


Age and 17β-Estradiol (E2) Facilitate Nuclear Export and Argonaute Loading of microRNAs in the Female Brain

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
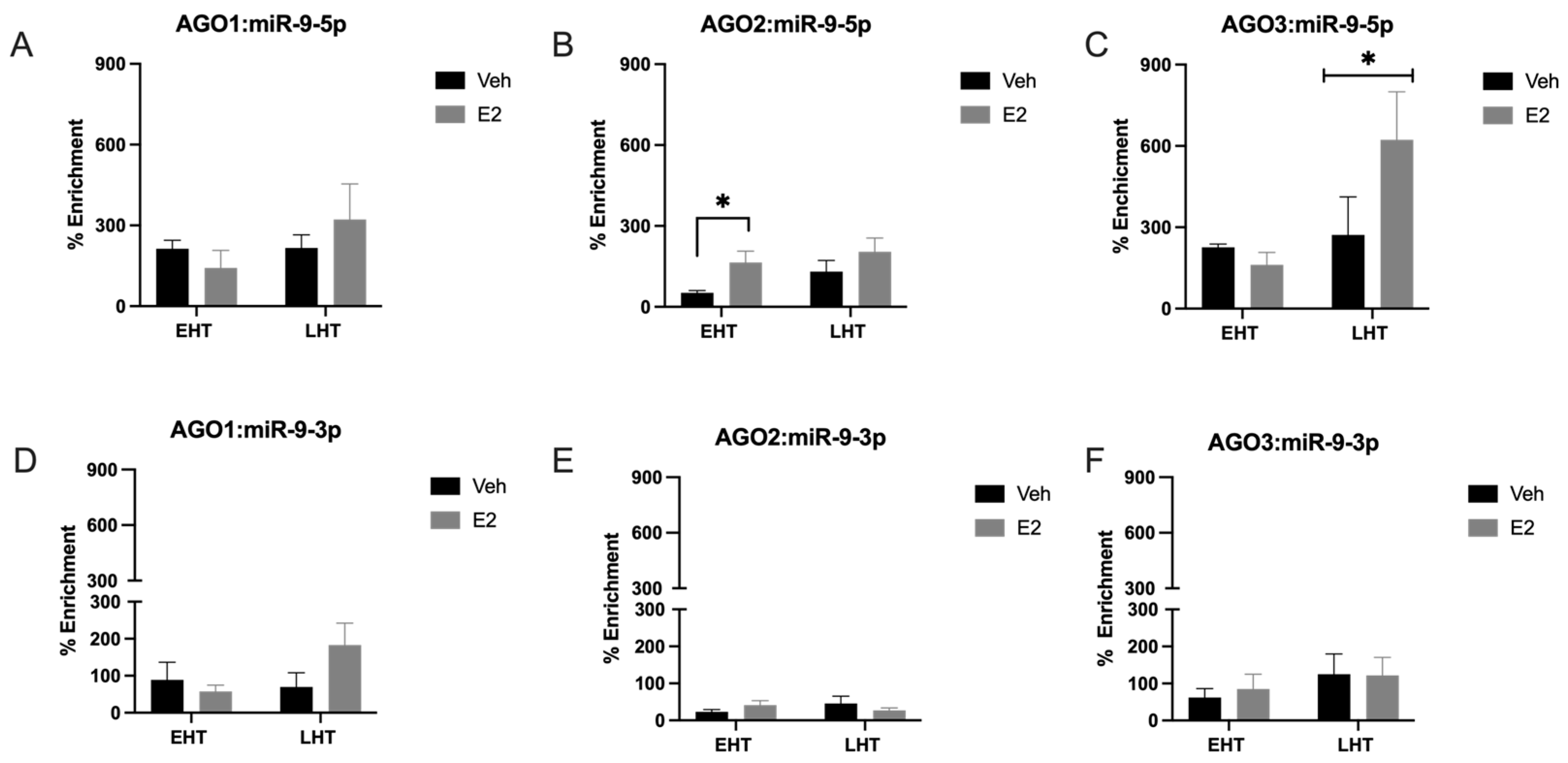
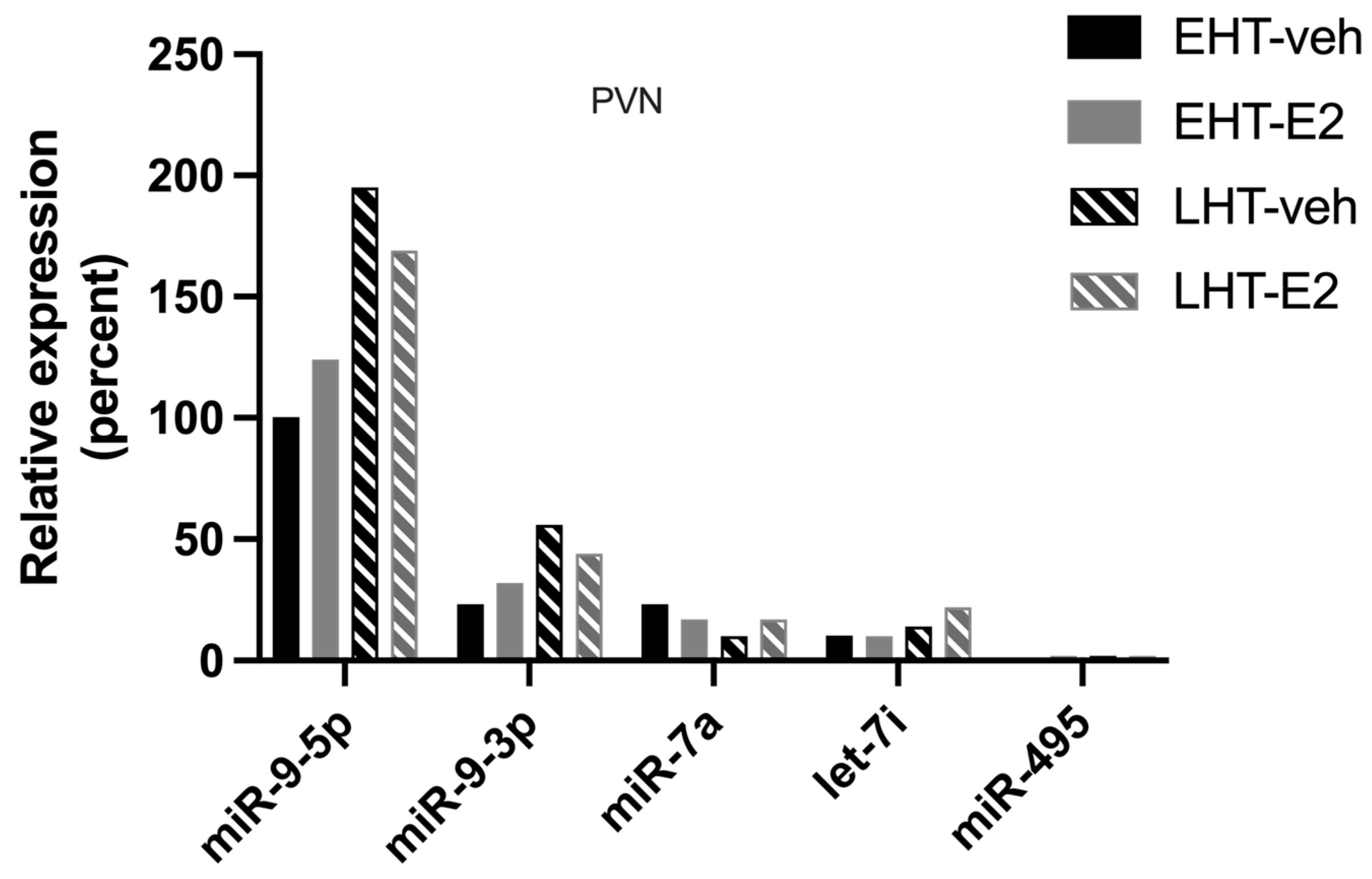
2.1. The Length of Time Following Ovarian Hormone Deprivation Altered Which AGO Protein Was Most Associated with miR-9-5p in the Paraventricular Nucleus (PVN) of the Hypothalamus
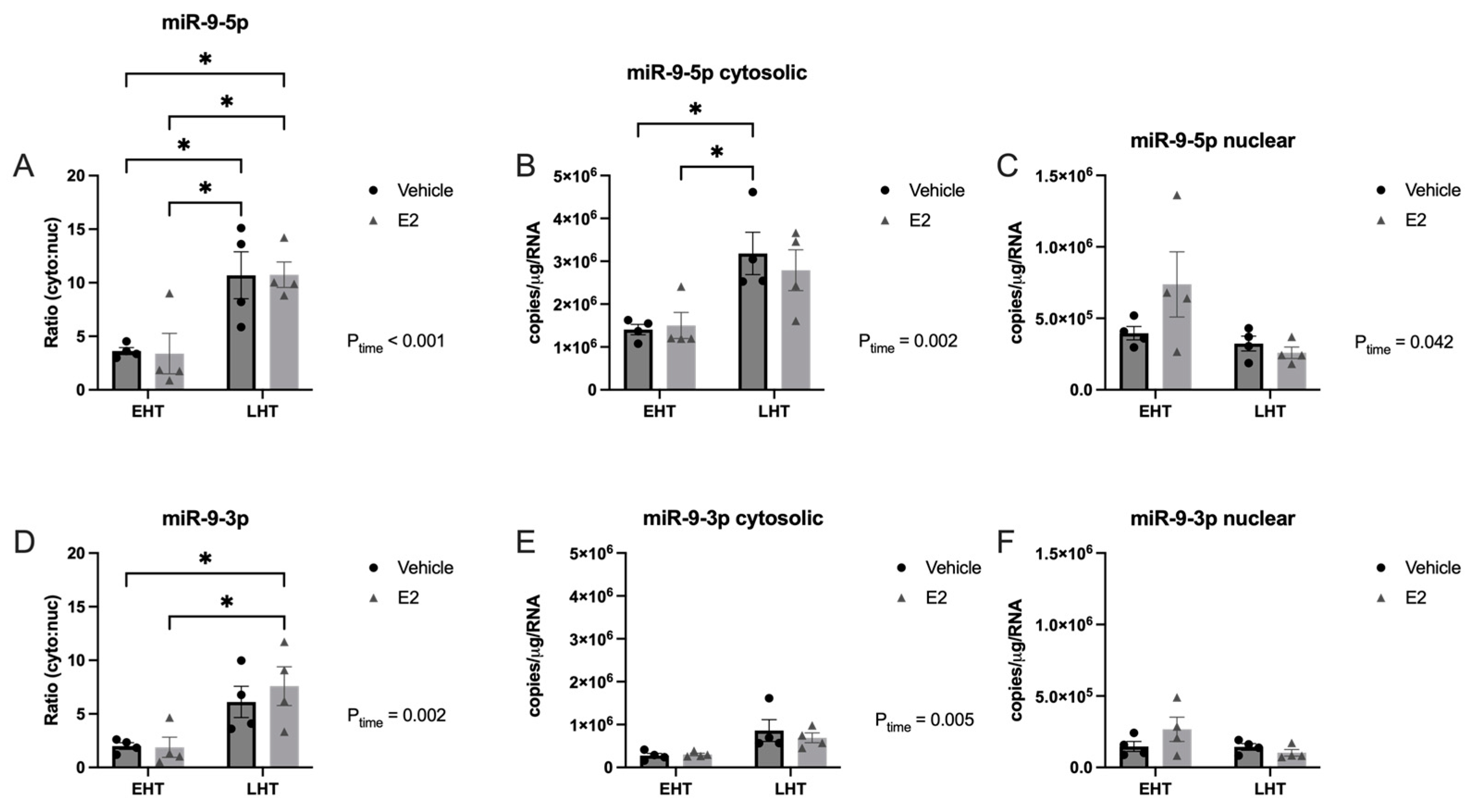
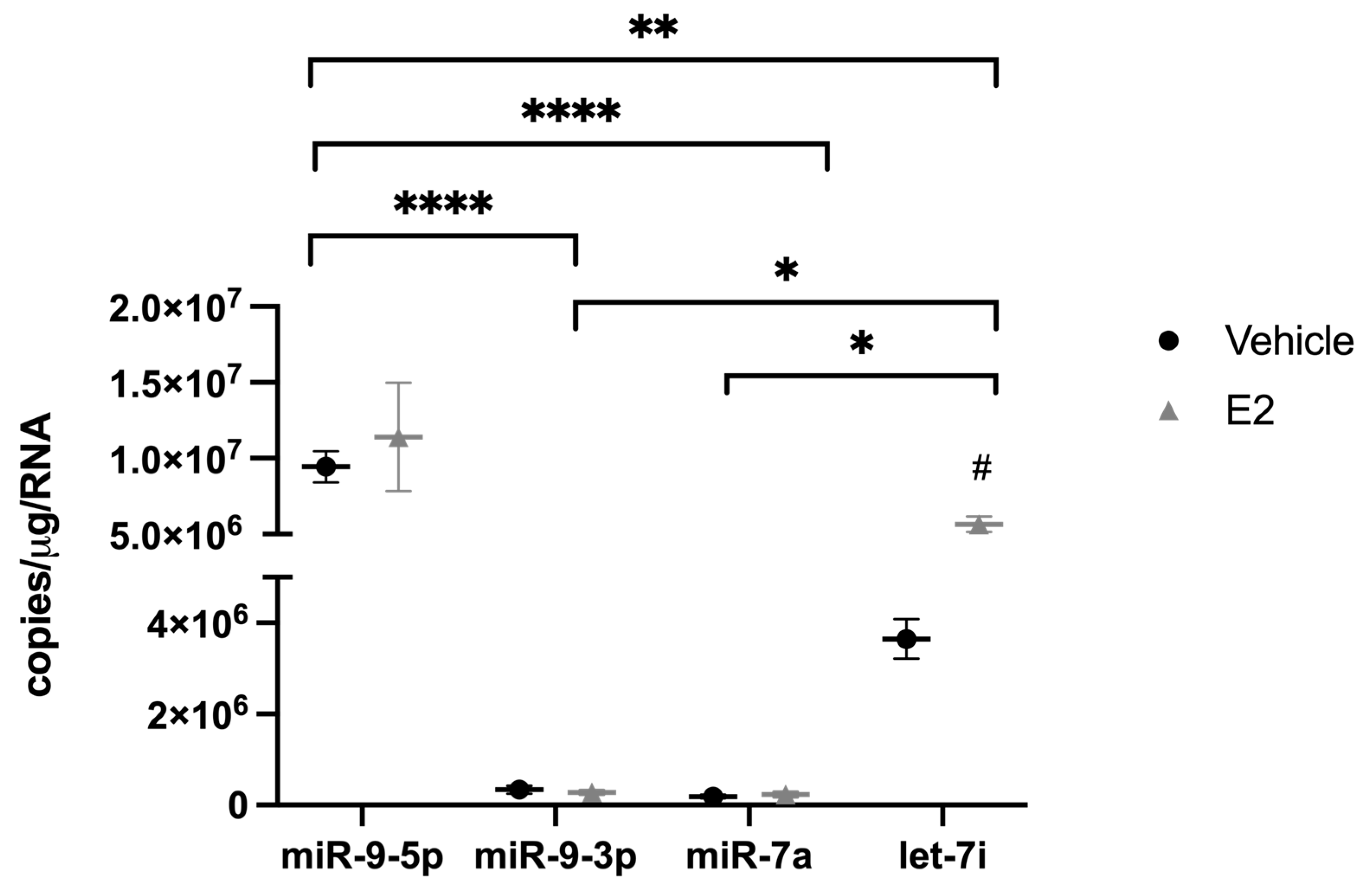
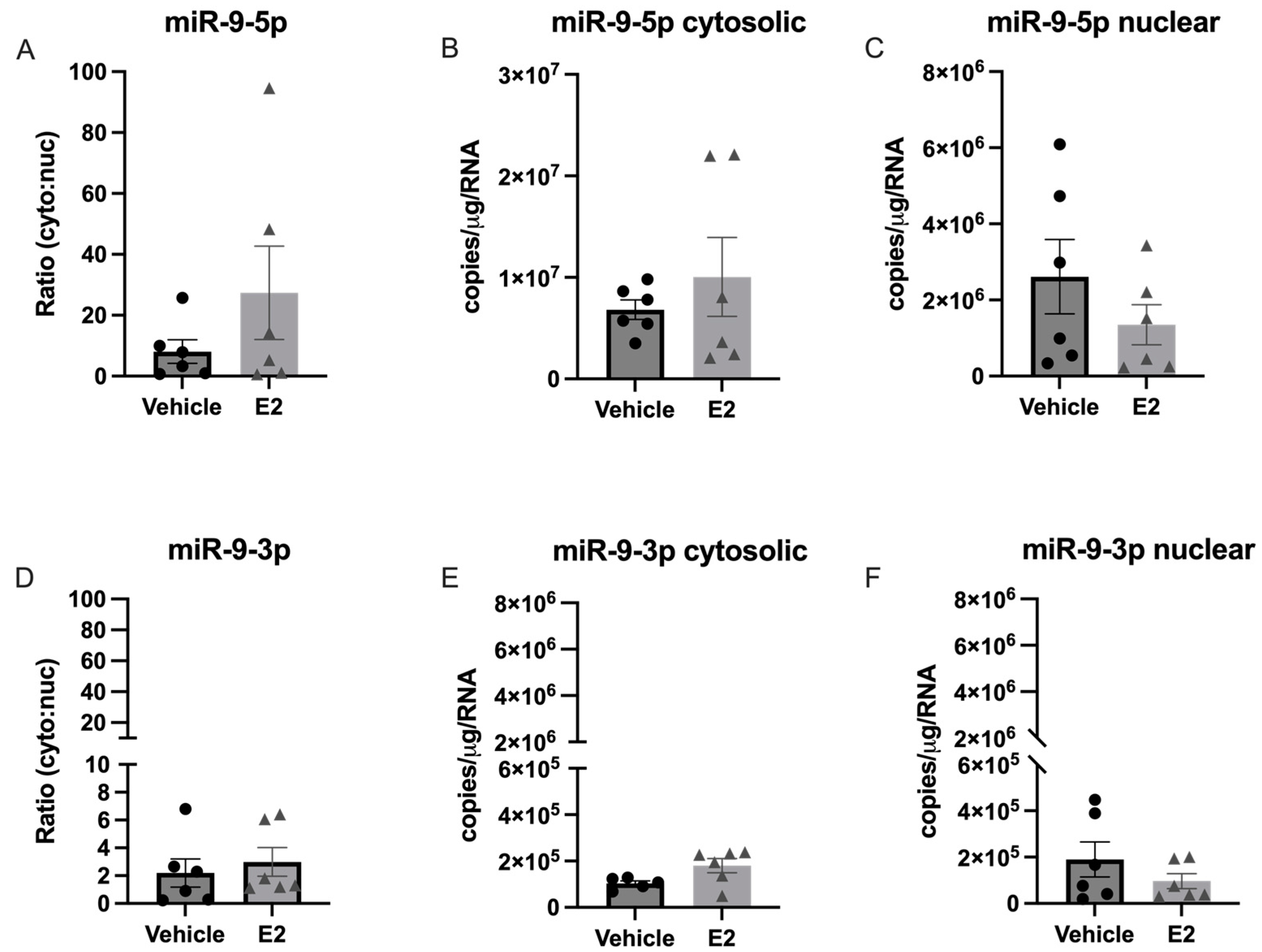
2.2. The Length of Time Following Ovarian Hormone Deprivation Altered Subcellular miR Localization in PVN-Derived Neuronal Cells
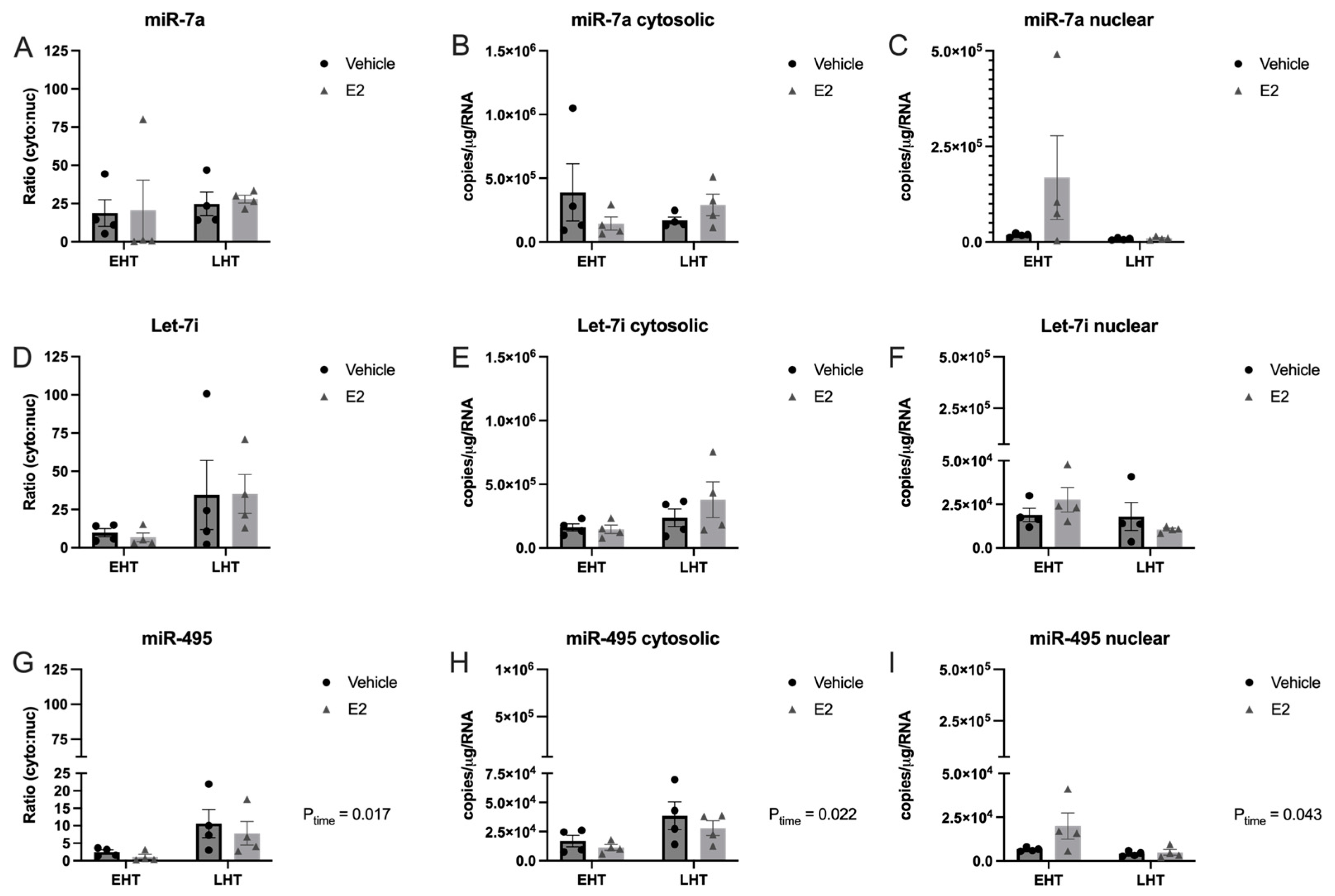
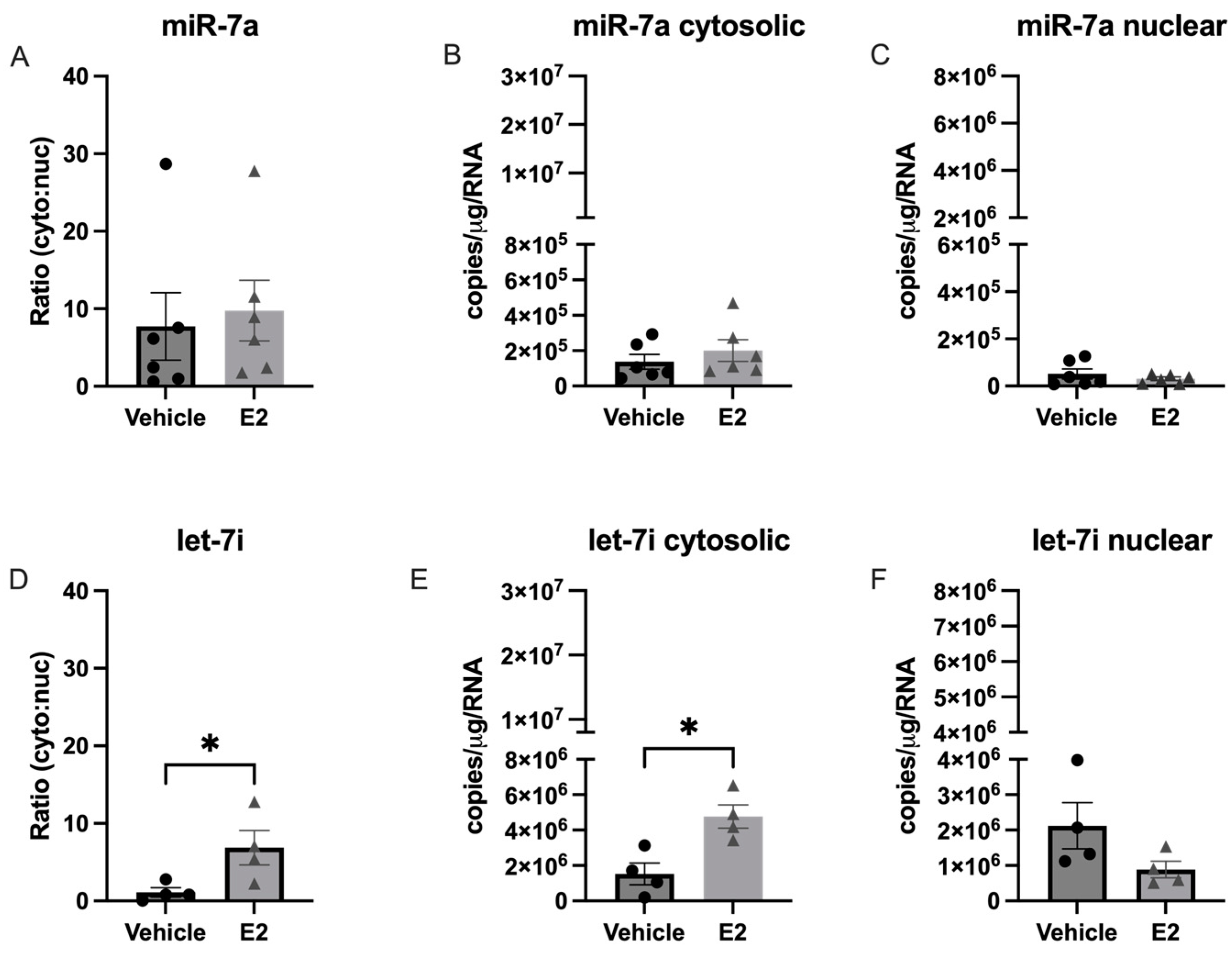
2.3. E2 Treatment Altered the Subcellular Localization of let-7i in a Hypothalamic Neuronal Cell Line Derived from the Paraventricular Nucleus (PVN) of the Hypothalamus
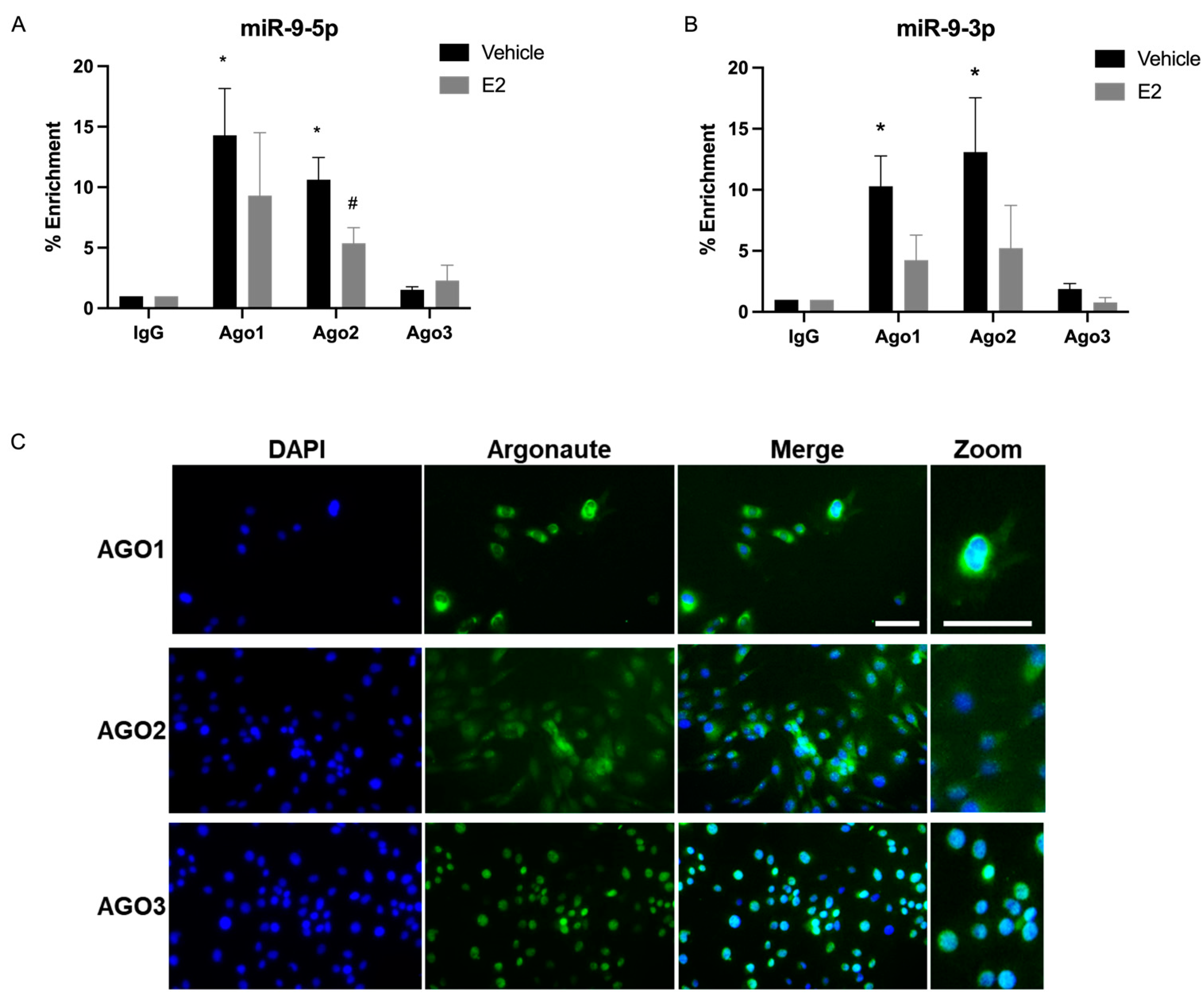
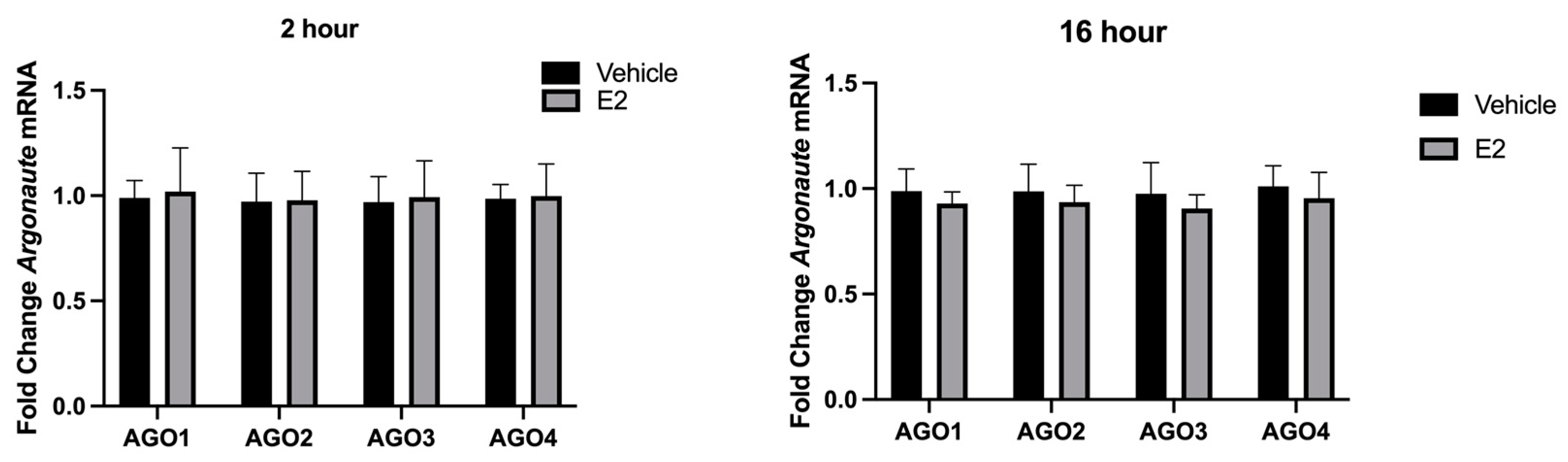
2.4. E2 Treatment Decreased miRNA Association with AGO1 and AGO2 in the Nucleus in PVN-Derived Neuronal Cells
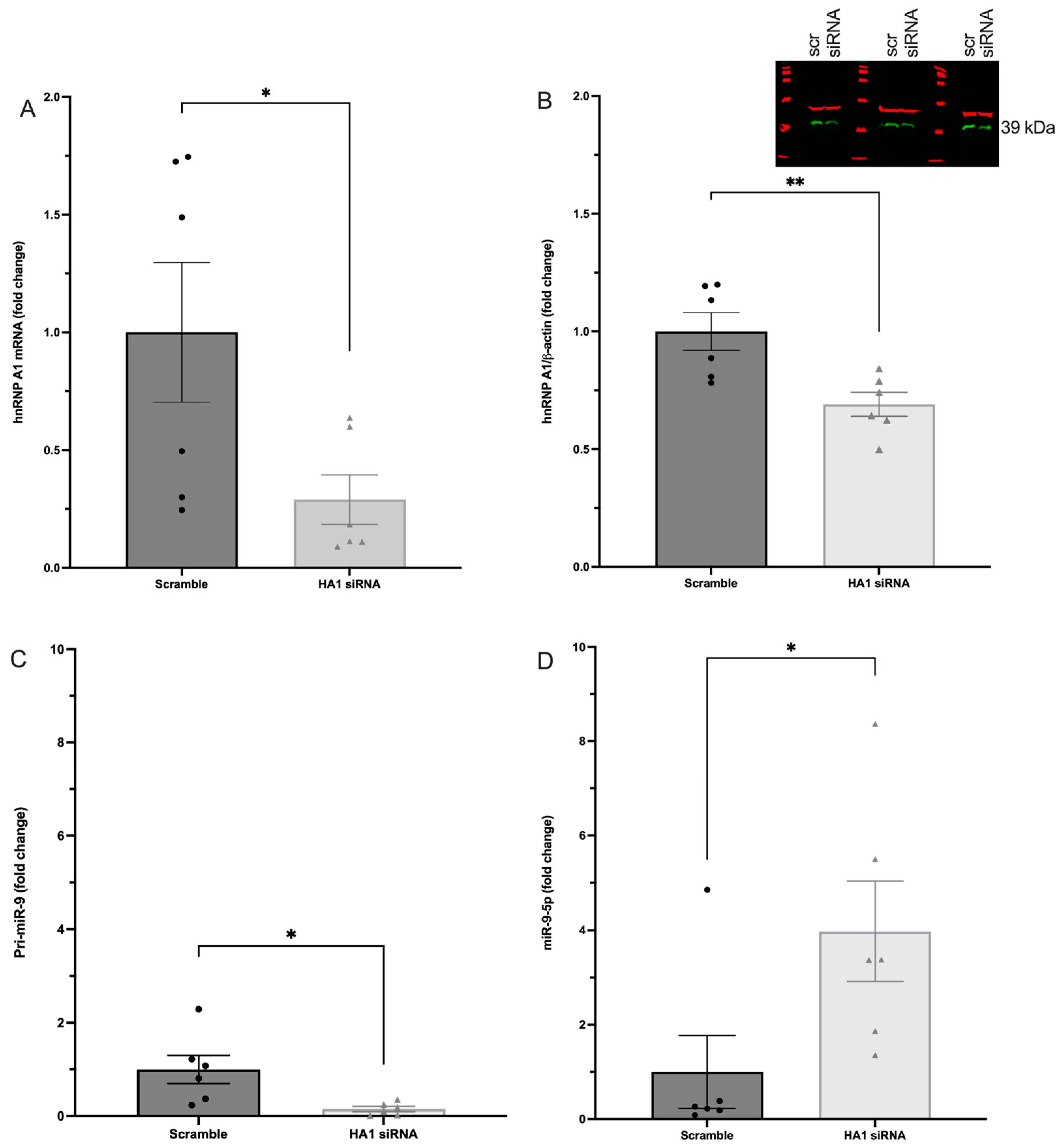
2.5. hnRNP A1 Decreased Mature miR-9-5p Expression in PVN-Derived Neuronal Cells
3. Discussion
3.1. Summary of Major Findings
3.2. Length of Ovarian Hormone Deprivation Regulates mi-9-5p AGO Switching in the Aged Female Brain
3.3. miR-9-5p and miR-9-3p Preferentially Associate with Cytoplasmic, and Not Nuclear, AGO3 in a Hypothalamic-Derived Neuronal Cell Line
3.4. Length of Ovarian Hormone Deprivation Alters miRNA Subcellular Localization in the Aged Female Brain
3.5. The RNA Binding Protein, hnRNP A1, Is an Important Regulator of miR-9 Expression in Hypothalamic PVN-Derived Neurons
4. Materials and Methods
4.1. Animals
4.1.1. Ovariectomy
4.1.2. Tissue Processing
