
Activity-Dependent Non-Coding RNA MAPK Interactome of the Human Epileptic Brain



Abstract
:1. Introduction
2. Results
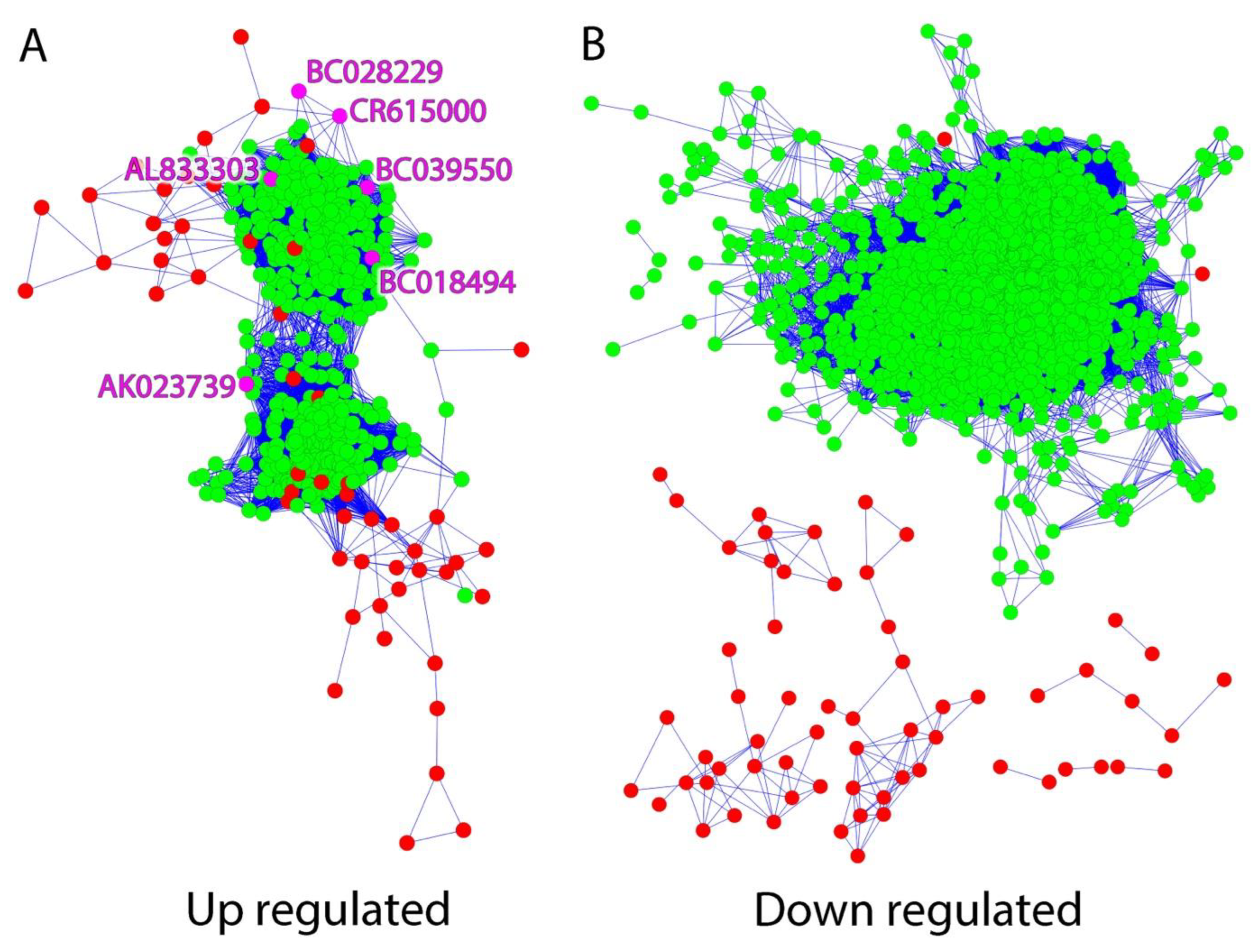
2.1. LncRNAs Are Co-Expressed within the MAPK Signaling Interactome in Human Epileptic Neocortical Tissue
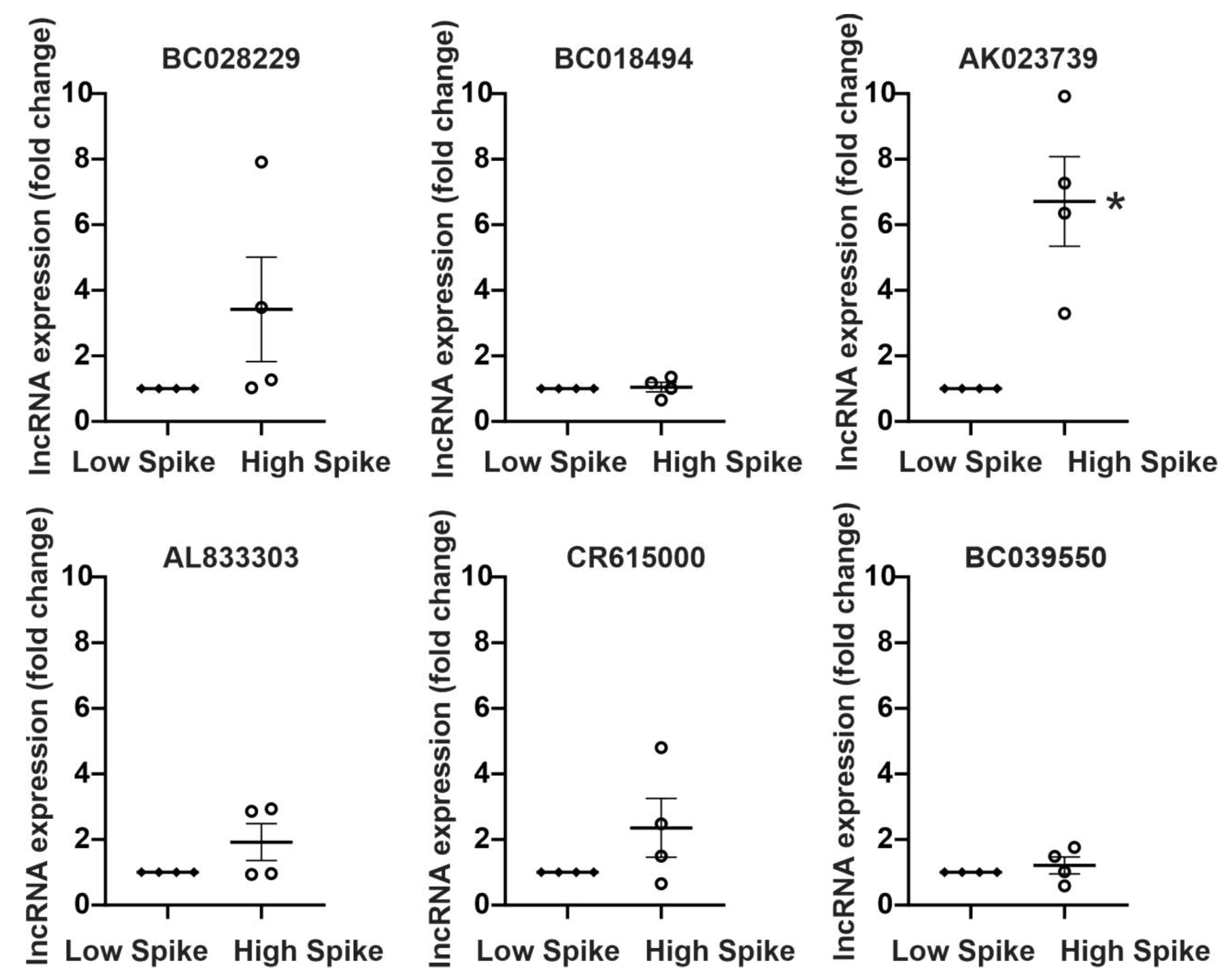
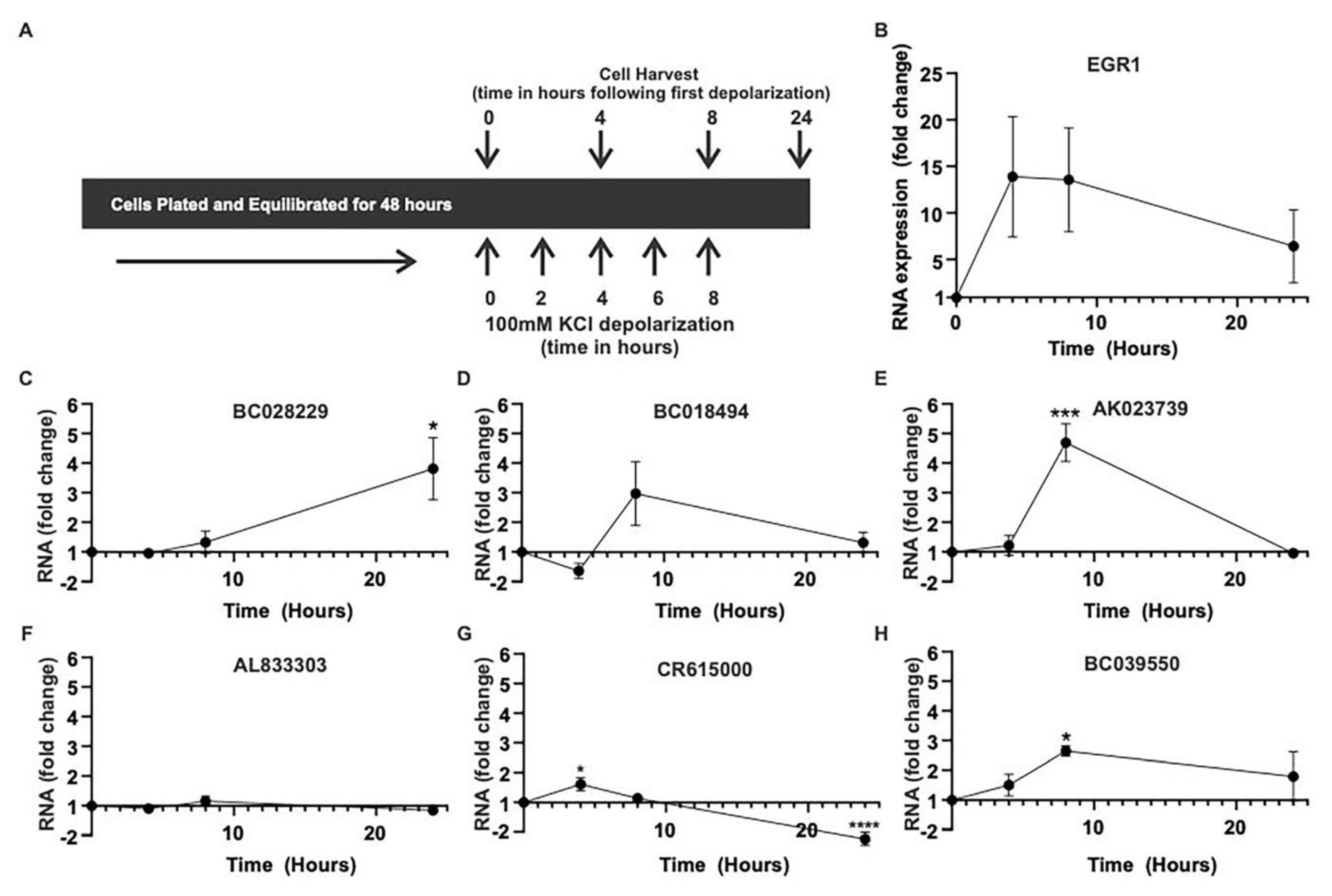
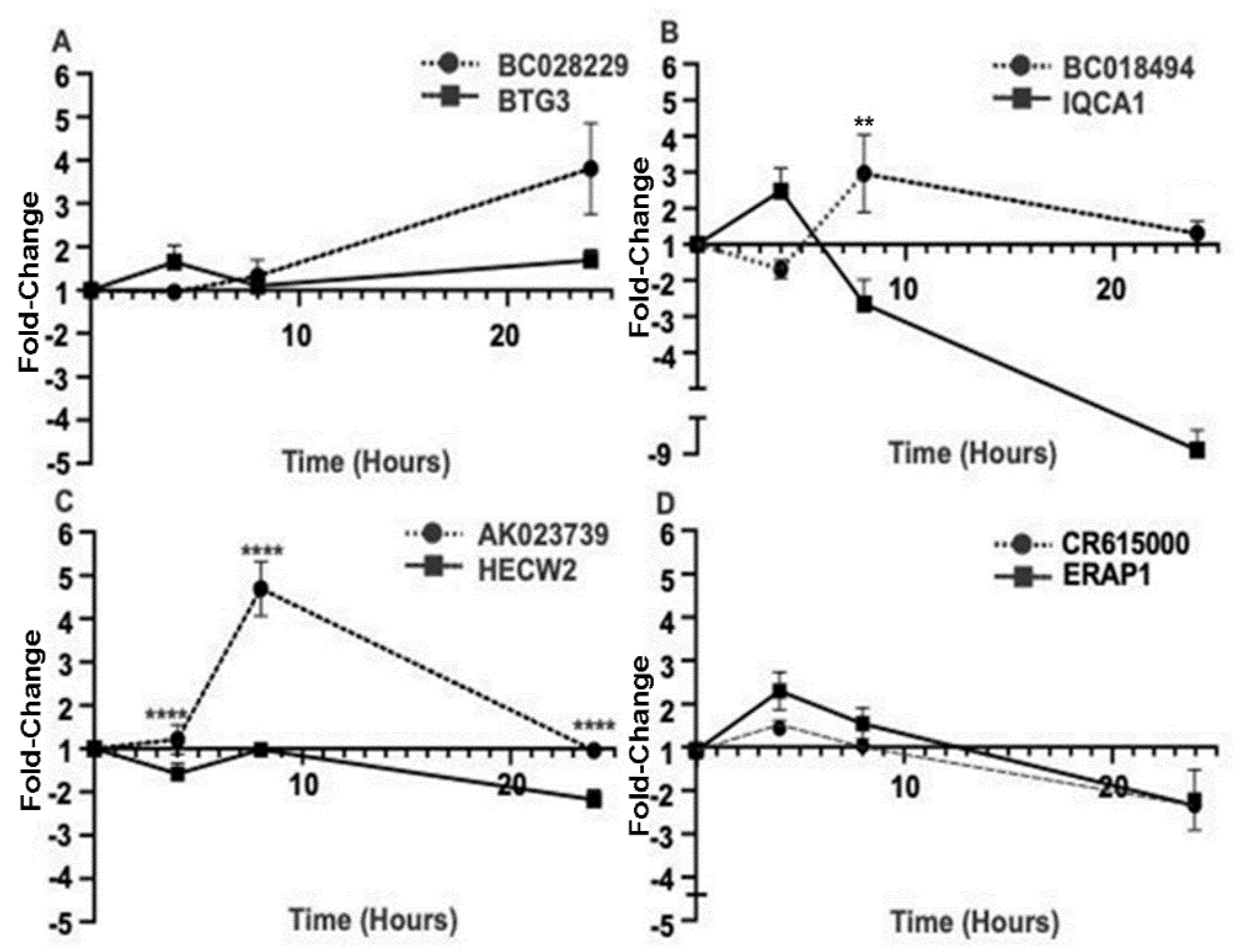
2.2. Activity-Dependent MAPK Signaling Increases the Levels of Specific lncRNAs In Vitro
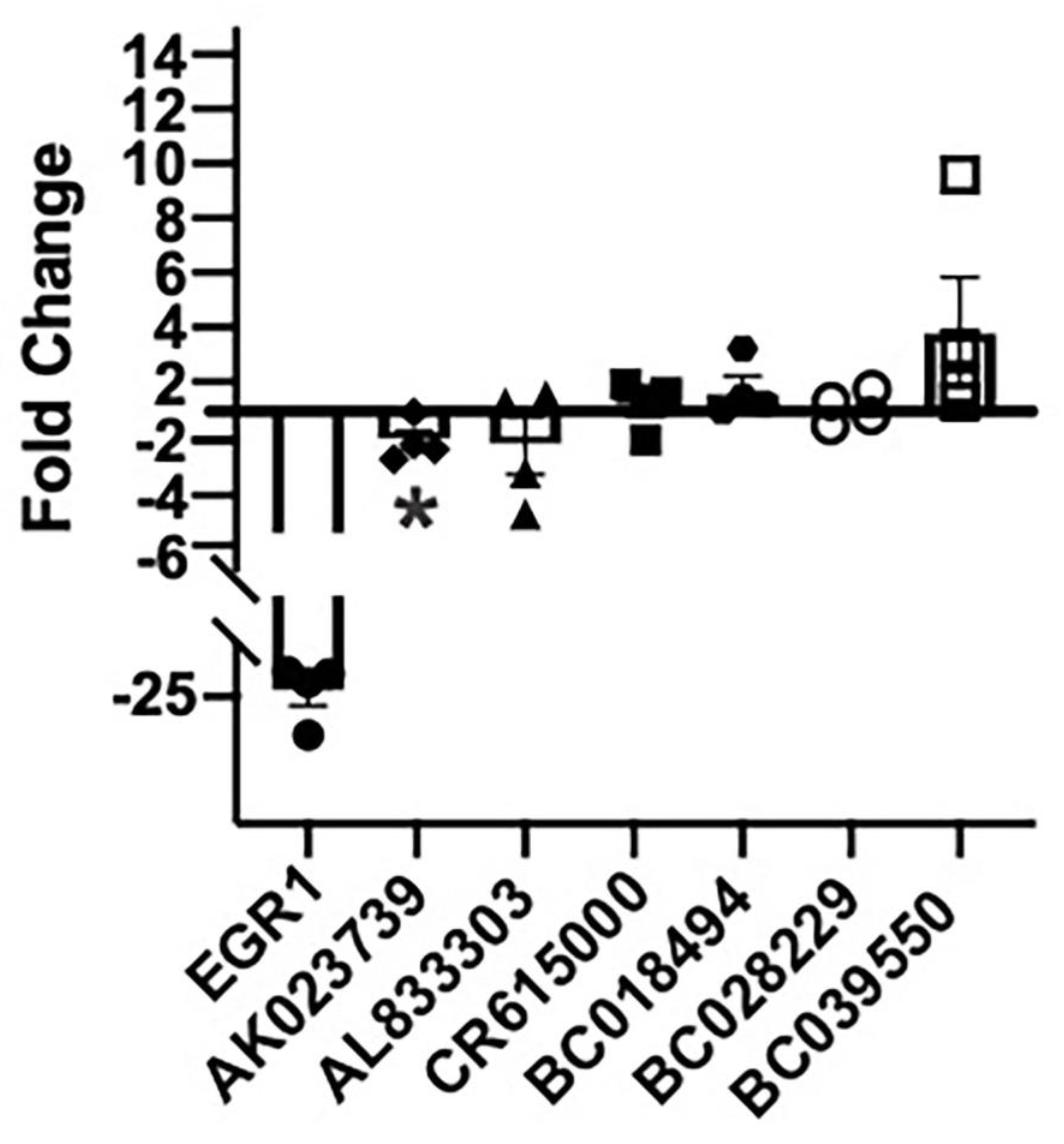
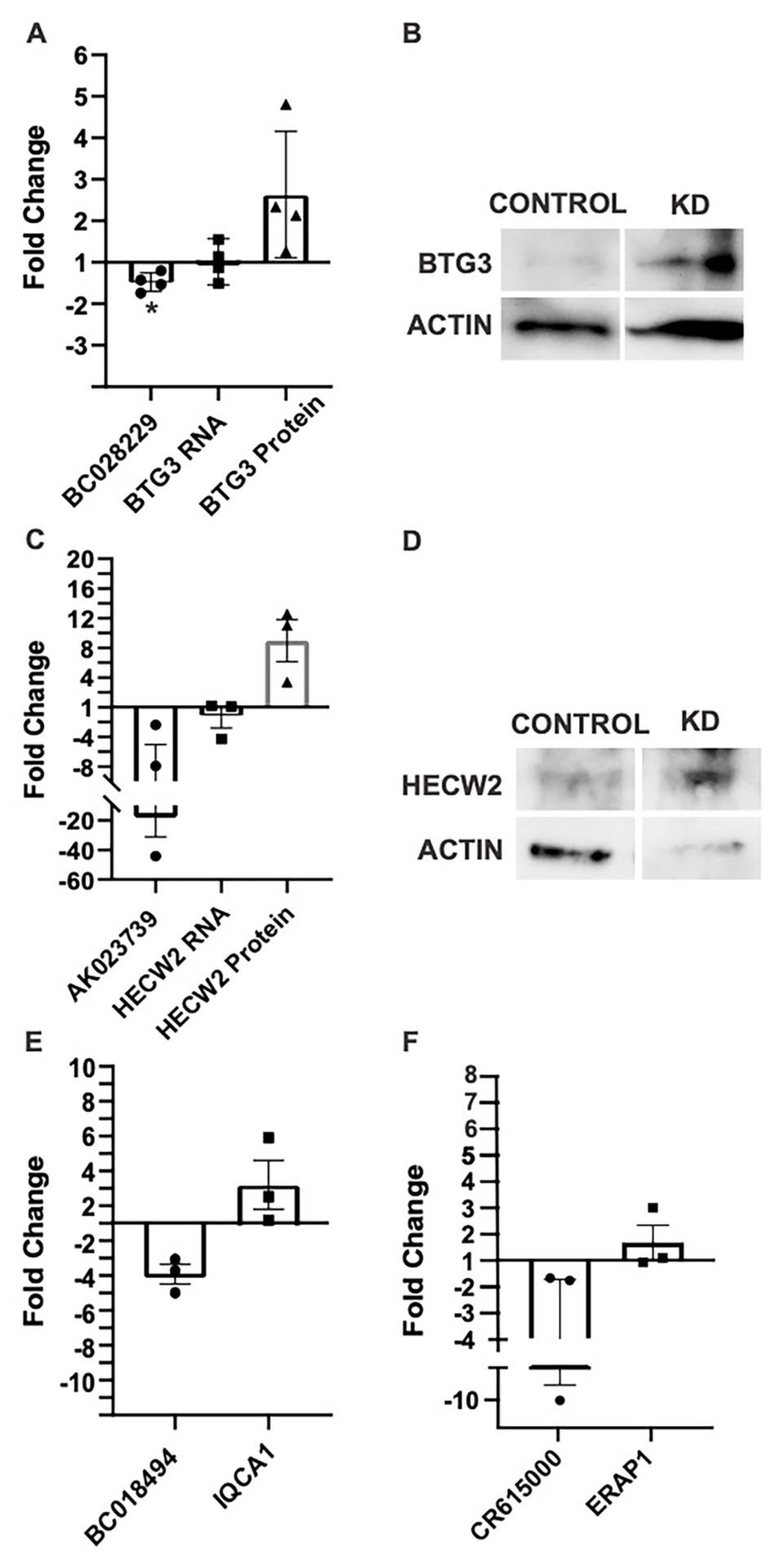
2.3. Antisense MAPK lncRNAs Downregulate Their Overlapping Protein-Coding Genes
3. Discussion
4. Materials and Methods
4.1. Electrically Mapped Human Epileptic Neocortical Tissues
4.2. Microarray and Gene Clustering
4.3. Cell Culture, Reagents, Depolarizations, and Electroporation
4.4. RNA Extraction, Reverse Transcription, and qPCR
4.5. Western Blots
4.6. Statistical Analysis and Experimental Design
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carninci, P.; Hayashizaki, Y. Noncoding RNA transcription beyond annotated genes. Curr. Opin. Genet. Dev. 2006, 17, 139–144. [Google Scholar] [CrossRef]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef]
- Dinger, M.E.; Pang, K.C.; Mercer, T.R.; Mattick, J.S. Differentiating Protein-Coding and Noncoding RNA: Challenges and Ambiguities. PLoS Comput. Biol. 2008, 4, e1000176. [Google Scholar] [CrossRef] [Green Version]
