.; Lee, J.C.; Saunders, A.J. MicroRNAs Can Regulate Human APP Levels. Mol. Neurodegener. 2008, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Hébert, S.S.; Horré, K.; Nicolaï, L.; Bergmans, B.; Papadopoulou, A.S.; Delacourte, A.; de Strooper, B. MicroRNA Regulation of Alzheimer’s Amyloid Precursor Protein Expression. Neurobiol. Dis. 2009, 33, 422–428. [Google Scholar] [CrossRef]
- Long, J.M.; Maloney, B.; Rogers, J.T.; Lahiri, D.K. Novel Upregulation of Amyloid-β Precursor Protein (APP) by MicroRNA-346 via Targeting of APP MRNA 5′-Untranslated Region: Implications in Alzheimer’s Disease. Mol. Psychiatry 2018, 24, 345–363. [Google Scholar] [CrossRef] [Green Version]
- Maqbool, M.; Mobashir, M.; Hoda, N. Pivotal Role of Glycogen Synthase Kinase-3: A Therapeutic Target for Alzheimer’s Disease. Eur. J. Med. Chem. 2016, 107, 63–81. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, Y.; Yu, M. MicroRNA-4722-5p and MicroRNA-615-3p Serve as Potential Biomarkers for Alzheimer’s Disease. Exp. Ther. Med. 2022, 23, 241. [Google Scholar] [CrossRef]
- Poursaei, E.; Abolghasemi, M.; Bornehdeli, S.; Shanehbandi, D.; Asadi, M.; Sadeghzadeh, M.; Rahmanpour, D.; Sadeh, R.N. Evaluation of Hsa-Let-7d-5p, Hsa-Let-7g-5p and Hsa-MiR-15b-5p Plasma Levels in Patients with Alzheimer’s Disease. Psychiatry Genet. 2022, 32, 25–29. [Google Scholar] [CrossRef]
- García-Fonseca, Á.; Martin-Jimenez, C.; Barreto, G.E.; Pachón, A.F.A.; González, J. The Emerging Role of Long Non-Coding RNAs and MicroRNAs in Neurodegenerative Diseases: A Perspective of Machine Learning. Biomolecules 2021, 11, 1132. [Google Scholar] [CrossRef]
- Choi, D.C.; Yoo, M.; Kabaria, S.; Junn, E. MicroRNA-7 Facilitates the Degradation of Alpha-Synuclein and Its Aggregates by Promoting Autophagy. Neurosci. Lett. 2018, 678, 118. [Google Scholar] [CrossRef]
- Li, S.; Bi, G.; Han, S.; Huang, R. MicroRNAs Play a Role in Parkinson’s Disease by Regulating Microglia Function: From Pathogenetic Involvement to Therapeutic Potential. Front. Mol. Neurosci. 2022, 14, 358. [Google Scholar] [CrossRef]
- Gong, X.; Huang, M.; Chen, L. Mechanism of MiR-132-3p Promoting Neuroinflammation and Dopaminergic Neurodegeneration in Parkinson’s Disease. eNeuro 2022, 9, 1–17. [Google Scholar] [CrossRef]
- Tang, X.; Ren, H.; Guo, M.; Qian, J.; Yang, Y.; Gu, C. Review on Circular RNAs and New Insights into Their Roles in Cancer. Comput. Struct. Biotechnol. J. 2021, 19, 910–928. [Google Scholar] [CrossRef]
- Nisar, S.; Bhat, A.A.; Singh, M.; Karedath, T.; Rizwan, A.; Hashem, S.; Bagga, P.; Reddy, R.; Jamal, F.; Uddin, S.; et al. Insights into the Role of CircRNAs: Biogenesis, Characterization, Functional, and Clinical Impact in Human Malignancies. Front. Cell Dev. Biol. 2021, 9, 617281. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z. Efficient Backsplicing Produces Translatable Circular MRNAs. RNA 2015, 21, 172–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, M.; Zheng, M.; Xu, Y.; Ma, S.; Zhang, W.; Ju, S. CircRNAs and Their Regulatory Roles in Cancers. Mol. Med. 2021, 27, 94. [Google Scholar] [CrossRef]
- Patop, I.L.; Wüst, S.; Kadener, S. Past, Present, and Future of Circ RNA s. EMBO J. 2019, 38, e100836. [Google Scholar] [CrossRef]
- Huang, A.; Zheng, H.; Wu, Z.; Chen, M.; Huang, Y. Circular RNA-Protein Interactions: Functions, Mechanisms, and Identification. Theranostics 2020, 10, 3506–3517. [Google Scholar] [CrossRef]
- Misir, S.; Wu, N.; Yang, B.B. Specific Expression and Functions of Circular RNAs. Cell Death Differ. 2022, 29, 481–491. [Google Scholar] [CrossRef]
