
Succinic Acid: Technology Development and Commercialization


Abstract
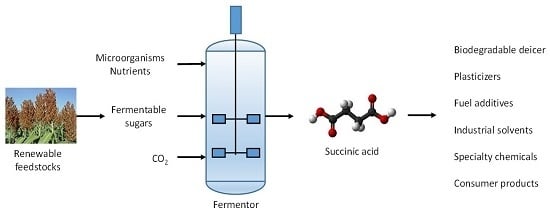
:
1. Introduction
2. Succinic Acid-Producing Microorganisms
3. Succinic Acid Recovery
4. Succinic Acid Applications
5. Commercialization of Bio-Based Succinic Acid
Conflicts of Interest
References
- Fumagalli, C. Succinic acid and succinic anhydride. In Kirk-Othmer Encyclopedia of Chemical Technology, 4th ed.; Kroschwitz, J.I., Howe-Grant, M., Eds.; Wiley: New York, NY, USA, 1997; Volume 22, pp. 1074–1102. [Google Scholar]
- Glassner, D.A.; Datta, R. Process for the Production and Purification of Succinic Acid. U.S. Patent 5143834, 1992. [Google Scholar]
- Guettler, M.V.; Jain, M.K. Method for Making Succinic Acid, Anaerobiospirillum Succiniciproducens Variants for Use in Process and Methods for Obtaining Variants. U.S. Patent 5521075, 1996. [Google Scholar]
- Global Succinic Acid Market Analysis & Trends 2013–2017 - Industry Forecast to 2025: $1.76 Billion Growth Opportunities/Investment Opportunities—Research and Markets. Available online: https://ceo.ca/@newswire/global-succinic-acid-market-analysis-trends-2013-2017 (accessed on 6 March 2017).
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass–Volume 1: Results of Screening for Potential Candidates from Sugars and Synthesis Gas; Office of Biomass Program, United States Department of Energy: Washington, DC, USA, 2004.
- Nghiem, N.P.; Hicks, K.B.; Johnston, D.B. Integration of succinic acid and ethanol production with potential application in a corn or barley biorefinery. Appl. Biochem. Biotechnol. 2010, 162, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- McKinlay, J.B.; Vieille, C.; Zeikus, J.G. Prospects for a bio-based succinate industry. Appl. Microbiol. Biotechnol. 2007, 76, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Samuelov, N.S.; Lamed, R.; Lowe, S.; Zeikus, J.G. Influence of CO2-HCO3−levels and pH on growth, succinate production, and enzyme activities of Anaerobiospirillum succiniciproducens. Appl. Environ. Microbiol. 1991, 57, 3013–3019. [Google Scholar] [PubMed]
- Guettler, M.V.; Rumler, D.; Jain, M.K. Actinobacillus succinogenes sp. nov., a novel succinic acid-producing strain from the bovine rumen. Int. J. Syst. Evol. Microbiol. 1999, 49, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Der Werf, M.J.V.; Guettler, M.V.; Jain, M.K.; Zeikus, J.G. Environmental and physiological factors affecting the succinate product ratio during carbohydrate fermentation by Actinobacillus sp. 130Z. Arch. Microbiol. 1997, 167, 332–342. [Google Scholar] [CrossRef]
- Zheng, P.; Fang, L.; Xu, Y.; Dong, J.J.; Ni, Y.; Sun, Z.H. Succinic acid production from corn stover by simultaneous saccharification and fermentation using Actinobacillus succinogenes. Bioresour. Technol. 2010, 101, 7889–7894. [Google Scholar] [CrossRef] [PubMed]
- McKinlay, J.B.; Laivenieks, M.; Schindler, B.D.; McKinlay, A.A.; Siddaramappa, S.; Challacombe, J.F.; Lowry, S.R.; Clum, A.; Lapidus, A.L.; Burkhart, K.B.; et al. A genomic perspective on the potential of Actinobacillus succinogenes for industrial succinate production. BMC Genom. 2010, 1, 680. [Google Scholar] [CrossRef] [PubMed]
- Jian, Y.; Kleff, S.; Guettler, M.V. Recombinant Microorganisms for Increased Production of Organic Acids. U.S. Patent 8431373, 2013. [Google Scholar]