- Bánfai, B.; Jia, H.; Khatun, J.; Wood, E.; Risk, B.; Gundling, J.; William, E.; Kundaje, A.; Gunawardena, H.P.; Yu, Y.; et al. Long noncoding RNAs are rarely translated in two human cell lines. Genome Res. 2012, 22, 1646–1657. [Google Scholar] [CrossRef] [Green Version]
- Choo, S.-W.; Zhong, Y.; Sendler, E.; Goustin, A.-S.; Cai, J.; Ju, D.; Kosir, M.A.; Giordo, R.; Lipovich, L. Estrogen distinctly regulates transcription and translation of lncRNAs and pseudogenes in breast cancer cells. Genomics 2022, 114, 110421. [Google Scholar] [CrossRef]
- Babak, T.; Blencowe, B.J.; Hughes, T.R. A systematic search for new mammalian noncoding RNAs indicates little conserved intergenic transcription. BMC Genom. 2005, 6, 104. [Google Scholar] [CrossRef] [Green Version]
- Harrow, J.; Frankish, A.; Gonzalez, J.M.; Tapanari, E.; Diekhans, M.; Kokocinski, F.; Aken, B.L.; Barrell, D.; Zadissa, A.; Searle, S.; et al. GENCODE: The reference human genome annotation for The ENCODE Project. Genome Res. 2012, 22, 1760–1774. [Google Scholar] [CrossRef] [Green Version]
- Carninci, P.; Sandelin, A.; Lenhard, B.; Katayama, S.; Shimokawa, K.; Ponjavic, J.; Semple, C.A.; Taylor, M.S.; Engström, P.G.; Frith, M.C.; et al. Genome-wide analysis of mammalian promoter architecture and evolution. Nat. Genet. 2006, 38, 626–635. [Google Scholar] [CrossRef]
- Necsulea, A.; Kaessmann, H. Evolutionary dynamics of coding and non-coding transcriptomes. Nat. Rev. Genet. 2014, 15, 734–748. [Google Scholar] [CrossRef]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef]
