


Long Non-Coding RNA Expression in Alpha-1 Antitrypsin Deficient Monocytes Pre- and Post-AAT Augmentation Therapy



Abstract
:1. Introduction
2. Results
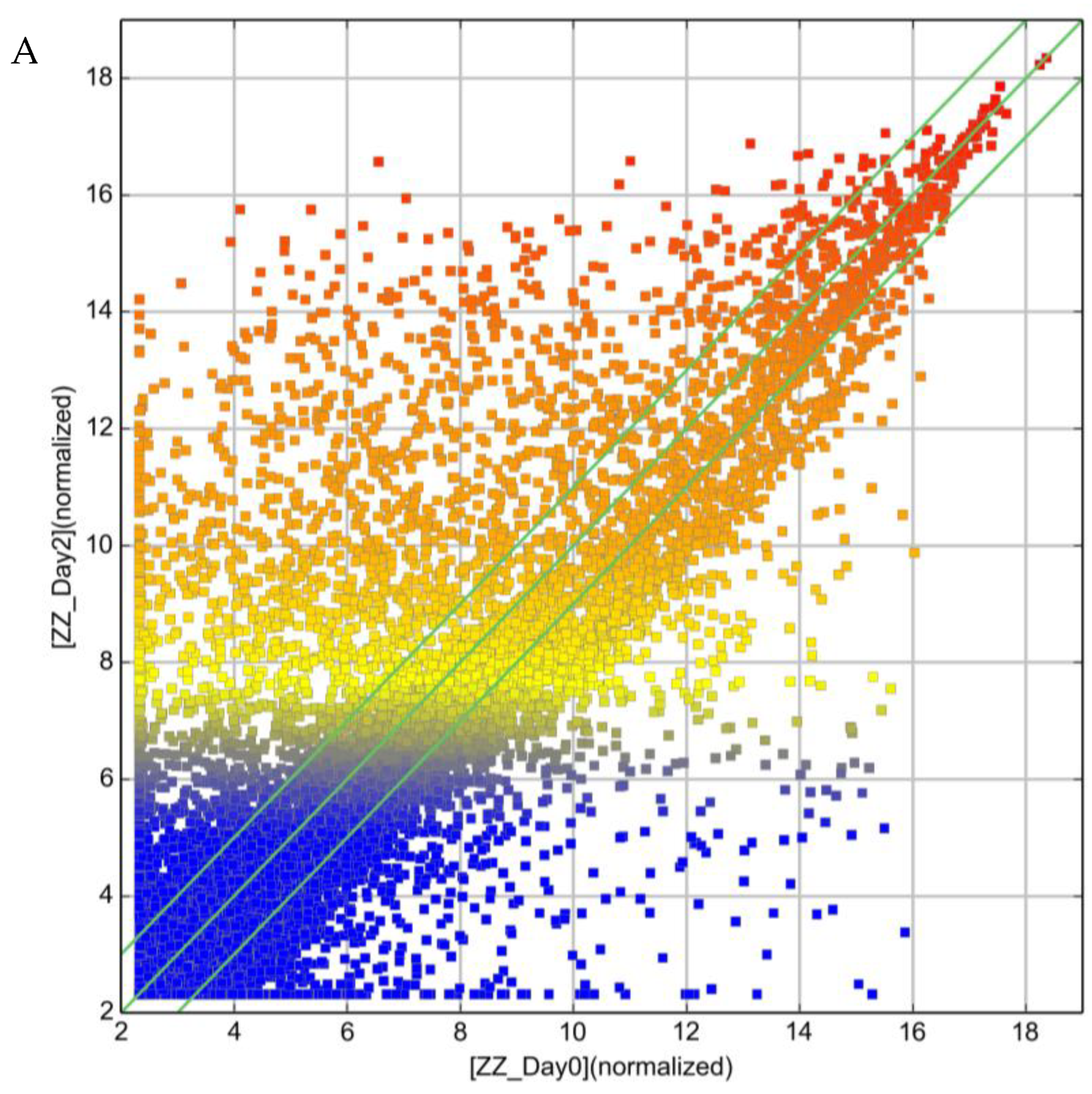
2.1. LncRNA Expression Profile of ZZ Monocytes
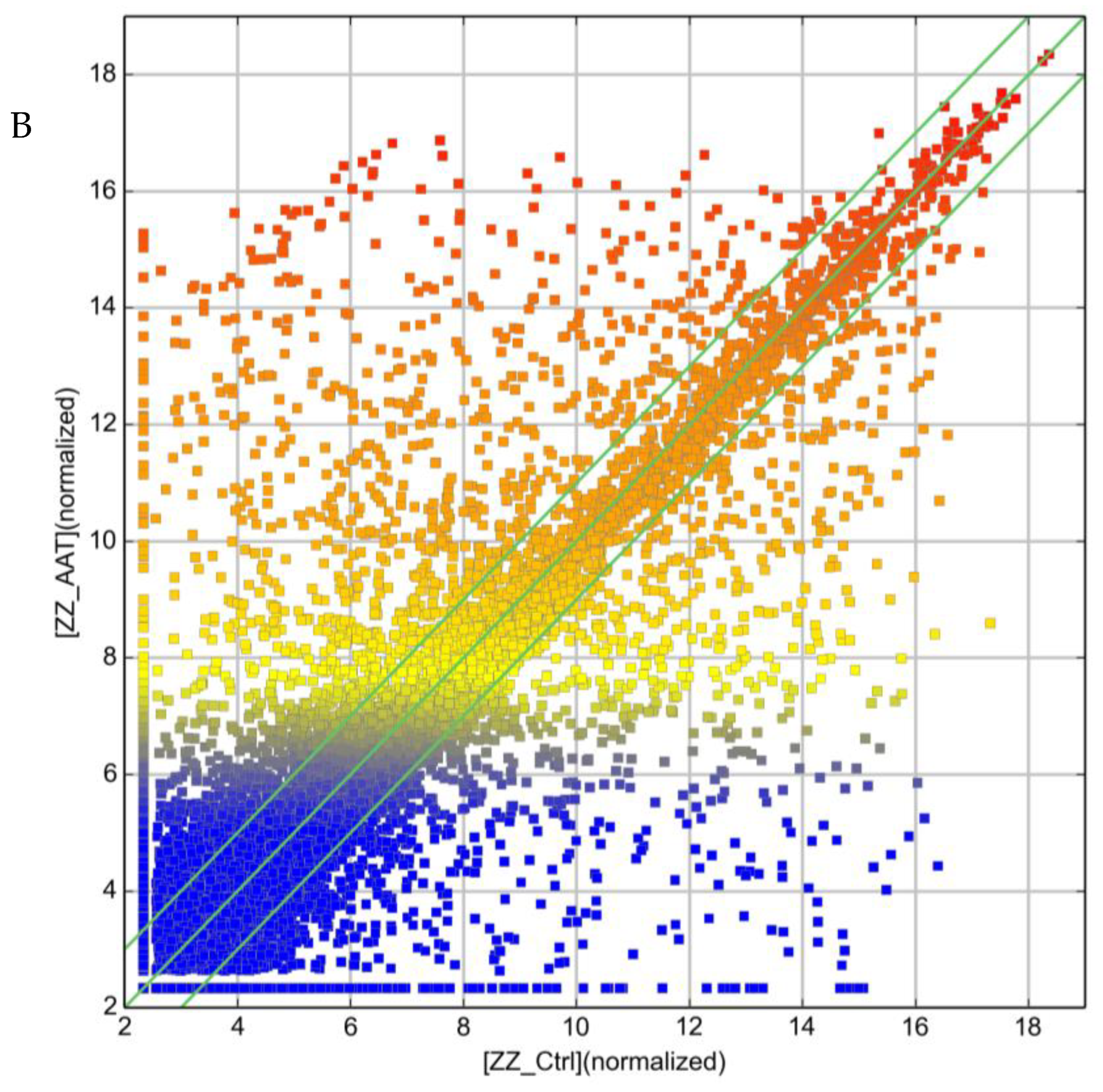
2.2. Effect of AAT Augmentation Therapy on lncRNA Profile
2.3. Ex Vivo Treatment of ZZ Monocytes Alters lncRNA Expression
2.4. Comparison of In Vivo and Ex Vivo Effects of AAT on ZZ Monocytes
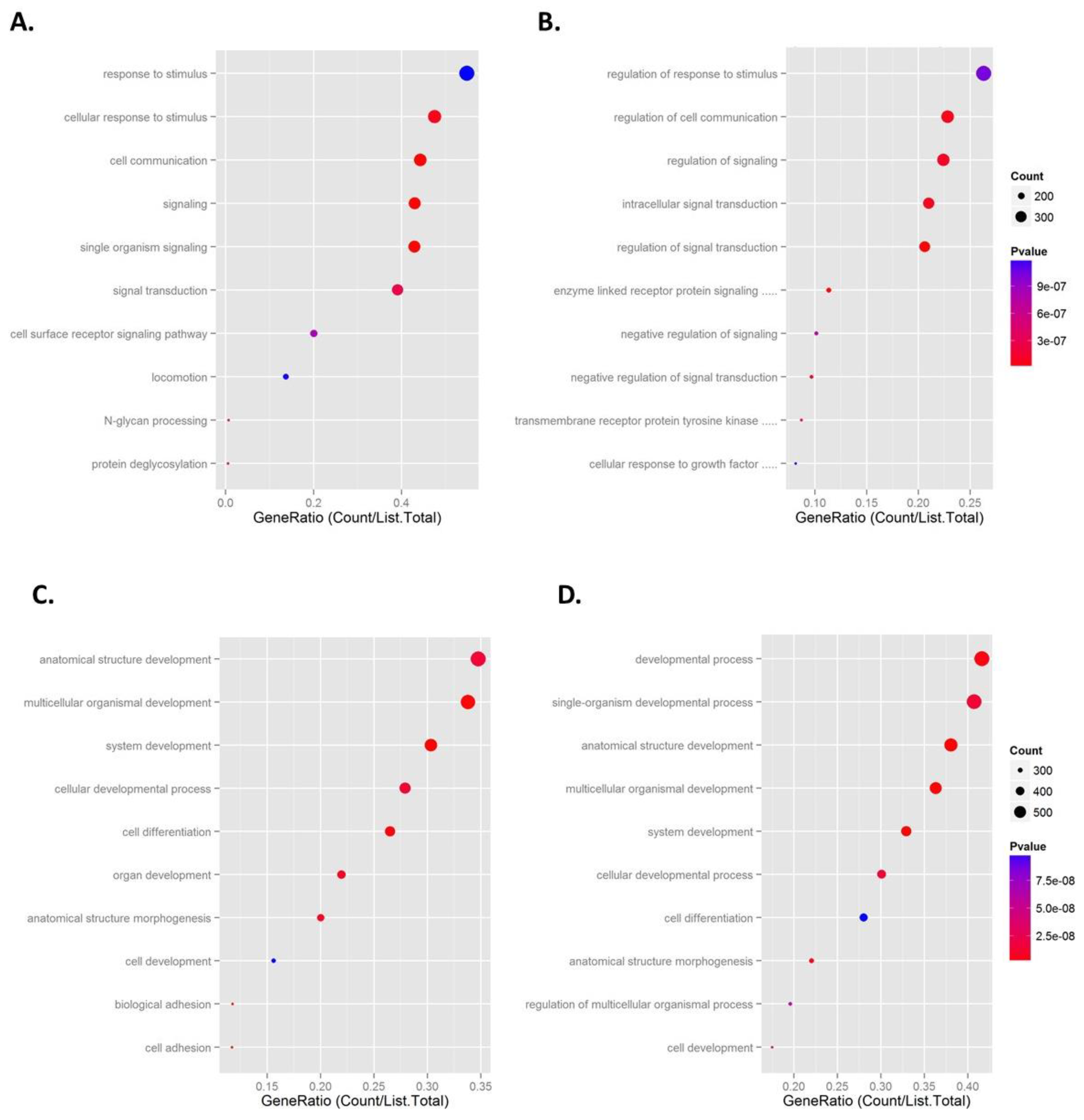
2.5. Pathway Analysis
3. Discussion
4. Methods
4.1. Study Population
4.2. Isolation, Culture and Treatment of Peripheral Blood Monocytes
4.3. Isolation of RNA, RNA Labeling and Array Hybridization
4.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Greene, C.M.; Marciniak, S.J.; Teckman, J.; Ferrarotti, I.; Brantly, M.L.; Lomas, D.A.; Stoller, J.K.; McElvaney, N.G. α1-Antitrypsin deficiency. Nat. Rev. Dis. Prim. 2016, 2, 16051, Erratum in Nat. Rev. Dis. Prim. 2018, 4, 40. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.E.; Murray, G.; Gogoi, D.; Yusuf, A.; McCarthy, C.; Wormald, M.R.; Casey, M.; Gabillard-Lefort, C.; McElvaney, N.G.; Reeves, E.P. A Review of Alpha-1 Antitrypsin Binding Partners for Immune Regulation and Potential Therapeutic Application. Int. J. Mol. Sci. 2022, 23, 2441. [Google Scholar] [CrossRef] [PubMed]
- Laurell, C.B.; Eriksson, S. The electrophoretic α1-globulin pattern of serum in α1-antitrypsin deficiency. 