
T2DB: A Web Database for Long Non-Coding RNA Genes in Type II Diabetes


 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
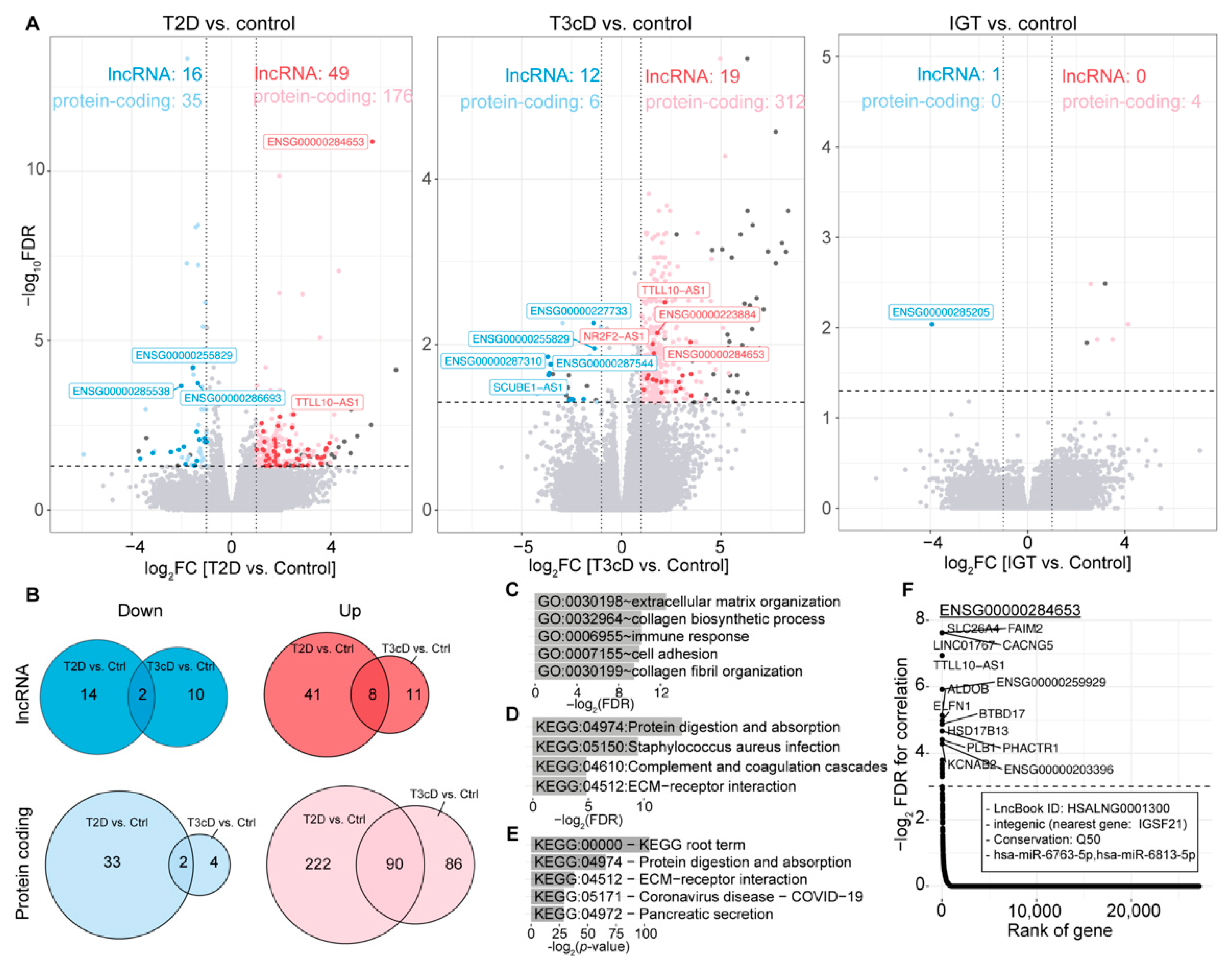
2.1. Many lncRNAs Are Differentially Expressed in T2D Patients
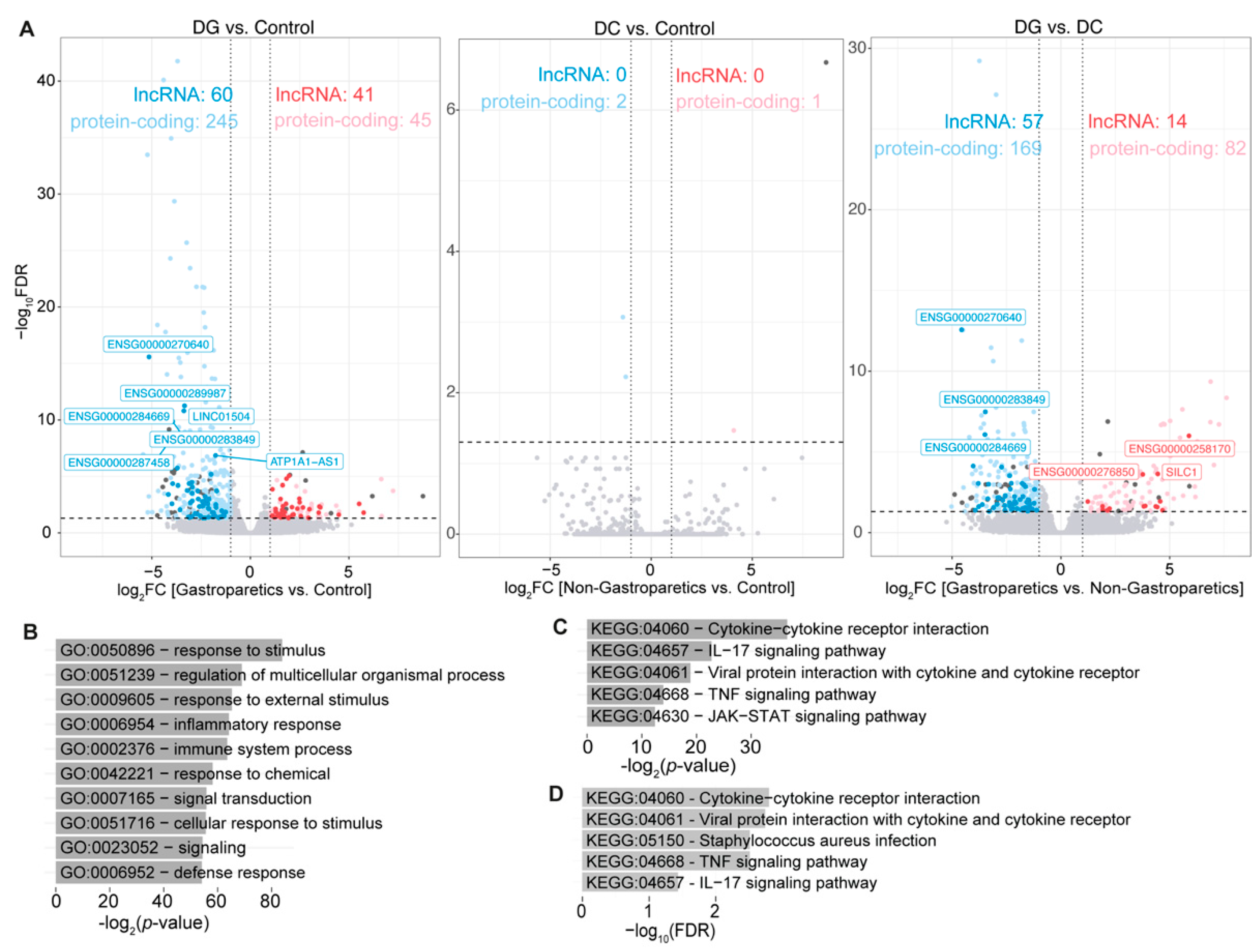
2.2. Diabetic Gastroparesis Results in Significant Down-Regulation in Gene Expressions
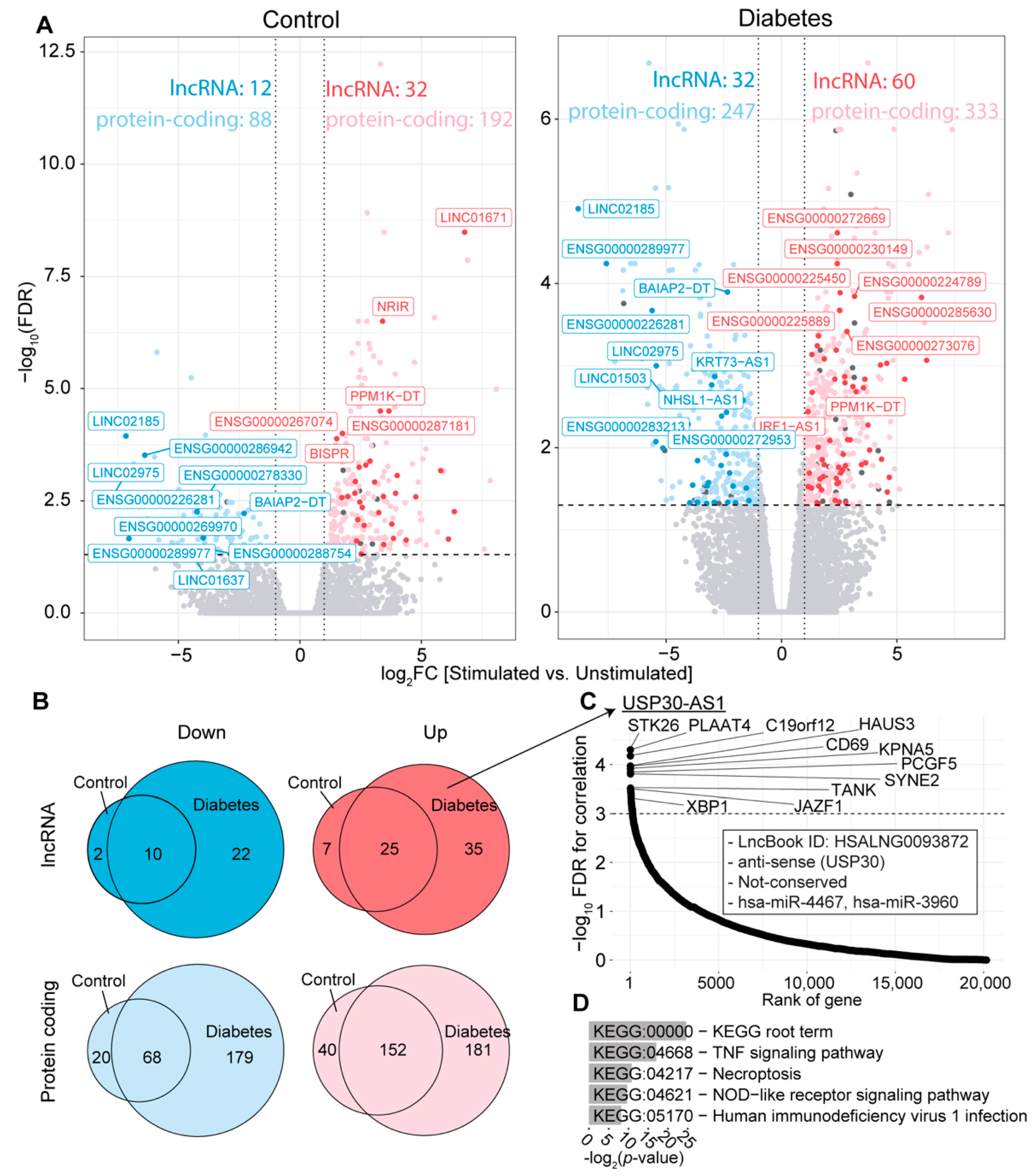
2.3. Many lncRNAs Are Differentially Expressed in Cultured Macrophages from T2D Patients
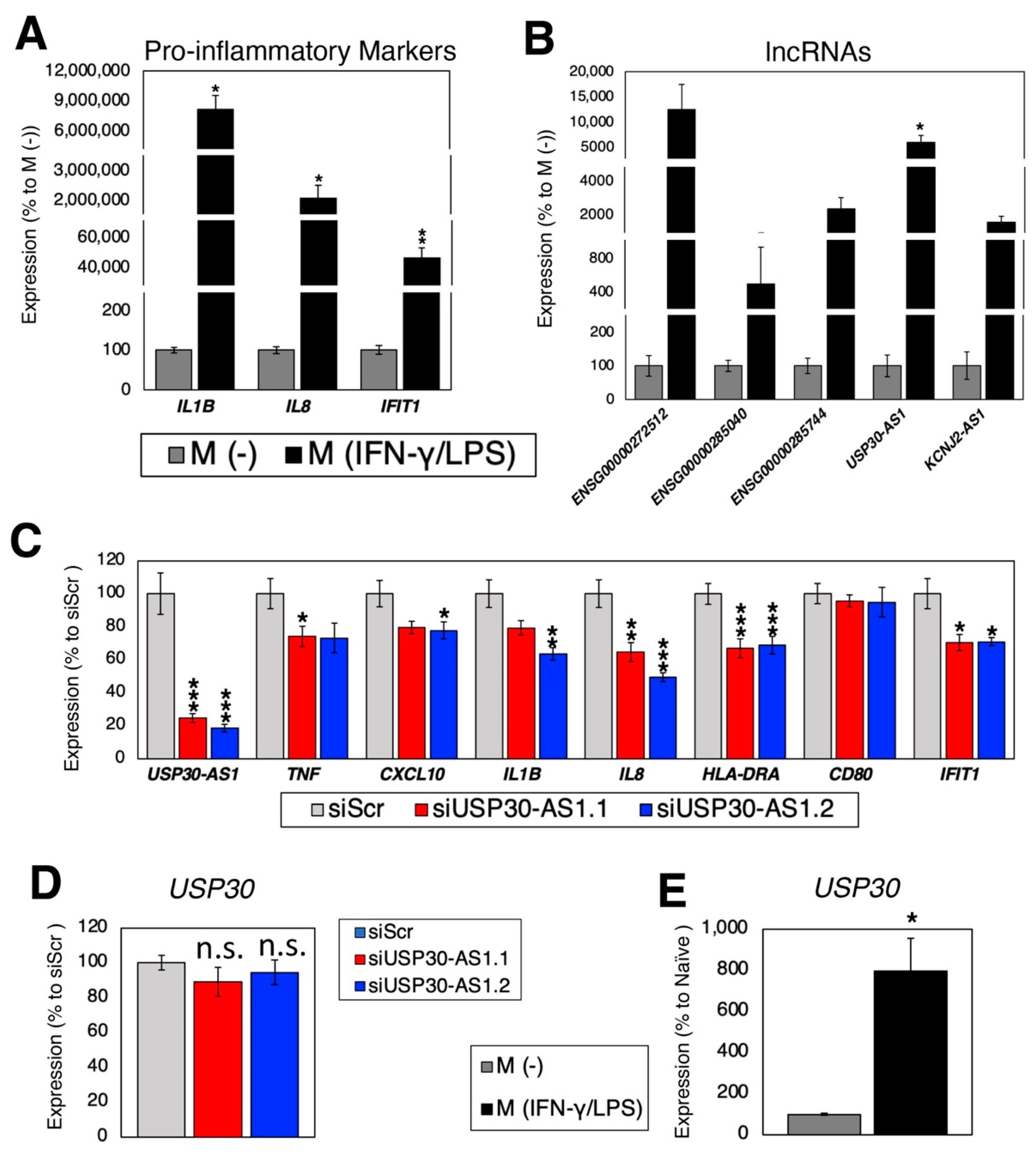
2.4. Loss-of-Function Experiments in Polarized Macrophages
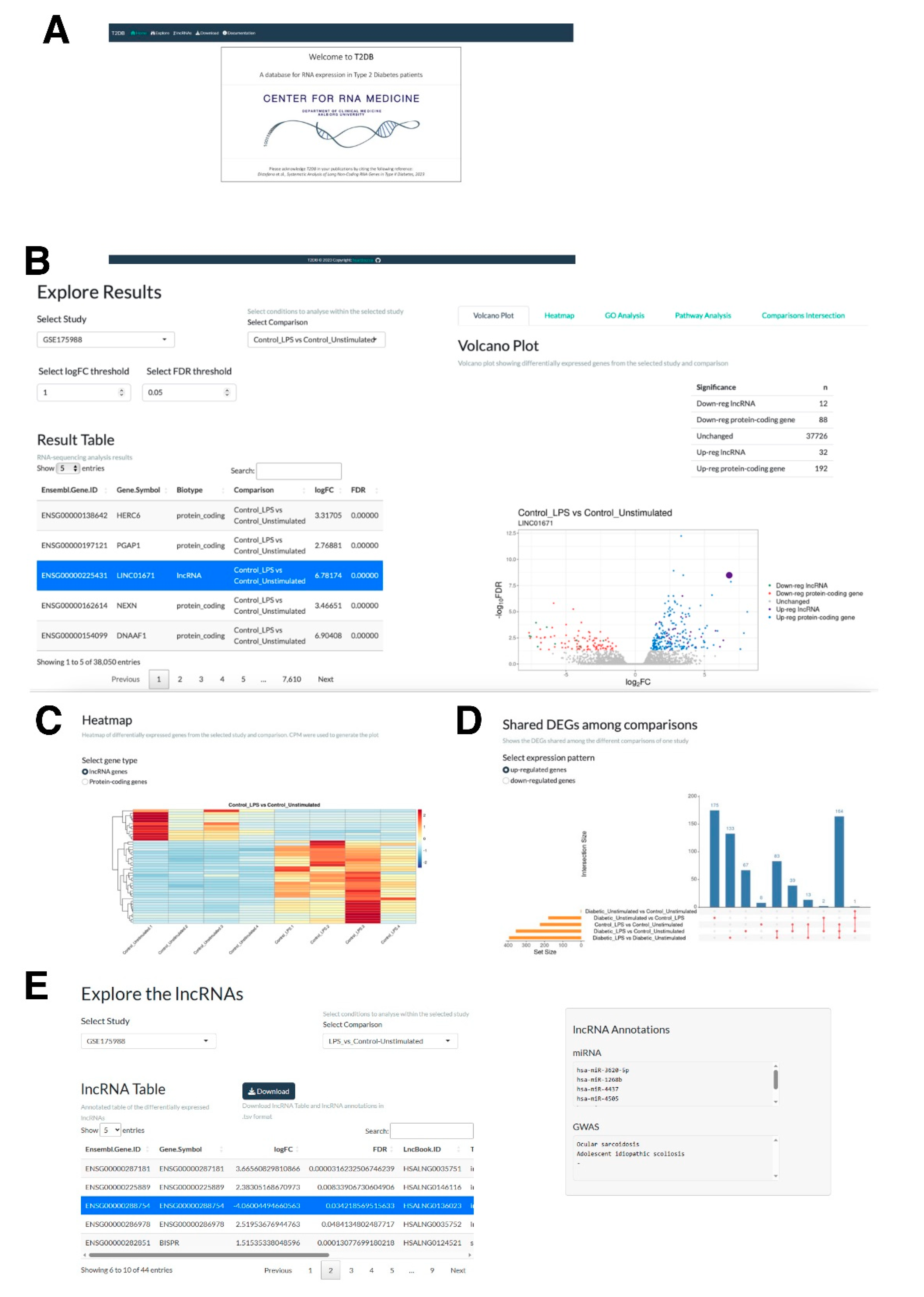
2.5. The Web Application, T2DB, for Expression Analysis of Protein-Coding and lncRNA Genes
3. Discussion
4. Materials and Methods
4.1. RNA-Seq Data Analysis and Visualization
4.2. Cell Culture and Treatments
4.3. Isolation of Total RNA and RT-PCR
4.4. T2DB Web Application
4.5. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Seal, R.L.; Chen, L.L.; Griffiths-Jones, S.; Lowe, T.M.; Mathews, M.B.; O’Reilly, D.; Pierce, A.J.; Stadler, P.F.; Ulitsky, I.; Wolin, S.L.; et al. A guide to naming human non-coding RNA genes. EMBO J. 2020, 39, e103777. [Google Scholar] [CrossRef]
- Uchida, S.; Adams, J.C. Physiological roles of non-coding RNAs. Am. J. Physiol. Cell. Physiol. 2019, 317, C1–C2. [Google Scholar] [CrossRef]
- Nojima, T.; Proudfoot, N.J. Mechanisms of lncRNA biogenesis as revealed by nascent transcriptomics. Nat. Rev. Mol. Cell. Biol. 2022, 23, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Klapproth, C.; Sen, R.; Stadler, P.F.; Findeiss, S.; Fallmann, J. Common Features in lncRNA Annotation and Classification: A Survey. Noncoding RNA 2021, 7, 77. [Google Scholar] [CrossRef]
- Seal, R.L.; Tweedie, S.; Bruford, E.A. A standardised nomenclature for long non-coding RNAs. IUBMB Life 2022, 75, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.W. A short guide to long non-coding RNA gene nomenclature. Human Genomics 2014, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, J.; Li, Y.; Song, T.; Wu, Y.; Fang, S.; Bu, D.; Li, H.; Sun, L.; Pei, D.; et al. NONCODEV6: An updated database dedicated to long non-coding RNA annotation in both animals and plants. Nucleic Acids Res. 2021, 49, D165–D171. [Google Scholar] [CrossRef]
- Uszczynska-Ratajczak, B.; Lagarde, J.; Frankish, A.; Guigo, R.; Johnson, R. Towards a complete map of the human long non-coding RNA transcriptome. Nat. Rev. Genet. 2018, 19, 535–548. [Google Scholar] [CrossRef]
- Sommerauer, C.; Kutter, C. Noncoding RNAs and RNA-binding proteins: Emerging governors of liver physiology and metabolic diseases. Am. J. Physiol. Cell. Physiol. 2022, 323, C1003–C1017. [Google Scholar] [CrossRef] [PubMed]
- Philip, M.; Chen, T.; Tyagi, S. A Survey of Current Resources to Study lncRNA-Protein Interactions. Noncoding RNA 2021, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Pisignano, G.; Ladomery, M. Epigenetic Regulation of Alternative Splicing: How LncRNAs Tailor the Message. Noncoding RNA 2021, 7, 21. [Google Scholar] [CrossRef]
- Karakas, D.; Ozpolat, B. The Role of LncRNAs in Translation. Noncoding RNA 2021, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell. Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, G.K.; Khullar, N.; Sidhu, I.S.; Navik, U.S.; Reddy, A.P.; Reddy, P.H.; Bhatti, J.S. Emerging role of non-coding RNA in health and disease. Metab. Brain Dis. 2021, 36, 1119–1134. [Google Scholar] [CrossRef]
- Lopez-Jimenez, E.; Andres-Leon, E. The Implications of ncRNAs in the Development of Human Diseases. Noncoding RNA 2021, 7, 17. [Google Scholar] [CrossRef]
- Sparber, P.; Filatova, A.; Khantemirova, M.; Skoblov, M. The role of long non-coding RNAs in the pathogenesis of hereditary diseases. BMC Med. Genomics 2019, 12, 42. [Google Scholar] [CrossRef]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- Khan, M.A.B.; Hashim, M.J.; King, J.K.; Govender, R.D.; Mustafa, H.; Al Kaabi, J. Epidemiology of Type 2 Diabetes—Global Burden of Disease and Forecasted Trends. J. Epidemiol. Glob. Health 2020, 10, 107–111. [Google Scholar] [CrossRef]
