
A Machine Learning Model Based on microRNAs for the Diagnosis of Essential Hypertension

 ,
,  and
and 
Abstract
:
1. Introduction
2. Results
2.1. Study Participants
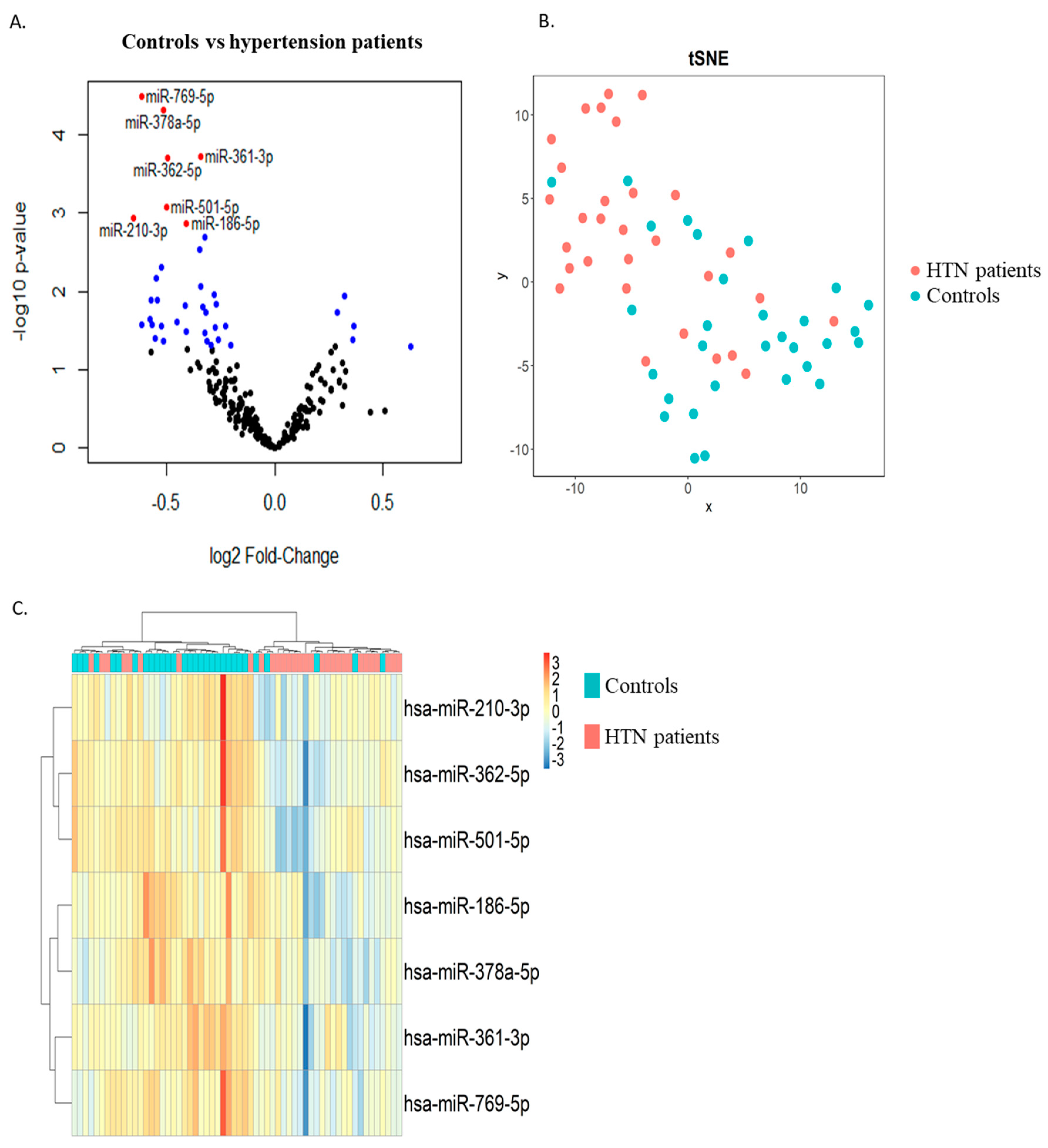
2.2. Discovery Phase
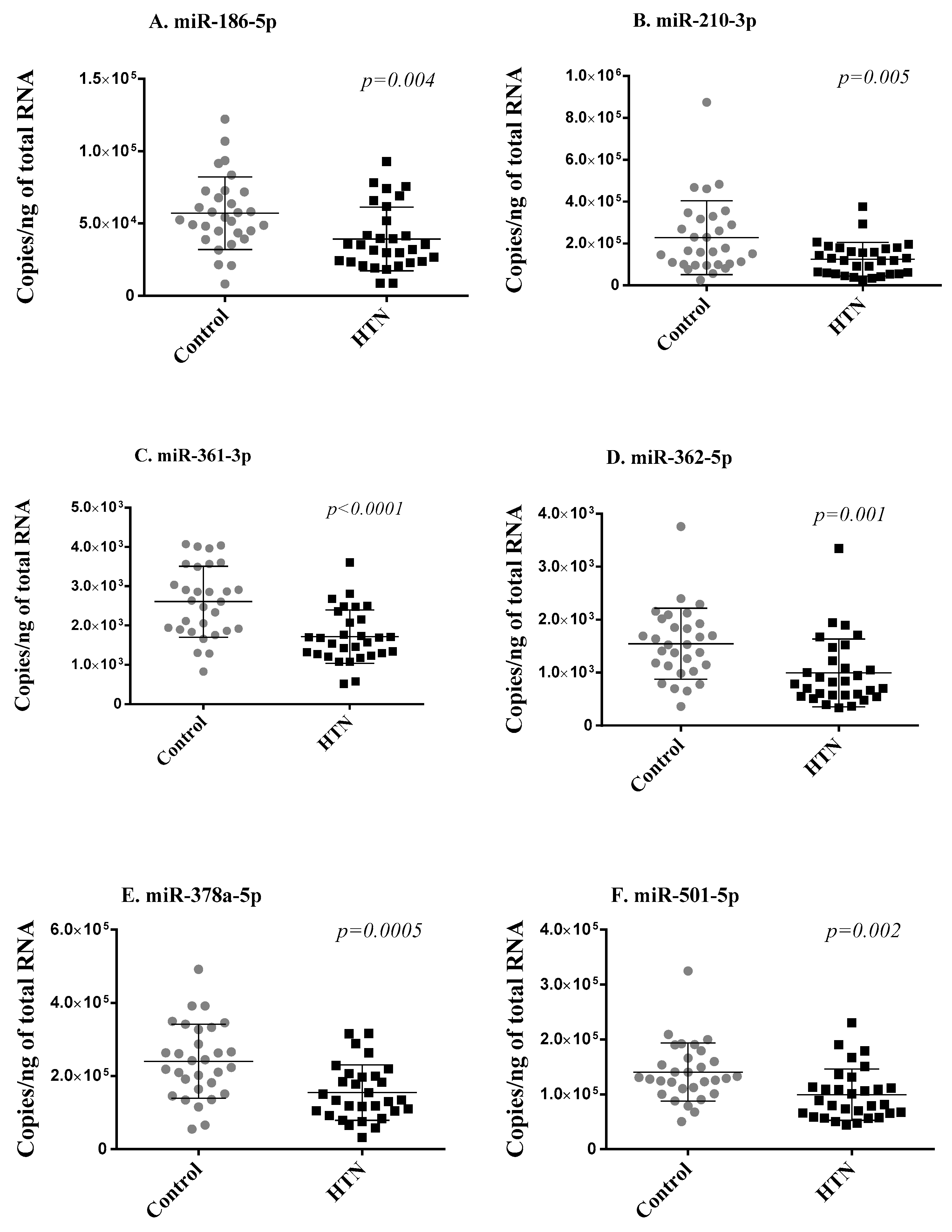
2.3. Replication Phase
2.4. Validation Phase
2.5. Effect of Medication
2.6. Correlation between miRNA Levels and Clinical Features
2.7. Machine Learning Model for Hypertension Prediction
3. Discussion
4. Perspectives
5. Methods
5.1. Study Design and Procedures
5.2. Whole Blood Collection and RNA Isolation
5.3. Small RNA Sequencing
5.4. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
5.5. Machine Learning Model Selection and Classification
5.6. Statistical Analysis
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 10CV | 10-fold cross-validation |
| AH | Arterial hypertension |
| AUC | Area under the receiver operating characteristic curve |
| AI | Artificial intelligence |
| BP | Blood pressure |
| ECG | Electrocardiogram |
| HFpEF | Heart failure with preserved Ejection Fraction |
| XGBoost, XGB | Extreme gradient boosting |
| kNN | k-nearest neighbor’s |
| LOOCV | Leave-One-Out Cross-Validation |
| LinearSVC | Linear support vector machine |
| Logit | Logistic regression |
| ML | Machine learning |
| miRNAs | MicroRNAs |
| MLP | Multi-layer perceptron |
| ncRNA | Non-coding RNA |
| NRT | No reverse transcriptase control |
| NTC | No template control |
| RF | Random forest |
| RT-qPCR | Real-time quantitative polymerase chain reaction |
| SVM | Support vector machine |
| t-SNE | t-distributed stochastic neighbor embedding |
References
- NCD Risk Factor Collaboration NCD-RisC. Worldwide trends in hypertension prevalence and progress in treatment and control from 1990 to 2019: A pooled analysis of 1201 population-representative studies with 104 million participants. Lancet 2021, 398, 957–980. [Google Scholar] [CrossRef]
- Huguet, N.; Larson, A.; Angier, H.; Marino, M.; Green, B.B.; Moreno, L.; DeVoe, J.E. Rates of Undiagnosed Hypertension and Diagnosed Hypertension Without Anti-hypertensive Medication Following the Affordable Care Act. Am. J. Hypertens. 2021, 34, 989–998. [Google Scholar] [CrossRef]
- Mahdi, A.; Armitage, L.C.; Tarassenko, L.; Watkinson, P. Estimated Prevalence of Hypertension and Undiagnosed Hypertension in a Large Inpatient Population: A Cross-sectional Observational Study. Am. J. Hypertens. 2021, 34, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Ivicevic Uhernik, A.; Kralj, V.; Cukelj, P.; Brkic-Bilos, I.; Erceg, M.; Benjak, T.; Stevanovic, R. Undiagnosed hypertension in Croatia. Croat. Med. J. 2023, 64, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Palano, F.; Paneni, F.; Sciarretta, S.; Tocci, G.; Volpe, M. The progression from hypertension to congestive heart failure. Recenti Prog. Med. 2011, 102, 461–467. [Google Scholar] [CrossRef]