1963. COPD 2013, 10, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Wewers, M.D.; Casolaro, M.A.; Sellers, S.E.; Swayze, S.C.; McPhaul, K.M.; Wittes, J.T.; Crystal, R.G. Replacement therapy for alpha 1-antitrypsin deficiency associated with emphysema. N. Engl. J. Med. 1987, 316, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.G.; Patel, M.; Bayliss, S.; Crossley, D.; Sapey, E.; Turner, A.M. Treatment of lung disease in alpha-1 antitrypsin deficiency: A systematic review. Int. J. Chronic Obstr. Pulm. Dis. 2017, 12, 1295–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, K.R.; Burdon, J.G.; Piitulainen, E.; Sandhaus, R.A.; Seersholm, N.; Stocks, J.M.; Stoel, B.C.; Huang, L.; Yao, Z.; Edelman, J.M.; et al. Intravenous augmentation treatment and lung density in severe alpha1 antitrypsin deficiency (RAPID): A randomised, double-blind, placebo-controlled trial. Lancet 2015, 386, 360–368. [Google Scholar] [CrossRef] [PubMed]
- McElvaney, N.G.; Burdon, J.; Holmes, M.; Glanville, A.; Wark, P.A.; Thompson, P.J.; Hernandez, P.; Chlumsky, J.; Teschler, H.; Ficker, J.H.; et al. Long-term efficacy and safety of alpha1 proteinase inhibitor treatment for emphysema caused by severe alpha1 antitrypsin deficiency: An open-label extension trial (RAPID-OLE). Lancet Respir. Med. 2017, 5, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P.; McEnery, T.; McQuillan, K.; McElvaney, O.F.; McElvaney, O.J.; Landers, S.; Coleman, O.; Bussayajirapong, A.; Hawkins, P.; Henry, M.; et al. α1 Antitrypsin therapy modulates the neutrophil membrane proteome and secretome. Eur. Respir. J. 2020, 55, 1901678. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.E.; Fee, L.; Browne, N.; Carroll, T.P.; Meleady, P.; Henry, M.; McQuillan, K.; Murphy, M.P.; Logan, M.; McCarthy, C.; et al. Activation of complement component 3 is associated with airways disease and