- Clark, M.B.; Mercer, T.R.; Bussotti, G.; Leonardi, T.; Haynes, K.R.; Crawford, J.; Brunck, M.E.; Le Cao, K.-A.; Thomas, G.P.; Chen, W.Y.; et al. Quantitative gene profiling of long noncoding RNAs with targeted RNA sequencing. Nat. Methods 2015, 12, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Lipovich, L.; Dachet, F.; Cai, J.; Bagla, S.; Balan, K.; Jia, H.; Loeb, J.A. Activity-Dependent Human Brain Coding/Noncoding Gene Regulatory Networks. Genetics 2012, 192, 1133–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dachet, F.; Brown, J.B.; Valyi-Nagy, T.; Narayan, K.D.; Serafini, A.; Boley, N.; Gingeras, T.R.; Celniker, S.E.; Mohapatra, G.; Loeb, J.A. Selective time-dependent changes in activity and cell-specific gene expression in human postmortem brain. Sci. Rep. 2021, 11, 6078. [Google Scholar] [CrossRef]
- Dachet, F.; Loeb, J.A. Effect of the post-mortem interval on lncRNA stability in the human brain. In Proceedings of the International Conference on Bioinformatics and Biomedicine Workshop Paper, Houston, TX, USA, 09–12 December 2021. (Virtual). [Google Scholar]
- Lipovich, L.; Johnson, R.; Lin, C.Y. MacroRNA underdogs in a microRNA world: Evolutionary, regulatory, and biomedical significance of mammalian long non-protein-coding RNA. Biochim. Biophys. Acta 2010, 1799, 597–615. [Google Scholar] [CrossRef] [PubMed]
- Sahagan, B.G.; Finch, C.E.; Wood, D.E.; St. Laurent, G., III; Modarresi, F.; Faghihi, M.A. Expression of a noncoding RNA is elevated in Alzheimer’s disease and drives rapid feed-forward regulation of β-secretase. Nat. Med. 2008, 14, 723–730. [Google Scholar]