- Levy, D.; Larson, M.G.; Vasan, R.S.; Kannel, W.B.; Ho, K.K. The progression from hypertension to congestive heart failure. JAMA 1996, 275, 1557–1562. [Google Scholar] [CrossRef] [PubMed]
- Jusic, A.; Thomas, P.B.; Wettinger, S.B.; Dogan, S.; Farrugia, R.; Gaetano, C.; Tuna, B.G.; Pinet, F.; Robinson, E.L.; Tual-Chalot, S.; et al. Noncoding RNAs in age-related cardiovascular diseases. Ageing Res. Rev. 2022, 77, 101610. [Google Scholar] [CrossRef] [PubMed]
- Batkai, S.; Thum, T. MicroRNAs in hypertension: Mechanisms and therapeutic targets. Curr. Hypertens. Rep. 2012, 14, 79–87. [Google Scholar] [CrossRef]
- Jusic, A.; Devaux, Y.; EU-CardioRNA COST Action. Noncoding RNAs in Hypertension. Hypertension 2019, 74, 477–492. [Google Scholar] [CrossRef]
- Tocci, G.; Paneni, F.; Ponziani, B.; Volpe, M. Use of predictive markers to improve cardiovascular protection. Future Cardiol. 2007, 3, 447–456. [Google Scholar] [CrossRef]
- Yushko, K.; Koval, S.; Snihurska, I.; Mysnychenko, O. The associations of circulating microRNA-133a with parameters of hypertension heart disease in patients with arterial hypertension and obesity. Eur. Heart J. 2020, 41, ehaa946.2712. [Google Scholar] [CrossRef]
- Zhang, Z.; Joyce, B.T.; Kresovich, J.K.; Zheng, Y.; Zhong, J.; Patel, R.; Zhang, W.; Liu, L.; Dou, C.; McCracken, J.P.; et al. Blood pressure and expression of microRNAs in whole blood. PLoS ONE 2017, 12, e0173550. [Google Scholar] [CrossRef]
- Ozkan, G.; Ulusoy, S.; Geyik, E.; Erdem, Y. Down-regulation of miRNA 145 and up-regulation of miRNA 4516 may be associated with primary hypertension. J. Clin. Hypertens. 2019, 21, 1724–1731. [Google Scholar] [CrossRef] [PubMed]
- Matshazi, D.M.; Weale, C.J.; Erasmus, R.T.; Kengne, A.P.; Davids, S.F.G.; Raghubeer, S.; Hector, S.; Davison, G.M.; Matsha, T.E. MicroRNA Profiles in Normotensive and Hypertensive South African Individuals. Front. Cardiovasc. Med. 2021, 8, 645541. [Google Scholar] [CrossRef] [PubMed]
- Perez-Hernandez, J.; Riffo-Campos, A.L.; Ortega, A.; Martinez-Arroyo, O.; Perez-Gil, D.; Olivares, D.; Solaz, E.; Martinez, F.; Martinez-Hervas, S.; Chaves, F.J.; et al. Urinary- and Plasma-Derived Exosomes Reveal a Distinct MicroRNA Signature Associated With Albuminuria in Hypertension. Hypertension 2021, 77, 960–971. [Google Scholar] [CrossRef]