- McKinlay, J.B.; Zeikus, J.G.; Vieille, C. Insights into Actinobacillus succinogenes fermentative metabolism in a chemically defined growth medium. Appl. Environ. Microbiol. 2005, 71, 6651–6656. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Song, H.; Lee, S.Y. Genome-based Metabolic Engineering of Mannheimia succiniciproducens for Succinic Acid Production. Appl. Environ. Microbiol. 2006, 72, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Kim, J.S.; Lee, S.Y.; In, Y.H.; Choi, S.S.; Rih, J.-K.; Kim, C.H.; Jeong, H.; Hur, C.G.; Kim, J.J. The genome sequence of the capnophilic rumen bacterium Mannheimia succiniciproducens. Nat. Biotechnol. 2004, 22, 1275–1281. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Kim, J.M.; Song, H.; Lee, J.W.; Kim, T.Y.; Jang, Y.-S. From genome sequence to integrated bioprocess for succinic acid production by Mannheimia succiniciproducens. Appl. Microbiol. Biotechnol. 2008, 79, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Kim, T.Y.; Choi, B.-K.; Choi, S.J.; Nielsen, L.K.; Chang, H.N.; Lee, S.Y. Development of a chemically defined medium for Mannheimia succiniciproducens based on its genome sequence. Appl. Microbiol. Biotechnol. 2008, 79, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, P.; Scholten, E.; Haefner, S.; Mayor, D.; Frey, J. Basfia succiniciproducens gen. nov., sp. nov., a new member of the family Pasteurellaceae isolated from bovine rumen. Int. J. Syst. Evol. Microbiol. 2010, 60, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.; Reinefeld, J.; Stellmacher, R.; Schaefer, R.; Lange, A.; Meyer, H.; Lalk, M.; Zelder, O.; Abendroth, G.; Schroeder, H.; et al. System-wide analysis and engineering of metabolic pathway fluxes in bio-succinate producing Basfia succiniciproducens. Biotechnol. Bioeng. 2013, 110, 3013–3023. [Google Scholar] [CrossRef] [PubMed]
- Scholten, E.; Renz, T.; Thomas, J. Continuous cultivation approach for fermentative succinic acid production from crude glycerol using Basfia succiniciproducens. Biotechnol. Lett. 2009, 31, 1947–1951. [Google Scholar] [CrossRef] [PubMed]
- Nghiem, N.P.; Donnelly, M.; Millard, C.S.; Stols, L. Method for the Production of Dicarboxylic Acids. U.S. Patent 5869301, 1999. [Google Scholar]
- Donnelly, M.I.; Sanville-Millard, C.Y.; Nghiem, N.P. Method to Produce Succinic Acid from Raw Hydrolysates. U.S. Patent 6743610, 2004. [Google Scholar]
- Stols, L.; Donnelly, M.I. Production of succinic acid through overexpression of NAD+-dependent malic enzyme in an Escherichia coli mutant. Appl. Environ. Microbiol. 1997, 63, 2695–2701. [Google Scholar] [PubMed]
- Gokarn, R.R.; Eiteman, M.A.; Altman, E. Expression of pyruvate carboxylase enhances succinate production in Escherichia coli without affecting glucose uptake. Biotechnol. Lett. 1998, 20, 795–798. [Google Scholar] [CrossRef]
- Vemuri, G.N.; Eiteman, M.A.; Altman, E. Effects of growth mode and pyruvate carboxylase on succinic acid production by metabolically engineered strains of Escherichia coli. Appl. Environ. Microbiol. 2002, 68, 1715–1727. [Google Scholar] [CrossRef] [PubMed]
- Vemuri, G.N.; Eiteman, M.A.; Altman, E. Succinate production in dual-phase Escherichia coli fermentations depends on the time of transition from aerobic to anaerobic conditions. J. Ind. Microbiol. Biotechnol. 2002, 28, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Bennett, G.N.; San, K.-Y. Metabolic engineering of aerobic succinate production systems in Escherichia coli to improve productivity and achieve the maximum theoretical succinate yield. Metab. Eng. 2005, 7, 116–127. [Google Scholar] [CrossRef] [PubMed]
- San, K.-Y.; Bennett, G.N.; Lin, H.; Sanchez, A. High Succinate Producing Bacteria. WO Patent application 2006034156 A2, 2006. [Google Scholar]
- Zhang, X.; Jantama, K.; Moore, J.C.; Jarboe, L.R.; Shanmugam, K.T.; Ingram, L.O. Metabolic evolution of energy-conserving pathways for succinate production in Escherichia coli. Proc. Nat. Acad. Sci. USA 2009, 106, 20180–20185. [Google Scholar] [CrossRef] [PubMed]
