1. Introduction:
The
KALRN gene is located on chromosome three of the human genome and gives rise to the Dbl family Guanine nucleotide exchange factor protein called Kalirin. It was originally identified as P-CIP10 in a screen for protein-protein interactions [
1] and subsequently named Kalirin [
2]. Later, it was also described as an interacting partner of Huntingtin-associated protein1 and called HAPIP [
3]. It is expressed selectively in the brain and central nervous system, while highly similar proteins like DUO and TRIO show expression in other tissues too [
4]. Kalirin protein consists of many protein-protein interaction domains that allow it to interact with several partners in the dendritic spine. Most of its functions are dependent on the RAC and Rho GTPase functions, which result in the release of GDP from RAC and Rho and activation of GTP signaling in the cell. For instance, the Kalirin-7 (Kal7) isoform localizes to the dendritic spines and interacts with PDZ domain-containing proteins using a specific region in its C-terminal to regulate dendritic morphogenesis [
5]. Kalirin protein isoforms are known to affect the formation of dendritic spines, dendritic arborization, axonal growth, synapse formation, and plasticity [
4,
6,
7].
Mutations in the
KALRN gene that are deleterious to protein function are associated with several neurodevelopmental diseases like autism and intellectual disability, as well as neuropsychiatric diseases like schizophrenia [
8,
9,
10,
11,
12]. Intronic variants that do not change the protein product but result in altered expression have been linked to addiction and reward anticipation [
13].
KALRN expression was also found to be reduced in post-mortem Alzheimer’s brain samples. Overall, the disease phenotypes seen in humans agree well with the loss of expression or function of Kalirin protein and resultant behavioral effects seen in mouse models. An interesting complication is the disparate effects of the Rac and Rho GEF domains: mutations in the Rac GEF domain reduce RAC signaling, resulting in reduced neuronal branching and spine density, and lead to a decrease in cortical thickness, while a mutation in the Rho GEF domain had similar effects in spite of activating Rho GTP signaling [
7,
14].
The multi-function Kalirin protein exerts its diverse effects through its many isoforms. The locus consists of 60 exons in humans, which give rise to 52 transcript isoforms. The regulated expression of
Kalrn isoforms is thought to be driven by differential promoter usage, although these promoters are yet to be characterized in detail. The transcript isoforms can be clustered into two major groups: One group of relatively short isoforms, mostly including the exons at the 5′ end, includes the Kal7 isoform that has been studied extensively in mice and cell culture models. Kal7 knockout also resulted in schizophrenia-like effects in adolescent mice [
15]. The Kal7 isoform includes the RacGEF domain but not the RhoGEF domain. Restoring levels of Kal7 can reverse the Alzheimer’s-like effects shown by mice with reduced Kal7 [
16,
17]. Besides the catalytic RacGEF domain that is critical to its function, it also contains a C-terminal PDZ domain, which is thought to target it to post-synaptic regions, and this facilitates its interactions with other proteins in this subcellular compartment. The longer isoforms, which include the RhoGEF domain, form a distinct group comprising Kalirin-9 (Kal9) and Kalirin-12 (Kal12), which play important roles during neuronal development [
18,
19,
20]. Although it is clear from mutations linked to diseases and phenotypes in mouse models that the correct spatio-temporal expression of Kalirin isoforms is as critical as the final protein product itself, there is relatively little information on the regulation of the genomic locus.
Long non-coding RNAs are RNA transcripts more than 200 nts in length that do not code for functional proteins but may exert a variety of regulatory roles ranging from miRNA sponging and clearance to providing scaffolds for assembling ribo-nucleoprotein complexes and tethers for chromatin modifiers [
21,
22]. In zebrafish, we have reported that a maternally inherited long non-coding RNA, named durga, is the transcript expressed earliest in the developmental time frame, from the
Kalrn locus. The non-coding RNA partially overlaps with the first exon of protein-coding
Kalrn isoforms and is expressed during an early developmental window and later in certain regions of the adult zebrafish brain [
23].