- Kontaraki, J.E.; Marketou, M.E.; Zacharis, E.A.; Parthenakis, F.I.; Vardas, P.E. MicroRNA-9 and microRNA-126 expression levels in patients with essential hypertension: Potential markers of target-organ damage. J. Am. Soc. Hypertens. 2014, 8, 368–375. [Google Scholar] [CrossRef]
- Tan, P.P.S.; Hall, D.; Chilian, W.M.; Chia, Y.C.; Mohd Zain, S.; Lim, H.M.; Kumar, D.N.; Ching, S.M.; Low, T.Y.; Md Noh, M.F.; et al. Exosomal microRNAs in the development of essential hypertension and its potential as biomarkers. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H1486–H1497. [Google Scholar] [CrossRef]
- Rana, I.; Velkoska, E.; Patel, S.K.; Burrell, L.M.; Charchar, F.J. MicroRNAs mediate the cardioprotective effect of angiotensin-converting enzyme inhibition in acute kidney injury. Am. J. Physiol. Renal Physiol. 2015, 309, F943–F954. [Google Scholar] [CrossRef]
- Vaskova, E.; Ikeda, G.; Tada, Y.; Wahlquist, C.; Mercola, M.; Yang, P.C. Sacubitril/Valsartan Improves Cardiac Function and Decreases Myocardial Fibrosis Via Downregulation of Exosomal miR-181a in a Rodent Chronic Myocardial Infarction Model. J. Am. Heart Assoc. 2020, 9, e015640. [Google Scholar] [CrossRef]
- Srivastava, S.P.; Goodwin, J.E.; Kanasaki, K.; Koya, D. Inhibition of Angiotensin-Converting Enzyme Ameliorates Renal Fibrosis by Mitigating DPP-4 Level and Restoring Antifibrotic MicroRNAs. Genes 2020, 11, 211. [Google Scholar] [CrossRef]
- Christou, C.D.; Mitsas, A.C.; Vlachavas, I.; Tsoulfas, G. The Use of Machine Learning in MicroRNA Diagnostics: Current Perspectives. Microrna 2022, 11, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Ríos, E.; Montesinos, L.; Alfaro-Ponce, M.; Pecchia, L. A review of machine learning in hypertension detection and blood pressure estimation based on clinical and physiological data. Biomed. Signal Process. Control. 2021, 68, 102813. [Google Scholar] [CrossRef]
- Padmanabhan, S.; Tran, T.Q.B.; Dominiczak, A.F. Artificial Intelligence in Hypertension: Seeing Through a Glass Darkly. Circ. Res. 2021, 128, 1100–1118. [Google Scholar] [CrossRef] [PubMed]
- Reel, P.S.; Reel, S.; van Kralingen, J.C.; Langton, K.; Lang, K.; Erlic, Z.; Larsen, C.K.; Amar, L.; Pamporaki, C.; Mulatero, P.; et al. Machine learning for classification of hypertension subtypes using multi-omics: A multi-centre, retrospective, data-driven study. EBioMedicine 2022, 84, 104276. [Google Scholar] [CrossRef]
- Angelaki, E.; Barmparis, G.D.; Kochiadakis, G.; Maragkoudakis, S.; Savva, E.; Kampanieris, E.; Kassotakis, S.; Kalomoirakis, P.; Vardas, P.; Tsironis, G.P.; et al. Artificial intelligence-based opportunistic screening for the detection of arterial hypertension through ECG signals. J. Hypertens. 2022, 40, 2494–2501. [Google Scholar] [CrossRef]