- Desai, T.A.; Rao, C.V. Regulation of Arabinose and Xylose metabolism in E. coli. Appl. Environ. Microbiol. 2010, 76, 1524–1532. [Google Scholar] [CrossRef] [PubMed]
- Okino, S.; Noburyu, R.; Suda, M.; Jojima, T.; Inui, M.; Yukawa, H. An efficient succinic acid production process in a metabolically engineered Corynebacterium glutamicum strain. Appl. Microbiol. Biotechnol. 2008, 81, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Yuzbachev, T.V.; Yuzbasheva, E.Y.; Laptev, I.A.; Sobolevskaya, T.I.; Vybornaya, T.V.; Larina, A.S.; Gvilava, I.T.; Antonova, S.V.; Sineoky, S.P. Is it possible to produce succinic acid at a low pH? Bioeng. Bugs 2011, 2, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Camarasa, C.; Grivet, J.-P.; Dequin, S. Investigation by 13C-NMR and tricarboxylic acid (TCA) deletion mutant analysis of pathways for succinate formation in Saccharomyces cerevisiae during anaerobic fermentation. Microbiology 2003, 149, 2669–2678. [Google Scholar] [CrossRef] [PubMed]
- Van Maris, A.J.; Geertman, J.-M.; Vermeulen, A.; Groothuizen, M.K.; Winkler, A.A.; Piper, M.D.W.; Van Dijken, J.P.; Pronk, J.T. Directed evolution of pyruvate decarboxylase-negative Saccharomyces cerevisiae yielding a C2-independent, glucose-tolerant, and pyruvate-hyperproducing yeast. Appl. Environ. Microbiol. 2004, 70, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Beauprez, J.J.; De Mey, M.; Soetaert, W.S. Microbial succinic acid production: Natural versus metabolic engineered producers. Process Biochem. 2010, 45, 1103–1114. [Google Scholar] [CrossRef]
- Schröder, H.; Haefner, S.; Von Abendroth, G.; Hollmann, R.; Raddatz, A.; Ernst, H.; Gurski, H. Microbial Succinic Acid Producers and Purification of Succinic Acid. U.S. Patent 9023632 B2, 2015. [Google Scholar]
- Krawczyk, J.M.; Haefner, S.; Schröder, H.; Costa, E.D.; Zelder, O.; Von Abendroth, G.; Wittmann, C.; Stellmacher, R.; Lange, A.; Lyons, B. J.; Lyons, T.J.; Crecy, E.; Hughes, E. Microorganisms for Succinic Acid Production. U.S. Patent application 20160348082, 2016. [Google Scholar]
- Dunuwila, D.; Cockrem, M. Methods and Systems of Producing Dicarboxylic Acids. U.S. Patent application 20120259138, 2012. [Google Scholar]
- Fruchey, O.S.; Manzer, L.E.; Dunuwila, D.; Keen, B.T.; Albin, B.A.; Clinton, N.A.; Dombek, B.D. Processes for the Production of Hydrogenated Products. U.S. Patent 8193375, 2012. [Google Scholar]
- Boit, B.; Fiey, G.; van, D.G.M.J. Process for Manufacturing Succinic Acid from A Fermentation broth Using Nano Filtration to Purify Recycled Mother Liquor. WO Patent application 2016087408, 2016. [Google Scholar]
- Mori, Y.; Takahashi, G.; Suda, H.; Yoshida, S. Processes for Producing Succinic Acid. U.S. Patent 9035095, 2015. [Google Scholar]
- Tosukhowong, T. A Process for Preparing Succinic Acid and Succinate Ester. WO Patent application 2015085198, 2015. [Google Scholar]
- Krieken, J.V.; Breugel, J.V. Process for the Preparation of A Monovalent Succinate Salt. U.S. Patent application 20110244534, 2011. [Google Scholar]
- De Jong, E.; Higson, A.; Walsh, P.; Wellisch, M. Bio-based Chemicals–Value Added Products from Biorefineries. IEA Bioenergy–Task 42 Biorefinery. Available online: http://www.ieabioenergy.com/wp-content/uploads/2013/10/Task-42-Biobased-Chemicals-value-added-products-from-biorefineries.pdf (accessed on 17 March 2017).
- Bio-conversion and Separation Technology (2013). WP 8.1. Determination of Market Potential for Selected Platform Chemicals–Itaconic acid, Succinic acid, 2,5-Furandicarboxylic acid. Available online: http://www.bioconsept.eu/wp-content/uploads/BioConSepT_Market-potential-for-selected-platform-chemicals_report1.pdf (accessed on 17 March 2017).