To identify similar non-coding RNAs from the mammalian Kalrn locus, we carefully annotated the transcripts reported in transcriptomic studies. By combining data from genome and transcriptome data repositories, we built a detailed map of the non-coding transcripts arising from the mouse Kalrn locus, including seven linear and one circular RNA. The linear RNAs are henceforth called Kalnc1 to 7 in the order of their positions from the 5′ to 3′ ends of the mouse Kalrn gene. Kalnc2 arises from the 5′ end close to the three known alternative start sites of Kalrn mRNA. During the in vitro differentiation of mouse primary cortical neurons, Kalnc2 was down-regulated in mature neurons. We used RNA from post-mortem human tissues and a human neuronal cell line to show that the human KALRN locus also gives rise to a potentially non-coding transcript arising from the 5′ end of the KALRN gene, which shares short stretches of sequence similarity and functional features like chromatin association with Kalnc2 and shows selective enrichment in immature neurons. In a mouse neuronal cell line, Kalnc2, overexpressed, leads to the downregulation of all major protein-coding transcripts of the Kalrn gene.
2. Results
The full-length mRNA transcript (15.4 kb) arising from the mouse
Kalrn gene consists of 60 exons spread over 604.4 kb of the genome (
Figure 1) and matches the complexity of the human
Kalrn gene. The start codon occurs within the first exon, and the stop codon of the largest isoform occurs in the 60th exon. Alternative splicing of these protein-coding transcripts gives rise to about 14 coding variants, of which Kal7, Kal9, and Kal12 have been characterized in relatively greater detail. Besides these coding transcripts, we found seven presumably non-coding transcripts, referred to here as
Kalnc1 to
Kalnc7, all expressed in the same direction as the protein-coding transcripts. The protein-coding transcripts of the
Kalrn gene are known to have at least four alternate start sites, often referred to as A, B, C, and D. Some of these transcripts comprise many exons and are several kilobases in length. For instance,
Kalnc1 consists of 37 exons collectively account for 6.1 kb in length after removal of introns. Although it overlaps significantly with the protein-coding transcripts (35/37 exons), the use of distinct exons at the termini, avoiding the canonical start codon, results in a low coding potential. Some other ncRNAs from the locus were as short as 373 nts, comprising only 2 exons. We next checked their coding potential and reliability of the annotation (
Figure 1A). For coding potential analysis, we used the CPAT algorithm, which recommends a score of less than 0.44 for non-coding transcripts.
Kalnc2 and
Kalnc5 showed low coding potential scores of 0.15 and 0.09, respectively, whereas the other ncRNAs arising from the
Kalrn locus have scores ranging from 0.75 to 1.00, perhaps attributable to the overlap with coding exons of the
Kalrn gene. Four of the linear ncRNAs,
Kalnc1,
Kalnc2,
Kalnc3, and
Kalnc6, are bioinformatically assembled transcripts, while
Kalnc4 has a relatively better level of confidence, with every splice junction being supported by at least one transcript. Interestingly, we found that a 294 nts circular RNA, apparently formed by the back splicing of exons 53 and 54, has also been detected in high-throughput sequencing but has not been verified further [
24,
25]. Taken together, bioinformatics predictions suggested that the
Kalrn gene locus gives rise to seven linear and one circular RNA (
Figure 1B), calling for a thorough experimental validation that is essential for studying the regulatory potential of this region.
Using total RNA isolated from the brain of adult mice and primers designed to differentiate between isoforms, we carried out RT-PCR to check the presence of these novel transcripts of unknown function. We also included a pair of pan-
Kalrn primers that would detect the coding transcripts. As expected, the pan-
Kalrn primers support the notion that
Kalrn is expressed across tissues, and
Kalnc1, the largest non-coding isoform, was also similar (
Figure 2A). Even with this single pair of primers, it was obvious that
Kalnc1 shows further splicing heterogeneity.
Kalnc6, a relatively small ncRNA, was also expressed in all the tissues tested. The predicted circular RNA [
24,
25] was detected using divergent PCR primers (
Figure 2B). The size of the observed band was 217 nts, slightly smaller than the predicted 294 nts. It was readily detected in the cerebrum, cerebellum, spinal cord, and ovary of adult mice. During development, it was detected only in the hippocampus at postnatal day 1.
Our group has previously shown that in zebrafish, a lncRNA arising from the
kalrna gene locus is maternally inherited [
23]. To test if the mammalian locus also had potentially inherited lncRNAs, we included RNA from the mouse ovary in our RT-PCR experiments. All the ncRNAs barring
Kalnc7 showed expression in the ovary besides being expressed in one or more parts of the central nervous system.
Kalnc3 and
Kalnc4 were particularly interesting because they were specifically expressed in the cerebrum and cerebellum, respectively.