- Lv, Q.; Wang, Z.; Zhong, Z.; Huang, W. Role of Long Noncoding RNAs in Parkinson’s Disease: Putative Biomarkers and Therapeutic Targets. Park. Dis. 2020, 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bilinovich, S.M.; Lewis, K.; Grepo, N.; Campbell, D.B. The Long Noncoding RNA RPS10P2-AS1 Is Implicated in Autism Spectrum Disorder Risk and Modulates Gene Expression in Human Neuronal Progenitor Cells. Front. Genet. 2019, 10, 970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmani, Z. APRO4 negatively regulates Src tyrosine kinase activity in PC12 cells. J. Cell Sci. 2006, 119, 646–658. [Google Scholar] [CrossRef] [Green Version]
- Beaumont, T.L.; Yao, B.; Shah, A.; Kapatos, G.; Loeb, J.A. Layer-Specific CREB Target Gene Induction in Human Neocortical Epilepsy. J. Neurosci. 2012, 32, 14389–14401. [Google Scholar] [CrossRef] [Green Version]
- Barkmeier, D.T.; Senador, D.; Leclercq, K.; Pai, D.; Hua, J.; Boutros, N.N.; Kaminski, R.M.; Loeb, J.A. Electrical, molecular and behavioral effects of interictal spiking in the rat. Neurobiol. Dis. 2012, 47, 92–101. [Google Scholar] [CrossRef] [Green Version]
- English, J.D.; Sweatt, J.D. Activation of p42 Mitogen-activated Protein Kinase in Hippocampal Long Term Potentiation. J. Biol. Chem. 1996, 271, 24329–24332. [Google Scholar] [CrossRef]
- English, J.D.; Sweatt, J.D. A Requirement for the Mitogen-activated Protein Kinase Cascade in Hippocampal Long Term Potentiation. J. Biol. Chem. 1997, 272, 19103–19106. [Google Scholar] [CrossRef] [Green Version]