- Schnurr, T.M.; Jakupovic, H.; Carrasquilla, G.D.; Angquist, L.; Grarup, N.; Sorensen, T.I.A.; Tjonneland, A.; Overvad, K.; Pedersen, O.; Hansen, T.; et al. Obesity, unfavourable lifestyle and genetic risk of type 2 diabetes: A case-cohort study. Diabetologia 2020, 63, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Kyrou, I.; Tsigos, C.; Mavrogianni, C.; Cardon, G.; Van Stappen, V.; Latomme, J.; Kivela, J.; Wikstrom, K.; Tsochev, K.; Nanasi, A.; et al. Sociodemographic and lifestyle-related risk factors for identifying vulnerable groups for type 2 diabetes: A narrative review with emphasis on data from Europe. BMC Endocr. Disord. 2020, 20, 134. [Google Scholar] [CrossRef] [PubMed]
- Leong, A.; Porneala, B.; Dupuis, J.; Florez, J.C.; Meigs, J.B. Type 2 Diabetes Genetic Predisposition, Obesity, and All-Cause Mortality Risk in the U.S.: A Multiethnic Analysis. Diabetes Care 2016, 39, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Martin-Timon, I.; Sevillano-Collantes, C.; Segura-Galindo, A.; Del Canizo-Gomez, F.J. Type 2 diabetes and cardiovascular disease: Have all risk factors the same strength? World J. Diabetes 2014, 5, 444–470. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhou, T.; Heianza, Y.; Li, X.; Fan, M.; Fonseca, V.A.; Qi, L. Type 2 Diabetes and Hypertension. Circ. Res. 2019, 124, 930–937. [Google Scholar] [CrossRef] [PubMed]
- Duh, E.J.; Sun, J.K.; Stitt, A.W. Diabetic retinopathy: Current understanding, mechanisms, and treatment strategies. JCI Insight 2017, 2, e93751. [Google Scholar] [CrossRef]
- McGill, J.B.; Haller, H.; Roy-Chaudhury, P.; Cherrington, A.; Wada, T.; Wanner, C.; Ji, L.; Rossing, P. Making an impact on kidney disease in people with type 2 diabetes: The importance of screening for albuminuria. BMJ Open Diabetes Res. Care 2022, 10, e002806. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Primers 2015, 1, 15019. [Google Scholar] [CrossRef]
- Olokoba, A.B.; Obateru, O.A.; Olokoba, L.B. Type 2 diabetes mellitus: A review of current trends. Oman Med. J. 2012, 27, 269–273. [Google Scholar] [CrossRef]
- Rorsman, P.; Ashcroft, F.M. Pancreatic beta-Cell Electrical Activity and Insulin Secretion: Of Mice and Men. Physiol. Rev. 2018, 98, 117–214. [Google Scholar] [CrossRef]
- Cerf, M.E. Beta cell dysfunction and insulin resistance. Front. Endocrinol. 2013, 4, 37. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Eckel, J. Organ Crosstalk and the Modulation of Insulin Signaling. Cells 2021, 10, 2082. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R. Insulin signaling in health and disease. J. Clin. Investig. 2021, 131, e142241. [Google Scholar] [CrossRef] [PubMed]
- Boucher, J.; Kleinridders, A.; Kahn, C.R. Insulin receptor signaling in normal and insulin-resistant states. Cold Spring Harb. Perspect. Biol. 2014, 6, 9191. [Google Scholar] [CrossRef]
- Formichi, C.; Nigi, L.; Grieco, G.E.; Maccora, C.; Fignani, D.; Brusco, N.; Licata, G.; Sebastiani, G.; Dotta, F. Non-Coding RNAs: Novel Players in Insulin Resistance and Related Diseases. Int. J. Mol. Sci. 2021, 22, 7716. [Google Scholar] [CrossRef]
- Tello-Flores, V.A.; Beltran-Anaya, F.O.; Ramirez-Vargas, M.A.; Esteban-Casales, B.E.; Navarro-Tito, N.; Alarcon-Romero, L.D.C.; Luciano-Villa, C.A.; Ramirez, M.; Del Moral-Hernandez, O.; Flores-Alfaro, E. Role of Long Non-Coding RNAs and the Molecular Mechanisms Involved in Insulin Resistance. Int. J. Mol. Sci. 2021, 22, 7256. [Google Scholar] [CrossRef]
- Zhang, T.N.; Wang, W.; Yang, N.; Huang, X.M.; Liu, C.F. Regulation of Glucose and Lipid Metabolism by Long Non-coding RNAs: Facts and Research Progress. Front. Endocrinol. 2020, 11, 457. [Google Scholar] [CrossRef]
- Degirmenci, U.; Li, J.; Lim, Y.C.; Siang, D.T.C.; Lin, S.; Liang, H.; Sun, L. Silencing an insulin-induced lncRNA, LncASIR, impairs the transcriptional response to insulin signalling in adipocytes. Sci. Rep. 2019, 9, 5608. [Google Scholar] [CrossRef]
- Chi, T.; Lin, J.; Wang, M.; Zhao, Y.; Liao, Z.; Wei, P. Non-Coding RNA as Biomarkers for Type 2 Diabetes Development and Clinical Management. Front. Endocrinol. 2021, 12, 630032. [Google Scholar] [CrossRef]
- Dieter, C.; Lemos, N.E.; Correa, N.R.F.; Assmann, T.S.; Crispim, D. The Impact of lncRNAs in Diabetes Mellitus: A Systematic Review and In Silico Analyses. Front. Endocrinol. 2021, 12, 602597. [Google Scholar] [CrossRef]