Since these transcripts overlap with coding transcripts and each other, in situ hybridization using larger probes cannot be used to resolve the spatial expression pattern in greater detail. To look at the brain expression pattern in greater detail, we carried out quantitative RT-PCR on RNA from the cortex and hippocampus in mice of postnatal day 1,3, 5 and 18 followed by 3 weeks, 1 month and 4 months. As shown in
Figure 2B,
Kalnc4 was readily detectable at all developmental stages in the cortex and hippocampus. This level of abundance is higher than expected for lncRNAs since it roughly correlates to one or two orders of magnitude less than the abundant
Gapdh mRNA. All the other lncRNAs were far less abundant, but a general trend in the expression pattern was that all the lncRNAs from this locus were least abundant at P3 and P5 (postnatal days 3 and 5), a period that coincides with the switch from neurogenesis to astrogenesis (
Figure 2C). During the later stages, when synapses mature, the expression of the
Kalrn locus lncRNAs is again relatively high.
Further, we analyzed 209 publicly available RNA-Seq datasets pertaining to the disease conditions that are already linked to the
Kalrn locus (addiction-17, autism-120, schizophrenia-38, and epilepsy-34). The datasets were internally normalized, and fold changes were calculated with respect to corresponding control samples (see methods for details). We found
Kalnc4 is upregulated in several autism-related datasets, while it is downregulated in mice addicted to nicotine (
Figure 2D). The largest lncRNA,
Kalnc1, was upregulated in a number of conditions. Interestingly,
Kalnc2 resembled protein-coding isoforms of Kalirin in that it was downregulated in the nucleus accumbens of mice addicted to nicotine. Thus, lncRNAs arising from the
Kalrn locus have specific developmental expression profiles and are differentially expressed in disease models.
Kalnc2 was of particular interest because of its location of origin and its temporal expression pattern. As shown in