4.2. Cell Culture
4.3. Molecular Biology Assays
4.3.1. RNA Isolation and cDNA Synthesis
4.3.2. Reverse Transcription Quantitative PCR (RT-qPCR)
4.3.3. RNA Immunoprecipitation (RIP)
4.3.4. Nuclear RNA Immunoprecipitation (nRIP)
4.3.5. siRNA Knockdown of hnRNP A1
4.4. Protein Assays
4.4.1. Western Blot
4.4.2. Immunofluorescence
4.5. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Kai, Z.S.; Pasquinelli, A.E. MicroRNA assassins: Factors that regulate the disappearance of miRNAs. Nat. Struct. Mol. Biol. 2010, 17, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.S.; Mott, N.N.; Wang, Y.; Chung, W.C.; Pak, T.R. MicroRNAs in the aging female brain: A putative mechanism for age-specific estrogen effects. Endocrinology 2013, 154, 2795–2806. [Google Scholar] [CrossRef]
- Rao, Y.S.; Pak, T.R. microRNAs and the adolescent brain: Filling the knowledge gap. Neurosci. Biobehav. Rev. 2016, 70, 313–322. [Google Scholar] [CrossRef]
- Kim, C.K.; Linscott, M.L.; Flury, S.; Zhang, M.; Newby, M.L.; Pak, T.R. 17beta-Estradiol Regulates miR-9-5p and miR-9-3p Stability and Function in the Aged Female Rat Brain. Noncoding RNA 2021, 7, 53. [Google Scholar] [CrossRef]
- Rao, Y.S.; Shults, C.L.; Pinceti, E.; Pak, T.R. Prolonged ovarian hormone deprivation alters the effects of 17β-estradiol on microRNA expression in the aged female rat hypothalamus. Oncotarget 2015, 6, 36965–36983. [Google Scholar] [CrossRef]
- Kim, C.K.; Asimes, A.; Zhang, M.; Son, B.T.; Kirk, J.A.; Pak, T.R. Differential Stability of miR-9-5p and miR-9-3p in the Brain Is Determined by Their Unique Cis- and Trans-Acting Elements. eNeuro 2020, 7. [Google Scholar] [CrossRef]
- Trabucchi, M.; Mategot, R. Subcellular Heterogeneity of the microRNA Machinery. Trends Genet. 2019, 35, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef] [PubMed]
