
Carbon Nanomaterials-Based Electrically Conductive Scaffolds to Repair the Ischaemic Heart Tissue

 , , and
, , and
Abstract
:1. Introduction
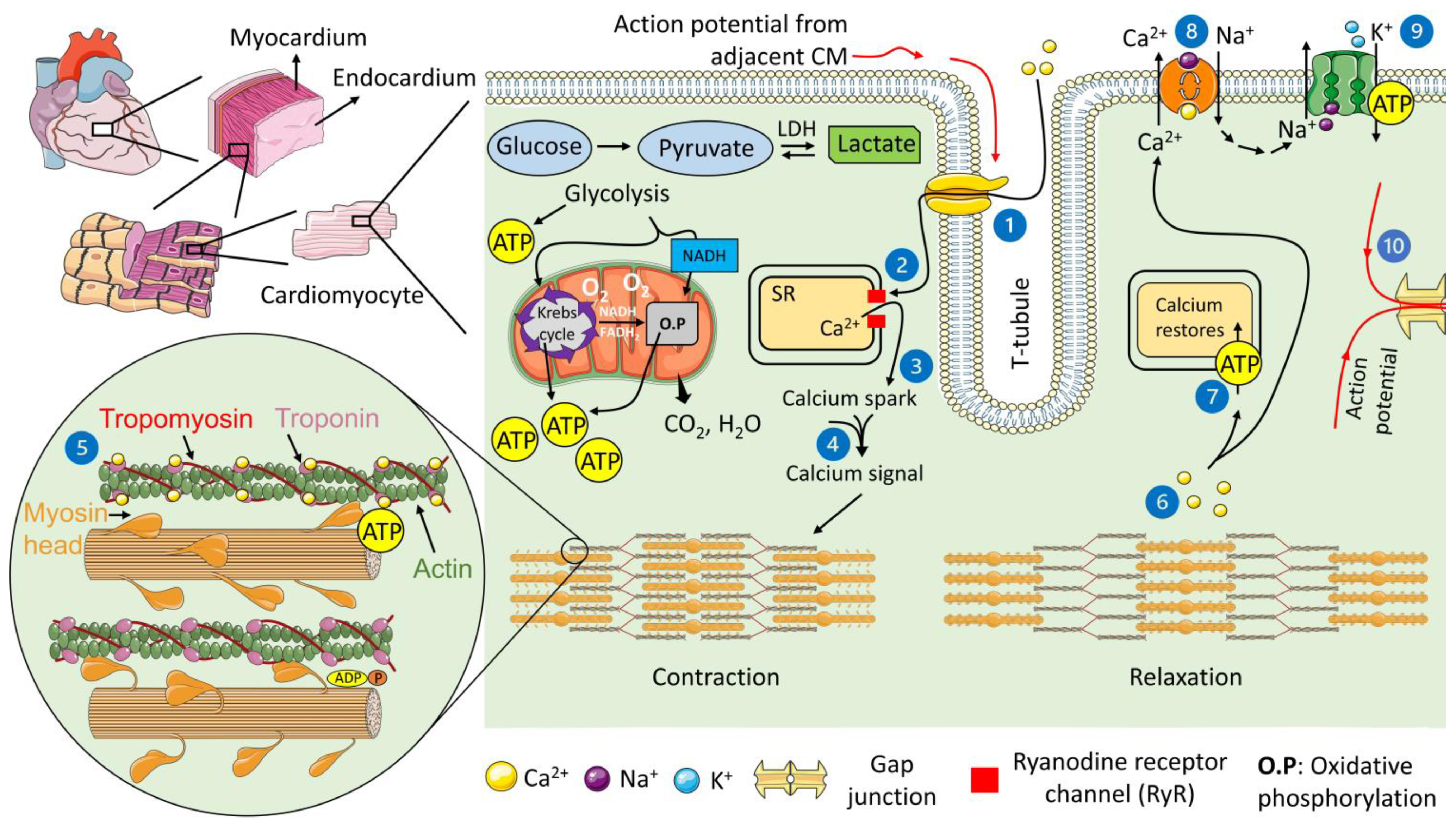
2. The Contractile Machinery of the Heart
3. The Damage to the Contractile Machinery due to Ischaemia
4. The Scar Formation and Its Electrophysiological Remodelling
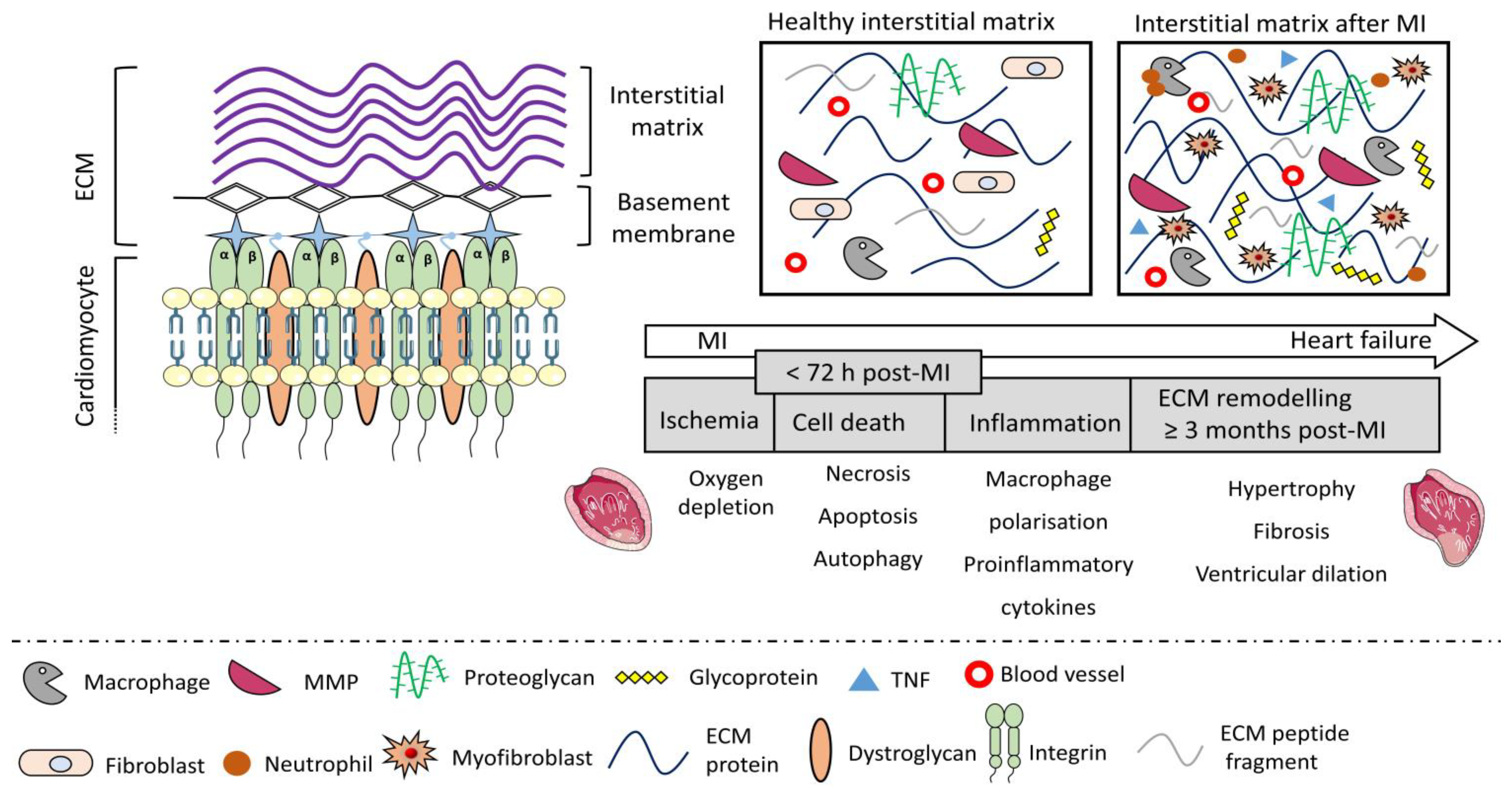
5. Hostile Microenvironment of the Injured Myocardium and the Post-MI Remodelling
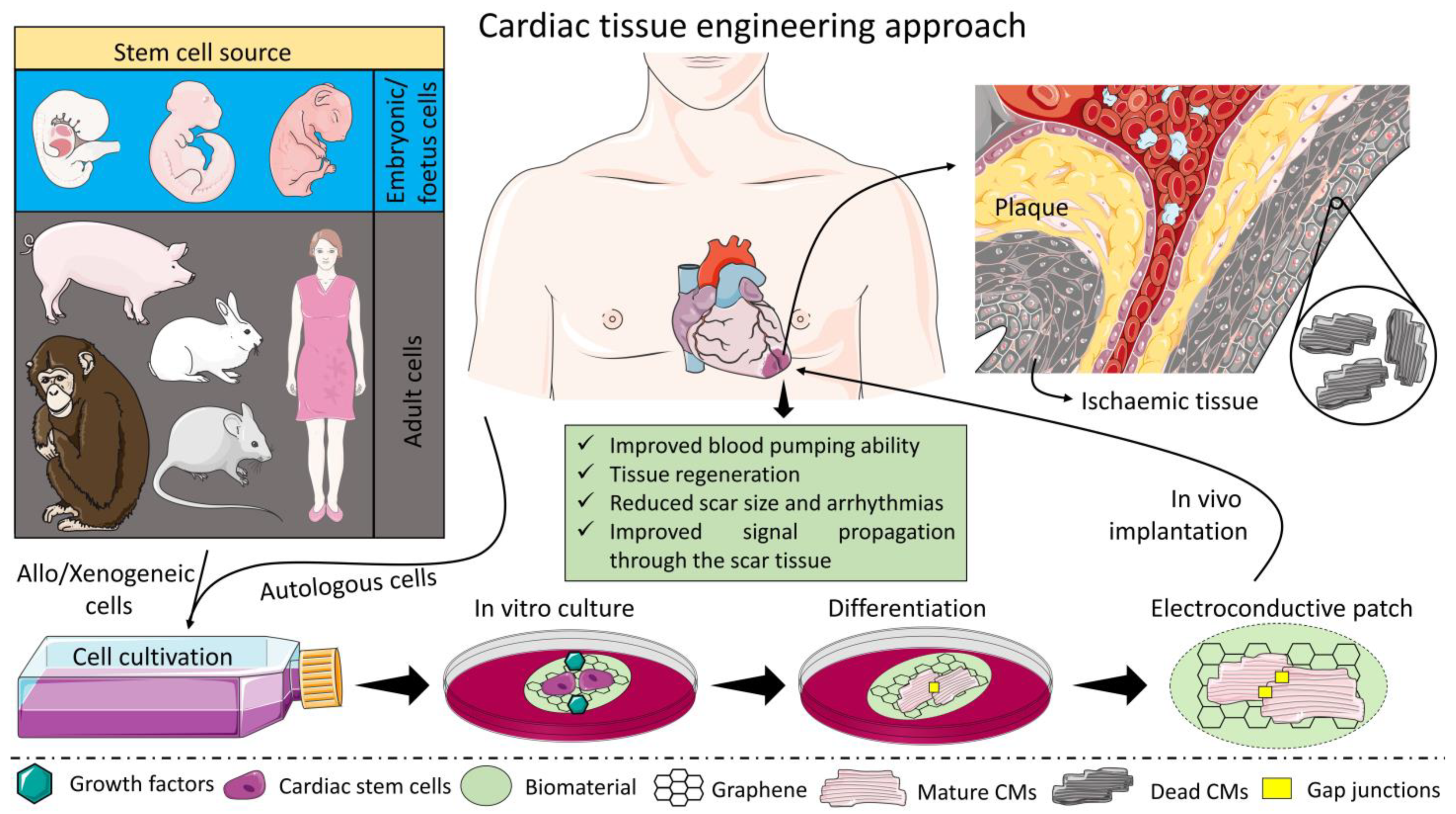
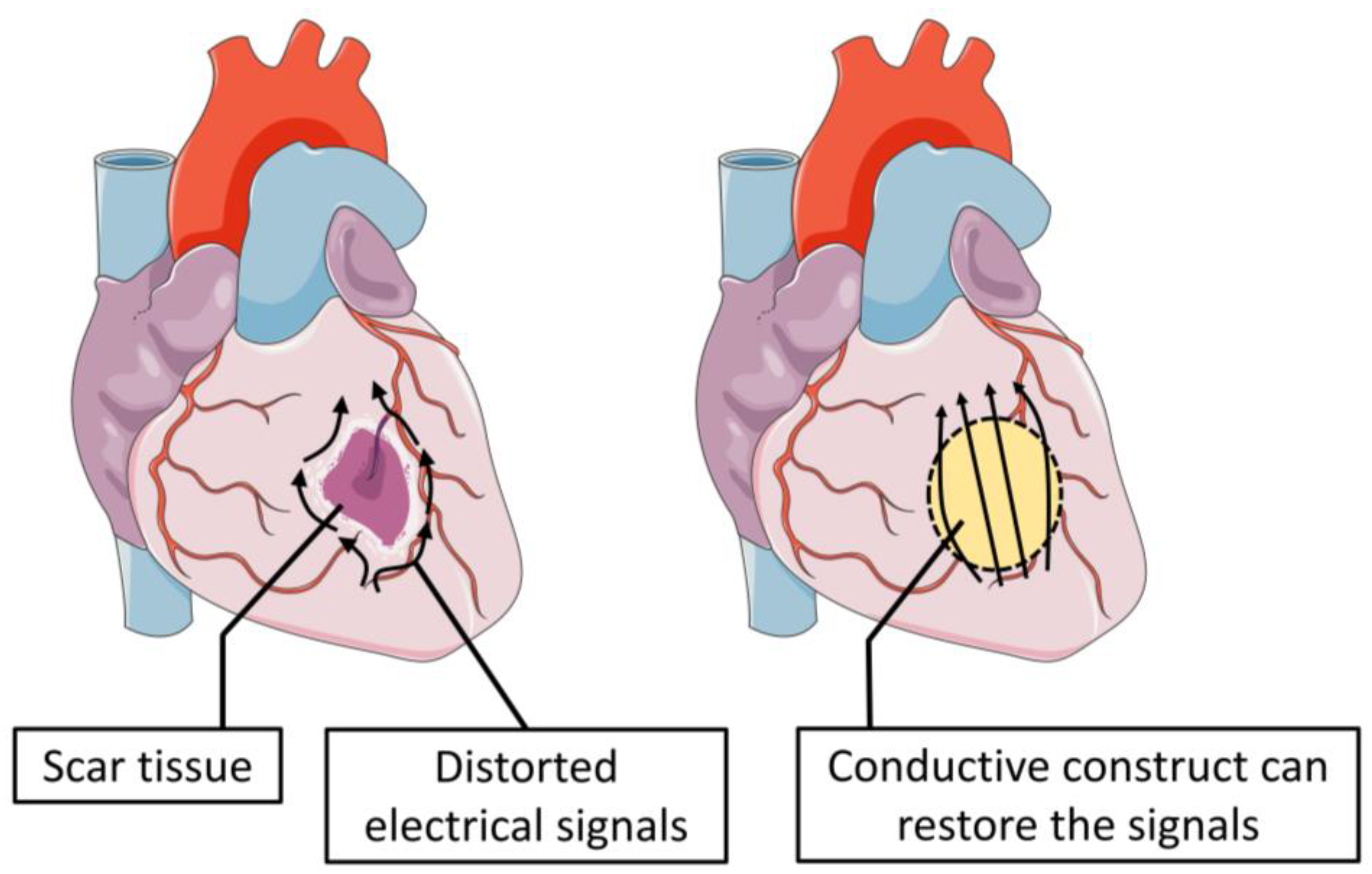
6. How Has T.E Helped So Far to Mitigate the Problem?
7. The Conductive Scaffold
7.1. Carbon Nanotubes
7.2. Graphene
7.3. Fullerene
7.4. Carbon Nanofibers
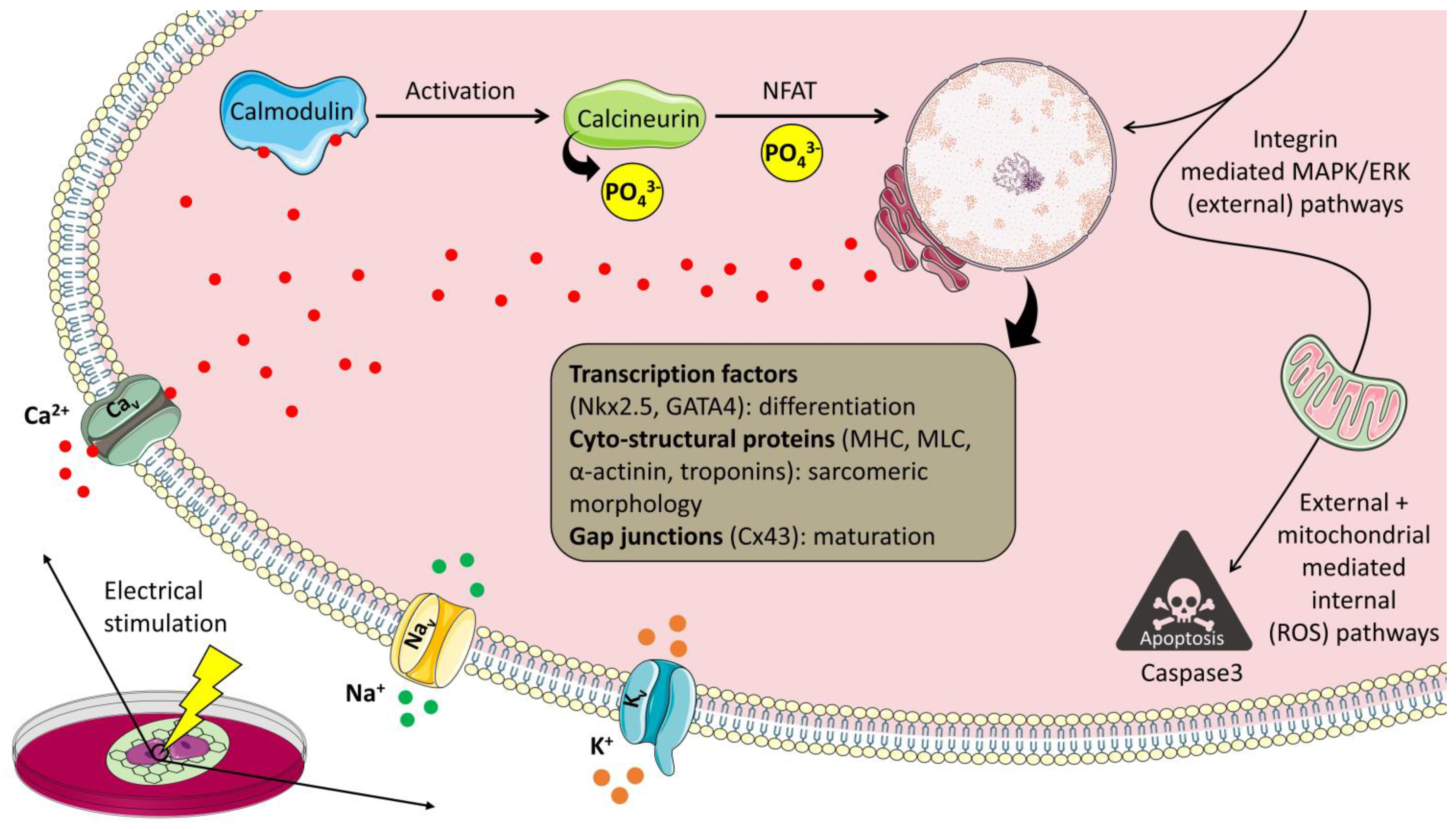
8. Possible Cell-Matrix Interactions
9. In Vivo Efficacy of the Conductive Constructs
10. Drawbacks

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbon Nanomaterial | Properties | Dose | Animal | Outcomes |
|---|---|---|---|---|
| SWCNTs [112] | Diameter: 0.8–1.2 nm Length: 100–1000 nm Surface area: 508 m2/g | 5 mg/m3 | Female C57BL/6 mice | Four days of exposure led to inflammation with oxidative, fibrotic, and mutagenic responses. |
| SWCNTs [113] | Diameter: 2 nm Length: 0.5–40 μm Surface area: 300–600 m2/g | 2 mg/kg | F344 rats | Presence of alveolar lesions after 21 days of exposure. |
| SWCNTs [114] | Length < 1 μm | 0.064–0.64 mg/kg | Female F344 rats | Oxidative damage to DNA in the liver and lungs. |
| MWCNTs [115] | Diameter: 10 nm Length: 200–300 nm Surface area: 253 m2/g | 11 and 241 mg/m3 | Male Webster rats | Fibrosis, oxidative stress, and inflammation at 241 mg/m3 concentration while no fibrosis at 11 mg/m3. |
| MWCNTs [116] | Outer diameter: 17.8 ± 6.4 nm, Length: 825 ± 174 nm Surface modification: carboxyl and hydroxyl groups Zeta potential: −48.9 ± 8 mV | Powder, <100 μg/rat | Wister strain rats | Two years post subcutaneous implantation large agglomerates did not degrade, but smaller ones underwent lysosomal degradation. The animals did not show any sign of cancer or severe inflammation. |
| MWCNTs [117] | Diameter: 50 nm Length: 10 μm Surface area: 280 m2/g | 32.6 mg/m3 | Female Kunming mice | Inhalation at the rate of 6 h/day resulted in no toxic effects after 30 days. However, severe pulmonary damage was observed after 60 days. |
| MWCNTs [118] | Diameter: 49 nm, Length: 3.86 μm | ≥40 μg/mouse | Male C57BL/5 mice | Inflammation and pulmonary fibrosis with granuloma formation over 56 days of exposure. |
| C-60 fullerene [119] | Size: 0.7 nm | 0.2 mg/mouse | Male C57BL/6J mice | DNA damage in the lungs. |