Figure 1B,
Kalnc2 is a relatively small 523-nt lncRNA comprising 2 exons with a very low coding potential. Its location coincides with the 5′ end of the
Kalrn protein coding isoforms, similar to the zebrafish durga lncRNA that we have previously identified. To understand the role of this lncRNA in greater detail, we studied its sub-cellular localization in primary cortical neurons in culture at the time of plating and at 15 days of culture (
Figure 3A,B). As is the case with zebrafish
durga, it was largely present in the nucleus. To resolve the intranuclear localization in further detail, we used the rodent neuroblastoma cell line Neuro2A. Here too,
Kalnc2 was localized to the nucleus, and further, it was enriched in the chromatin fraction (
Figure 3C,D).
Malat1 and
Gapdh showed the expected nuclear and cytoplasmic localization, respectively, ruling out the mixing of fractions during the experiment.
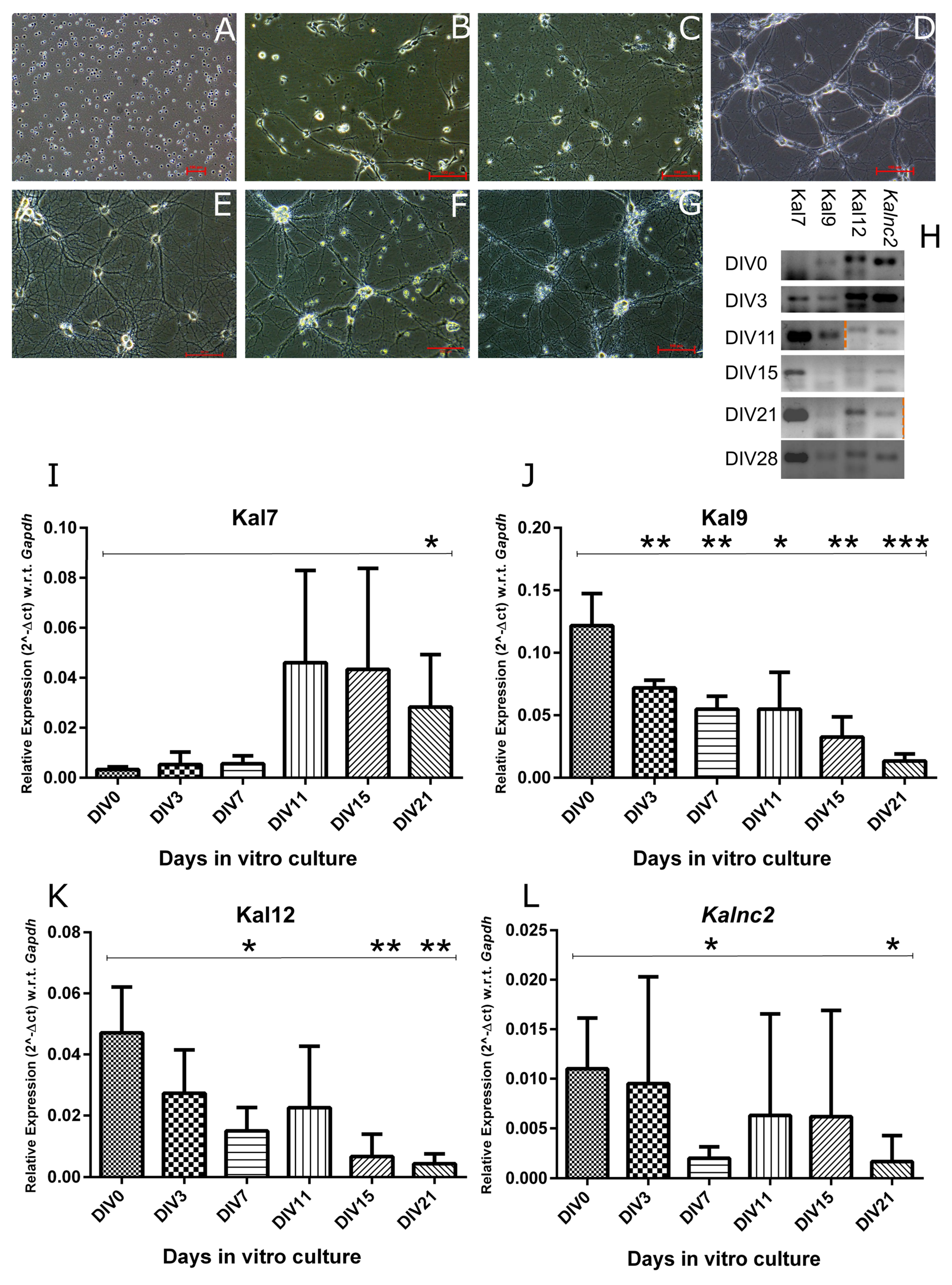
Next, we characterized the expression profiles of
Kalnc2 and the major
Kalrn mRNA isoforms, Kal7, 9, and 12, during in vitro differentiation of cortical neurons (
Figure 4). In agreement with previous reports, Kal9 and Kal12 are expressed during the early proliferative stages, but as neurons start forming synapses and these synapses mature, their expression is reduced. In sharp contrast, the expression of the Kal7 isoform, rises only in the later stages. In vivo, it has been shown that Kal9 and Kal12 dominate during embryonic development, while Kal7 is largely found in the adult brain.
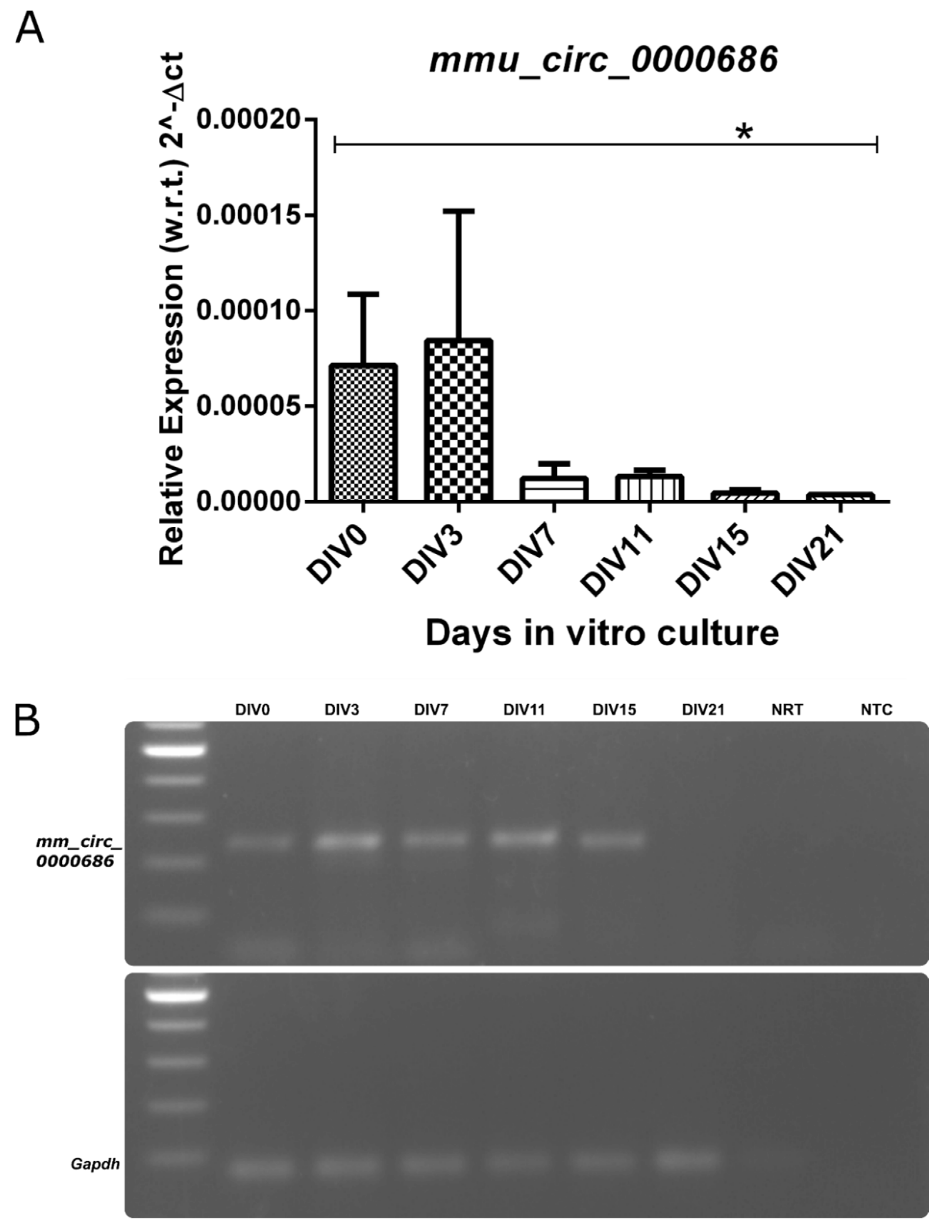
Kalnc2 showed a strong similarity in expression pattern to Kal12, which is essential for axonal outgrowth. Like Kal12, it was expressed most strongly in the early stages of culture and decreased gradually with the formation of the neuronal network. The circular RNA arising from a back splicing of exon 54 and exon 53 of the