- Wang, M.; Yu, F.; Li, P. Circular RNAs: Characteristics, Function and Clinical Significance in Hepatocellular Carcinoma. Cancers 2018, 10, 258. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-Intron Circular RNAs Regulate Transcription in the Nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular Intronic Long Noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Lu, T.; Wang, Q.; Liu, J.; Jiao, W. Circular RNAs: Crucial Regulators in the Human Body (Review). Oncol. Rep. 2018, 40, 3119–3135. [Google Scholar] [CrossRef]
- Geng, X.; Jia, Y.; Zhang, Y.; Shi, L.; Li, Q.; Zang, A.; Wang, H. Circular RNA: Biogenesis, Degradation, Functions and Potential Roles in Mediating Resistance to Anticarcinogens. Epigenomics 2020, 12, 267–283. [Google Scholar] [CrossRef]
- Lei, D.; Wang, Y.; Zhang, L.; Wang, Z. Circ_0010729 Regulates Hypoxia-Induced Cardiomyocyte Injuries by Activating TRAF5 via Sponging MiR-27a-3p. Life Sci. 2020, 262, 118511. [Google Scholar] [CrossRef]
- Dang, R.Y.; Liu, F.L.; Li, Y. Circular RNA Hsa_circ_0010729 Regulates Vascular Endothelial Cell Proliferation and Apoptosis by Targeting the MiR-186/HIF-1α Axis. Biochem. Biophys. Res. Commun. 2017, 490, 104–110. [Google Scholar] [CrossRef]
- Liu, Z.; He, Q.; Liu, Y.; Zhang, Y.; Cui, M.; Peng, H.; Wang, Y.; Chen, S.; Li, D.; Chen, L.; et al. Hsa_circ_0005915 Promotes N,N-Dimethylformamide-Induced Oxidative Stress in HL-7702 Cells through NRF2/ARE Axis. Toxicology 2021, 458, 152838. [Google Scholar] [CrossRef]
- Vikhe, P.P.; Tateossian, H.; Bharj, G.; Brown, S.D.M.; Hood, D.W. Mutation in Fbxo11 Leads to Altered Immune Cell Content in Jeff Mouse Model of Otitis Media. Front. Genet. 2020, 11, 50. [Google Scholar] [CrossRef]
- Qiao, D.; Hu, C.; Li, Q.; Fan, J. Circ-Rbms1 Knockdown Alleviates CSE-Induced Apoptosis, Inflammation and Oxidative Stress via up-Regulating FBXO11 through MiR-197-3p in 16HBE Cells. Int. J. COPD 2021, 16, 2105–2118. [Google Scholar] [CrossRef]
- Chen, N.; Zhao, G.; Yan, X.; Lv, Z.; Yin, H.; Zhang, S.; Song, W.; Li, X.; Li, L.; Du, Z.; et al. A Novel FLI1 Exonic Circular RNA Promotes Metastasis in Breast Cancer by Coordinately Regulating TET1 and DNMT1. Genome Biol. 2018, 19, 218. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Zhao, Y.; Dang, S.; Wang, Y.; Li, X.; Yu, X.; Li, Z.; Wei, J.; Liu, M.; Li, G. Circular RNA Circ-DONSON Facilitates Gastric Cancer Growth and Invasion via NURF Complex Dependent Activation of Transcription Factor SOX4. Mol. Cancer 2019, 18, 45. [Google Scholar] [CrossRef]
- Yang, Q.; Du, W.W.; Wu, N.; Yang, W.; Awan, F.M.; Fang, L.; Ma, J.; Li, X.; Zeng, Y.; Yang, Z.; et al. A Circular RNA Promotes Tumorigenesis by Inducing C-Myc Nuclear Translocation. Cell Death Differ. 2017, 24, 1609–1620. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Yuan, Z.; Du, K.Y.; Fang, L.; Lyu, J.; Zhang, C.; He, A.; Eshaghi, E.; Zeng, K.; Ma, J.; et al. Translation of Yes-Associated Protein (YAP) Was Antagonized by Its Circular RNA via Suppressing the Assembly of the Translation Initiation Machinery. Cell Death Differ. 2019, 26, 2758–2773. [Google Scholar] [CrossRef] [Green Version]
- Rahmati, Y.; Asemani, Y.; Aghamiri, S.; Ezzatifar, F.; Najafi, S. CiRS-7/CDR1as; An Oncogenic Circular RNA as a Potential Cancer Biomarker. Pathol. Res. Pract. 2021, 227, 153639. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, D.; Wei, Y. Overexpressed CDR1as Functions as an Oncogene to Promote the Tumor Progression via MiR-7 in Non-Small-Cell Lung Cancer. OncoTargets Ther. 2018, 11, 3979–3987. [Google Scholar] [CrossRef] [Green Version]
- Weng, W.; Wei, Q.; Toden, S.; Yoshida, K.; Nagasaka, T.; Fujiwara, T.; Cai, S.; Qin, H.; Ma, Y.; Goel, A. Circular RNA CiRS-7—A Promising Prognostic Biomarker and a Potential Therapeutic Target in Colorectal Cancer. Clin. Cancer Res. 2017, 23, 3918–3928. [Google Scholar] [CrossRef] [Green Version]