- de Gonzalo-Calvo, D.; Martinez-Camblor, P.; Bar, C.; Duarte, K.; Girerd, N.; Fellstrom, B.; Schmieder, R.E.; Jardine, A.G.; Massy, Z.A.; Holdaas, H.; et al. Improved cardiovascular risk prediction in patients with end-stage renal disease on hemodialysis using machine learning modeling and circulating microribonucleic acids. Theranostics 2020, 10, 8665–8676. [Google Scholar] [CrossRef]
- Errington, N.; Iremonger, J.; Pickworth, J.A.; Kariotis, S.; Rhodes, C.J.; Rothman, A.M.; Condliffe, R.; Elliot, C.A.; Kiely, D.G.; Howard, L.S.; et al. A diagnostic miRNA signature for pulmonary arterial hypertension using a consensus machine learning approach. EBioMedicine 2021, 69, 103444. [Google Scholar] [CrossRef]
- Singh, D.D.; Kim, Y.; Choi, S.A.; Han, I.; Yadav, D.K. Clinical Significance of MicroRNAs, Long Non-Coding RNAs, and CircRNAs in Cardiovascular Diseases. Cells 2023, 12, 1629. [Google Scholar] [CrossRef]
- Wu, G.; Jose, P.A.; Zeng, C. Noncoding RNAs in the Regulatory Network of Hypertension. Hypertension 2018, 72, 1047–1059. [Google Scholar] [CrossRef]
- Poller, W.; Dimmeler, S.; Heymans, S.; Zeller, T.; Haas, J.; Karakas, M.; Leistner, D.M.; Jakob, P.; Nakagawa, S.; Blankenberg, S.; et al. Non-coding RNAs in cardiovascular diseases: Diagnostic and therapeutic perspectives. Eur. Heart J. 2018, 39, 2704–2716. [Google Scholar] [CrossRef]
- Santulli, G. MicroRNAs and Endothelial (Dys) Function. J. Cell Physiol. 2016, 231, 1638–1644. [Google Scholar] [CrossRef] [PubMed]
- Parthenakis, F.; Marketou, M.; Kontaraki, J.; Patrianakos, A.; Nakou, H.; Touloupaki, M.; Vernardos, M.; Kochiadakis, G.; Chlouverakis, G.; Vardas, P. Low Levels of MicroRNA-21 Are a Marker of Reduced Arterial Stiffness in Well-Controlled Hypertension. J. Clin. Hypertens. 2017, 19, 235–240. [Google Scholar] [CrossRef]
- Chen, T.; Sun, M.R.; Zhou, Q.; Guzman, A.M.; Ramchandran, R.; Chen, J.; Ganesh, B.; Raj, J.U. Extracellular vesicles derived from endothelial cells in hypoxia contribute to pulmonary artery smooth muscle cell proliferation in-vitro and pulmonary hypertension in mice. Pulm. Circ. 2022, 12, e12014. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Hou, Y.; Cai, X. MiR-210-3p Enhances Cardiomyocyte Apoptosis and Mitochondrial Dysfunction by Targeting the NDUFA4 Gene in Sepsis-Induced Myocardial Dysfunction. Int. Heart J. 2021, 62, 636–646. [Google Scholar] [CrossRef] [PubMed]
- Biro, O.; Alasztics, B.; Molvarec, A.; Joo, J.; Nagy, B.; Rigo, J., Jr. Various levels of circulating exosomal total-miRNA and miR-210 hypoxamiR in different forms of pregnancy hypertension. Pregnancy Hypertens. 2017, 10, 207–212. [Google Scholar] [CrossRef]