- Yuva-Aydemir, Y.; Simkin, A.; Gascon, E.; Gao, F.B. MicroRNA-9: Functional evolution of a conserved small regulatory RNA. RNA Biol. 2011, 8, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Griffin, K.N.; Walters, B.W.; Li, H.; Wang, H.; Biancon, G.; Tebaldi, T.; Kaya, C.B.; Kanyo, J.; Lam, T.T.; Cox, A.L.; et al. Widespread association of the Argonaute protein AGO2 with meiotic chromatin suggests a distinct nuclear function in mammalian male reproduction. Genome Res. 2022, 32, 1655–1668. [Google Scholar] [CrossRef]
- Chu, Y.; Yokota, S.; Liu, J.; Kilikevicius, A.; Johnson, K.C.; Corey, D.R. Argonaute binding within human nuclear RNA and its impact on alternative splicing. RNA 2021, 27, 991–1003. [Google Scholar] [CrossRef]
- Janowski, B.A.; Huffman, K.E.; Schwartz, J.C.; Ram, R.; Nordsell, R.; Shames, D.S.; Minna, J.D.; Corey, D.R. Involvement of AGO1 and AGO2 in mammalian transcriptional silencing. Nat. Struct. Mol. Biol. 2006, 13, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Ameyar-Zazoua, M.; Rachez, C.; Souidi, M.; Robin, P.; Fritsch, L.; Young, R.; Morozova, N.; Fenouil, R.; Descostes, N.; Andrau, J.C.; et al. Argonaute proteins couple chromatin silencing to alternative splicing. Nat. Struct. Mol. Biol. 2012, 19, 998–1004. [Google Scholar] [CrossRef]
- Allo, M.; Agirre, E.; Bessonov, S.; Bertucci, P.; Gomez Acuna, L.; Buggiano, V.; Bellora, N.; Singh, B.; Petrillo, E.; Blaustein, M.; et al. Argonaute-1 binds transcriptional enhancers and controls constitutive and alternative splicing in human cells. Proc. Natl. Acad. Sci. USA 2014, 111, 15622–15629. [Google Scholar] [CrossRef]
- Kooshapur, H.; Choudhury, N.R.; Simon, B.; Muhlbauer, M.; Jussupow, A.; Fernandez, N.; Jones, A.N.; Dallmann, A.; Gabel, F.; Camilloni, C.; et al. Structural basis for terminal loop recognition and stimulation of pri-miRNA-18a processing by hnRNP A1. Nat. Commun. 2018, 9, 2479. [Google Scholar] [CrossRef]