- Zeng, K.; Chen, X.; Xu, M.; Liu, X.; Hu, X.; Xu, T.; Sun, H.; Pan, Y.; He, B.; Wang, S. CircHIPK3 Promotes Colorectal Cancer Growth and Metastasis by Sponging MiR-7. Cell Death Dis. 2018, 9, 417. [Google Scholar] [CrossRef]
- Kai, D.; Yannian, L.; Yitian, C.; Dinghao, G.; Xin, Z.; Wu, J. Circular RNA HIPK3 Promotes Gallbladder Cancer Cell Growth by Sponging MicroRNA-124. Biochem. Biophys. Res. Commun. 2018, 503, 863–869. [Google Scholar] [CrossRef]
- Yu, Q.; Chen, W.; Li, Y.; He, J.; Wang, Y.; Yang, S.; Zhou, J. The Novel Circular RNA HIPK3 Accelerates the Proliferation and Invasion of Hepatocellular Carcinoma Cells by Sponging the Micro RNA-124 or Micro RNA-506/Pyruvate Dehydrogenase Kinase 2 Axis. Bioengineered 2022, 13, 4717–4729. [Google Scholar] [CrossRef]
- Huang, G.; Zhu, H.; Shi, Y.; Wu, W.; Cai, H.; Chen, X. Cir-ITCH Plays an Inhibitory Role in Colorectal Cancer by Regulating the Wnt/β-Catenin Pathway. PLoS ONE 2015, 10, e0131225. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Sang, J.; Zhang, Y.; Gao, L.; Zhao, D.; Cao, H. Circular RNA ITCH Attenuates the Progression of Nasopharyngeal Carcinoma by Inducing PTEN Upregulation via MiR-214. J. Gene Med. 2022, 24. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, S.; Chen, X.; Li, N.; Li, J.; Jia, R.; Pan, Y.; Liang, H. CircNT5E Acts as a Sponge of MiR-422a to Promote Glioblastoma Tumorigenesis. Cancer Res. 2018, 78, 4812–4825. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Zheng, J.; Gao, Y.; Zhou, X.; Song, W.; Huang, J. The Circular RNA NT5E Promotes Non-Small Cell Lung Cancer Cell Growth via Sponging MicroRNA-134. Aging 2020, 12, 3936. [Google Scholar] [CrossRef]
- Yang, J.; Liu, X.; Dai, G.; Qu, L.; Tan, B.; Zhu, B.; Qi, F.; Gai, X.; Cheng, B. CircNT5E Promotes the Proliferation and Migration of Bladder Cancer via Sponging MiR-502-5p. J. Cancer 2021, 12, 2430. [Google Scholar] [CrossRef]
- Su, H.; Zou, D.; Sun, Y.; Dai, Y. Hypoxia-Associated CircDENND2A Promotes Glioma Aggressiveness by Sponging MiR-625-5p. Cell. Mol. Biol. Lett. 2019, 24, 24. [Google Scholar] [CrossRef]
- Ren, S.; Liu, J.; Feng, Y.; Li, Z.; He, L.; Li, L.; Cao, X.; Wang, Z.; Zhang, Y. Knockdown of CircDENND4C Inhibits Glycolysis, Migration and Invasion by up-Regulating MiR-200b/c in Breast Cancer under Hypoxia. J. Exp. Clin. Cancer Res. 2019, 38, 388. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Qin, Y.; Li, S.; Li, L.; He, J.; Chen, Y.; Zhou, X.; Liu, H. CircDENND4C Promotes Proliferation and Metastasis of Lung Cancer by Upregulating BRD4 Signaling Pathway. J. Oncol. 2021, 2021, 2469691. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Khoshbakht, T.; Taheri, M.; Jamali, E. CircITCH: A Circular RNA With Eminent Roles in the Carcinogenesis. Front. Oncol. 2021, 11, 4304. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, W.; Zhou, Q.; Chen, C.; Yuan, W.; Liu, J.; Li, X.; Sun, Z. Roles of CircRNAs in the Tumour Microenvironment. Mol. Cancer 2020, 19, 14. [Google Scholar] [CrossRef] [Green Version]
- Lou, Y.Y.; Wang, Q.D.; Lu, Y.T.; Tu, M.Y.; Xu, X.; Xia, Y.; Peng, Y.; Lai, M.M.; Zheng, X.Q. Differential CircRNA Expression Profiles in Latent Human Cytomegalovirus Infection and Validation Using Clinical Samples. Physiol. Genom. 2019, 51, 51–58. [Google Scholar] [CrossRef]
- Chen, T.-C.; Tallo-Parra, M.; Cao, Q.M.; Kadener, S.; Böttcher, R.; Pérez-Vilaró, G.; Boonchuen, P.; Somboonwiwat, K.; Díez, J.; Sarnow, P. Host-Derived Circular RNAs Display Proviral Activities in Hepatitis C Virus-Infected Cells. PLOS Pathog. 2020, 16, e1008346. [Google Scholar] [CrossRef]