- Hao, H.; Yan, S.; Zhao, X.; Han, X.; Fang, N.; Zhang, Y.; Dai, C.; Li, W.; Yu, H.; Gao, Y.; et al. Atrial myocyte-derived exosomal microRNA contributes to atrial fibrosis in atrial fibrillation. J. Transl. Med. 2022, 20, 407. [Google Scholar] [CrossRef]
- Virga, F.; Cappellesso, F.; Stijlemans, B.; Henze, A.T.; Trotta, R.; Van Audenaerde, J.; Mirchandani, A.S.; Sanchez-Garcia, M.A.; Vandewalle, J.; Orso, F.; et al. Macrophage miR-210 induction and metabolic reprogramming in response to pathogen interaction boost life-threatening inflammation. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef]
- Ye, H.; Ling, S.; Castillo, A.C.; Thomas, B.; Long, B.; Qian, J.; Perez-Polo, J.R.; Ye, Y.; Chen, X.; Birnbaum, Y. Nebivolol induces distinct changes in profibrosis microRNA expression compared with atenolol, in salt-sensitive hypertensive rats. Hypertension 2013, 61, 1008–1013. [Google Scholar] [CrossRef]
- Solayman, M.H.; Langaee, T.Y.; Gong, Y.; Shahin, M.H.; Turner, S.T.; Chapman, A.B.; Gums, J.G.; Boerwinkle, E.; Beitelshees, A.L.; El-Hamamsy, M.; et al. Effect of plasma MicroRNA on antihypertensive response to beta blockers in the Pharmacogenomic Evaluation of Antihypertensive Responses (PEAR) studies. Eur. J. Pharm. Sci. 2019, 131, 93–98. [Google Scholar] [CrossRef]
- Mone, P.; Lombardi, A.; Kansakar, U.; Varzideh, F.; Jankauskas, S.S.; Pansini, A.; Marzocco, S.; De Gennaro, S.; Famiglietti, M.; Macina, G.; et al. Empagliflozin Improves the MicroRNA Signature of Endothelial Dysfunction in Patients with Heart Failure with Preserved Ejection Fraction and Diabetes. J. Pharmacol. Exp. Ther. 2023, 384, 116–122. [Google Scholar] [CrossRef]
- Qi, H.; Liu, Z.; Liu, B.; Cao, H.; Sun, W.; Yan, Y.; Zhang, L. micro-RNA screening and prediction model construction for diagnosis of salt-sensitive essential hypertension. Medicine 2017, 96, e6417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhao, D. MicroRNA-362-5p promotes the proliferation and inhibits apoptosis of trophoblast cells via targeting glutathione-disulfide reductase. Bioengineered 2021, 12, 2410–2419. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ding, M.; Duan, X.; Feng, X.; Wang, P.; Jiang, Q.; Cheng, Z.; Zhang, W.; Yu, S.; Yao, W.; et al. Diagnostic Value of Plasma MicroRNAs for Lung Cancer Using Support Vector Machine Model. J. Cancer 2019, 10, 5090–5098. [Google Scholar] [CrossRef]
- Su, C.-T.; Yang, C.-H. Feature selection for the SVM: An application to hypertension diagnosis. Expert. Syst. Appl. 2008, 34, 754–763. [Google Scholar] [CrossRef]