- Michlewski, G.; Caceres, J.F. Antagonistic role of hnRNP A1 and KSRP in the regulation of let-7a biogenesis. Nat. Struct. Mol. Biol. 2010, 17, 1011–1018. [Google Scholar] [CrossRef]
- Michlewski, G.; Guil, S.; Caceres, J.F. Stimulation of pri-miR-18a processing by hnRNP A1. Adv. Exp. Med. Biol. 2010, 700, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Naqvi, A.; Hendriks, G.J.; Feltzin, V.; Zhu, Y.; Grigoriev, A.; Bonini, N.M. Impact of age-associated increase in 2’-O-methylation of miRNAs on aging and neurodegeneration in Drosophila. Genes. Dev. 2014, 28, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, J.; Kim, K.; Chang, H.; You, K.; Kim, V.N. Bias-minimized quantification of microRNA reveals widespread alternative processing and 3’ end modification. Nucleic Acids Res. 2019, 47, 2630–2640. [Google Scholar] [CrossRef] [PubMed]
- Park, M.S.; Phan, H.D.; Busch, F.; Hinckley, S.H.; Brackbill, J.A.; Wysocki, V.H.; Nakanishi, K. Human Argonaute3 has slicer activity. Nucleic Acids Res. 2017, 45, 11867–11877. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.; Diederichs, S. Argonaute-3 activates the let-7a passenger strand microRNA. RNA Biol. 2013, 10, 1631–1643. [Google Scholar] [CrossRef] [PubMed]
- Azuma-Mukai, A.; Oguri, H.; Mituyama, T.; Qian, Z.R.; Asai, K.; Siomi, H.; Siomi, M.C. Characterization of endogenous human Argonautes and their miRNA partners in RNA silencing. Proc. Natl. Acad. Sci. USA 2008, 105, 7964–7969. [Google Scholar] [CrossRef] [PubMed]