- Cai, Z.; Lu, C.; He, J.; Liu, L.; Zou, Y.; Zhang, Z.; Zhu, Z.; Ge, X.; Wu, A.; Jiang, T.; et al. Identification and Characterization of CircRNAs Encoded by MERS-CoV, SARS-CoV-1 and SARS-CoV-2. Brief. Bioinform. 2021, 22, 1297–1308. [Google Scholar] [CrossRef]
- Yang, M.; Qi, M.; Xu, L.; Huang, P.; Wang, X.; Sun, J.; Shi, J.; Hu, Y. Differential Host CircRNA Expression Profiles in Human Lung Epithelial Cells Infected with SARS-CoV-2. Infect. Genet. Evol. 2021, 93, 104923. [Google Scholar] [CrossRef]
- Zhang, M.; Bian, Z. The Emerging Role of Circular RNAs in Alzheimer’s Disease and Parkinson’s Disease. Front. Aging Neurosci. 2021, 13, 426. [Google Scholar] [CrossRef]
- Doxakis, E. Insights into the Multifaceted Role of Circular RNAs: Implications for Parkinson’s Disease Pathogenesis and Diagnosis. NPJ Parkinson’s Dis. 2022, 8, 7. [Google Scholar] [CrossRef]
- Cochran, K.R.; Veeraraghavan, K.; Kundu, G.; Mazan-Mamczarz, K.; Coletta, C.; Thambisetty, M.; Gorospe, M.; De, S. Systematic Identification of CircRNAs in Alzheimer’s Disease. Genes 2021, 12, 1258. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Safari, M.; Taheri, M.; Samadian, M. Expression of Linear and Circular LncRNAs in Alzheimer’s Disease. J. Mol. Neurosci. 2021, 72, 187–200. [Google Scholar] [CrossRef]
- Hanan, M.; Simchovitz, A.; Yayon, N.; Vaknine, S.; Cohen-Fultheim, R.; Karmon, M.; Madrer, N.; Rohrlich, T.M.; Maman, M.; Bennett, E.R.; et al. A Parkinson’s Disease Circ RNA s Resource Reveals a Link between Circ SLC 8A1 and Oxidative Stress. EMBO Mol. Med. 2020, 12, e11942. [Google Scholar] [CrossRef]
- Falaleeva, M.; Stamm, S. Processing of SnoRNAs as a New Source of Regulatory Non-Coding RNAs: SnoRNA Fragments Form a New Class of Functional RNAs. BioEssays 2013, 35, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Martens-Uzunova, E.S.; Olvedy, M.; Jenster, G. Beyond MicroRNA-Novel RNAs Derived from Small Non-Coding RNA and Their Implication in Cancer. Cancer Lett. 2013, 340, 201–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mleczko, A.M.; Bąkowska-Żywicka, K. When Small RNAs Become Smaller: Non-Canonical Functions of SnoRNAs and Their Derivatives. Acta Biochim. Pol. 2016, 63, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Wajahat, M.; Bracken, C.P.; Orang, A. Emerging Functions for SnoRNAs and SnoRNA-Derived Fragments. Int. J. Mol. Sci. 2021, 22, 10193. [Google Scholar] [CrossRef] [PubMed]
- Kishore, S.; Khanna, A.; Zhang, Z.; Hui, J.; Balwierz, P.J.; Stefan, M.; Beach, C.; Nicholls, R.D.; Zavolan, M.; Stamm, S. The SnoRNA MBII-52 (SNORD 115) Is Processed into Smaller RNAs and Regulates Alternative Splicing. Hum. Mol. Genet. 2010, 19, 1153–1164. [Google Scholar] [CrossRef] [Green Version]
- Ender, C.; Krek, A.; Friedländer, M.R.; Beitzinger, M.; Weinmann, L.; Chen, W.; Pfeffer, S.; Rajewsky, N.; Meister, G. A Human SnoRNA with MicroRNA-Like Functions. Mol. Cell 2008, 32, 519–528. [Google Scholar] [CrossRef]
- Abel, Y.; Rederstorff, M. SnoRNAs and the Emerging Class of SdRNAs: Multifaceted Players in Oncogenesis. Biochimie 2019, 164, 17–21. [Google Scholar] [CrossRef]
- Bratkovič, T.; Rogelj, B. The Many Faces of Small Nucleolar RNAs. Biochim. Et Biophys. Acta 2014, 1839, 438–443. [Google Scholar] [CrossRef]
- Taft, R.J.; Glazov, E.A.; Lassmann, T.; Hayashizaki, Y.; Carninci, P.; Mattick, J.S. Small RNAs Derived from SnoRNAs. RNA 2009, 15, 1233–1240. [Google Scholar] [CrossRef] [Green Version]
- Langenberger, D.; Çakir, M.V.; Hoffmann, S.; Stadler, P.F. Dicer-Processed Small RNAs: Rules and Exceptions. J. Exp. Zool. B Mol. Dev. Evol. 2013, 320, 35–46. [Google Scholar] [CrossRef]