- Suzuki, K.; Yamada, H.; Fujii, R.; Munetsuna, E.; Ando, Y.; Ohashi, K.; Ishikawa, H.; Yamazaki, M.; Maeda, K.; Hashimoto, S.; et al. Association between circulating vascular-related microRNAs and an increase in blood pressure: A 5-year longitudinal population-based study. J. Hypertens. 2021, 39, 84–89. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Y.; Chen, G.; Zhou, Y.; Yao, H.; Tan, H. Upregulation of miR-361-3p suppresses serotonin-induced proliferation in human pulmonary artery smooth muscle cells by targeting SERT. Cell Mol. Biol. Lett. 2020, 25, 45. [Google Scholar] [CrossRef]
- Ye, X.; Song, H.; Hu, H.; Zhou, C.; Chen, Q.; Hong, L.; Huang, M.; Zhu, H. MiR-361-3p alleviates cerebral ischemia-reperfusion injury by targeting NACC1 through the PINK1/Parkin pathway. J. Mol. Histol. 2022, 53, 357–367. [Google Scholar] [CrossRef]
- Krist, B.; Florczyk, U.; Pietraszek-Gremplewicz, K.; Jozkowicz, A.; Dulak, J. The Role of miR-378a in Metabolism, Angiogenesis, and Muscle Biology. Int. J. Endocrinol. 2015, 2015, 281756. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, P.; Deng, C. miR-378a-5p regulates CAMKK2/AMPK pathway to contribute to cerebral ischemia/reperfusion injury-induced neuronal apoptosis. Folia Histochem. Cytobiol. 2021, 59, 57–65. [Google Scholar] [CrossRef]
- Wang, B.; Zhao, X.; Xiao, L.; Chen, Y. FoxO1 Silencing Facilitates Neurological Function Recovery in Intracerebral Hemorrhage Mice via the lncRNA GAS5/miR-378a-5p/Hspa5 Axis. J. Stroke Cerebrovasc. Dis. 2022, 31, 106443. [Google Scholar] [CrossRef]
- Webb, A.J.S.; Werring, D.J. New Insights Into Cerebrovascular Pathophysiology and Hypertension. Stroke 2022, 53, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Devaux, Y. MicroRNAs as biomarkers in the brain-heart axis? Eur. Heart J. Acute Cardiovasc. Care 2022, 11, 617–619. [Google Scholar] [CrossRef] [PubMed]
- Miao, R.; Dong, X.B.; Gong, J.N.; Li, J.F.; Pang, W.Y.; Liu, Y.Y.; Yang, Y.H. Analysis of significant microRNA associated with chronic thromboembolic pulmonary hypertension. Zhonghua Yi Xue Za Zhi 2018, 98, 1397–1402. [Google Scholar] [CrossRef] [PubMed]
- Iacomino, G.; Russo, P.; Marena, P.; Lauria, F.; Venezia, A.; Ahrens, W.; De Henauw, S.; De Luca, P.; Foraita, R.; Gunther, K.; et al. Circulating microRNAs are associated with early childhood obesity: Results of the I.Family Study. Genes. Nutr. 2019, 14, 2. [Google Scholar] [CrossRef]
- de Gonzalo-Calvo, D.; Perez-Boza, J.; Curado, J.; Devaux, Y.; EU-CardioRNA COST Action CA17129. Challenges of microRNA-based biomarkers in clinical application for cardiovascular diseases. Clin. Transl. Med. 2022, 12, e585. [Google Scholar] [CrossRef]