| Graphene oxide [120] | Thickness: 20–30 layers | 0.12, 0.47 and 1.88 mg/L | Male SD rats | No toxic effects 28 days post-exposure |
| Reduced graphene oxide [121] | Thickness: 5 nm Zeta potential: 25 mV | 7 mg/kg | Male Wister rats | No toxic effect on hippocampus, liver, blood, and lungs 7 days post-exposure. |
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
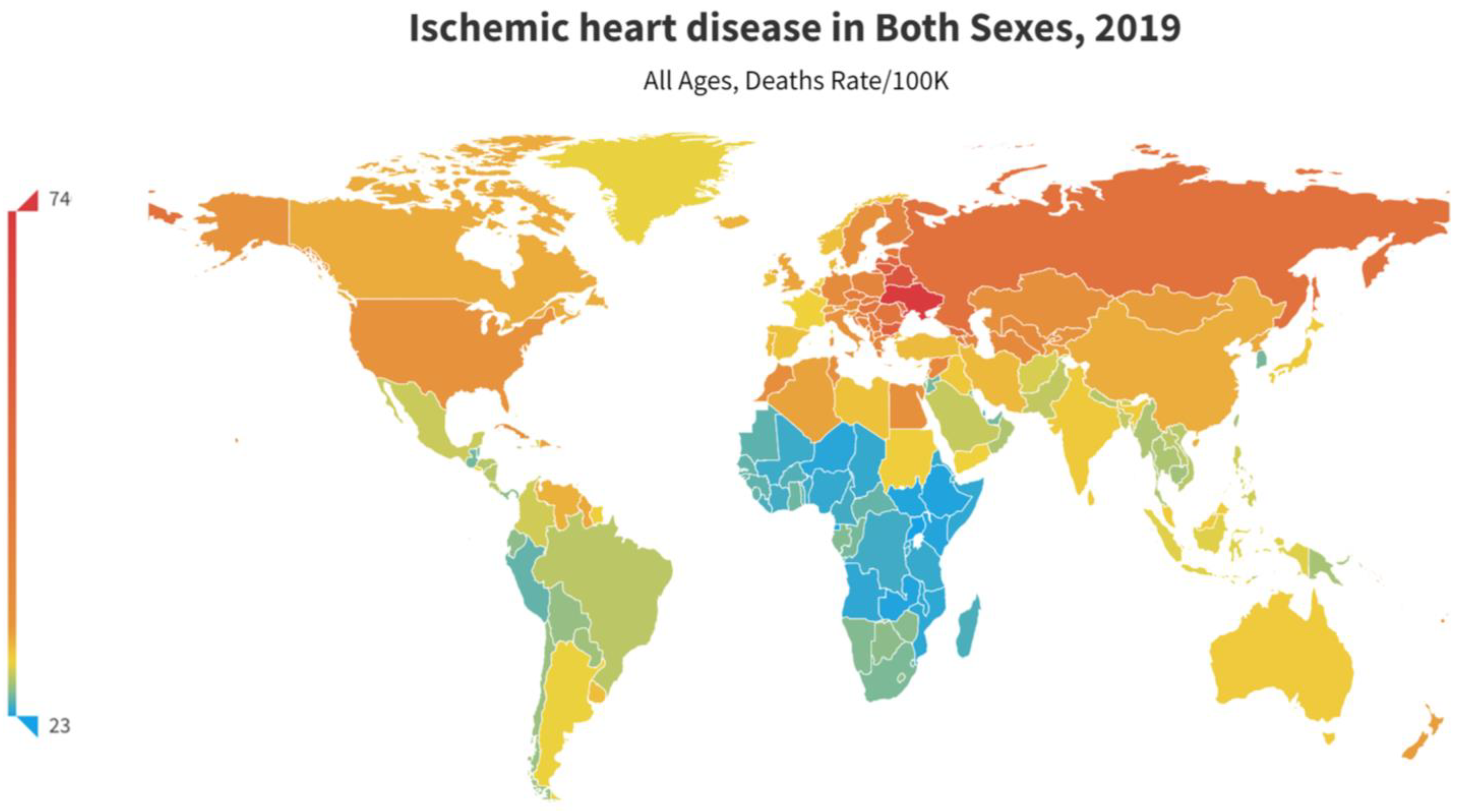
- Khan, M.A.; Hashim, M.J.; Mustafa, H.; Baniyas, M.Y.; Suwaidi, S.K.B.M.A.; AlKatheeri, R.; Alblooshi, F.M.K.; Almatrooshi, M.E.A.H.; Alzaabi, M.E.H.; Darmaki, R.S.A.; et al. Global Epidemiology of Ischemic Heart Disease: Results from the Global Burden of Disease Study. Cureus 2020, 12, e9349. [Google Scholar] [CrossRef]
- Ambrose, J.A.; Singh, M. Pathophysiology of Coronary Artery Disease Leading to Acute Coronary Syndromes. F1000Prime Rep. 2015, 7, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, O.; Zdunek, S.; Felker, A.; Salehpour, M.; Alkass, K.; Bernard, S.; Sjostrom, S.L.; Szewczykowska, M.; Jackowska, T.; dos Remedios, C.; et al. Dynamics of Cell Generation and Turnover in the Human Heart. Cell 2015, 161, 1566–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadahiro, T. Cardiac Regeneration with Pluripotent Stem Cell-Derived Cardiomyocytes and Direct Cardiac Reprogramming. Regen. Ther. 2019, 11, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Burke, A.P.; Virmani, R. Pathophysiology of Acute Myocardial Infarction. Med. Clin. N. Am. 2007, 91, 553–572. [Google Scholar] [CrossRef]
- Heusch, G.; Gersh, B.J. The Pathophysiology of Acute Myocardial Infarction and Strategies of Protection beyond Reperfusion: A Continual Challenge. Eur. Heart J. 2017, 38, 774–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangalore, S.; Fakheri, R.; Wandel, S.; Toklu, B.; Wandel, J.; Messerli, F.H. Renin Angiotensin System Inhibitors for Patients with Stable Coronary Artery Disease without Heart Failure: Systematic Review and Meta-Analysis of Randomized Trials. BMJ 2017, 356, j4. [Google Scholar] [CrossRef] [Green Version]
- Freemantle, N.; Cleland, J.; Young, P.; Mason, J.; Harrison, J. β Blockade after Myocardial Infarction: Systematic Review and Meta Regression Analysis. BMJ 1999, 318, 1730–1737. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Xiang, J.; Wang, X.; Liu, H.; Hu, B.; Feng, M.; Fu, Q. Β2-Adrenoceptor Agonist Clenbuterol Reduces Infarct Size and Myocardial Apoptosis after Myocardial Ischaemia/Reperfusion in Anaesthetized Rats. Br. J. Pharmacol. 2010, 160, 1561–1572. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Miao, B.; Charles, E.J.; Wu, D.; Kron, I.L.; French, B.A.; Yang, Z. Stimulation of the Beta2 Adrenergic Receptor at Reperfusion Limits Myocardial Reperfusion Injury via an Interleukin-10-Dependent Anti-Inflammatory Pathway in the Spleen. Circ. J. Off. J. Jpn. Circ. Soc. 2018, 82, 2829–2836. [Google Scholar] [CrossRef]