- Brameier, M.; Herwig, A.; Reinhardt, R.; Walter, L.; Gruber, J. Human Box C/D SnoRNAs with MiRNA like Functions: Expanding the Range of Regulatory RNAs. Nucleic Acids Res. 2011, 39, 675–686. [Google Scholar] [CrossRef]
- Scott, M.S.; Avolio, F.; Ono, M.; Lamond, A.I.; Barton, G.J. Human MiRNA Precursors with Box H/ACA SnoRNA Features. PLOS Comput. Biol. 2009, 5, e1000507. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, T.; del Gaudio, D.; German, J.R.; Shinawi, M.; Peters, S.U.; Person, R.E.; Garnica, A.; Cheung, S.W.; Beaudet, A.L. Prader-Willi Phenotype Caused by Paternal Deficiency for the HBII-85 C/D Box Small Nucleolar RNA Cluster. Nat. Genet. 2008, 40, 719–721. [Google Scholar] [CrossRef] [Green Version]
- Bazeley, P.S.; Shepelev, V.; Talebizadeh, Z.; Butler, M.G.; Fedorova, L.; Filatov, V.; Fedorov, A. SnoTARGET Shows That Human Orphan SnoRNA Targets Locate Close to Alternative Splice Junctions. Gene 2008, 408, 172–179. [Google Scholar] [CrossRef]
- Scott, M.S.; Ono, M.; Yamada, K.; Endo, A.; Barton, G.J.; Lamond, A.I. Human Box C/D SnoRNA Processing Conservation across Multiple Cell Types. Nucleic Acids Res. 2012, 40, 3676–3688. [Google Scholar] [CrossRef] [Green Version]
- Ono, M.; Yamada, K.; Avolio, F.; Scott, M.S.; van Koningsbruggen, S.; Barton, G.J.; Lamond, A.I. Analysis of Human Small Nucleolar RNAs (SnoRNA) and the Development of SnoRNA Modulator of Gene Expression Vectors. Mol. Biol. Cell. 2010, 21, 1569–1584. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Shi, Q.; Shen, Q.; Zhang, Q.; Cao, X. Dicer-Independent SnRNA/SnoRNA-Derived Nuclear RNA 3 Regulates Tumor-Associated Macrophage Function by Epigenetically Repressing Inducible Nitric Oxide Synthase Transcription. Cancer Commun. 2021, 41, 140–153. [Google Scholar] [CrossRef]
- Mleczko, A.M.; Machtel, P.; Walkowiak, M.; Wasilewska, A.; Pietras, P.J.; Bąkowska-Żywicka, K. Levels of SdRNAs in Cytoplasm and Their Association with Ribosomes Are Dependent upon Stress Conditions but Independent from SnoRNA Expression. Sci. Rep. 2019, 9, 18397. [Google Scholar] [CrossRef] [Green Version]
- Chow, R.D.; Chen, S. Sno-Derived RNAs Are Prevalent Molecular Markers of Cancer Immunity. Oncogene 2018, 37, 6442–6462. [Google Scholar] [CrossRef]
- Martens-Uzunova, E.S.; Hoogstrate, Y.; Kalsbeek, A.; Pigmans, B.; Vredenbregt-Van Den Berg, M.; Dits, N.; Nielsen, S.J.; Baker, A.; Visakorpi, T.; Bangma, C.; et al. C/D-Box SnoRNA-Derived RNA Production Is Associated with Malignant Transformation and Metastatic Progression in Prostate Cancer. Oncotarget 2015, 6, 17430–17444. [Google Scholar] [CrossRef] [Green Version]
- Mourtada-Maarabouni, M.; Pickard, M.R.; Hedge, V.L.; Farzaneh, F.; Williams, G.T. GAS5, a Non-Protein-Coding RNA, Controls Apoptosis and Is Downregulated in Breast Cancer. Oncogene 2008, 28, 195–208. [Google Scholar] [CrossRef] [Green Version]
- Patterson, D.G.; Roberts, J.T.; King, V.M.; Houserova, D.; Barnhill, E.C.; Crucello, A.; Polska, C.J.; Brantley, L.W.; Kaufman, G.C.; Nguyen, M.; et al. Human SnoRNA-93 Is Processed into a MicroRNA-like RNA That Promotes Breast Cancer Cell Invasion. NPJ Breast Cancer 2017, 3, 25. [Google Scholar] [CrossRef]
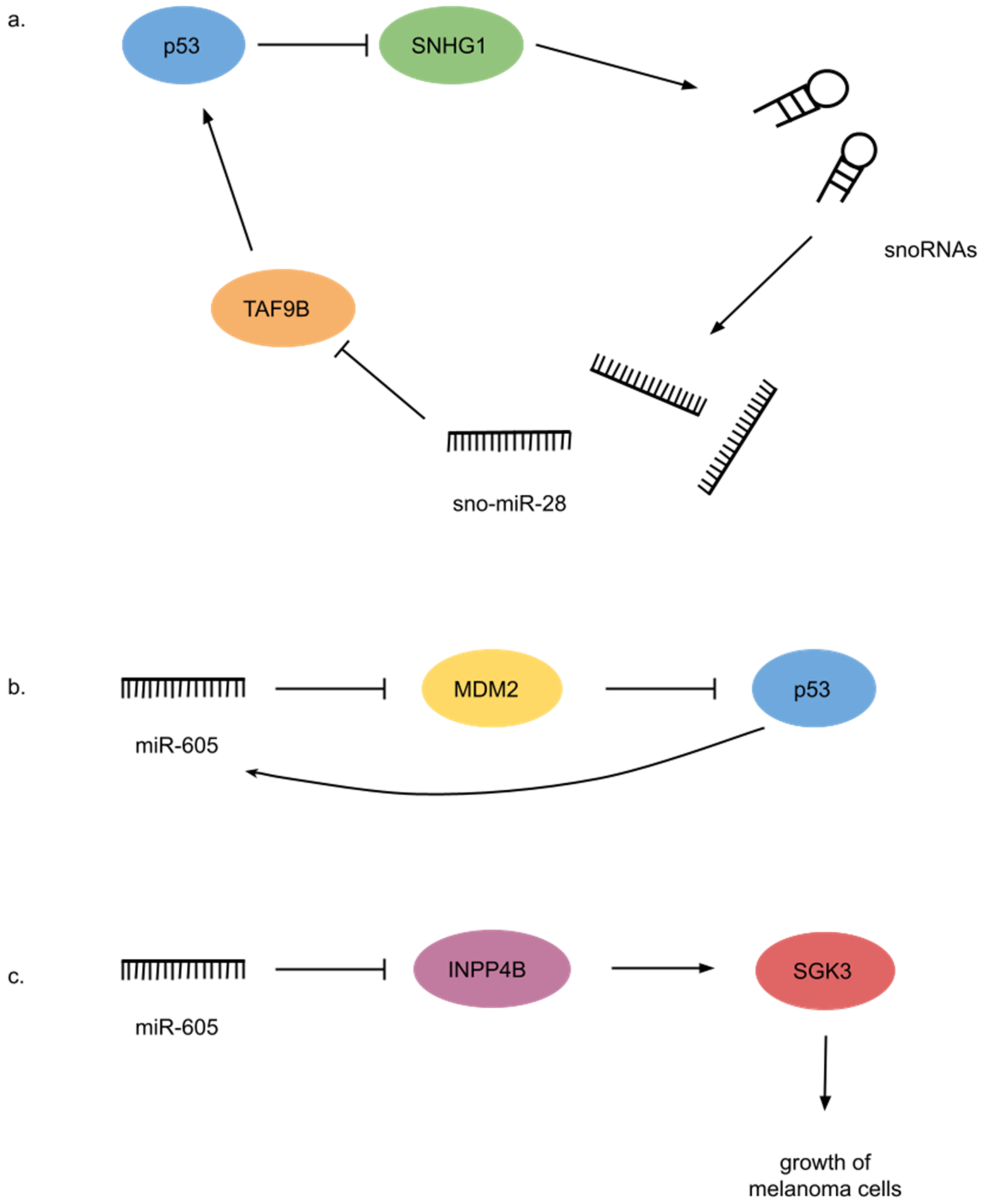
- Yu, F.; Bracken, C.P.; Pillman, K.A.; Lawrence, D.M.; Goodall, G.J.; Callen, D.F.; Neilsen, P.M. P53 Represses the Oncogenic Sno-MiR-28 Derived from a SnoRNA. PLoS ONE 2015, 10, e0129190. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Lin, H.; Luo, X.; Luo, X.; Wang, Z. MiR-605 Joins P53 Network to Form a P53:MiR-605:Mdm2 Positive Feedback Loop in Response to Stress. EMBO J. 2011, 30, 524. [Google Scholar] [CrossRef]