pulmonary emphysema in alpha-1 antitrypsin deficiency. Thorax 2020, 75, 321–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEnery, T.; White, M.M.; Gogoi, D.; Coleman, O.; Bergin, D.; Jundi, B.; Flannery, R.; Alsaif, F.A.T.; Landers, S.A.; Casey, M.; et al. Alpha-1 Antitrypsin Therapy Modifies Neutrophil Adhesion in Patients with Obstructive Lung Disease. Am. J. Respir. Cell Mol. Biol. 2022, 67, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Hassan, T.; Smith, S.G.; Gaughan, K.; Oglesby, I.K.; O’Neill, S.; McElvaney, N.G.; Greene, C.M. Isolation and identification of cell-specific microRNAs targeting a messenger RNA using a biotinylated anti-sense oligonucleotide capture affinity technique. Nucleic Acids Res. 2013, 41, e71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, G.L.; Sun, L.Q.; Sun, M.H.; Xu, H.M. LncRNA SLC8A1-AS1 protects against myocardial damage through activation of cGMP-PKG signaling pathway by inhibiting SLC8A1 in mice models of myocardial infarction. J. Cell. Physiol. 2019, 234, 9019–9032. [Google Scholar] [CrossRef] [PubMed]
- Neubert, P.; Homann, A.; Wendelborn, D.; Bär, A.L.; Krampert, L.; Trum, M.; Schröder, A.; Ebner, S.; Weichselbaum, A.; Schatz, V.; et al. NCX1 represents an ionic Na+ sensing mechanism in macrophages. PLoS Biol. 2020, 18, e3000722. [Google Scholar] [CrossRef]
- Molloy, K.; Hersh, C.P.; Morris, V.B.; Carroll, T.P.; O’Connor, C.A.; Lasky-Su, J.A.; Greene, C.M.; O’Neill, S.J.; Silverman, E.K.; McElvaney, N.G. Clarification of the risk of chronic obstructive pulmonary disease in α1-antitrypsin deficiency PiMZ heterozygotes. Am. J. Respir. Crit. Care Med. 2014, 189, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Hassan, T.; de Santi, C.; Mooney, C.; McElvaney, N.G.; Greene, C.M. Alpha-1 antitrypsin augmentation therapy decreases miR-199a-5p, miR-598 and miR-320a expression in monocytes via inhibition of NFκB. Sci. Rep. 2017, 7, 13803. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe Name | Fold Change | Log2 FC | Up-/Downregulated | Sequence Name | Gene Symbol |
|---|---|---|---|---|---|
| ASHGA5P037206 | 3809.9 | 11.9 | Up | ENST00000430815 | C21orf91-OT1 |
| ASHGA5P031008 | 3229.1 | 11.7 | Up | ENST00000448643 | RP11-479J7.1 |
| ASHGA5P056270 | 3169.5 | 11.6 | Up | TCONS_00001566 | XLOC_000905 |
| ASHGA5P034590 | 2695.8 | 11.4 | Up | ENST00000452073 | AC092295.5 |
| ASHGA5P015625 | 2468.78 | 11.3 | Up | NR_051994 | DIAPH3-AS1 |
| ASHGA5P047974 | 2205.78 | 11.1 | Up | NR_049725 | NAV2-AS5 |
| ASHGA5P058933 | 2088.8 | 11.03 | Up | uc.92+ | uc.92 |
| ASHGA5P055648 | 2051.5 | 11.0 | Up | ENST00000417460 | AC003986.7 |
| ASHGA5P029317 | 1351.3 | 10.4 | Up | TCONS_00010873 | XLOC_004700 |
| ASHGA5P035595 | 1191.5 | 10.2 | Up | uc001bgg.1 | AK025975 |
| ASHGA5P030089 | −7931.1 | −12.95 | Down | ENST00000559861 | RP11-265M18.2 |
| ASHGA5P036057 | −5954.6 | −12.93 | Down | TCONS_00026835 | XLOC_013154 |
| ASHGA5P039386 | −5670.7 | −12.5 | Down | ENST00000509449 | RP11-360F5.1 |
| ASHGA5P017396 | −1561.1 | −10.6 | Down | ENST00000441018 | KCNMB2-IT1 |
| ASHGA5P023626 | −1283.9 | −10.3 | Down | HMlincRNA1110+ | HMlincRNA1110 |
| ASHGA5P031124 | −1036.6 | −10.0 | Down | ENST00000569199 | RP11-1437A8.6 |
| ASHGA5P037608 | −933.0 | −9.86 | Down | ENST00000430707 | AP000347.4 |
| ASHGA5P055474 | −902.4 | −9.82 | Down | TCONS_00015528 | XLOC_007235 |