- Burroughs, A.M.; Ando, Y.; de Hoon, M.J.; Tomaru, Y.; Suzuki, H.; Hayashizaki, Y.; Daub, C.O. Deep-sequencing of human Argonaute-associated small RNAs provides insight into miRNA sorting and reveals Argonaute association with RNA fragments of diverse origin. RNA Biol. 2011, 8, 158–177. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, A.; Burwinkel, B. Distinct AGO1 and AGO2 associated miRNA profiles in human cells and blood plasma. RNA Biol. 2012, 9, 1066–1075. [Google Scholar] [CrossRef]
- Bofill-De Ros, X.; Kasprzak, W.K.; Bhandari, Y.; Fan, L.; Cavanaugh, Q.; Jiang, M.; Dai, L.; Yang, A.; Shao, T.J.; Shapiro, B.A.; et al. Structural Differences between Pri-miRNA Paralogs Promote Alternative Drosha Cleavage and Expand Target Repertoires. Cell Rep. 2019, 26, 447–459 e444. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, J.; Koppetsch, B.S.; Wang, J.; Tipping, C.; Ma, S.; Weng, Z.; Theurkauf, W.E.; Zamore, P.D. Heterotypic piRNA Ping-Pong requires qin, a protein with both E3 ligase and Tudor domains. Mol. Cell 2011, 44, 572–584. [Google Scholar] [CrossRef]
- Webster, A.; Li, S.; Hur, J.K.; Wachsmuth, M.; Bois, J.S.; Perkins, E.M.; Patel, D.J.; Aravin, A.A. Aub and Ago3 Are Recruited to Nuage through Two Mechanisms to Form a Ping-Pong Complex Assembled by Krimper. Mol. Cell 2015, 59, 564–575. [Google Scholar] [CrossRef]
- Kalantari, R.; Hicks, J.A.; Li, L.; Gagnon, K.T.; Sridhara, V.; Lemoff, A.; Mirzaei, H.; Corey, D.R. Stable association of RNAi machinery is conserved between the cytoplasm and nucleus of human cells. RNA 2016, 22, 1085–1098. [Google Scholar] [CrossRef] [PubMed]
- Meister, G.; Landthaler, M.; Patkaniowska, A.; Dorsett, Y.; Teng, G.; Tuschl, T. Human Argonaute2 mediates RNA cleavage targeted by miRNAs and siRNAs. Mol. Cell 2004, 15, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Jie, M.; Feng, T.; Huang, W.; Zhang, M.; Feng, Y.; Jiang, H.; Wen, Z. Subcellular Localization of miRNAs and Implications in Cellular Homeostasis. Genes 2021, 12, 856. [Google Scholar] [CrossRef] [PubMed]