Kalrn gene corresponding to Kal9/12 isoforms was also expressed highly in the initial stages but decreased during differentiation until it was undetectable at 21 days in vitro (
Figure 5).
In parallel, we explored the role of a lncRNA arising from a comparable location in the
KALRN locus of the human genome. The human
KALRN locus is comparable to the mouse locus in complexity, with 29 ncRNAs arising from the 692 kb locus. We found a transcript (ENST00000684441.1/ENST00000488825.5) (
supplementary data Figure S1) that was annotated in the direction of the coding mRNAs at the 5′ end—a position corresponding to the
durga lncRNA in zebrafish and
Kalnc2 in mouse. Since the first exon of this transcript showed sequence conservation across vertebrates, this lncRNA is henceforth referred to as
hsKALNC2 (
supplementary data Figure S2). We used directional reverse transcriptase PCR to validate the expression and confirm the orientation of this lncRNA. In agreement with the observation in mice, this RNA is also expressed in the cerebrum and ovary, but is excluded from tissues like cerebellum, liver, and heart where
Kalrn mRNA is readily detected (
Figure 6A). In human tissues and in the cell line SH-SY5Y, the lncRNA was of much lower abundance when compared to
KALRN mRNA transcripts (
Figure 6A). The
hsKALNC2 transcript was closely associated with the chromatin fraction (
Figure 6B) in SH-SY5Y cells where it was expressed along with
KALRN mRNA (
Figure 6C). Taken together, we propose that mammalian genomes express a syntenically conserved lncRNA from the 5′ end of the
Kalrn protein-coding gene, which is nuclear localized and selectively enriched in specific brain and ovary regions. The
mmKalnc2 and
hsKALNC2 genes share an identical 200-nts stretch at the 5′ end suggesting that mutational studies in the future should focus on this region. Owing to their conservation in mammals and relative synteny with respect to zebrafish
durga lncRNA, we reasoned that they may be involved in similar functions.