- Kirchner, A.; Dachet, F.; Loeb, J.A. Identifying targets for preventing epilepsy using systems biology of the human brain. Neuropharmacology 2019, 168, 107757. [Google Scholar] [CrossRef]
- Kirchner, A.; Bagla, S.; Dachet, F.; Loeb, J. DUSP4 appears to be a highly localized endogenous inhibitor of epileptic signaling in human neocortex. Neurobiol. Dis. 2020, 145, 105073. [Google Scholar] [CrossRef]
- Rosikiewicz, W.; Makałowska, I. Biological Functions of Natural Antisense Transcripts. Acta Biochim. Pol. 2017, 63, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Berko, E.R.; Cho, M.T.; Eng, C.; Shao, Y.; A Sweetser, D.; Waxler, J.; Robin, N.H.; Brewer, F.; Donkervoort, S.; Mohassel, P.; et al. De novo missense variants in HECW2 are associated with neurodevelopmental delay and hypotonia. J. Med. Genet. 2017, 54, 84–86. [Google Scholar] [CrossRef] [Green Version]
- Errichiello, E.; Novara, F.; Cremante, A.; Verri, A.; Galli, J.; Fazzi, E.; Bellotti, D.; Losa, L.; Cisternino, M.; Zuffardi, O. Dissection of partial 21q monosomy in different phenotypes: Clinical and molecular characterization of five cases and review of the literature. Mol. Cytogenet. 2016, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Ullman, N.L.; Smith-Hicks, C.L.; Desai, S.; Stafstrom, C.E. De Novo HECW2 Mutation Associated With Epilepsy, Developmental Decline, and Intellectual Disability: Case Report and Review of Literature. Pediatr. Neurol. 2018, 85, 76–78. [Google Scholar] [CrossRef]