- Koester, S.K.; Dougherty, J.D. A Proposed Role for Interactions between Argonautes, miRISC, and RNA Binding Proteins in the Regulation of Local Translation in Neurons and Glia. J. Neurosci. 2022, 42, 3291–3301. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Saetrom, P.; Snove, O., Jr.; Rossi, J.J. MicroRNA-directed transcriptional gene silencing in mammalian cells. Proc. Natl. Acad. Sci. USA 2008, 105, 16230–16235. [Google Scholar] [CrossRef] [PubMed]
- Cernilogar, F.M.; Onorati, M.C.; Kothe, G.O.; Burroughs, A.M.; Parsi, K.M.; Breiling, A.; Lo Sardo, F.; Saxena, A.; Miyoshi, K.; Siomi, H.; et al. Chromatin-associated RNA interference components contribute to transcriptional regulation in Drosophila. Nature 2011, 480, 391–395. [Google Scholar] [CrossRef]
- Zhang, Y.; Fan, M.; Zhang, X.; Huang, F.; Wu, K.; Zhang, J.; Liu, J.; Huang, Z.; Luo, H.; Tao, L.; et al. Cellular microRNAs up-regulate transcription via interaction with promoter TATA-box motifs. RNA 2014, 20, 1878–1889. [Google Scholar] [CrossRef] [PubMed]
- Noren Hooten, N.; Martin-Montalvo, A.; Dluzen, D.F.; Zhang, Y.; Bernier, M.; Zonderman, A.B.; Becker, K.G.; Gorospe, M.; de Cabo, R.; Evans, M.K. Metformin-mediated increase in DICER1 regulates microRNA expression and cellular senescence. Aging Cell 2016, 15, 572–581. [Google Scholar] [CrossRef]
- Ohno, S.I.; Oikawa, K.; Tsurui, T.; Harada, Y.; Ono, K.; Tateishi, M.; Mirza, A.; Takanashi, M.; Kanekura, K.; Nagase, K.; et al. Nuclear microRNAs release paused Pol II via the DDX21-CDK9 complex. Cell Rep. 2022, 39, 110673. [Google Scholar] [CrossRef]
- Clarke, J.P.; Thibault, P.A.; Salapa, H.E.; Levin, M.C. A Comprehensive Analysis of the Role of hnRNP A1 Function and Dysfunction in the Pathogenesis of Neurodegenerative Disease. Front. Mol. Biosci. 2021, 8, 659610. [Google Scholar] [CrossRef] [PubMed]