To test for a direct regulatory role of lncRNA in the expression of
Kalrn mRNAs, we modulated the expression level of
mmKalnc2. The
mmKalnc2-specific transcript was expressed in sense and antisense orientation (as a control) under a bidirectional promoter such that transfected cells would coexpress a Red Fluorescent Protein (RFP) allowing their visualization. This clone was transfected into Neuro2A cells using lipofectamine (see materials and methods for details), and total RNA was collected 48 h post-transfection. We measured the steady-state expression of mouse Kal7, 9, and 12 using isoform-specific primers [
27] in qRT-PCR. We could change the expression level of the lncRNA by >100 fold, irrespective of the orientation (
Figure 7E). As shown in
Figure 7B,D when
Kalnc2 (sense transcript) was overexpressed, we find that Kal7, Kal9 and Kal12 mRNAs were downregulated by about 50% (
Figure 7F–H).
3. Discussion
Our results reveal that the Kalrn locus gives rise to several linear and one circular ncRNA that are selectively expressed in the adult mouse nervous system. The circular RNA, mmu_circ_0000686, mmKalnc2, and hsKALNC2 from the human Kalrn gene locus are also expressed during the early stages of development. The same pattern is recapitulated during in vitro differentiation, showing a gradual decrease with time.
The circular RNA,
mmu_circ_0000686, and the linear RNAs,
Kalnc2 and
Kalnc5, were clearly non-coding. The other RNAs described here showed some coding potential in our bioinformatics analysis but were exclusively localized to the nucleus. In our opinion, the ambiguity in the coding potential of
Kalnc1, 3, 4,
and 7 is due to the overlap with protein-coding RNAs. The inclusion of introns, which usually contain stop codons, may convert a coding transcript into a non-coding variant.
Kalnc6 showed some coding potential and is at least partially localized to the cytoplasm. Further studies are required to verify the non-coding nature of this transcript. The ultimate evidence for the coding status of a transcript can only be provided by the demonstration of the protein product. Association with ribosomes and the presence of short ORFs are both now known to be only indicators of coding potential [
28]. Further, certain transcripts may perform non-coding RNA functions and also give rise to small peptides or proteins [
29].
Compared to mRNAs, lncRNAs are usually much lower abundance. Among the non-coding RNAs arising from the Kalrn gene locus, only Kalnc4 is expressed at high levels, comparable to the relatively abundant mRNA Gapdh. All the other lncRNAs are expressed at a baseline about two orders of magnitude lesser than Kalnc4. However, these RNAs are transiently induced several folds in the mouse cortex around postnatal day 18 but are barely detectable in the 1-month-old mouse brain. All the non-coding RNAs are again readily detectable in the 4-month-old mouse brain. Since mouse puberty occurs at around 38 days, we speculate that the non-coding RNAs of the Kalrn locus may be induced during puberty, besides a transient spike in expression during development.
The mammalian
Kalrn gene locus has been extensively mined for mutations associated with several neurodevelopmental and neuropsychiatric diseases [
9,
10,
11]. Meta-analysis of gene expression data reported by various groups studying mouse models of these diseases showed that
Kalnc1,
2,
4, and
7 show altered expression in models of autism and nicotine addiction. The protein-coding isoforms arising from the locus have several interaction domains that provide spatiotemporal heterogeneity to the core Rho GTPase role of the Kalirin protein. Many protein-coding isoforms depend on different promoters for their expression, while the relationship between regulatory elements and non-coding RNAs has not been explored. Certain isoforms promote axonal outgrowth, while others determine dendrite numbers, dendritic arborization, and the maturation of dendritic spines. As neurons mature, the expression of the longer protein-coding isoforms, Kal9 and Kal12, decreases with a concomitant increase in Kal7. We used overexpression of
Kalnc2 to explore the possibility that it regulates the expression of protein-coding
Kalrn transcripts. When we ectopically overexpressed it in N2A cells, we found Kal7, Kal9, and Kal12 mRNAs were downregulated. However, it appears that
Kalnc2 may have no effect on the switch from longer to shorter
Kalrn protein-coding isoforms during neuron differentiation and maturation.
There are several caveats to unequivocally establishing the function of Kalnc2. More refined experiments addressing the role of Kalnc2 in specific neuronal subtypes or in specific regions of the brain may reveal a regulatory role for these lncRNAs in vivo. Loss of function studies through knock-down and mutations are needed to establish the function of this lncRNA. We cannot rule out the possibility that localization and RNA modification of endogenous Kalnc2 may not be replicated when overexpressed ectopically.