| ASHGA5P044777 | −898.4 | −9.81 | Down | ENST00000518535 | RP11-723D22.2 |
| ASHGA5P021038 | −836.7 | −9.7 | Down | ENST00000523781 | CTB-17P3.4 |
| Probe Name | Fold Change | Log2 FC | Up-/Downregulated | Sequence Name | Gene Symbol |
|---|---|---|---|---|---|
| ASHGA5P022322 | 7893.8 | 12.95 | Up | ENST00000554529 | RP11-14J7.6 |
| ASHGA5P042735 | 7344.9 | 12.8 | Up | ENST00000439207 | RP11-568A7.2 |
| ASHGA5P011052 | 6757.5 | 12.72 | Up | ENST00000422819 | CCDC162P |
| ASHGA5P029573 | 6669.8 | 12.70 | Up | ENST00000557322 | CTD-2002H8.2 |
| ASHGA5P057456 | 4693.6 | 12.2 | Up | TCONS_00017630 | XLOC_008282 |
| ASHGA5P035826 | 4089.4 | 12.0 | Up | ENST00000439606 | AC007254.3 |
| ASHGA5P035237 | 3294.4 | 11.7 | Up | ENST00000453478 | AC068491.1 |
| ASHGA5P027333 | 2953.9 | 11.6 | Up | TCONS_00017353 | XLOC_008198 |
| ASHGA5P033331 | 2315.7 | 11.18 | Up | ENST00000433856 | AC003051.1 |
| ASHGA5P058916 | 2272.5 | 11.15 | Up | uc.268+ | uc.268 |
| ASHGA5P056269 | −6780.7 | −12.7 | Down | TCONS_00001565 | XLOC_000905 |
| ASHGA5P030398 | −6063.4 | −12.6 | Down | TCONS_00009189 | XLOC_004205 |
| ASHGA5P032563 | −5563.0 | −12.4 | Down | ENST00000576014 | RP11-166P13.4 |
| ASHGA5P032930 | −5105.5 | −12.3 | Down | ENST00000580225 | AC004702.2 |
| ASHGA5P045942 | −3974.7 | −11.96 | Down | ENST00000433572 | RP11-542K23.7 |
| ASHGA5P050627 | −3961.0 | −11.95 | Down | uc002tjp.2 | DKFZp686E10196 |
| ASHGA5P030018 | −3461.2 | −11.8 | Down | NR_026891 | FLJ10038 |
| ASHGA5P027135 | −2799.5 | −11.5 | Down | NR_038227 | LOC100506451 |
| ASHGA5P058034 | −2775.1 | −11.4 | Down | TCONS_00025566 | XLOC_012393 |