- Chen, Y.; He, Y.; Zhou, H. The potential role of lncRNAs in diabetes and diabetic microvascular complications. Endocr. J. 2020, 67, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ma, T.K.; Wen, S.; Li, L.L.; Xu, L.; Zhu, X.W.; Zhang, C.X.; Liu, N.; Wang, X.; Fan, Q.L. LncRNA ARAP1-AS2 promotes high glucose-induced human proximal tubular cell injury via persistent transactivation of the EGFR by interacting with ARAP1. J. Cell. Mol. Med. 2020, 24, 12994–13009. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Yang, Y.; Chen, X.; Liu, Y.; Hu, Q.; Huang, B.; Liu, Y.; Pan, Y.; Zhang, Y.; Liu, D.; et al. The long non-coding RNA betaFaar regulates islet beta-cell function and survival during obesity in mice. Nat. Commun. 2021, 12, 3997. [Google Scholar] [CrossRef]
- Wang, M.; Wang, S.; Yao, D.; Yan, Q.; Lu, W. A novel long non-coding RNA CYP4B1-PS1-001 regulates proliferation and fibrosis in diabetic nephropathy. Mol. Cell. Endocrinol. 2016, 426, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Reddy, M.A.; Senapati, P.; Stapleton, K.; Lanting, L.; Wang, M.; Amaram, V.; Ganguly, R.; Zhang, L.; Devaraj, S.; et al. Diabetes Mellitus-Induced Long Noncoding RNA Dnm3os Regulates Macrophage Functions and Inflammation via Nuclear Mechanisms. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 1806–1820. [Google Scholar] [CrossRef]
- Reddy, M.A.; Amaram, V.; Das, S.; Tanwar, V.S.; Ganguly, R.; Wang, M.; Lanting, L.; Zhang, L.; Abdollahi, M.; Chen, Z.; et al. lncRNA DRAIR is downregulated in diabetic monocytes and modulates the inflammatory phenotype via epigenetic mechanisms. JCI Insight 2021, 6, e143289. [Google Scholar] [CrossRef]
- Ge, X.; Xu, B.; Xu, W.; Xia, L.; Xu, Z.; Shen, L.; Peng, W.; Huang, S. Long noncoding RNA GAS5 inhibits cell proliferation and fibrosis in diabetic nephropathy by sponging miR-221 and modulating SIRT1 expression. Aging 2019, 11, 8745–8759. [Google Scholar] [CrossRef]
- Cui, X.; Tan, J.; Shi, Y.; Sun, C.; Li, Y.; Ji, C.; Wu, J.; Zhang, Z.; Chen, S.; Guo, X.; et al. The long non-coding RNA Gm10768 activates hepatic gluconeogenesis by sequestering microRNA-214 in mice. J. Biol. Chem. 2018, 293, 4097–4109. [Google Scholar] [CrossRef]
- Luo, R.; Xiao, F.; Wang, P.; Hu, Y.X. lncRNA H19 sponging miR-93 to regulate inflammation in retinal epithelial cells under hyperglycemia via XBP1s. Inflamm. Res. 2020, 69, 255–265. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Y.; Liu, Z.; Lin, B.; Deng, X.; Xiao, Q.; Chen, Z.; Ye, H.; Chen, D.; Su, Y.; et al. Downregulation of Kcnq1ot1 attenuates beta-cell proliferation and insulin secretion via the miR-15b-5p/Ccnd1 and Ccnd2 axis. Acta Diabetol. 2022, 59, 885–899. [Google Scholar] [CrossRef]
- Niu, B.; Yao, L.; Zhang, Y.; Xia, X.; Su, H. LncRNA KCNQ1OT1 promoted hepatitis C virus-induced pyroptosis of beta-cell through mediating the miR-223-3p/NLRP3 axis. Ann. Transl. Med. 2021, 9, 1387. [Google Scholar] [CrossRef]
- Cao, Z.; Yao, F.; Lang, Y.; Feng, X. Elevated Circulating LINC-P21 Serves as a Diagnostic Biomarker of Type 2 Diabetes Mellitus and Regulates Pancreatic beta-cell Function by Sponging miR-766-3p to Upregulate NR3C2. Exp. Clin. Endocrinol. Diabetes 2022, 130, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Geng, J.; Li, X.; Wan, J.; Liu, J.; Zhou, Z.; Liu, X. Long Noncoding RNA LINC01619 Regulates MicroRNA-27a/Forkhead Box Protein O1 and Endoplasmic Reticulum Stress-Mediated Podocyte Injury in Diabetic Nephropathy. Antioxid. Redox Signal. 2018, 29, 355–376. [Google Scholar] [CrossRef]
- Wang, J.; Yang, W.; Chen, Z.; Chen, J.; Meng, Y.; Feng, B.; Sun, L.; Dou, L.; Li, J.; Cui, Q.; et al. Long Noncoding RNA lncSHGL Recruits hnRNPA1 to Suppress Hepatic Gluconeogenesis and Lipogenesis. Diabetes 2018, 67, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Liu, Y.; Cui, J.; Lv, J.; Li, C.; Lu, J.; Huo, X.; Dou, J.; Bai, Z.; Chen, Z.; et al. LncRNA LYPLAL1-DT screening from type 2 diabetes with macrovascular complication contributes protective effects on human umbilical vein endothelial cells via regulating the miR-204-5p/SIRT1 axis. Cell. Death Discov. 2022, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Yao, J.; Liu, J.Y.; Li, X.M.; Wang, X.Q.; Li, Y.J.; Tao, Z.F.; Song, Y.C.; Chen, Q.; Jiang, Q. lncRNA-MIAT regulates microvascular dysfunction by functioning as a competing endogenous RNA. Circ. Res. 2015, 116, 1143–1156. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, S.H.; Liu, S.; Wang, Q. Role of lncRNA NEAT1 mediated by YY1 in the development of diabetic cataract via targeting the microRNA-205-3p/MMP16 axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5863–5870. [Google Scholar] [CrossRef]