- Miyazaki, K.; Ozaki, T.; Kato, C.; Hanamoto, T.; Fujita, T.; Irino, S. A novel HECT-type E3 ubiquitin ligase, NEDL2, stabilizes p73 and enhances its transcriptional activity. Biochem. Biophys. Res. Commun. 2003, 308, 106–113. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, X.; Gao, Y. The long noncoding RNA MEG3 regulates Ras-MAPK pathway through RASA1 in trophoblast and is associated with unexplained recurrent spontaneous abortion. Mol. Med. 2021, 27, 70. [Google Scholar] [CrossRef]
- Wang, P.; Fu, H.; Cui, J.; Chen, X. Differential lncRNA-mRNA co-expression network analysis revealing the potential regulatory roles of lncRNAs in myocardial infarction. Mol. Med. Rep. 2016, 13, 1195–1203. [Google Scholar] [CrossRef]
- Dashti, S.; Taherian-Esfahani, Z.; Kholghi-Oskooei, V.; Noroozi, R.; Arsang-Jang, S.; Ghafouri-Fard, S. In silico identification of MAPK14-related lncRNAs and assessment of their expression in breast cancer samples. Sci. Rep. 2020, 10, 8316. [Google Scholar] [CrossRef]
- Luo, R.; Huang, X.; Yan, Z.; Gao, X.; Wang, P.; Yang, Q.; Wang, W.; Xie, K.; Gun, S. Identification and Characterization of MAPK Signaling Pathway Genes and Associated lncRNAs in the Ileum of Piglets Infected by Clostridium perfringens Type C. BioMed Res. Int. 2020, 2020, 8496872–8496912. [Google Scholar] [CrossRef]
- Washietl, S.; Kellis, M.; Garber, M. Evolutionary dynamics and tissue specificity of human long noncoding RNAs in six mammals. Genome Res. 2014, 24, 616–628. [Google Scholar] [CrossRef] [Green Version]