- Roacho-Pérez, J.A.; Garza-Treviño, E.N.; Moncada-Saucedo, N.K.; Carriquiry-Chequer, P.A.; Valencia-Gómez, L.E.; Matthews, E.R.; Gómez-Flores, V.; Simental-Mendía, M.; Delgado-Gonzalez, P.; Delgado-Gallegos, J.L.; et al. Artificial Scaffolds in Cardiac Tissue Engineering. Life 2022, 12, 1117. [Google Scholar] [CrossRef]
- Perea-Gil, I.; Prat-Vidal, C.; Bayes-Genis, A. In Vivo Experience with Natural Scaffolds for Myocardial Infarction: The Times They Are a-Changin’. Stem Cell Res. Ther. 2015, 6, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ul Haq, A.; Carotenuto, F.; De Matteis, F.; Prosposito, P.; Francini, R.; Teodori, L.; Pasquo, A.; Di Nardo, P. Intrinsically Conductive Polymers for Striated Cardiac Muscle Repair. Int. J. Mol. Sci. 2021, 22, 8550. [Google Scholar] [CrossRef] [PubMed]
- Ul Haq, A.; Carotenuto, F.; Di Nardo, P.; Francini, R.; Prosposito, P.; Pescosolido, F.; De Matteis, F. Extrinsically Conductive Nanomaterials for Cardiac Tissue Engineering Applications. Micromachines 2021, 12, 914. [Google Scholar] [CrossRef]
- Ryoo, S.M.; Kim, W.Y. Clinical Applications of Lactate Testing in Patients with Sepsis and Septic Shock. J. Emerg. Crit. Care Med. 2018, 2, 14. [Google Scholar] [CrossRef]
- Braasch, W.; Gudbjarnason, S.; Puri, P.S.; Ravens, K.G.; Bing, R.J. Early Changes in Energy Metabolism in the Myocardium Following Acute Coronary Artery Occlusion in Anesthetized Dogs. Circ. Res. 1968, 23, 429–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frangogiannis, N.G. Pathophysiology of Myocardial Infarction. In Comprehensive Physiology; American Physiological Society: Bethesda, MD, USA, 2015; pp. 1841–1875. [Google Scholar] [CrossRef]
- Carmeliet, E. Cardiac Ionic Currents and Acute Ischemia: From Channels to Arrhythmias. Physiol. Rev. 1999, 79, 917–1017. [Google Scholar] [CrossRef] [PubMed]
- Kléber, A.G. Resting Membrane Potential, Extracellular Potassium Activity, and Intracellular Sodium Activity during Acute Global Ischemia in Isolated Perfused Guinea Pig Hearts. Circ. Res. 1983, 52, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Coraboeuf, E. Acidosis-Induced Abnormal Repolarization and Repetitive Activity in Isolated Dog Purkinje Fibers. ACIDOSIS-Induc. Abnorm. REPOLARIZATION REPETITIVE Act. Isol. DOG PURKINJE FIBERS. 1980. Available online: https://pascal-francis.inist.fr/vibad/index.php?action=getRecordDetail&idt=PASCAL8050486952 (accessed on 25 November 2022).