- Chen, Q.; He, Y.; Wang, X.; Zhu, Y.; Huang, Y.; Cao, J.; Yan, R. LncRNA PTGS2 regulates islet beta-cell function through the miR-146a-5p/RBP4 axis and its diagnostic value in type 2 diabetes mellitus. Am. J. Transl. Res. 2021, 13, 11316–11328. [Google Scholar]
- Cai, Q.; Wang, C.; Huang, L.; Wu, C.; Yan, B.; Chen, T.; Li, Q.; Wang, L. Long Non-Coding RNA Small Nucleolar RNA Host Gene 5 (SNHG5) Regulates Renal Tubular Damage in Diabetic Nephropathy via Targeting MiR-26a-5p. Horm. Metab. Res. 2021, 53, 818–824. [Google Scholar] [CrossRef]
- Zhang, P.; Li, Y.N.; Tu, S.; Cheng, X.B. SP1-induced lncRNA TUG1 regulates proliferation and apoptosis in islet cells of type 2 diabetes mellitus via the miR-188-3p/FGF5 axis. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 1959–1966. [Google Scholar] [CrossRef]
- Zhou, A.X.; Mondal, T.; Tabish, A.M.; Abadpour, S.; Ericson, E.; Smith, D.M.; Knoll, R.; Scholz, H.; Kanduri, C.; Tyrberg, B.; et al. The long noncoding RNA TUNAR modulates Wnt signaling and regulates human beta-cell proliferation. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E846–E857. [Google Scholar] [CrossRef] [PubMed]
- Vanormelingen, C.; Tack, J.; Andrews, C.N. Diabetic gastroparesis. Br. Med. Bull. 2013, 105, 213–230. [Google Scholar] [CrossRef] [PubMed]
- Young, C.F.; Moussa, M.; Shubrook, J.H. Diabetic Gastroparesis: A Review. Diabetes Spectr. 2020, 33, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Grover, M.; Gibbons, S.J.; Nair, A.A.; Bernard, C.E.; Zubair, A.S.; Eisenman, S.T.; Wilson, L.A.; Miriel, L.; Pasricha, P.J.; Parkman, H.P.; et al. Transcriptomic signatures reveal immune dysregulation in human diabetic and idiopathic gastroparesis. BMC Med. Genomics 2018, 11, 62. [Google Scholar] [CrossRef]
- Wigger, L.; Barovic, M.; Brunner, A.D.; Marzetta, F.; Schoniger, E.; Mehl, F.; Kipke, N.; Friedland, D.; Burdet, F.; Kessler, C.; et al. Multi-omics profiling of living human pancreatic islet donors reveals heterogeneous beta cell trajectories towards type 2 diabetes. Nat. Metab. 2021, 3, 1017–1031. [Google Scholar] [CrossRef] [PubMed]
- Hart, P.A.; Bellin, M.D.; Andersen, D.K.; Bradley, D.; Cruz-Monserrate, Z.; Forsmark, C.E.; Goodarzi, M.O.; Habtezion, A.; Korc, M.; Kudva, Y.C.; et al. Type 3c (pancreatogenic) diabetes mellitus secondary to chronic pancreatitis and pancreatic cancer. Lancet Gastroenterol. Hepatol. 2016, 1, 226–237. [Google Scholar] [CrossRef]
- Rao, S.S.; Disraeli, P.; McGregor, T. Impaired glucose tolerance and impaired fasting glucose. Am. Fam. Physician 2004, 69, 1961–1968. [Google Scholar]
- Li, Z.; Liu, L.; Feng, C.; Qin, Y.; Xiao, J.; Zhang, Z.; Ma, L. LncBook 2.0: Integrating human long non-coding RNAs with multi-omics annotations. Nucleic Acids Res. 2023, 51, D186–D191. [Google Scholar] [CrossRef]
- Corella, D.; Sorli, J.V.; Gonzalez, J.I.; Ortega, C.; Fito, M.; Bullo, M.; Martinez-Gonzalez, M.A.; Ros, E.; Aros, F.; Lapetra, J.; et al. Novel association of the obesity risk-allele near Fas Apoptotic Inhibitory Molecule 2 (FAIM2) gene with heart rate and study of its effects on myocardial infarction in diabetic participants of the PREDIMED trial. Cardiovasc. Diabetol. 2014, 13, 5. [Google Scholar] [CrossRef]
- Moriggi, M.; Belloli, S.; Barbacini, P.; Murtaj, V.; Torretta, E.; Chaabane, L.; Canu, T.; Penati, S.; Malosio, M.L.; Esposito, A.; et al. Skeletal Muscle Proteomic Profile Revealed Gender-Related Metabolic Responses in a Diet-Induced Obesity Animal Model. Int. J. Mol. Sci. 2021, 22, 4680. [Google Scholar] [CrossRef]
- Jia, G.; Sowers, J.R. Targeting CITED2 for Angiogenesis in Obesity and Insulin Resistance. Diabetes 2016, 65, 3535–3536. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lockhart, S.M.; Rathjen, T.; Albadawi, H.; Sorensen, D.; O’Neill, B.T.; Dwivedi, N.; Preil, S.R.; Beck, H.C.; Dunwoodie, S.L.; et al. Insulin Downregulates the Transcriptional Coregulator CITED2, an Inhibitor of Proangiogenic Function in Endothelial Cells. Diabetes 2016, 65, 3680–3690. [Google Scholar] [CrossRef] [PubMed]