- Lyu, Y.; Bai, L.; Qin, C. Long noncoding RNAs in neurodevelopment and Parkinson’s disease. Anim. Model. Exp. Med. 2019, 2, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q.; Chen, Y. Long noncoding RNAs and Alzheimer’s disease. Clin. Interv. Aging 2016, 11, 867–872. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Wu, Z.; Wu, T.; Huang, Y.; Cheng, Z.; Li, X.; Sun, T.; Xie, X.; Zhou, Y.; Du, Z. Tumor-suppressive function of long noncoding RNA MALAT1 in glioma cells by downregulation of MMP2 and inactivation of ERK/MAPK signaling. Cell Death Dis. 2016, 7, e2123. [Google Scholar] [CrossRef] [Green Version]
- Yanagida, S.; Taniue, K.; Sugimasa, H.; Nasu, E.; Takeda, Y.; Kobayashi, M.; Yamamoto, T.; Okamoto, A.; Akiyama, T. ASBEL, an ANA/BTG3 antisense transcript required for tumorigenicity of ovarian carcinoma. Sci. Rep. 2013, 3, 1305. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.M.; Bahal, R.; Rasmussen, T.P.; Manautou, J.E.; Zhong, X. The growth of siRNA-based therapeutics: Updated clinical studies. Biochem. Pharmacol. 2021, 189, 114432. [Google Scholar] [CrossRef]
- Ray, K.K.; Wright, R.S.; Kallend, D.; Koenig, W.; Leiter, L.A.; Raal, F.J.; Bisch, J.A.; Richardson, T.; Jaros, M.; Wijngaard, P.L.; et al. Two Phase 3 Trials of Inclisiran in Patients with Elevated LDL Cholesterol. N. Engl. J. Med. 2020, 382, 1507–1519. [Google Scholar] [CrossRef]
- Eyford, B.A.; Singh, C.S.B.; Abraham, T.; Munro, L.; Choi, K.B.; Hill, T. A Nanomule Peptide Carrier Delivers siRNA Across the Intact Blood-Brain Barrier to Attenuate Ischemic Stroke. Front. Mol. Biosci. 2021, 8, 611367. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Xiao, X.; Deng, X.; Zhang, F.; Zhang, X.; Hu, Q.; Sheng, W. Targeting long non-coding RNA ASBEL with oligonucleotide antagonist for breast cancer therapy. Biochem. Biophys. Res. Commun. 