- van den Borne, S.W.M.; Diez, J.; Blankesteijn, W.M.; Verjans, J.; Hofstra, L.; Narula, J. Myocardial Remodeling after Infarction: The Role of Myofibroblasts. Nat. Rev. Cardiol. 2010, 7, 30–37. [Google Scholar] [CrossRef]
- Thygesen, K.; Alpert, J.S.; Jaffe, A.S.; Simoons, M.L.; Chaitman, B.R.; White, H.D.; Writing Group on the Joint ESC/ACCF/AHA/WHF Task Force for the Universal Definition of Myocardial Infarction; Katus, H.A.; Apple, F.S.; Lindahl, B. Third Universal Definition of Myocardial Infarction. Eur. Heart J. 2012, 33, 2551–2567. [Google Scholar] [CrossRef] [Green Version]
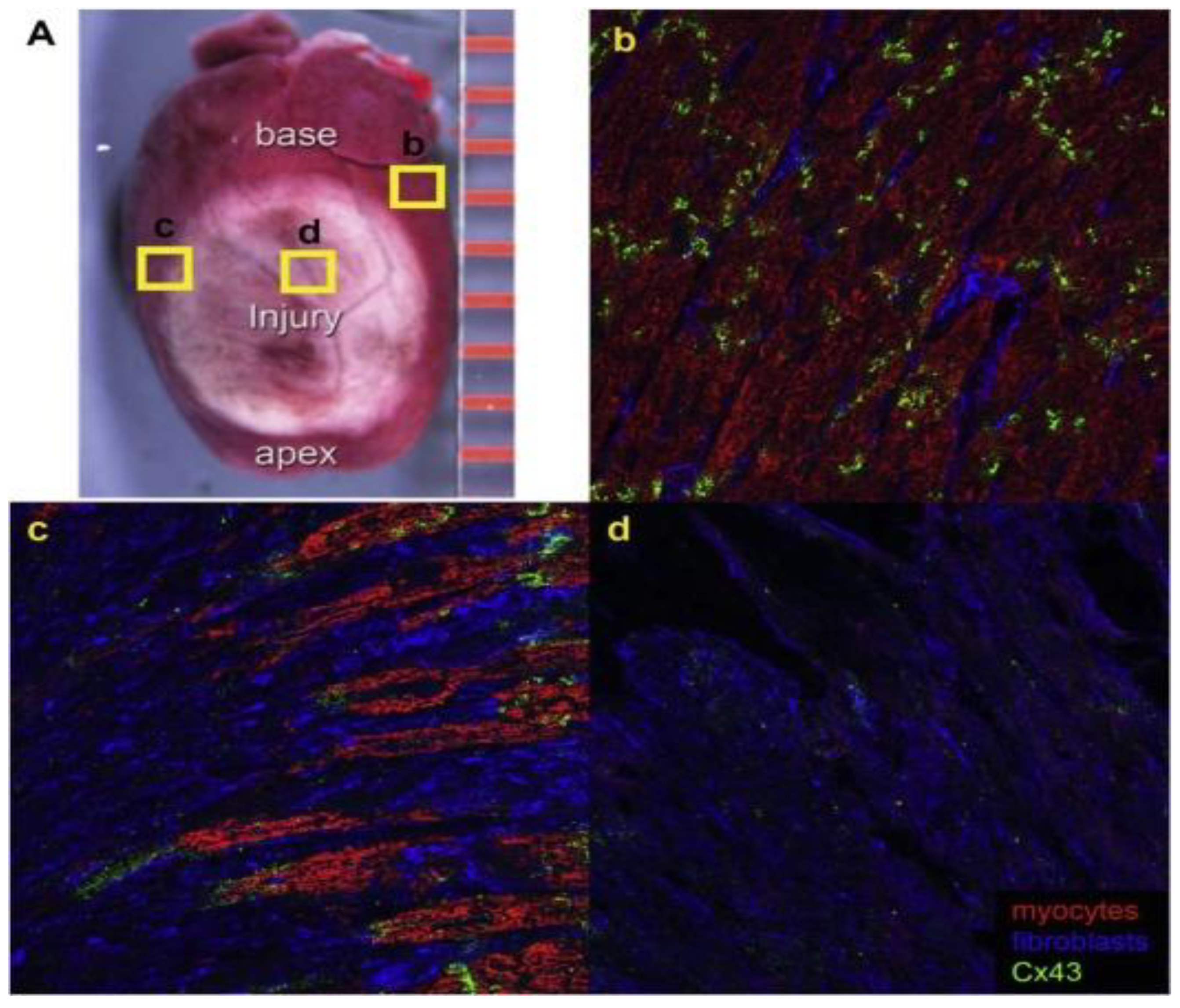
- Gourdie, R.G.; Severs, N.J.; Green, C.R.; Rothery, S.; Germroth, P.; Thompson, R.P. The Spatial Distribution and Relative Abundance of Gap-Junctional Connexin40 and Connexin43 Correlate to Functional Properties of Components of the Cardiac Atrioventricular Conduction System. J. Cell Sci. 1993, 105, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Beardslee, M.A.; Lerner, D.L.; Tadros, P.N.; Laing, J.G.; Beyer, E.C.; Yamada, K.A.; Kléber, A.G.; Schuessler, R.B.; Saffitz, J.E. Dephosphorylation and Intracellular Redistribution of Ventricular Connexin43 during Electrical Uncoupling Induced by Ischemia. Circ. Res. 2000, 87, 656–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, D.L.; Yamada, K.A.; Schuessler, R.B.; Saffitz, J.E. Accelerated Onset and Increased Incidence of Ventricular Arrhythmias Induced by Ischemia in Cx43-Deficient Mice. Circulation 2000, 101, 547–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, N.; Huang, Y.; Zheng, J.; Spencer, C.I.; Zhang, Y.; Fu, J.-D.; Nie, B.; Xie, M.; Zhang, M.; Wang, H.; et al. Conversion of Human Fibroblasts into Functional Cardiomyocytes by Small Molecules. Science 2016, 352, 1216–1220. [Google Scholar] [CrossRef] [Green Version]
- Roell, W.; Lewalter, T.; Sasse, P.; Tallini, Y.N.; Choi, B.-R.; Breitbach, M.; Doran, R.; Becher, U.M.; Hwang, S.-M.; Bostani, T.; et al. Engraftment of Connexin 43-Expressing Cells Prevents Post-Infarct Arrhythmia. Nature 2007, 450, 819–824. [Google Scholar] [CrossRef]
- Fomovsky, G.M.; Clark, S.A.; Parker, K.M.; Ailawadi, G.; Holmes, J.W. Anisotropic Reinforcement of Acute Anteroapical Infarcts Improves Pump Function. Circ. Heart Fail. 2012, 5, 515–522. [Google Scholar] [CrossRef] [Green Version]
- Ursell, P.C.; Gardner, P.I.; Albala, A.; Fenoglio, J.J.; Wit, A.L. Structural and Electrophysiological Changes in the Epicardial Border Zone of Canine Myocardial Infarcts during Infarct Healing. Circ. Res. 1985, 56, 436–451. [Google Scholar] [CrossRef] [Green Version]
- Driesen, R.B.; Verheyen, F.K.; Dijkstra, P.; Thoné, F.; Cleutjens, J.P.; Lenders, M.-H.; Ramaekers, F.C.S.; Borgers, M. Structural Remodelling of Cardiomyocytes in the Border Zone of Infarcted Rabbit Heart. Mol. Cell. Biochem. 2007, 302, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Jugdutt, B.I. Ventricular Remodeling After Infarction and the Extracellular Collagen Matrix. Circulation 2003, 108, 1395–1403. [Google Scholar] [CrossRef]
- Baba, S.; Dun, W.; Cabo, C.; Boyden, P.A. Remodeling in Cells From Different Regions of the Reentrant Circuit During Ventricular Tachycardia. Circulation 2005, 112, 2386–2396. [Google Scholar] [CrossRef] [Green Version]
- Dun, W.; Lowe, J.S.; Wright, P.; Hund, T.J.; Mohler, P.J.; Boyden, P.A. Ankyrin-G Participates in INa Remodeling in Myocytes from the Border Zones of Infarcted Canine Heart. PLoS ONE 2013, 8, e78087. [Google Scholar] [CrossRef] [PubMed]
- Lue, W.M.; Boyden, P.A. Abnormal Electrical Properties of Myocytes from Chronically Infarcted Canine Heart. Alterations in Vmax and the Transient Outward Current. Circulation 1992, 85, 1175–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, J.M.B.; Boyden, P.A. Electrical Remodeling in Ischemia and Infarction. Cardiovasc. Res. 1999, 42, 284–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ongstad, E.L.; Gourdie, R.G. Can Heart Function Lost to Disease Be Regenerated by Therapeutic Targeting of Cardiac Scar Tissue? Semin. Cell Dev. Biol. 2016, 58, 41–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bers, D.M. Calcium Cycling and Signaling in Cardiac Myocytes. Annu. Rev. Physiol. 2008, 70, 23–49. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, D.S.; Jackson, L.E.; Smith, J.M.; Garan, H.; Ruskin, J.N.; Cohen, R.J. Electrical Alternans and Vulnerability to Ventricular Arrhythmias. N. Engl. J. Med. 1994, 330, 235–241. [Google Scholar] [CrossRef]
- Iijima, K.; Geshi, E.; Nomizo, A.; Arata, Y.; Katagiri, T. Alterations in Sarcoplasmic Reticulum and Angiotensin II Type 1 Receptor Gene Expression after Myocardial Infarction in Rats. Jpn. Circ. J. 1998, 62, 449–454. [Google Scholar] [CrossRef] [Green Version]
- Frangogiannis, N.G.; Smith, C.W.; Entman, M.L. The Inflammatory Response in Myocardial Infarction. Cardiovasc. Res. 2002, 53, 31–47. [Google Scholar] [CrossRef]
- Rienks, M.; Papageorgiou, A.-P.; Frangogiannis, N.G.; Heymans, S. Myocardial Extracellular Matrix: An Ever-Changing and Diverse Entity. Circ. Res. 2014, 114, 872–888. [Google Scholar] [CrossRef]
- Khodayari, S.; Khodayari, H.; Amiri, A.Z.; Eslami, M.; Farhud, D.; Hescheler, J.; Nayernia, K. Inflammatory Microenvironment of Acute Myocardial Infarction Prevents Regeneration of Heart with Stem Cells Therapy. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2019, 53, 887–909. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; de Castro Brás, L.E.; Toba, H.; Iyer, R.P.; Hall, M.E.; Winniford, M.D.; Lange, R.A.; Tyagi, S.C.; Lindsey, M.L. Myofibroblasts and the Extracellular Matrix Network in Post-Myocardial Infarction Cardiac Remodeling. Pflugers Arch. 2014, 466, 1113–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Y.; Lv, A.; Wang, L.; Yan, X.; Zhao, W.; Cao, F. Engineered Myocardial Tissues Constructed in Vivo Using Cardiomyocyte-like Cells Derived from Bone Marrow Mesenchymal Stem Cells in Rats. J. Biomed. Sci. 2012, 19, 6. [Google Scholar] [CrossRef] [Green Version]
- Morritt, A.N.; Bortolotto, S.K.; Dilley, R.J.; Han, X.; Kompa, A.R.; McCombe, D.; Wright, C.E.; Itescu, S.; Angus, J.A.; Morrison, W.A. Cardiac Tissue Engineering in an In Vivo Vascularized Chamber. Circulation 2007, 115, 353–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Kupfer, M.E.; Jung, J.P.; Yang, L.; Zhang, P.; Da Sie, Y.; Tran, Q.; Ajeti, V.; Freeman, B.T.; Fast, V.G.; et al. Myocardial Tissue Engineering With Cells Derived From Human-Induced Pluripotent Stem Cells and a Native-Like, High-Resolution, 3-Dimensionally Printed Scaffold. Circ. Res. 2017, 120, 1318–1325. [Google Scholar] [CrossRef] [Green Version]
- Madden, L.R.; Mortisen, D.J.; Sussman, E.M.; Dupras, S.K.; Fugate, J.A.; Cuy, J.L.; Hauch, K.D.; Laflamme, M.A.; Murry, C.E.; Ratner, B.D. Proangiogenic Scaffolds as Functional Templates for Cardiac Tissue Engineering. Proc. Natl. Acad. Sci. USA 2010, 107, 15211–15216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendel, J.S.; Ye, L.; Zhang, P.; Tranquillo, R.T.; Zhang, J.J. Functional Consequences of a Tissue-Engineered Myocardial Patch for Cardiac Repair in a Rat Infarct Model. Tissue Eng. Part A 2014, 20, 1325–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoppel, W.L.; Hu, D.; Domian, I.J.; Kaplan, D.L.; Black, L.D. Anisotropic Silk Biomaterials Containing Cardiac Extracellular Matrix for Cardiac Tissue Engineering. Biomed. Mater. 2015, 10, 034105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuckey, D.J.; Ishii, H.; Chen, Q.-Z.; Boccaccini, A.R.; Hansen, U.; Carr, C.A.; Roether, J.A.; Jawad, H.; Tyler, D.J.; Ali, N.N.; et al. Magnetic Resonance Imaging Evaluation of Remodeling by Cardiac Elastomeric Tissue Scaffold Biomaterials in a Rat Model of Myocardial Infarction. Tissue Eng. Part A 2010, 16, 3395–3402. [Google Scholar] [CrossRef]