- Kamma, H.; Portman, D.S.; Dreyfuss, G. Cell type-specific expression of hnRNP proteins. Exp. Cell Res. 1995, 221, 187–196. [Google Scholar] [CrossRef]
- Jones, A.N.; Walbrun, A.; Falleroni, F.; Rief, M.; Sattler, M. Conformational Effects of a Cancer-Linked Mutation in Pri-miR-30c RNA. J. Mol. Biol. 2022, 434, 167705. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T.; Kuwano, Y.; Takahara, Y.; Nishida, K.; Rokutan, K. HnRNPA1 interacts with G-quadruplex in the TRA2B promoter and stimulates its transcription in human colon cancer cells. Sci. Rep. 2019, 9, 10276. [Google Scholar] [CrossRef] [PubMed]
- Lemieux, B.; Blanchette, M.; Monette, A.; Mouland, A.J.; Wellinger, R.J.; Chabot, B. A Function for the hnRNP A1/A2 Proteins in Transcription Elongation. PLoS ONE 2015, 10, e0126654. [Google Scholar] [CrossRef] [PubMed]
- Paramasivam, M.; Membrino, A.; Cogoi, S.; Fukuda, H.; Nakagama, H.; Xodo, L.E. Protein hnRNP A1 and its derivative Up1 unfold quadruplex DNA in the human KRAS promoter: Implications for transcription. Nucleic Acids Res. 2009, 37, 2841–2853. [Google Scholar] [CrossRef] [PubMed]
- Michlewski, G.; Guil, S.; Semple, C.A.; Caceres, J.F. Posttranscriptional regulation of miRNAs harboring conserved terminal loops. Mol. Cell 2008, 32, 383–393. [Google Scholar] [CrossRef]
- Guil, S.; Caceres, J.F. The multifunctional RNA-binding protein hnRNP A1 is required for processing of miR-18a. Nat. Struct. Mol. Biol. 2007, 14, 591–596. [Google Scholar] [CrossRef]
- Zhang, M.; Flury, S.; Kim, C.K.; Chung, W.C.J.; Kirk, J.A.; Pak, T.R. Absolute quantification of phosphorylated ERβ amino acid residues in the hippocampus of women and in a rat model of menopause. Endocrinology 2021, 162, bqab122. [Google Scholar] [CrossRef]