- Chen, L.; Cao, Y.; Rong, D.; Wang, Y.; Cao, Y. MicroRNA-605 Functions as a Tumor Suppressor by Targeting INPP4B in Melanoma. Oncol. Rep. 2017, 38, 1276–1286. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.-Z.; Zhou, A.; Hu, Z.; Yu, A.-M. Small Nucleolar RNA-Derived MicroRNA Hsa-MiR-1291 Modulates Cellular Drug Disposition through Direct Targeting of ABC Transporter ABCC1. Drug Metab. Dispos. 2013, 41, 1744–1751. [Google Scholar] [CrossRef]
- Zong, T.; Yang, Y.; Zhao, H.; Li, L.; Liu, M.; Fu, X.; Tang, G.; Zhou, H.; Aung, L.H.H.; Li, P.; et al. TsRNAs: Novel Small Molecules from Cell Function and Regulatory Mechanism to Therapeutic Targets. Cell Prolif. 2021, 54, e12977. [Google Scholar] [CrossRef]
- Yu, X.; Xie, Y.; Zhang, S.; Song, X.; Xiao, B.; Yan, Z. TRNA-Derived Fragments: Mechanisms Underlying Their Regulation of Gene Expression and Potential Applications as Therapeutic Targets in Cancers and Virus Infections. Theranostics 2020, 11, 461–469. [Google Scholar] [CrossRef]
- Liu, B.; Cao, J.; Wang, X.; Guo, C.; Liu, Y.; Wang, T. Deciphering the TRNA-Derived Small RNAs: Origin, Development, and Future. Cell Death Dis. 2022, 13, 24. [Google Scholar] [CrossRef]
- Kumar, P.; Kuscu, C.; Dutta, A. Biogenesis and Function of Transfer RNA-Related Fragments (TRFs). Trends Biochem. Sci. 2016, 41, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Han, T.; Li, G. Novel Insights into the Roles of TRNA-Derived Small RNAs. RNA Biol. 2021, 18, 2157–2167. [Google Scholar] [CrossRef]
- Telonis, A.G.; Loher, P.; Honda, S.; Jing, Y.; Palazzo, J.; Kirino, Y.; Rigoutsos, I. Dissecting TRNA-Derived Fragment Complexities Using Personalized Transcriptomes Reveals Novel Fragment Classes and Unexpected Dependencies. Oncotarget 2015, 6, 24797–24822. [Google Scholar] [CrossRef] [Green Version]
- Thompson, D.M.; Lu, C.; Green, P.J.; Parker, R. TRNA Cleavage Is a Conserved Response to Oxidative Stress in Eukaryotes. RNA 2008, 14, 2095–2103. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Yeom, J.H.; Kay, M.A. Transfer RNA-Derived Small RNAs: Another Layer of Gene Regulation and Novel Targets for Disease Therapeutics. Mol. Ther. 2020, 28, 2340–2357. [Google Scholar] [CrossRef]
- Yamasaki, S.; Ivanov, P.; Hu, G.F.; Anderson, P. Angiogenin Cleaves TRNA and Promotes Stress-Induced Translational Repression. J. Cell Biol. 2009, 185, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Lyons, S.M.; Achorn, C.; Kedersha, N.L.; Anderson, P.J.; Ivanov, P. YB-1 Regulates TiRNA-Induced Stress Granule Formation but Not Translational Repression. Nucleic Acids Res. 2016, 44, 6949–6960. [Google Scholar] [CrossRef]
- Li, S.; Xu, Z.; Sheng, J. TRNA-Derived Small RNA: A Novel Regulatory Small Non-Coding RNA. Genes 2018, 9, 246. [Google Scholar] [CrossRef] [Green Version]
- Farny, N.G.; Kedersha, N.L.; Silver, P.A. Metazoan Stress Granule Assembly Is Mediated by P-EIF2α-Dependent and -Independent Mechanisms. RNA 2009, 15, 1814–1821. [Google Scholar] [CrossRef] [Green Version]
- Lyons, S.M.; Kharel, P.; Akiyama, Y.; Ojha, S.; Dave, D.; Tsvetkov, V.; Merrick, W.; Ivanov, P.; Anderson, P. EIF4G Has Intrinsic G-Quadruplex Binding Activity That Is Required for TiRNA Function. Nucleic Acids Res. 2020, 48, 6223–6233. [Google Scholar] [CrossRef]
- Shaukat, A.N.; Kaliatsi, E.G.; Stamatopoulou, V.; Stathopoulos, C. Mitochondrial TRNA-Derived Fragments and Their Contribution to Gene Expression Regulation. Front. Physiol. 2021, 12, 1425. [Google Scholar] [CrossRef]