| ASHGA5P051790 | −2246.8 | −11.1 | Down | ENST00000461063 | SOX2-OT |
| Sequence Name | Log2 FC In Vivo | Log2 FC Ex Vivo | Features |
|---|---|---|---|
| ENST00000452073 | 11.3964871 | 7.6158142 | c/s 19, natural antisense |
| NR_049725 | 11.1070238 | 6.3263702 | c/s 11, intronic antisense |
| uc.92+ | 11.0284561 | 1.7144114 | c/s 2, intronic antisense |
| Sequence Name | Log2 FC Ex Vivo | Log2 FC In Vivo | Features |
|---|---|---|---|
| ENST00000554529 | 12.9464952 | 8.2979554 | c/s 14, intronic antisense |
| ENST00000439606 | 11.997679 | 4.0002684 | c/s 2, intergenic |
| ENST00000453478 | 11.6858148 | 7.375152 | c/s 2, intergenic |
| TCONS_00017353 | 11.5284172 | 4.525568 | X c/s, intergenic |
| uc.268+ | 11.150079 | 6.470752 | c/s 9, intronic antisense |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, S.G.J.; Greene, C.M. Long Non-Coding RNA Expression in Alpha-1 Antitrypsin Deficient Monocytes Pre- and Post-AAT Augmentation Therapy. Non-Coding RNA 2023, 9, 6. https://doi.org/10.3390/ncrna9010006
Smith SGJ, Greene CM. Long Non-Coding RNA Expression in Alpha-1 Antitrypsin Deficient Monocytes Pre- and Post-AAT Augmentation Therapy. Non-Coding RNA. 2023; 9(1):6. https://doi.org/10.3390/ncrna9010006
Chicago/Turabian StyleSmith, Stephen G. J., and Catherine M. Greene. 2023. "Long Non-Coding RNA Expression in Alpha-1 Antitrypsin Deficient Monocytes Pre- and Post-AAT Augmentation Therapy" Non-Coding RNA 9, no. 1: 6. https://doi.org/10.3390/ncrna9010006
APA StyleSmith, S. G. J., & Greene, C. M. (2023). Long Non-Coding RNA Expression in Alpha-1 Antitrypsin Deficient Monocytes Pre- and Post-AAT Augmentation Therapy. Non-Coding RNA, 9(1), 6. https://doi.org/10.3390/ncrna9010006