- Ban, K.; Park, H.-J.; Kim, S.; Andukuri, A.; Cho, K.-W.; Hwang, J.W.; Cha, H.J.; Kim, S.Y.; Kim, W.-S.; Jun, H.-W.; et al. Cell Therapy with Embryonic Stem Cell-Derived Cardiomyocytes Encapsulated in Injectable Nanomatrix Gel Enhances Cell Engraftment and Promotes Cardiac Repair. ACS Nano 2014, 8, 10815–10825. [Google Scholar] [CrossRef]
- Sun, H.; Lü, S.; Jiang, X.-X.; Li, X.; Li, H.; Lin, Q.; Mou, Y.; Zhao, Y.; Han, Y.; Zhou, J.; et al. Carbon Nanotubes Enhance Intercalated Disc Assembly in Cardiac Myocytes via the Β1-Integrin-Mediated Signaling Pathway. Biomaterials 2015, 55, 84–95. [Google Scholar] [CrossRef]
- Ahmadi, P.; Nazeri, N.; Derakhshan, M.A.; Ghanbari, H. Preparation and Characterization of Polyurethane/Chitosan/CNT Nanofibrous Scaffold for Cardiac Tissue Engineering. Int. J. Biol. Macromol. 2021, 180, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, J.-N.; Tsai, C.-J.; Nussinov, R.; Ma, B. Diameters of Single-Walled Carbon Nanotubes (SWCNTs) and Related Nanochemistry and Nanobiology. Front. Mater. Sci. China 2010, 4, 17–28. [Google Scholar] [CrossRef]
- Sakurai, S.; Inaguma, M.; Futaba, D.N.; Yumura, M.; Hata, K. A Fundamental Limitation of Small Diameter Single-Walled Carbon Nanotube Synthesis-A Scaling Rule of the Carbon Nanotube Yield with Catalyst Volume. Materials 2013, 6, 2633–2641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanone, S.; Andujar, P.; Kermanizadeh, A.; Boczkowski, J. Determinants of Carbon Nanotube Toxicity. Adv. Drug Deliv. Rev. 2013, 65, 2063–2069. [Google Scholar] [CrossRef] [PubMed]
- Saito, N.; Haniu, H.; Usui, Y.; Aoki, K.; Hara, K.; Takanashi, S.; Shimizu, M.; Narita, N.; Okamoto, M.; Kobayashi, S.; et al. Safe Clinical Use of Carbon Nanotubes as Innovative Biomaterials. Chem. Rev. 2014, 114, 6040–6079. [Google Scholar] [CrossRef] [PubMed]
- Roshanbinfar, K.; Hilborn, J.; Varghese, P.O.; Oommen, P.O. Injectable and Thermoresponsive Pericardial Matrix Derived Conductive Scaffold for Cardiac Tissue Engineering. RSC Adv. 2017, 7, 31980–31988. [Google Scholar] [CrossRef] [Green Version]
- Ramón-Azcón, J.; Ahadian, S.; Estili, M.; Liang, X.; Ostrovidov, S.; Kaji, H.; Shiku, H.; Ramalingam, M.; Nakajima, K.; Sakka, Y.; et al. Dielectrophoretically Aligned Carbon Nanotubes to Control Electrical and Mechanical Properties of Hydrogels to Fabricate Contractile Muscle Myofibers. Adv. Mater. 2013, 25, 4028–4034. [Google Scholar] [CrossRef]
- Abedi, A.; Bakhshandeh, B.; Babaie, A.; Mohammadnejad, J.; Vahdat, S.; Mombeiny, R.; Moosavi, S.R.; Amini, J.; Tayebi, L. Concurrent Application of Conductive Biopolymeric Chitosan/ Polyvinyl Alcohol/ MWCNTs Nanofibers, Intracellular Signaling Manipulating Molecules and Electrical Stimulation for More Effective Cardiac Tissue Engineering. Mater. Chem. Phys. 2021, 258, 123842. [Google Scholar] [CrossRef]
- Ahadian, S.; Yamada, S.; Ramón-Azcón, J.; Estili, M.; Liang, X.; Nakajima, K.; Shiku, H.; Khademhosseini, A.; Matsue, T. Hybrid Hydrogel-Aligned Carbon Nanotube Scaffolds to Enhance Cardiac Differentiation of Embryoid Bodies. Acta Biomater. 2016, 31, 134–143. [Google Scholar] [CrossRef]
- Ahadian, S.; Yamada, S.; Estili, M.; Liang, X.; Banan Sadeghian, R.; Nakajima, K.; Shiku, H.; Matsue, T.; Khademhosseini, A. Carbon Nanotubes Embedded in Embryoid Bodies Direct Cardiac Differentiation. Biomed. Microdevices 2017, 19, 57. [Google Scholar] [CrossRef]
- Mehrotra, S.; Singh, R.D.; Bandyopadhyay, A.; Janani, G.; Dey, S.; Mandal, B.B. Engineering Microsphere-Loaded Non-Mulberry Silk-Based 3D Bioprinted Vascularized Cardiac Patches with Oxygen-Releasing and Immunomodulatory Potential. ACS Appl. Mater. Interfaces 2021, 13, 50744–50759. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.R.; Jung, S.M.; Zalabany, M.; Kim, K.; Zorlutuna, P.; Kim, S.B.; Nikkhah, M.; Khabiry, M.; Azize, M.; Kong, J.; et al. Carbon-Nanotube-Embedded Hydrogel Sheets for Engineering Cardiac Constructs and Bioactuators. ACS Nano 2013, 7, 2369–2380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Zhou, J.; Huang, Z.; Qu, L.; Lin, N.; Liang, C.; Dai, R.; Tang, L.; Tian, F. Carbon Nanotube-Incorporated Collagen Hydrogels Improve Cell Alignment and the Performance of Cardiac Constructs. Int. J. Nanomed. 2017, 12, 3109–3120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehdikhani, M.; Ghaziof, S. Electrically Conductive Poly-$${\upepsilon }$$-Caprolactone/Polyethylene Glycol/Multi-Wall Carbon Nanotube Nanocomposite Scaffolds Coated with Fibrin Glue for Myocardial Tissue Engineering. Appl. Phys. A 2018, 124, 77. [Google Scholar] [CrossRef]
- Ren, J.; Xu, Q.; Chen, X.; Li, W.; Guo, K.; Zhao, Y.; Wang, Q.; Zhang, Z.; Peng, H.; Li, Y.-G. Superaligned Carbon Nanotubes Guide Oriented Cell Growth and Promote Electrophysiological Homogeneity for Synthetic Cardiac Tissues. Adv. Mater. 2017, 29, 1702713. [Google Scholar] [CrossRef]
- Martinelli, V.; Cellot, G.; Toma, F.M.; Long, C.S.; Caldwell, J.H.; Zentilin, L.; Giacca, M.; Turco, A.; Prato, M.; Ballerini, L.; et al. Carbon Nanotubes Promote Growth and Spontaneous Electrical Activity in Cultured Cardiac Myocytes. Nano Lett. 2012, 12, 1831–1838. [Google Scholar] [CrossRef]
- Kharaziha, M.; Shin, S.R.; Nikkhah, M.; Topkaya, S.N.; Masoumi, N.; Annabi, N.; Dokmeci, M.R.; Khademhosseini, A. Tough and Flexible CNT-Polymeric Hybrid Scaffolds for Engineering Cardiac Constructs. Biomaterials 2014, 35, 7346–7354. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Dong, Y.; Chen, P.; Chen, R.; Li, Y.; Du, W.; Liu, B.-F. Reduced Graphene Oxide Foam Templated by Nickel Foam for Organ-on-a-Chip Engineering of Cardiac Constructs. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 117, 111344. [Google Scholar] [CrossRef]
- Karimi Hajishoreh, N.; Baheiraei, N.; Naderi, N.; Salehnia, M. Reduced Graphene Oxide Facilitates Biocompatibility of Alginate for Cardiac Repair. J. Bioact. Compat. Polym. 2020, 35, 363–377. [Google Scholar] [CrossRef]
- Norahan, M.H.; Pourmokhtari, M.; Saeb, M.R.; Bakhshi, B.; Soufi Zomorrod, M.; Baheiraei, N. Electroactive Cardiac Patch Containing Reduced Graphene Oxide with Potential Antibacterial Properties. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 104, 109921. [Google Scholar] [CrossRef]
- Zhou, J.; Yang, X.; Liu, W.; Wang, C.; Shen, Y.; Zhang, F.; Zhu, H.; Sun, H.; Chen, J.; Lam, J.; et al. Injectable OPF/Graphene Oxide Hydrogels Provide Mechanical Support and Enhance Cell Electrical Signaling after Implantation into Myocardial Infarct. Theranostics 2018, 8, 3317–3330. [Google Scholar] [CrossRef] [PubMed]
- Talebi, A.; Labbaf, S.; Karimzadeh, F.; Masaeli, E.; Nasr Esfahani, M.-H. Electroconductive Graphene-Containing Polymeric Patch: A Promising Platform for Future Cardiac Repair. ACS Biomater. Sci. Eng. 2020, 6, 4214–4224. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.R.; Zihlmann, C.; Akbari, M.; Assawes, P.; Cheung, L.; Zhang, K.; Manoharan, V.; Zhang, Y.S.; Yüksekkaya, M.; Wan, K.-T.; et al. Reduced Graphene Oxide-GelMA Hybrid Hydrogels as Scaffolds for Cardiac Tissue Engineering. Small Weinh. Bergstr. Ger. 2016, 12, 3677–3689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsui, J.H.; Leonard, A.; Camp, N.D.; Long, J.T.; Nawas, Z.Y.; Chavanachat, R.; Smith, A.S.T.; Choi, J.S.; Dong, Z.; Ahn, E.H.; et al. Tunable Electroconductive Decellularized Extracellular Matrix Hydrogels for Engineering Human Cardiac Microphysiological Systems. Biomaterials 2021, 272, 120764. [Google Scholar] [CrossRef]