- Sakai, M.; Matsumoto, M.; Tujimura, T.; Yongheng, C.; Noguchi, T.; Inagaki, K.; Inoue, H.; Hosooka, T.; Takazawa, K.; Kido, Y.; et al. CITED2 links hormonal signaling to PGC-1alpha acetylation in the regulation of gluconeogenesis. Nat. Med. 2012, 18, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Edgar, L.; Akbar, N.; Braithwaite, A.T.; Krausgruber, T.; Gallart-Ayala, H.; Bailey, J.; Corbin, A.L.; Khoyratty, T.E.; Chai, J.T.; Alkhalil, M.; et al. Hyperglycemia Induces Trained Immunity in Macrophages and Their Precursors and Promotes Atherosclerosis. Circulation 2021, 144, 961–982. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Zhang, X.; Chao, L.; Zheng, Y.; Tan, Y.; Wang, L.; Zhang, W. Comprehensive analysis of key genes, microRNAs and long non-coding RNAs in hepatocellular carcinoma. FEBS Open Bio. 2018, 8, 1424–1436. [Google Scholar] [CrossRef]
- Gupta, M.; Martens, K.; Metz, L.M.; de Koning, A.J.; Pfeffer, G. Long noncoding RNAs associated with phenotypic severity in multiple sclerosis. Mult. Scler. Relat. Disord. 2019, 36, 101407. [Google Scholar] [CrossRef]
- Yan, Q.; Forno, E.; Herrera-Luis, E.; Pino-Yanes, M.; Qi, C.; Rios, R.; Han, Y.Y.; Kim, S.; Oh, S.; Acosta-Perez, E.; et al. A genome-wide association study of severe asthma exacerbations in Latino children and adolescents. Eur. Respir. J. 2021, 57, 2002693. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Lin, H.C.; Liu, H.; Gan, D.; Jin, W.; Cui, C.; Yan, Y.; Qian, Y.; Han, C.; Wang, Z. A 6 lncRNA-Based Risk Score System for Predicting the Recurrence of Colon Adenocarcinoma Patients. Front. Oncol. 2020, 10, 81. [Google Scholar] [CrossRef]
- Cheng, C.; Wang, Q.; Zhu, M.; Liu, K.; Zhang, Z. Integrated analysis reveals potential long non-coding RNA biomarkers and their potential biological functions for disease free survival in gastric cancer patients. Cancer Cell. Int. 2019, 19, 123. [Google Scholar] [CrossRef]
- Yin, X.; Yang, Z.; Zhu, M.; Chen, C.; Huang, S.; Li, X.; Zhong, H.; Wen, H.; Sun, Q.; Yu, X.; et al. ILF2 Contributes to Hyperproliferation of Keratinocytes and Skin Inflammation in a KLHDC7B-DT-Dependent Manner in Psoriasis. Front. Genet. 2022, 13, 890624. [Google Scholar] [CrossRef]
- Werba, G.; Gonda, T.A. Missing links—Epigenetic regulators of the pancreatic cancer-associated inflammation. Clin. Sci. 2021, 135, 1289–1293. [Google Scholar] [CrossRef]
- Li, M.X.; Wang, H.Y.; Yuan, C.H.; Ma, Z.L.; Jiang, B.; Li, L.; Zhang, L.; Xiu, D.R. KLHDC7B-DT aggravates pancreatic ductal adenocarcinoma development via inducing cross-talk between cancer cells and macrophages. Clin. Sci. 2021, 135, 629–649. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Ding, Y.; Wang, S.; Xu, L.; Yu, H. The Construction and Analysis of the Aberrant lncRNA-miRNA-mRNA Network in Adipose Tissue from Type 2 Diabetes Individuals with Obesity. J. Diabetes Res. 2020, 2020, 3980742. [Google Scholar] [CrossRef]
- Gao, W.Z.; Guo, L.M.; Xu, T.Q.; Yin, Y.H.; Jia, F. Identification of a multidimensional transcriptome signature for survival prediction of postoperative glioblastoma multiforme patients. J. Transl. Med. 2018, 16, 368. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, X.; Bu, X. LINC02381 contributes to cell proliferation and hinders cell apoptosis in glioma by transcriptionally enhancing CBX5. Brain Res. Bull. 2021, 176, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Priyanka, P.; Sharma, M.; Das, S.; Saxena, S. The lncRNA HMS recruits RNA-binding protein HuR to stabilize the 3′-UTR of HOXC10 mRNA. J. Biol. Chem. 2021, 297, 100997. [Google Scholar] [CrossRef] [PubMed]
- Nemati, H.; Fakhre-Taha, M.; Javanmard, A.R.; Jahanbakhshi, A.; Mowla, S.J.; Soltani, B.M. LINC02381-ceRNA exerts its oncogenic effect through regulation of IGF1R signaling pathway in glioma. J. Neurooncol. 2022, 158, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Huang, P.; Wu, H.; Wang, S.; Liu, G. LINC02381 aggravates breast cancer through the miR-1271-5p/FN1 axis to activate PI3K/AKT pathway. Mol. Carcinog. 2022, 61, 346–358. [Google Scholar] [CrossRef]
- Zhao, G.; Luo, W.D.; Yuan, Y.; Lin, F.; Guo, L.M.; Ma, J.J.; Chen, H.B.; Tang, H.; Shu, J. LINC02381, a sponge of miR-21, weakens osteogenic differentiation of hUC-MSCs through KLF12-mediated Wnt4 transcriptional repression. J. Bone Miner. Metab. 2022, 40, 66–80. [Google Scholar] [CrossRef]