- Schmidt, G.; Andersson, S.B.; Nordle, O.; Johansson, C.J.; Gunnarsson, P.O. Release of 17-beta-oestradiol from a vaginal ring in postmenopausal women: Pharmacokinetic evaluation. Gynecol. Obstet. Investig. 1994, 38, 253–260. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Hard Cover Edition; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Schmittgen, T.D.; Zakrajsek, B.A. Effect of experimental treatment on housekeeping gene expression: Validation by real-time, quantitative RT-PCR. J. Biochem. Biophys. Methods 2000, 46, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Shults, C.L.; Dingwall, C.B.; Kim, C.K.; Pinceti, E.; Rao, Y.S.; Pak, T.R. 17beta-estradiol regulates the RNA-binding protein Nova1, which then regulates the alternative splicing of estrogen receptor beta in the aging female rat brain. Neurobiol. Aging 2018, 61, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linscott, M.L.; Yildiz, Y.; Flury, S.; Newby, M.L.; Pak, T.R. Age and 17β-Estradiol (E2) Facilitate Nuclear Export and Argonaute Loading of microRNAs in the Female Brain. Non-Coding RNA 2023, 9, 74. https://doi.org/10.3390/ncrna9060074
Linscott ML, Yildiz Y, Flury S, Newby ML, Pak TR. Age and 17β-Estradiol (E2) Facilitate Nuclear Export and Argonaute Loading of microRNAs in the Female Brain. Non-Coding RNA. 2023; 9(6):74. https://doi.org/10.3390/ncrna9060074
Chicago/Turabian StyleLinscott, Megan L., Yoldas Yildiz, Sarah Flury, Mikayla L. Newby, and Toni R. Pak. 2023. "Age and 17β-Estradiol (E2) Facilitate Nuclear Export and Argonaute Loading of microRNAs in the Female Brain" Non-Coding RNA 9, no. 6: 74. https://doi.org/10.3390/ncrna9060074
APA StyleLinscott, M. L., Yildiz, Y., Flury, S., Newby, M. L., & Pak, T. R. (2023). Age and 17β-Estradiol (E2) Facilitate Nuclear Export and Argonaute Loading of microRNAs in the Female Brain. Non-Coding RNA, 9(6), 74. https://doi.org/10.3390/ncrna9060074