- Honda, S.; Loher, P.; Shigematsu, M.; Palazzo, J.P.; Suzuki, R.; Imoto, I.; Rigoutsos, I.; Kirino, Y. Sex Hormone-Dependent TRNA Halves Enhance Cell Proliferation in Breast and Prostate Cancers. Proc. Natl. Acad. Sci. USA 2015, 112, E3816–E3825. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, H.; Liu, X.; Nguyen, H.C.B.; Zhang, S.; Fish, L.; Tavazoie, S.F. Endogenous TRNA-Derived Fragments Suppress Breast Cancer Progression via YBX1 Displacement. Cell 2015, 161, 790–802. [Google Scholar] [CrossRef] [Green Version]
- Lasham, A.; Samuel, W.; Cao, H.; Patel, R.; Mehta, R.; Stern, J.L.; Reid, G.; Woolley, A.G.; Miller, L.D.; Black, M.A.; et al. YB-1, the E2F Pathway, and Regulation of Tumor Cell Growth. J. Natl. Cancer Inst. 2012, 104, 133. [Google Scholar] [CrossRef] [Green Version]
- Maute, R.L.; Schneider, C.; Sumazin, P.; Holmes, A.; Califano, A.; Basso, K.; Dalla-Favera, R. TRNA-Derived MicroRNA Modulates Proliferation and the DNA Damage Response and Is down-Regulated in B Cell Lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 1404–1409. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Shi, J.; Wu, Z.; Gao, P.; Zhang, W.; Qu, B.; Wang, X.; Song, Y.; Wang, Z. A 3′-TRNA-Derived Fragment Enhances Cell Proliferation, Migration and Invasion in Gastric Cancer by Targeting FBXO47. Arch. Biochem. Biophys. 2020, 690, 108467. [Google Scholar] [CrossRef]
- Lu, Z.; Su, K.; Wang, X.; Zhang, M.; Ma, S.; Li, H.; Qiu, Y. Expression Profiles of TRNA-Derived Small RNAs and Their Potential Roles in Primary Nasopharyngeal Carcinoma. Front. Mol. Biosci. 2021, 8, 780621. [Google Scholar] [CrossRef]
- Chen, H.; Xu, Z.; Cai, H.; Peng, Y.; Yang, L.; Wang, Z. Identifying Differentially Expressed TRNA-Derived Small Fragments as a Biomarker for the Progression and Metastasis of Colorectal Cancer. Dis Markers 2022, 2022, 8. [Google Scholar] [CrossRef]
- Panoutsopoulou, K.; Dreyer, T.; Dorn, J.; Obermayr, E.; Mahner, S.; van Gorp, T.; Braicu, I.; Zeillinger, R.; Magdolen, V.; Avgeris, M.; et al. TRNAGlyGCC-Derived Internal Fragment (i-TRF-GlyGCC) in Ovarian Cancer Treatment Outcome and Progression. Cancers 2021, 14, 24. [Google Scholar] [CrossRef]
- Yeung, M.L.; Bennasser, Y.; Watashi, K.; Le, S.-Y.; Houzet, L.; Jeang, K.-T. Pyrosequencing of Small Non-Coding RNAs in HIV-1 Infected Cells: Evidence for the Processing of a Viral-Cellular Double-Stranded RNA Hybrid. Nucleic Acids Res. 2009, 37, 6575–6586. [Google Scholar] [CrossRef]
- Ruggero, K.; Guffanti, A.; Corradin, A.; Sharma, V.K.; de Bellis, G.; Corti, G.; Grassi, A.; Zanovello, P.; Bronte, V.; Ciminale, V.; et al. Small Noncoding RNAs in Cells Transformed by Human T-Cell Leukemia Virus Type 1: A Role for a TRNA Fragment as a Primer for Reverse Transcriptase. J. Virol. 2014, 88, 3612–3622. [Google Scholar] [CrossRef] [Green Version]
- Wang, E.; Thombre, R.; Shah, Y.; Latanich, R.; Wang, J. G-Quadruplexes as Pathogenic Drivers in Neurodegenerative Disorders. Nucleic Acids Res. 2021, 49, 4816. [Google Scholar] [CrossRef]
- Ivanov, P.; O’Day, E.; Emara, M.M.; Wagner, G.; Lieberman, J.; Anderson, P. G-Quadruplex Structures Contribute to the Neuroprotective Effects of Angiogenin-Induced TRNA Fragments. Proc. Natl. Acad. Sci. USA 2014, 111, 18201–18206. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, M.; Pollex, T.; Hanna, K.; Tuorto, F.; Meusburger, M.; Helm, M.; Lyko, F. RNA Methylation by Dnmt2 Protects Transfer RNAs against Stress-Induced Cleavage. Genes Dev. 2010, 24, 1590–1595. [Google Scholar] [CrossRef] [Green Version]
- Blanco, S.; Dietmann, S.; Flores, J.V.; Hussain, S.; Kutter, C.; Humphreys, P.; Lukk, M.; Lombard, P.; Treps, L.; Popis, M.; et al. Aberrant Methylation of TRNAs Links Cellular Stress to Neuro-Developmental Disorders. EMBO J. 2014, 33, 2020–2039. [Google Scholar] [CrossRef]