- Hitscherich, P.; Aphale, A.; Gordan, R.; Whitaker, R.; Singh, P.; Xie, L.-H.; Patra, P.; Lee, E.J. Electroactive Graphene Composite Scaffolds for Cardiac Tissue Engineering. J. Biomed. Mater. Res. A 2018, 106, 2923–2933. [Google Scholar] [CrossRef]
- Saravanan, S.; Sareen, N.; Abu-El-Rub, E.; Ashour, H.; Sequiera, G.L.; Ammar, H.I.; Gopinath, V.; Shamaa, A.A.; Sayed, S.S.E.; Moudgil, M.; et al. Graphene Oxide-Gold Nanosheets Containing Chitosan Scaffold Improves Ventricular Contractility and Function After Implantation into Infarcted Heart. Sci. Rep. 2018, 8, 15069. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-P.; Qu, K.-Y.; Zhou, B.; Zhang, F.; Wang, Y.-Y.; Abodunrin, O.D.; Zhu, Z.; Huang, N.-P. Electrical Stimulation of Neonatal Rat Cardiomyocytes Using Conductive Polydopamine-Reduced Graphene Oxide-Hybrid Hydrogels for Constructing Cardiac Microtissues. Colloids Surf. B Biointerfaces 2021, 205, 111844. [Google Scholar] [CrossRef]
- Zhao, G.; Feng, Y.; Xue, L.; Cui, M.; Zhang, Q.; Xu, F.; Peng, N.; Jiang, Z.; Gao, D.; Zhang, X. Anisotropic Conductive Reduced Graphene Oxide/Silk Matrices Promote Post-Infarction Myocardial Function by Restoring Electrical Integrity. Acta Biomater. 2022, 139, 190–203. [Google Scholar] [CrossRef]
- Goodarzi, S.; Da Ros, T.; Conde, J.; Sefat, F.; Mozafari, M. Fullerene: Biomedical Engineers Get to Revisit an Old Friend. Mater. Today 2017, 20, 460–480. [Google Scholar] [CrossRef]
- Hao, T.; Zhou, J.; Lü, S.; Yang, B.; Wang, Y.; Fang, W.; Jiang, X.; Lin, Q.; Li, J.; Wang, C. Fullerene Mediates Proliferation and Cardiomyogenic Differentiation of Adipose-Derived Stem Cells via Modulation of MAPK Pathway and Cardiac Protein Expression. Int. J. Nanomed. 2016, 11, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Hao, T.; Li, J.; Yao, F.; Dong, D.; Wang, Y.; Yang, B.; Wang, C. Injectable Fullerenol/Alginate Hydrogel for Suppression of Oxidative Stress Damage in Brown Adipose-Derived Stem Cells and Cardiac Repair. ACS Nano 2017, 11, 5474–5488. [Google Scholar] [CrossRef] [PubMed]
- Minami, K.; Kasuya, Y.; Yamazaki, T.; Ji, Q.; Nakanishi, W.; Hill, J.P.; Sakai, H.; Ariga, K. Highly Ordered 1D Fullerene Crystals for Concurrent Control of Macroscopic Cellular Orientation and Differentiation toward Large-Scale Tissue Engineering. Adv. Mater. 2015, 27, 4020–4026. [Google Scholar] [CrossRef] [PubMed]
- Tashakori-Miyanroudi, M.; Rakhshan, K.; Ramez, M.; Asgarian, S.; Janzadeh, A.; Azizi, Y.; Seifalian, A.; Ramezani, F. Conductive Carbon Nanofibers Incorporated into Collagen Bio-Scaffold Assists Myocardial Injury Repair. Int. J. Biol. Macromol. 2020, 163, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Mehrabi, A.; Baheiraei, N.; Adabi, M.; Amirkhani, Z. Development of a Novel Electroactive Cardiac Patch Based on Carbon Nanofibers and Gelatin Encouraging Vascularization. Appl. Biochem. Biotechnol. 2020, 190, 931–948. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.M.; Eng, G.; Caridade, S.G.; Mano, J.F.; Reis, R.L.; Vunjak-Novakovic, G. Electrically Conductive Chitosan/Carbon Scaffolds for Cardiac Tissue Engineering. Biomacromolecules 2014, 15, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Ping, P.; Wang, F.; Luo, L. Synthesis, Secretion, Function, Metabolism and Application of Natriuretic Peptides in Heart Failure. J. Biol. Eng. 2018, 12, 2. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Xiao, B.; Wu, F.; Sun, L.; Li, B.; Guo, W.; Hu, X.; Xu, X.; Wen, T.; Liu, J.; et al. Co-Axial Fibrous Scaffolds Integrating with Carbon Fiber Promote Cardiac Tissue Regeneration Post Myocardial Infarction. Mater. Today Bio 2022, 16, 100415. [Google Scholar] [CrossRef]
- Asiri, A.M.; Marwani, H.M.; Khan, S.B.; Webster, T.J. Greater Cardiomyocyte Density on Aligned Compared with Random Carbon Nanofibers in Polymer Composites. Int. J. Nanomed. 2014, 9, 5533–5539. [Google Scholar] [CrossRef] [Green Version]
- Stoppel, W.L.; Kaplan, D.L.; Black, L.D. Electrical and Mechanical Stimulation of Cardiac Cells and Tissue Constructs. Adv. Drug Deliv. Rev. 2016, 96, 135–155. [Google Scholar] [CrossRef]
- Passier, R.; Zeng, H.; Frey, N.; Naya, F.J.; Nicol, R.L.; McKinsey, T.A.; Overbeek, P.; Richardson, J.A.; Grant, S.R.; Olson, E.N. CaM Kinase Signaling Induces Cardiac Hypertrophy and Activates the MEF2 Transcription Factor in Vivo. J. Clin. Investig. 2000, 105, 1395–1406. [Google Scholar] [CrossRef] [Green Version]
- Adachi, A.; Takahashi, T.; Ogata, T.; Imoto-Tsubakimoto, H.; Nakanishi, N.; Ueyama, T.; Matsubara, H. NFAT5 Regulates the Canonical Wnt Pathway and Is Required for Cardiomyogenic Differentiation. Biochem. Biophys. Res. Commun. 2012, 426, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cao, X. NFAT Directly Regulates Nkx2-5 Transcription during Cardiac Cell Differentiation. Biol. Cell 2009, 101, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; McMillin, J.B.; Lewis, A.; Moore, M.; Zhu, W.G.; Williams, R.S.; Kellems, R.E. Electrical Stimulation of Neonatal Cardiac Myocytes Activates the NFAT3 and GATA4 Pathways and Up-Regulates the Adenylosuccinate Synthetase 1 Gene. J. Biol. Chem. 2000, 275, 1855–1863. [Google Scholar] [CrossRef] [Green Version]
- Watt, A.J.; Battle, M.A.; Li, J.; Duncan, S.A. GATA4 Is Essential for Formation of the Proepicardium and Regulates Cardiogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 12573–12578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saadane, N.; Alpert, L.; Chalifour, L.E. Expression of Immediate Early Genes, GATA-4, and Nkx-2.5 in Adrenergic-Induced Cardiac Hypertrophy and during Regression in Adult Mice. Br. J. Pharmacol. 1999, 127, 1165–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, B.; Jin, J.-P. TNNT1, TNNT2, and TNNT3: Isoform Genes, Regulation, and Structure–Function Relationships. Gene 2016, 582, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kotini, M.; Barriga, E.H.; Leslie, J.; Gentzel, M.; Rauschenberger, V.; Schambony, A.; Mayor, R. Gap Junction Protein Connexin-43 Is a Direct Transcriptional Regulator of N-Cadherin in Vivo. Nat. Commun. 2018, 9, 3846. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Chen, J.; Sun, H.; Qiu, X.; Mou, Y.; Liu, Z.; Zhao, Y.; Li, X.; Han, Y.; Duan, C.; et al. Engineering the Heart: Evaluation of Conductive Nanomaterials for Improving Implant Integration and Cardiac Function. Sci. Rep. 2014, 4, 3733. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhou, J.; Liu, Z.; Chen, J.; Lü, S.; Sun, H.; Li, J.; Lin, Q.; Yang, B.; Duan, C.; et al. A PNIPAAm-Based Thermosensitive Hydrogel Containing SWCNTs for Stem Cell Transplantation in Myocardial Repair. Biomaterials 2014, 35, 5679–5688. [Google Scholar] [CrossRef]
- Bao, R.; Tan, B.; Liang, S.; Zhang, N.; Wang, W.; Liu, W. A π-π Conjugation-Containing Soft and Conductive Injectable Polymer Hydrogel Highly Efficiently Rebuilds Cardiac Function after Myocardial Infarction. Biomaterials 2017, 122, 63–71. [Google Scholar] [CrossRef]
- Yuan, Z.; Qin, Q.; Yuan, M.; Wang, H.; Li, R. Development and Novel Design of Clustery Graphene Oxide Formed Conductive Silk Hydrogel Cell Vesicle to Repair and Routine Care of Myocardial Infarction: Investigation of Its Biological Activity for Cell Delivery Applications. J. Drug Deliv. Sci. Technol. 2020, 60, 102001. [Google Scholar] [CrossRef]
- Advancing Adverse Outcome Pathway (AOP) Development for Nanomaterial Risk Assessment and Categorisation Part 2: Case Study on Tissue Injury; Safety of Manufactured Nanomaterials. Available online: https://www.oecd.org/officialdocuments/publicdisplaydocumentpdf/?cote=env/jm/%20mono (accessed on 25 November 2022).