- Bian, X.; Sun, Y.M.; Wang, L.M.; Shang, Y.L. ELK1-induced upregulation lncRNA LINC02381 accelerates the osteosarcoma tumorigenesis through targeting CDCA4 via sponging miR-503-5p. Biochem. Biophys. Res. Commun. 2021, 548, 112–119. [Google Scholar] [CrossRef]
- Jafarzadeh, M.; Soltani, B.M. Long Noncoding RNA LOC400043 (LINC02381) Inhibits Gastric Cancer Progression through Regulating Wnt Signaling Pathway. Front. Oncol. 2020, 10, 562253. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, Q. Linc02381 Exacerbates Rheumatoid Arthritis through Adsorbing miR-590-5p and Activating the Mitogen-Activated Protein Kinase Signaling Pathway in Rheumatoid arthritis-fibroblast-like synoviocytes. Cell Transplant. 2020, 29, 963689720938023. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Z.; Ma, Y.; Su, H.; Xie, P.; Ran, J. LINC02381 Promoted Cell Viability and Migration via Targeting miR-133b in Cervical Cancer Cells. Cancer Manag. Res. 2020, 12, 3971–3979. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zou, X.; Mei, J. The Risk Correlation between N7-Methylguanosine Modification-Related lncRNAs and Survival Prognosis of Oral Squamous Cell Carcinoma Based on Comprehensive Bioinformatics Analysis. Appl. Bionics Biomech. 2022, 2022, 1666792. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhu, L.; Zhao, X.; Wu, X.; Yang, J.; Xu, B.; Zhao, Z.; Gu, L.; Su, L. LncRNA SERPINB9P1 expression and polymorphisms are associated with ischemic stroke in a Chinese Han population. Neurol. Sci. 2022, 43, 1143–1154. [Google Scholar] [CrossRef]
- Wang, S. Investigation of long non-coding RNA expression profiles in patients with post-menopausal osteoporosis by RNA sequencing. Exp. Ther. Med. 2020, 20, 1487–1497. [Google Scholar] [CrossRef]
- Liu, L.; Wang, H.; Zhang, X.; Chen, R. Identification of Potential Biomarkers in Neonatal Sepsis by Establishing a Competitive Endogenous RNA Network. Comb. Chem. High Throughput Screen. 2020, 23, 369–380. [Google Scholar] [CrossRef]
- Li, C.; Liang, X.; Liu, Y. lncRNA USP30-AS1 sponges miR-765 and modulates the progression of colon cancer. World J. Surg. Oncol. 2022, 20, 73. [Google Scholar] [CrossRef]
- Chen, M.; Chi, Y.; Chen, H.; Zhao, L. Long non-coding RNA USP30-AS1 aggravates the malignant progression of cervical cancer by sequestering microRNA-299-3p and thereby overexpressing PTP4A1. Oncol. Lett. 2021, 22, 505. [Google Scholar] [CrossRef]
- Zhou, W.; Xu, S.; Deng, T.; Zhou, R.; Wang, C. LncRNA USP30-AS1 promotes the survival of acute myeloid leukemia cells by cis-regulating USP30 and ANKRD13A. Hum. Cell 2022, 35, 360–378. [Google Scholar] [CrossRef]
- Lee, A.H.; Heidtman, K.; Hotamisligil, G.S.; Glimcher, L.H. Dual and opposing roles of the unfolded protein response regulated by IRE1alpha and XBP1 in proinsulin processing and insulin secretion. Proc. Natl. Acad. Sci. USA 2011, 108, 8885–8890. [Google Scholar] [CrossRef] [PubMed]
- Zeggini, E.; Weedon, M.N.; Lindgren, C.M.; Frayling, T.M.; Elliott, K.S.; Lango, H.; Timpson, N.J.; Perry, J.R.; Rayner, N.W.; Freathy, R.M.; et al. Replication of genome-wide association signals in UK samples reveals risk loci for type 2 diabetes. Science 2007, 316, 1336–1341. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Gao, Y.; Zhou, L.; Chen, J.; Xie, Z.; Ye, Z.; Wang, Y. USP30: Structure, Emerging Physiological Role, and Target Inhibition. Front. Pharmacol. 2022, 13, 851654. [Google Scholar] [CrossRef]
- Gil, N.; Ulitsky, I. Regulation of gene expression by cis-acting long non-coding RNAs. Nat. Rev. Genet. 2020, 21, 102–117. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, H.; Hu, D.; Liu, S. New algorithms based on autophagy-related lncRNAs pairs to predict the prognosis of skin cutaneous melanoma patients. Arch. Dermatol. Res. 2023, ahead of print. [Google Scholar] [CrossRef]
- Xiang, X.; Guo, Y.; Chen, Z.; Zhang, F.; Huang, J.; Qin, Y. A prognostic risk prediction model based on ferroptosis-related long non-coding RNAs in bladder cancer: A bulk RNA-seq research and scRNA-seq validation. Medicine 2022, 101, e32558. [Google Scholar] [CrossRef]
- Kang, Z.; Dou, Q.; Huang, T.; Tu, M.; Zhong, Y.; Wang, M.; Li, T. An angiogenesis-related lncRNA signature for the prognostic prediction of patients with bladder cancer and LINC02321 promotes bladder cancer progression via the VEGFA signaling pathway. Mol. Med. Rep. 2023, 27, 38. [Google Scholar] [CrossRef]