- Wu, W.; Lee, I.; Spratt, H.; Fang, X.; Bao, X. TRNA-Derived Fragments in Alzheimer’s Disease: Implications for New Disease Biomarkers and Neuropathological Mechanisms. J. Alzheimers Dis. 2021, 79, 793. [Google Scholar] [CrossRef]
- Li, Z.; Stanton, B.A. Transfer RNA-Derived Fragments, the Underappreciated Regulatory Small RNAs in Microbial Pathogenesis. Front. Microbiol. 2021, 12, 687632. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Function | miRNA Target | miRNA Mechanism of Action | References |
|---|---|---|---|---|
| miR-522 | TGS | CYP2E1 promoter | DNA:RNA hybrid with promoter | [24] |
| miR-223 | TGS | NFI-A promoter | DNA:RNA hybrid with promoter | [25] |
| let-7i | TGA | Interleukin-2 promoter | Binds to TATA-box and enhances promoter activities | [26] |
| miR-138 | TGA | Insulin promoter | Binds to TATA-box and enhances promoter activities | [26] |
| miR-92a | TGA | Calcitonin promoter | Binds to TATA-box and enhances promoter activities | [26] |
| miR-181d | TGA | C-myc promoter | Binds to TATA-box and enhances promoter activities | [26] |
| miR-373 | TGA | E-cadherin, CSDC2 | Pol II enrichment at promoters | [27] |
| miR-24-1 | TGA | FBP1, FANCC | Chromatin state alteration of the FBP1 enhancer | [28] |
| miRNA | miRNA Expression in Cancer | miRNA Target | Function | Type of Cancer | References |
|---|---|---|---|---|---|
| let-7 | Downregulated | PD-L1, HMGA1, apoptotic genes | tumor suppressor | various cancers | [45] |
| miR15/16 | Downregulated | BCL2 | tumor suppressor | CLL | [46] |
| miR-21 | Upregulated | Maps, PDCD4, TPM1, PTEN | oncogene | lung, breast, and bladder cancer | [47,48,49] |
| miR-155 | Upregulated | signaling pathways (TGF-β, JAK-STAT) | oncogene | breast cancer | [50,51] |
| miR-125b | upregulated/downregulated | multiple mRNAs with diverse functions in different tissues | oncogene/tumor suppressor | colon cancer, hematopoietic tumors/NSCLC, breast cancer | [52,53,54] |
| miR-17-92 | upregulated/downregulated | E2F transcription factors/AIB1, ETV1, ERBB3 | oncogene/tumor suppressor | lymphoma, lung cancer, colon cancer, pancreatic cancer, prostate cancer, HNSCC/breast cancer, HCC | [55,56,57,58,59] |
| circRNA | miRNA Target | Disease Associated | circRNA Expression | circRNA Function | References |
|---|---|---|---|---|---|
| ciRS-7 (CDR1as) | miR-7 | Several cancers including NSCLC and CRC | upregulated | oncogene | [94,115,116,117] |
| circHIPK3 | miR-7, miR-124, miR-506 | CRC, GBC, HCC | upregulated | oncogene | [118,119,120] |
| circITCH | miR-7, miR-214 | CRC, lung cancer, NPC | downregulated | tumor suppressor | [104,121,122] |
| circNT5E | miR-422a, miR-134, miR-502-5p | Glioma, NSCLC, bladder cancer | upregulated | oncogene | [123,124,125] |
| circDENND2A | miR-625-5p | Glioma | upregulated | oncogene | [126] |
| circDENND4C | miR-200b, miR-200c, miR-141-3p | Breast cancer, NSCLC, lung cancer | upregulated | oncogene | [127,128] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plawgo, K.; Raczynska, K.D. Context-Dependent Regulation of Gene Expression by Non-Canonical Small RNAs. Non-Coding RNA 2022, 8, 29. https://doi.org/10.3390/ncrna8030029
Plawgo K, Raczynska KD. Context-Dependent Regulation of Gene Expression by Non-Canonical Small RNAs. Non-Coding RNA. 2022; 8(3):29. https://doi.org/10.3390/ncrna8030029
Chicago/Turabian StylePlawgo, Kinga, and Katarzyna Dorota Raczynska. 2022. "Context-Dependent Regulation of Gene Expression by Non-Canonical Small RNAs" Non-Coding RNA 8, no. 3: 29. https://doi.org/10.3390/ncrna8030029
APA StylePlawgo, K., & Raczynska, K. D. (2022). Context-Dependent Regulation of Gene Expression by Non-Canonical Small RNAs. Non-Coding RNA, 8(3), 29. https://doi.org/10.3390/ncrna8030029