- Yang, S.-T.; Wang, X.; Jia, G.; Gu, Y.; Wang, T.; Nie, H.; Ge, C.; Wang, H.; Liu, Y. Long-Term Accumulation and Low Toxicity of Single-Walled Carbon Nanotubes in Intravenously Exposed Mice. Toxicol. Lett. 2008, 181, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Elgrabli, D.; Floriani, M.; Abella-Gallart, S.; Meunier, L.; Gamez, C.; Delalain, P.; Rogerieux, F.; Boczkowski, J.; Lacroix, G. Biodistribution and Clearance of Instilled Carbon Nanotubes in Rat Lung. Part. Fibre Toxicol. 2008, 5, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osmond-McLeod, M.J.; Poland, C.A.; Murphy, F.; Waddington, L.; Morris, H.; Hawkins, S.C.; Clark, S.; Aitken, R.; McCall, M.J.; Donaldson, K. Durability and Inflammogenic Impact of Carbon Nanotubes Compared with Asbestos Fibres. Part. Fibre Toxicol. 2011, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Cervantes, D.X.; Maes, H.M.; Schäffer, A.; Hollender, J.; Kohler, H.-P.E. Slow Biotransformation of Carbon Nanotubes by Horseradish Peroxidase. Environ. Sci. Technol. 2014, 48, 4826–4834. [Google Scholar] [CrossRef]
- Russier, J.; Ménard-Moyon, C.; Venturelli, E.; Gravel, E.; Marcolongo, G.; Meneghetti, M.; Doris, E.; Bianco, A. Oxidative Biodegradation of Single- and Multi-Walled Carbon Nanotubes. Nanoscale 2011, 3, 893–896. [Google Scholar] [CrossRef]
- Chen, H.H.; Yu, C.; Ueng, T.H.; Chen, S.; Chen, B.J.; Huang, K.J.; Chiang, L.Y. Acute and Subacute Toxicity Study of Water-Soluble Polyalkylsulfonated C60 in Rats. Toxicol. Pathol. 1998, 26, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Porter, A.E.; Muller, K.; Skepper, J.; Midgley, P.; Welland, M. Uptake of C60 by Human Monocyte Macrophages, Its Localization and Implications for Toxicity: Studied by High Resolution Electron Microscopy and Electron Tomography. Acta Biomater. 2006, 2, 409–419. [Google Scholar] [CrossRef]
- Shvedova, A.A.; Kisin, E.; Murray, A.R.; Johnson, V.J.; Gorelik, O.; Arepalli, S.; Hubbs, A.F.; Mercer, R.R.; Keohavong, P.; Sussman, N.; et al. Inhalation vs. Aspiration of Single-Walled Carbon Nanotubes in C57BL/6 Mice: Inflammation, Fibrosis, Oxidative Stress, and Mutagenesis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 295, L552–L565. [Google Scholar] [CrossRef]
- Mangum, J.B.; Turpin, E.A.; Antao-Menezes, A.; Cesta, M.F.; Bermudez, E.; Bonner, J.C. Single-Walled Carbon Nanotube (SWCNT)-Induced Interstitial Fibrosis in the Lungs of Rats Is Associated with Increased Levels of PDGF MRNA and the Formation of Unique Intercellular Carbon Structures That Bridge Alveolar Macrophages in Situ. Part. Fibre Toxicol. 2006, 3, 15. [Google Scholar] [CrossRef] [Green Version]
- Folkmann, J.K.; Risom, L.; Jacobsen, N.R.; Wallin, H.; Loft, S.; Møller, P. Oxidatively Damaged DNA in Rats Exposed by Oral Gavage to C60 Fullerenes and Single-Walled Carbon Nanotubes. Environ. Health Perspect. 2009, 117, 703–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellinger-Ziegelbauer, H.; Pauluhn, J. Pulmonary Toxicity of Multi-Walled Carbon Nanotubes (Baytubes) Relative to Alpha-Quartz Following a Single 6h Inhalation Exposure of Rats and a 3 Months Post-Exposure Period. Toxicology 2009, 266, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Yokoyama, A.; Nodasaka, Y.; Kohgo, T.; Motomiya, K.; Matsumoto, H.; Nakazawa, E.; Numata, T.; Zhang, M.; Yudasaka, M.; et al. Long-Term Biopersistence of Tangled Oxidized Carbon Nanotubes inside and Outside Macrophages in Rat Subcutaneous Tissue. Sci. Rep. 2013, 3, 2516. [Google Scholar] [CrossRef] [Green Version]
- Li, J.-G.; Li, W.-X.; Xu, J.-Y.; Cai, X.-Q.; Liu, R.-L.; Li, Y.-J.; Zhao, Q.-F.; Li, Q.-N. Comparative Study of Pathological Lesions Induced by Multiwalled Carbon Nanotubes in Lungs of Mice by Intratracheal Instillation and Inhalation. Environ. Toxicol. 2007, 22, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Porter, D.W.; Hubbs, A.F.; Mercer, R.R.; Wu, N.; Wolfarth, M.G.; Sriram, K.; Leonard, S.; Battelli, L.; Schwegler-Berry, D.; Friend, S.; et al. Mouse Pulmonary Dose- and Time Course-Responses Induced by Exposure to Multi-Walled Carbon Nanotubes. Toxicology 2010, 269, 136–147. [Google Scholar] [CrossRef]
- Totsuka, Y.; Higuchi, T.; Imai, T.; Nishikawa, A.; Nohmi, T.; Kato, T.; Masuda, S.; Kinae, N.; Hiyoshi, K.; Ogo, S.; et al. Genotoxicity of Nano/Microparticles in in Vitro Micronuclei, in Vivo Comet and Mutation Assay Systems. Part. Fibre Toxicol. 2009, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.K.; Shin, J.H.; Lee, J.S.; Hwang, J.H.; Lee, J.H.; Baek, J.E.; Kim, T.G.; Kim, B.W.; Kim, J.S.; Lee, G.H.; et al. 28-Day Inhalation Toxicity of Graphene Nanoplatelets in Sprague-Dawley Rats. Nanotoxicology 2016, 10, 891–901. [Google Scholar] [CrossRef]
- Mendonça, M.C.P.; Soares, E.S.; de Jesus, M.B.; Ceragioli, H.J.; Irazusta, S.P.; Batista, Â.G.; Vinolo, M.A.R.; Maróstica Júnior, M.R.; da Cruz-Höfling, M.A. Reduced Graphene Oxide: Nanotoxicological Profile in Rats. J. Nanobiotechnol. 2016, 14, 53. [Google Scholar] [CrossRef] [Green Version]
- Jain, S. Toxicity Issues Related to Biomedical Applications of Carbon Nanotubes. J. Nanomed. Nanotechnol. 2012, 3, 2. [Google Scholar] [CrossRef]
- Raja, I.S.; Song, S.-J.; Kang, M.S.; Lee, Y.B.; Kim, B.; Hong, S.W.; Jeong, S.J.; Lee, J.-C.; Han, D.-W. Toxicity of Zero- and One-Dimensional Carbon Nanomaterials. Nanomaterials 2019, 9, 1214. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Zhang, X.; Sun, L.; Wei, Y.; Wei, X. Cellular Toxicity and Immunological Effects of Carbon-Based Nanomaterials. Part. Fibre Toxicol. 2019, 16, 18. [Google Scholar] [CrossRef] [PubMed]







| Conductive Construct | Mechano-Electrical Properties | Electrical Stimulation | Cell Line | Cellular Behaviour |
|---|---|---|---|---|
| Collagen/SWCNTs composite [52] | σ = 1.72 × 10−9/Ω | - | NRVMs | Enhanced assembly of intercalated discs |
| PU/chitosan/CNTs membranes [53] | UTS = 21.9 MPa, E = 4.34 MPa, R = 0.17 kΩ/sq | - | HUVECs, H9c2 | Improved cell viability |
| Decellularised pericardium ECM, MWCNTs hydrogels [58] | G′ = 229 Pa, G″ = 150 Pa, σ = 15 × 10−3 S/cm | - | HL-1 | Three-fold increase in the proliferation rate |
| MWCNTs, GelMA hydrogels [59] | E = 23.4 kPa, Z = 400 kΩ | 10 ms AC pulses at 1 Hz and 8 V for 2 days | C2C12 | Myotube formation when stimulated electrically |
| CHI, PVA, MWCNTs membranes [60] | E = 941 MPa, εr = 3.8%, σ = 1.2 mS/cm | Monophasic pulses at 1.25 Hz and 10 V for 2 days | USSCs | Differentiation to cardiomyocyte-like cells |
| GelMA-CNTs hydrogels [61] | E = 30.6 kPa, Z = 56 kΩ at 0.2 Hz | 10 ms AC pulses at 1 Hz and 3V for 2 days | 129/SVE-derived mouse stem cells | Formation of beating embryoid bodies with enhanced expression of cTnT2 and Nkx2.5 |
| CNTs embedded inside of EBs [62] | E = 35.2 kPa, Z = 300 kΩ at 1 Hz | 10 ms AC pulses at 1 Hz and 3V for 2 days | 129/SVE-derived mouse stem cells | Several fold increase in the expression of Nkx2.5, acta2, cTnT2, MHC, MLC, and Cdh5 genes |