- Lakkisto, P.; Dalgaard, L.T.; Belmonte, T.; Pinto-Sietsma, S.J.; Devaux, Y.; de Gonzalo-Calvo, D.; CA, E.U.-C.C.A. Development of circulating microRNA-based biomarkers for medical decision-making: A friendly reminder of what should NOT be done. Crit. Rev. Clin. Lab. Sci. 2023, 60, 141–152. [Google Scholar] [CrossRef]
- Devaux, Y.; Salgado-Somoza, A.; Dankiewicz, J.; Boileau, A.; Stammet, P.; Schritz, A.; Zhang, L.; Vausort, M.; Gilje, P.; Erlinge, D.; et al. Incremental Value of Circulating MiR-122-5p to Predict Outcome after Out of Hospital Cardiac Arrest. Theranostics 2017, 7, 2555–2564. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
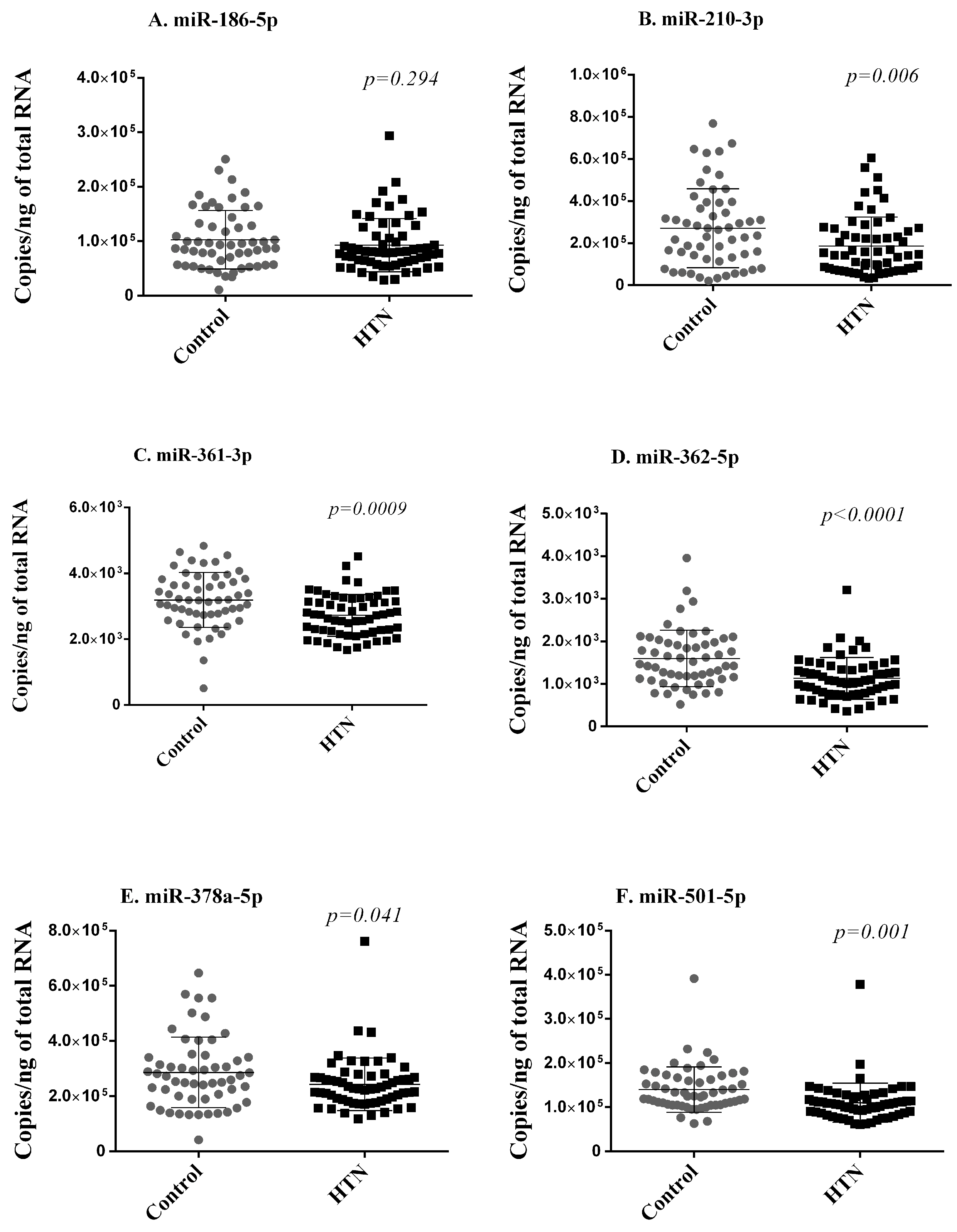{kind=link}
| Clinical Characteristics | Discovery Cohort (n = 60) | Validation Cohort (n = 114) | ||||
|---|---|---|---|---|---|---|
| Control Group (n = 30) | Hypertension Group (n = 30) | p | Control Group (n = 55) | Hypertension Group (n = 59) | p | |
| Mean age | 45.6 ± 13.167 | 45.7 ± 13.02 | 0.955 | 43.91 ± 15.44 | 48.57 ± 12.72 | 0.079 |
| Gender | ||||||
| Males, N (%) | 15 (50%) | 15 (50%) | - | 24 (43.64%) | 35 (59.32%) | 0.133 |
| Females, N (%) | 15 (50%) | 15 (50%) | - | 31 (56.36%) | 24 (40.68%) | |
| BMI (kg/m2) | 25.25 ± 2.238 | 30.52 ± 3.377 | 0.0001 | 25.27 ± 2.95 | 28.93 ± 3.14 | 0.0001 |
| Blood pressure | ||||||
| Systolic blood pressure (mmHg) | 120.43 ± 5.67 | 147.8 ± 13.67 | 0.0001 | 122.2 ± 7.91 | 150.5 ± 14.39 | 0.0001 |
| Diastolic blood pressure (mmHg) | 77.57 ± 6.24 | 98.33 ± 6.73 | 0.0001 | 78.0 ± 6.55 | 97.42 ± 5.38 | 0.0001 |
| Hypertension treatment | ||||||
| Untreated, N (%) | - | 6 (20%) | - | - | 14 (23.73%) | - |
| Treated, N (%) | 24 (80%) | - | - | 45 (76.27%) | ||
| Anti-hypertension therapy | ||||||
| No therapy, N (%) | - | 6 (20%) | - | - | 14 (23.73%) | - |
| Monotherapy, N (%) | - | 19 (63.33%) | - | - | 37 (62.71%) | |
| Combined therapy, N (%) | 5 (16.67%) | - | - | 8 (13.56%) | ||
| Complications caused by hypertension | ||||||
| No complications, N (%) | - | 27 (90%) | - | - | 54 (91.53%) | - |
| With complications, N (%) | 3 (10%) | - | - | 5 (8.47%) | ||
| Diabetes mellitus | ||||||
| Non-diabetic patients, N (%) | - | - | - | - | 50 (84.75%) | - |
| Diabetic patients, N (%) | - | - | - | - | 8 (13.56%) | - |
| Hospitalization due to uncontrolled hypertension, N (%) | - | - | - | - | 1 (1.69%) | - |
| Smoking status | ||||||
| Current smokers, N (%) | 5 (16.66%) | 17 (56.66%) | 0.005 | 13 (23.64%) | 27 (45.76%) | 0.003 |
| Former smokers, N (%) | 6 (20.00%) | 3 (10.00%) | 4 (7.27%) | 10 (16.95%) | ||