In spite of the apparent lack of sequence conservation, the lncRNAs
durga (zebrafish),
mmKalnc2, and
hsKALNC2 show striking similarities: they share the same protein-coding neighbor, associate with chromatin, and are expressed in immature neurons. In the future, chromatin immunoprecipitation can reveal the genomic region to which these lncRNAs bind, perhaps in association with chromatin remodeling complexes. They may also be involved in defining 3D chromatin organization. Neuronal differentiation and maturation are accompanied by large-scale changes in 3D chromatin structure. It will be interesting to study the role of lncRNAs, especially those from the
Kalrn locus, that could contribute to such spatial reorganization of chromatin in the nuclei of neurons during differentiation. Regulatory RNAs inherited through the maternal ooplasm are a form of epigenetic inheritance [
30]. Recent studies have shown that gametes are a rich source of regulatory RNAs, including miRNA [
30], tRNA fragments [
31], and lncRNA [
32]. A lncRNA arising from the locus is inherited through the maternal cytoplasm of oocytes in zebrafish [
23]. The expression pattern of the mammalian lncRNAs raises the possibility that they are also inherited, although it is premature to assume ovarian expression corresponds to expression in oocytes since we cannot rule out the possibility that the RNA is localized to the non-gametic tissue of the ovary. In summary, we report that several, hitherto unknown, non-coding RNAs are dynamically expressed from the mammalian
Kalrn locus, and one such RNA,
Kalnc2, can downregulate major
Kalrn protein-coding transcripts, highlighting the regulatory potential of this genomic region that is implicated in several neuropsychiatric and neurodevelopmental disease conditions.
4. Materials and Methods
4.1. Genome and Transcriptome Meta-Analysis
All the analysis and visualization for the mouse genome was carried out on (GRCm38/mm10), and similarly, the human genome (GRCh38/hg38) was used. The
KALRN genomic locus (mouse = chr16: 33,969,073-4,573,532; human = chr3: 124,033,369-124,726,325) was accessed with the tracks from Gencode v.38 (Ensembl 104) or Gencode v.M25 for mouse (Ensembl 100). Treating
Kalrn/ENST00000682506.1 as the longest transcript, all transcripts within its limits were then further analyzed. Five transcripts that were mentioned as “processed transcripts” and two that denoted “intron-retained” were observed. These transcript sequences were downloaded and used for coding potential analysis in the CPAT software [
26] (
http://lilab.research.bcm.edu/) (accessed on 30 June 2022). Oligonucleotide primers (
Supplementary Data Table S1) specific for transcripts and pan-
Kalrn were designed using Primer 3.
4.2. Animals
All experimental procedures were performed on Balb/c mice and approved by the Institutional Animal Ethics Committee of the CSIR-Institute of Genomics and Integrative Biology (IGIB) and followed appropriate guidelines for live animal use in research. All mice used in this study were bred in the CSIR-IGIB animal house facility. They were kept at a temperature of 24 ± 2 °C on a 12 h light/dark cycle, with ad libitum access to food and water. All institutional guidelines were followed for animal handling and experiments.
4.3. Primary Cortical Neuron Culture
Adult male and female mice were kept for mating at a 1:2 ratio, and pregnancy was confirmed by checking the vaginal plug, and after a few days of mating, it was re-confirmed by measuring the weight of a female. Pregnant females were anesthetized with thiopentone (40 mg/kg) at Embryonic days 16–8 (E16-18). The embryos were dissected out by making a straight incision in the abdomen. Embryos were immediately kept in cold HBBS (14025 Gibco, Billings, MT, USA), and brains were removed. Carefully, meninges and blood vessels were removed, and cortices were dissected out. They were transferred into the 15 mL tube containing cold HBSS. Cortices were washed three times with HBSS for 5 min at 500 g and 0.25% trypsin-EDTA (25200-0560, Gibco) was added, then triturated by a Pasteur pipette and kept at 37 °C for 5 min. Trypsin-EDTA was inactivated by (10082-147, Gibco), and tissue was centrifuged at 500× g for 5 min, then washed twice with HBSS, and a single cell suspension was made in complete neurobasal media (Neurobasal; 10888-022, Gibco + B27; A35828-01, Gibco + Glutamax; 35050-061, Gibco + Primocin; ant-pm-1, Invivogen, Waltham, MA, USA). Cells were counted and seeded on the poly-D lysine (P7405, Sigma, St. Louis, MO, USA) coated (0.1 mg/mL solution) plates. The first media change, replacing half of the medium, was done after 12 h of cell seeding, and thereafter twice a week. Cells were collected at different days in vitro culture (DIV): DIV0 (the day when cells were seeded), DIV3, DIV7, DIV11, DIV15, DIV21, and DIV28.