| Silk/CNTs patch [63] | E = 51 kPa, εr = 42%, R = 94 kΩ | AC square waveform at 1 Hz | neonatal rat CMs | Synchronous beating patterns with contractile phenotype |
| GelMA/CNTs patch [64] | E = 32 kPa, Z = 2 kΩ at 1000 Hz | AC 50 ms pulses between 0.5–3 Hz at 1 V/cm | neonatal rat CMs | Three times higher spontaneous contractions than GelMA |
| SWCNTs/collagen hydrogel [65] | Mechanical strength = 24 kPa, σ = 525 mS/m | - | CMs | Formation of engineered cardiac tissue with strong contraction potential |
| MWCNTs/PCL/PEG/ fibrin glue patch [66] | E = 47 MPa, σ = 0.45 S/m | - | Rat myoblasts | Improved cell viability and proliferation |
| Pristine CNTs films [67] | σ = 3.1 mS | 2 ms AC pulses at 1 Hz and 2 V/cm for 3 days | neonatal rat CMs | Synchronous beating patterns after stimulation |
| Pristine MWCNT films [68] | - | - | NRVMs, cardiac fibroblasts | Development of striated morphology with tight desmosomes like nano-connections |
| PGS-gelatin/CNTs membranes [69] | E = 373 kPa, Z = 7 kΩ at 40 Hz | 50 ms AC biphasic pulses, V = 0–7 volts, f = 1–3 Hz | Neonatal rat CMs | Twice the beating rate five days post-culture on the conductive membranes |
| Conductive Construct | Mechano-Electrical Properties | Electrical Stimulation | Cell Line | Cellular Behaviour |
|---|---|---|---|---|
| rGO foams [70] | G′ = 8 kPa, σ = 112 S/m | - | Neonatal rat CMs | A 3D cellular organisation within conductive foam that formed a large beating syncytium with 65 bpm |
| rGO/alginate hydrogels [71] | G′ = 1 kPa, σ = 1/9 ± 0.16 × 105 S/m | - | hBM-MSCs | Differentiation to CMs-like cells |
| rGO/collagen patch [72] | E = 340 kPa, σ = 22 μS/m | - | CMs | Elevated expression of Cx43 and cTnT2 |
| OPF/GO hydrogels [73] | σ = 4.24 mS/cm | - | Neonatal rat cardiac fibroblasts | Well-organised sarcomeres with enhanced expression of α-tubulin, and actinin |
| PCL/CHI/PPy/graphene patch [74] | E = 0.098 MPa, UTS = 1.27 MPa, εr = 8%, σ = 5.33 S/cm | - | mESCs-CMs | Elevated troponin levels with beating CMs |
| GelMA/rGO hydrogels [75] | E = 22.6 kPa, Z = 1.5 kΩ at 100 Hz | 50 ms AC biphasic pulses at 1–3 Hz and 3–6 V/cm | Primary CMs | Nine times faster beating rate compared to when encapsulated in non-conductive hydrogel |
| rGO/dECM hydrogels [76] | E = 17.5 kPa, σ = 3 S/m | - | hiPSCs-CMs and HS-27A stromal cells | Generation of engineered heart tissue with elevated expression of voltage-gated calcium and potassium channels |
| PCL, graphene composites [77] | σ = 1.5 × 10−8 S/m | 10 ms AC pulses at 1 Hz and 5 V | mESC-CMs | Average beating frequency 1.1 beats/second two weeks post-culture |
| Chitosan/GO-Au patch [78] | σ = 12 × 10−5 S/cm | - | hiPSCs-CMs | Improved cell adhesion |
| rGO/GelMA/ polydopamine hydrogels [79] | E = 23.6 kPa, Z = 1250 Ω at 1000 Hz | AC 2 ms triphasic pulses at 1.5 Hz | CMs | Improved calcium transients with 40 bpm |
| rGO/silk electrospun membranes [80] | Tensile strength = 6 MPa, σ = 0.3 S/cm | AC 5 ms square wave pulses at 100 Hz | CMs and myofibroblasts | Improved cell viability. However, the stimulation caused ventricular fibrillation |
| Conductive Construct | Mechano- Electrical Properties | Electrical Stimulation | Cell Line | Cellular Behaviour |
|---|---|---|---|---|
| Cell suspension + C-60 fullerene NPs [82] | - | - | BADSCs | Improved MAPK expression led to the differentiation of BADSCs to cardiomyocyte-like cells |
| Fullerenol/alginate hydrogels [83] | G′ = 700 Pa, G″ = 100 Pa | - | BADSCs | Inhibition of ROS environment by activating ERK/p38 pathways which led to better cell survival and cardiomyogenic differentiation |
| Fullerene whiskers [84] | - | - | C2C12 | Myogenic differentiation with elevated expression of MyoD and myogenin |
| Conductive Construct | Mechano-Electrical Properties | Electrical Stimulation | Cell Line | Cellular Behaviour |
|---|---|---|---|---|
| Collagen/CNF composites [85] | Mechanical strength = 3.5 N | - | H9c2 | Elevated expression of α-actinin |
| Gelatin/CNF patch [86] | E = 8.42 MPs, UTS = 5.32 MPa, σ = 84 µS/m | - | CMs | Three- to four-fold increase in actn4 and Cx43 expression. |
| Chitosan/CNF composites [87] | E = 28 kPa, σ = 0.25 S/m | - | Neonatal rat CMs | Cell maturation with contractile phenotype evident from elevated expression of cyto-skeletal Cx43, GATA4, troponins, Myh6, Myh7, and ANF |
| PLA/CNFs membranes [89] | E = 389 kPa, σ = 0.58 S/m | - | Neonatal rat CMs | Elevated expression of α-actinin, Cx43 and β-actin |
| PLGA/CNFs composites [90] | E = 7.9 MPa, σ = 0.1 S/m | - | CMs | Increased cell growth and density five days post-culture |
| Conductive Construct | Injured Heart | Implanted Heart |
|---|---|---|
| OPF/GO hydrogels [73] | Wall thickness = 0.37 mm | Wall thickness = 0.77 mm |
| Chitosan/GO-Au patch [78] | Ejection fraction = 40% Fraction shortening = 19% | Ejection fraction = 70% Fraction shortening = 40%, Shorter QRS interval |
| rGO/silk electrospun membranes [80] | Ejection fraction = 46% Fraction shortening = 23% Wall thickness = 0.7 mm | Ejection fraction = 68% Fraction shortening = 38% Wall thickness = 1.2 mm |
| Fullerenol/alginate hydrogels [83] | Wall thickness = 0.61 mm | Wall thickness = 1.3 mm |
| Collagen/CNFs composites [85] | High tissue degeneration with the damaged assembly of intercalated discs | Improved angiogenesis with the repair of intercalated discs assembly |
| PLA/CNFs membranes [89] | Ejection fraction = 50% Fraction shortening = 29% Wall thickness = 0.8 mm | Ejection fraction = 75% Fraction shortening = 45% Wall thickness = 2.4 mm |
| Gelatin/SWCNTs hydrogels [100] | Ejection fraction = 43% Fraction shortening = 19% | Ejection fraction = 49% Fraction shortening = 29% |
| PNIAA/SWCNTs hydrogels + BADSCs [101] | Wall thickness = 538 µm | Wall thickness = 863 µm |
| PEG-MEL/HA-SH/GO composites [102] | Wall thickness = 0.9 mm | Wall thickness = 1.9 mm |
| SF, GO hydrogels [103] | Wall thickness = 250 µm | Wall thickness = 280 µm |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ul Haq, A.; Carotenuto, F.; Trovalusci, F.; De Matteis, F.; Di Nardo, P. Carbon Nanomaterials-Based Electrically Conductive Scaffolds to Repair the Ischaemic Heart Tissue. C 2022, 8, 72. https://doi.org/10.3390/c8040072
Ul Haq A, Carotenuto F, Trovalusci F, De Matteis F, Di Nardo P. Carbon Nanomaterials-Based Electrically Conductive Scaffolds to Repair the Ischaemic Heart Tissue. C. 2022; 8(4):72. https://doi.org/10.3390/c8040072
Chicago/Turabian StyleUl Haq, Arsalan, Felicia Carotenuto, Federica Trovalusci, Fabio De Matteis, and Paolo Di Nardo. 2022. "Carbon Nanomaterials-Based Electrically Conductive Scaffolds to Repair the Ischaemic Heart Tissue" C 8, no. 4: 72. https://doi.org/10.3390/c8040072
APA StyleUl Haq, A., Carotenuto, F., Trovalusci, F., De Matteis, F., & Di Nardo, P. (2022). Carbon Nanomaterials-Based Electrically Conductive Scaffolds to Repair the Ischaemic Heart Tissue. C, 8(4), 72. https://doi.org/10.3390/c8040072