2017, 489, 386–392. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| lncRNA | Fold Change (FC) | False Discovery Rate (FDR) | lncRNA Classification | Overlapping Coding Gene | Mutations Associated with Epilepsy |
|---|---|---|---|---|---|
| BC028229 | 2.1 | 0.00052 | antisense | BTG3 | Yes |
| BC018494 | 1.7 | 1.8 | antisense | IQCA1 | No |
| AK023739 | 1.7 | 3.2 | antisense | HECW2 | Yes |
| AL833303 | 1.5 | 0.00025 | intergenic | N/A | No |
| CR615000 | 1.5 | 3.9 | antisense | ERAP1 | No |
| BC039550 | 1.3 | 0.03 | intergenic | N/A | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirchner, A.; Dachet, F.; Lipovich, L.; Loeb, J.A. Activity-Dependent Non-Coding RNA MAPK Interactome of the Human Epileptic Brain. Non-Coding RNA 2023, 9, 3. https://doi.org/10.3390/ncrna9010003
Kirchner A, Dachet F, Lipovich L, Loeb JA. Activity-Dependent Non-Coding RNA MAPK Interactome of the Human Epileptic Brain. Non-Coding RNA. 2023; 9(1):3. https://doi.org/10.3390/ncrna9010003
Chicago/Turabian StyleKirchner, Allison, Fabien Dachet, Leonard Lipovich, and Jeffrey A. Loeb. 2023. "Activity-Dependent Non-Coding RNA MAPK Interactome of the Human Epileptic Brain" Non-Coding RNA 9, no. 1: 3. https://doi.org/10.3390/ncrna9010003
APA StyleKirchner, A., Dachet, F., Lipovich, L., & Loeb, J. A. (2023). Activity-Dependent Non-Coding RNA MAPK Interactome of the Human Epileptic Brain. Non-Coding RNA, 9(1), 3. https://doi.org/10.3390/ncrna9010003