4.4. Neuro2A Cell Culture
N2A cells (originally sourced from the National Cell Repository) were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, Invitrogen) supplemented with 10% fetal bovine serum (FBS) and incubated in a humidified incubator with 5% CO2.
4.5. SH-SY5Y Differentiation
SH-SY5Y cells (source: European Collection of Authenticated Cell Cultures) were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, Invitrogen) supplemented with 10% fetal bovine serum (FBS) and incubated in a humidified incubator with 5% CO2. For differentiation of SH-SY5Y, cells at passage number 30–33 were treated with 10 µm of trans retinoic acid (RA) at day 1, cells were then grown for 4 days with media change at every alternative day and replaced with 10 ng/mL BDNF and 10µm RA at day5 and grown for 10 days. At day 10, DMEM was replaced with neurobasal media, and cells were collected at day 12.
4.6. Kalnc2 over-Expression
A Kalnc2-specific fragment of 321 nts (chr16:34,512,74034,513,074) and reporter mCherry were cloned under the mammalian CAG-bidirectional promoter and transfected by lipofectamine 2000 (116668-019, Invitrogen) into the Neuro2A cells. After transfection, cells were collected at different time points, and the expression of Kalnc2 along with Kalrn protein-coding transcripts was measured by quantitative RT-PCR.
4.7. RNA Isolation and RT-PCR
Mice were euthanized with sodium thiopental (40 mg/kg), and the brain and other body parts were removed. RNA was isolated using trizol (15596018, Ambion, Austin, TX, USA) from the whole brain, cortex, hippocampus, cerebellum, ovary, liver, heart, and primary cortical neurons. cDNA was prepared from 1 µg of total RNA, and then RT-PCR was done using the SYBR Green master mix (PKG025-A Genetix). Gapdh was used as a normalization control in RT-PCR data analysis. The RT-PCR products were visualized by agarose (2%) gel electrophoresis.
4.8. Subcellular Fractionation
Cells were washed with 1×PBS and then collected with 0.25% trypsin-EDTA, centrifuged (100–200×
g for 5 min), and counted. A 175 µL cytoplasmic lysis buffer (tris-HCl pH 8.0 50 mM, NaCl 140 mM, MgCl2 1.5 mM, NP-40 0.5%, DTT 1 mM, EDTA 5 mM) was added to 1 × 10
6 cells, suspended, and incubated at ice for 5 min, and then cell suspension was centrifuged at 300×
g for mins at 4 °C. The supernatant was transferred into the fresh tube, and it is a cytoplasmic fraction. The pellet was washed twice with 200 µL of cytoplasmic lysis buffer at 300 g for 2 min at 4 °C. 175 µL nuclear lysis buffer (Tris HCl pH 8.0, 50 mM, NaCl 500 mM, MgCl2 1.5 mM, NP-40 0.5%, and DTT 1 mM) were added per 1 × 10
6 cells pellet, suspended, and incubated at ice for 5 min. The cell suspension was then centrifuged at 16,400×
g for 2 min at 4 °C. The supernatant was collected which constituted soluble nuclear fraction, and the pellet corresponding to the chromatin fraction, were separated, and the RNA isolation, cDNA preparation, and RT-PCR were done for all three fractions. The RT-PCR products were visualized by agarose (2%) gel electrophoresis. Subcellular fractions were confirmed by respective fraction markers:
Gapdh and
RNA18S (cytoplasmic markers),
Malat1 (nuclear markers), and
RNA45S (chromatin markers), since 45S rRNA is chromosome-associated [
33].
4.9. Meta-Analysis of Expression Datasets
The 209 datasets associated with the query (addiction OR autism OR epilepsy OR schizophrenia) AND “Mus musculus” [porgn] AND (“Expression profiling by high-throughput sequencing” [Filter]) were downloaded from Gene Expression Omnibus (GEO). Raw SRA sample files were converted into fastq format by fastq-dump [
34], adapter trimming was performed by trimmometic [
34], and finally, quality control was done by fastqc [
35] software. Passed quality control samples were aligned on mouse assembly (GRCm38/mm10|: Chr16) by STAR [
36], reads were counted by RSEM [
37], and differential expression and fold change were calculated by EBSeq [
36].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