- Dai, Y.W.; Wen, Z.K.; Wu, Z.X.; Wu, H.D.; Lv, L.X.; Yan, C.Z.; Liu, C.H.; Wang, Z.Q.; Zheng, C. Amino Acid Metabolism-Related lncRNA Signature Predicts the Prognosis of Breast Cancer. Front. Genet. 2022, 13, 880387. [Google Scholar] [CrossRef]
- Qiu, Y.; Wang, H.T.; Zheng, X.F.; Huang, X.; Meng, J.Z.; Huang, J.P.; Wen, Z.P.; Yao, J. Autophagy-related long non-coding RNA prognostic model predicts prognosis and survival of melanoma patients. World J. Clin. Cases 2022, 10, 3334–3351. [Google Scholar] [CrossRef]
- Tao, S.; Tao, K.; Cai, X. Necroptosis-Associated lncRNA Prognostic Model and Clustering Analysis: Prognosis Prediction and Tumor-Infiltrating Lymphocytes in Breast Cancer. J. Oncol. 2022, 2022, 7099930. [Google Scholar] [CrossRef]
- Shi, G.J.; Zhou, Q.; Zhu, Q.; Wang, L.; Jiang, G.Q. A novel prognostic model associated with the overall survival in patients with breast cancer based on lipid metabolism-related long noncoding RNAs. J. Clin. Lab. Anal. 2022, 36, e24384. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Liu, L.; Li, P.; Yuan, Y.; Peng, M.; Jin, H.; Qin, D. Constructing a Novel Signature Based on Immune-Related lncRNA to Improve Prognosis Prediction of Cervical Squamous Cell Carcinoma Patients. Reprod. Sci. 2022, 29, 800–815. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, H.; Huang, Q.; Wu, J.; Zhang, M. A prognostic model based on immune-related long noncoding RNAs for patients with epithelial ovarian cancer. J. Ovarian Res. 2022, 15, 8. [Google Scholar] [CrossRef]
- Li, Z.; Wei, J.; Zheng, H.; Zhang, Y.; Song, M.; Cao, H.; Jin, Y. The new horizon of biomarker in melanoma patients: A study based on autophagy-related long non-coding RNA. Medicine 2022, 101, e28553. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Wu, P.; Zhao, X.; Jin, X.; Wang, J.; Shi, Y.; Yang, X.; She, Y.; Li, Y.; Li, C. Using Immune-Related lncRNA Signature for Prognosis and Response to Immunotherapy in Cutaneous Melanoma. Int. J. Gen. Med. 2021, 14, 6463–6475. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhou, Z.; Chai, Y.; Che, X.; Zhang, Y. Identification of a Nomogram from Ferroptosis-Related Long Noncoding RNAs Signature to Analyze Overall Survival in Patients with Bladder Cancer. J. Oncol. 2021, 2021, 8533464. [Google Scholar] [CrossRef]
- Ding, Y.; Li, T.; Li, M.; Tayier, T.; Zhang, M.; Chen, L.; Feng, S. A Novel Autophagy-Related lncRNA Gene Signature to Improve the Prognosis of Patients with Melanoma. Biomed. Res. Int. 2021, 2021, 8848227. [Google Scholar] [CrossRef]
- Mao, Y.; Lv, J.; Jiang, L.; Wang, Y. Integrative analysis of ceRNA network reveals functional lncRNAs associated with independent recurrent prognosis in colon adenocarcinoma. Cancer Cell. Int. 2021, 21, 352. [Google Scholar] [CrossRef]
- Wan, J.; Guo, C.; Fang, H.; Xu, Z.; Hu, Y.; Luo, Y. Autophagy-Related Long Non-coding RNA Is a Prognostic Indicator for Bladder Cancer. Front. Oncol. 2021, 11, 647236. [Google Scholar] [CrossRef]
- Gao, M.; Wang, X.; Han, D.; Lu, E.; Zhang, J.; Zhang, C.; Wang, L.; Yang, Q.; Jiang, Q.; Wu, J.; et al. A Six-lncRNA Signature for Immunophenotype Prediction of Glioblastoma Multiforme. Front. Genet. 2020, 11, 604655. [Google Scholar] [CrossRef]
- Chen, P.; Gao, Y.; Ouyang, S.; Wei, L.; Zhou, M.; You, H.; Wang, Y. A Prognostic Model Based on Immune-Related Long Non-Coding RNAs for Patients with Cervical Cancer. Front. Pharmacol. 2020, 11, 585255. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Li, T.; Gao, S.; Yin, H.; Cao, H.; He, W. An epithelial-mesenchymal transition-related long noncoding RNA signature correlates with the prognosis and progression in patients with bladder cancer. Biosci. Rep. 2021, 41, BSR20203944. [Google Scholar] [CrossRef]
- Meng, C.; Zhou, J.Q.; Liao, Y.S. Autophagy-related long non-coding RNA signature for ovarian cancer. J. Int. Med. Res. 2020, 48, 300060520970761. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Jing, C.; Xiao, C.; Li, T. An autophagy-related long non-coding RNA prognostic signature accurately predicts survival outcomes in bladder urothelial carcinoma patients. Aging 2020, 12, 15624–15637. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, L.; Chen, L.L. Life without A tail: New formats of long noncoding RNAs. Int. J. Biochem. Cell. Biol. 2014, 54, 338–349. [Google Scholar] [CrossRef]