| Non-smokers, N (%) | 19 (63.33%) | 10 (33.33% | 38 (69.09%) | 22 (37.29%) | ||
| Alcohol consumption, N (%) | 5 (16.66%) | 12 (40.00%) | 0.084 | 39 (70.91%) | 25 (42.37%) | 0.002 |
| Hypertension family history | ||||||
| Positive family history | 13 (43.33%) | 24 (80%) | 0.010 | 16 (29.09%) | 45 (76.27%) | 0.999 |
| Negative family history | 15 (50%) | 6 (20% | 37 (67.27%) | 14 (23.73%) | ||
| Unknown family history | 2 (6.67%) | 0 | 2 (3.64%) | 0 | ||
| Mean of Normalized Counts | |||||
|---|---|---|---|---|---|
| miRNA | Control Group (n = 30) | Hypertension Group (n = 30) | Log2 FC | p | FDR |
| hsa-miR-186-5p | 9620 | 7314 | −0.410 | 0.0013 | 0.043 |
| hsa-miR-210-3p | 130 | 81 | −0.653 | 0.0012 | 0.041 |
| hsa-miR-361-3p | 3917 | 3113 | −0.344 | 0.0002 | 0.028 |
| hsa-miR-362-5p | 450 | 317 | −0.494 | 0.0002 | 0.028 |
| hsa-miR-378a-5p | 349 | 248 | −0.515 | 0.00005 | 0.018 |
| hsa-miR-501-5p | 26 | 18 | −0.503 | 0.0008 | 0.038 |
| hsa-miR-769-5p | 47 | 30 | −0.616 | 0.00003 | 0.015 |
| AUC | Balanced Accuracy | F1 (Hypertension) | Precision (Hypertension) | Sensitivity (Hypertension) | Specificity (Hypertension) | |
|---|---|---|---|---|---|---|
| Test dataset | 0.90 | 0.87 | 0.87 | 0.91 | 0.83 | 0.91 |
| LOOCV | 0.89 | 0.83 | 0.83 | 0.87 | 0.80 | 0.86 |
| AUC | Balanced Accuracy | F1 (Hypertension) | Precision (Hypertension) | Sensitivity (Hypertension) | Specificity (Hypertension) | |
|---|---|---|---|---|---|---|
| Test dataset | 0.89 | 0.87 | 0.87 | 0.91 | 0.83 | 0.91 |
| LOOCV | 0.87 | 0.79 | 0.80 | 0.81 | 0.79 | 0.80 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jusic, A.; Junuzovic, I.; Hujdurovic, A.; Zhang, L.; Vausort, M.; Devaux, Y. A Machine Learning Model Based on microRNAs for the Diagnosis of Essential Hypertension. Non-Coding RNA 2023, 9, 64. https://doi.org/10.3390/ncrna9060064
Jusic A, Junuzovic I, Hujdurovic A, Zhang L, Vausort M, Devaux Y. A Machine Learning Model Based on microRNAs for the Diagnosis of Essential Hypertension. Non-Coding RNA. 2023; 9(6):64. https://doi.org/10.3390/ncrna9060064
Chicago/Turabian StyleJusic, Amela, Inela Junuzovic, Ahmed Hujdurovic, Lu Zhang, Mélanie Vausort, and Yvan Devaux. 2023. "A Machine Learning Model Based on microRNAs for the Diagnosis of Essential Hypertension" Non-Coding RNA 9, no. 6: 64. https://doi.org/10.3390/ncrna9060064
APA StyleJusic, A., Junuzovic, I., Hujdurovic, A., Zhang, L., Vausort, M., & Devaux, Y. (2023). A Machine Learning Model Based on microRNAs for the Diagnosis of Essential Hypertension. Non-Coding RNA, 9(6), 64. https://doi.org/10.3390/ncrna9060064