- Johnsson, P.; Lipovich, L.; Grander, D.; Morris, K.V. Evolutionary conservation of long non-coding RNAs; sequence, structure, function. Biochim. Biophys. Acta 2014, 1840, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Ilieva, M.; Dao, J.; Miller, H.E.; Madsen, J.H.; Bishop, A.J.R.; Kauppinen, S.; Uchida, S. Systematic Analysis of Long Non-Coding RNA Genes in Nonalcoholic Fatty Liver Disease. Noncoding RNA 2022, 8, 56. [Google Scholar] [CrossRef]
- Ilieva, M.; Miller, H.E.; Agarwal, A.; Paulus, G.K.; Madsen, J.H.; Bishop, A.J.R.; Kauppinen, S.; Uchida, S. FibroDB: Expression Analysis of Protein-Coding and Long Non-Coding RNA Genes in Fibrosis. Noncoding RNA 2022, 8, 13. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. In Use R! 2nd ed.; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.; Huber, W.; Pages, H.; Aboyoun, P.; Carlson, M.; Gentleman, R.; Morgan, M.T.; Carey, V.J. Software for computing and annotating genomic ranges. PLoS Comput. Biol. 2013, 9, e1003118. [Google Scholar] [CrossRef]
- Delprat, V.; Tellier, C.; Demazy, C.; Raes, M.; Feron, O.; Michiels, C. Cycling hypoxia promotes a pro-inflammatory phenotype in macrophages via JNK/p65 signaling pathway. Sci. Rep. 2020, 10, 882. [Google Scholar] [CrossRef]
- Chang, W.; Cheng, J.; Allaire, J.J.; Sievert, C.; Schloerke, B.; Xie, Y.; Allen, J.; McPherson, J.; Dipert, A.; Borges, B.; et al. Shiny: Web Application Framework for R. Available online: https://shiny.rstudio.com/ (accessed on 21 March 2023).
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Yu, G. Enrichplot: Visualization of Functional Enrichment Result. Available online: https://yulab-smu.top/biomedical-knowledge-mining-book/ (accessed on 21 March 2023).
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R package for the visualization of intersecting sets and their properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Official Gene Name | Official Gene Symbol and Alias | Reference |
|---|---|---|
| ARAP1 antisense RNA 2 | ARAP1-AS2 | [42] |
| TRAF3IP2 antisense RNA 1 | TRAF3IP2-AS1, also known as βFaar | [43] |
| cytochrome P450, family 4, subfamily b, polypeptide 1, pseudogene 1 | Cyp4b1-ps1, also known as CYP4B1-PS1-001 | [44] |
| DNM3 opposite strand/antisense RNA | Dnm3os | [45] |
| diabetes regulated anti-inflammatory lncRNA | DRAIR, also known as CPEB2-AS | [46] |
| growth arrest specific 5 | GAS5 | [47] |
| predicted gene 10768 | Gm10768 | [48] |
| H19 imprinted maternally expressed transcript | H19 | [49] |
| KCNQ1 opposite strand/antisense transcript 1 | KCNQ1OT1 | [50,51] |
| tumor protein p53 pathway corepressor 1 | TP53COR1, also known as LINC-P21 | [52] |
| long intergenic non-protein-coding RNA 1619 | LINC01619 | [53] |
| predicted gene, 19689 | Gm19689, also known as lncSHGL | [54] |
| LYPLAL1 divergent transcript | LYPLAL1-DT | [55] |
| myocardial infarction associated transcript | MIAT | [56] |
| nuclear paraspeckle assembly transcript 1 | NEAT1 | [57] |
| prostaglandin-endoperoxide synthase 2 | PTGS2 | [58] |
| small nucleolar RNA host gene 5 | SNHG5 | [59] |
| taurine up-regulated 1 | TUG1 | [60] |
| TCL1 upstream neural differentiation-associated RNA | TUNAR | [61] |
| ARAP1 antisense RNA 2 | ARAP1-AS2 | [42] |
| TRAF3IP2 antisense RNA 1 | TRAF3IP2-AS1, also known as βFaar | [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Distefano, R.; Ilieva, M.; Madsen, J.H.; Ishii, H.; Aikawa, M.; Rennie, S.; Uchida, S. T2DB: A Web Database for Long Non-Coding RNA Genes in Type II Diabetes. Non-Coding RNA 2023, 9, 30. https://doi.org/10.3390/ncrna9030030
Distefano R, Ilieva M, Madsen JH, Ishii H, Aikawa M, Rennie S, Uchida S. T2DB: A Web Database for Long Non-Coding RNA Genes in Type II Diabetes. Non-Coding RNA. 2023; 9(3):30. https://doi.org/10.3390/ncrna9030030
Chicago/Turabian StyleDistefano, Rebecca, Mirolyuba Ilieva, Jens Hedelund Madsen, Hideshi Ishii, Masanori Aikawa, Sarah Rennie, and Shizuka Uchida. 2023. "T2DB: A Web Database for Long Non-Coding RNA Genes in Type II Diabetes" Non-Coding RNA 9, no. 3: 30. https://doi.org/10.3390/ncrna9030030
APA StyleDistefano, R., Ilieva, M., Madsen, J. H., Ishii, H., Aikawa, M., Rennie, S., & Uchida, S. (2023). T2DB: A Web Database for Long Non-Coding RNA Genes in Type II Diabetes. Non-Coding RNA, 9(3), 30. https://doi.org/10.3390/ncrna9030030