- Varadarajan, S.; Miller, D.J. Catalytic upgrading of fermentation-derived organic acids. Biotechnol. Prog. 1999, 15, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Delhomme, C.; Weuster-Boltz, D.; Kühn, F.E. Succinic acid from renewable resources as a C4 building-block chemical—A review of the catalytic possibilities in aqueous media. Green Chem. 2008, 11, 13–26. [Google Scholar] [CrossRef]
- Burgard, A.P.; van Dien, S.J.; Burk, M. Methods and Organisms for the Growth-Coupled Production of 1,4-butanediol. U.S. Patent 7947483, 2011. [Google Scholar]
- Berglund, K.A.; Alizadeh, H.; Dunuwila, D.D. Deicing Compositions and Methods of Use. U.S. Patent 6287480, 2001. [Google Scholar]
- Facklam, T. Mixed Alkyl Benzyl Esters of Succinic Acid Used as Plasticizers. US Patent 9080032, 2015. [Google Scholar]
- Broz, J.; Seon, A.; Simoes-Nunes, C. Use of Succinic Acid. U.S. Patent application 20110189347, 2011. [Google Scholar]
- Tosukhowong, T. A Process for Preparing Succinic Acid and Succinate Ester. U.S. Patent application 20160304431, 2016. [Google Scholar]
- Rush, B.J.; Watts, K.T.; McIntosh, V.L., Jr.; Fosmer, A.M.; Poynter, G.M.; McMullin, T.W. Yeast Cells Having Reductive TCA Pathway from Pyruvate to Succinate and Overexpressing An Exogenous NAD(P)+ Transhydrogenase Enzyme. U.S. Patent application 20150203877, 2015. [Google Scholar]
- BioAmber website. Available online: https://www.bio-amber.com/bioamber/en/company (accessed on 20 March 2017).
- Jantama, K.; Haupt, M.J.; Svoronos, S.A.; Zhang, X.; Moore, J.C.; Shanmugham, K.T.; Ingram, L.O. Combining metabolic engineeringand metabolic evolutions to develop nonrecombinant strains of Escherichia coli C that produce succinate and malate. Biotechnol. Bioeng. 2008, 99, 1140–1153. [Google Scholar] [CrossRef] [PubMed]
- Jantama, K.; Zhang, X.; Moore, J.C.; Shanmugham, K.T.; Svoronos, S.A.; Ingram, L.O. Eliminating side products and increasing succinate yields in engineered strains of Escherichia coli C. Biotechnol. Bioeng. 2008, 101, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Grabar, T.; Gong, W.; Yocum, R. Metabolic Evolution of Escherichia coli Strains That Produce Organic Acids. U.S. Patent 8871489, 2014. [Google Scholar]
- Myriant Website. Available online: http://myriant.com/our-company/index.cfm (accessed on 20 March 2017).
- Myriant in Talks to Build Bio-BDO Plant in Asia. Available online: https://www.icis.com/resources/news/2013/07/02/9684067/myriant-in-talks-to-build-bio-bdo-plant-in-asia-exec/ (accessed on 20 March 2017).
- Reverdia Website. Available online: http://www.reverdia.com/ (accessed on 20 March 2017).
- Verwaal, R.; Wu, L.; Damveld, R.A.; Sagt, C.M.J. Dicarboxylic Acid Production in Eukaryotes. U.S. Patent 9340804, 2016. [Google Scholar]
- Jansen, M.L.A.; Van De Graaf, M.J.; Verwaal, R. Dicarboxylic acid production process. U.S. Patent application US 20120040422, 2012. [Google Scholar]
- Succinity Website. Available online: http://succinity.com/ (accessed 20 March 2017).
- Succinity Produces First Commercial Quantities of Biobased Succinic Acid. Available online: https://www.basf.com/en/company/news-and-media/news-releases/2014/03/p-14-0303-ci.html (accessed 20 March 2017).
- Moussa, H.I.; Elkamel, A.; Young, S.B. Assessing energy performance of bio-based succinic acid production using LCA. J. Clean. Prod. 2016, 139, 761–769. [Google Scholar] [CrossRef]
- Pinazo, J.M.; Domine, M.E.; Parvulescu, V.; Petru, F. Sustainability metrics for succinic acid production: A comparison between biomass-based and petrochemical routes. Catal. Today 2015, 239, 17–24. [Google Scholar] [CrossRef]



{kind=link}
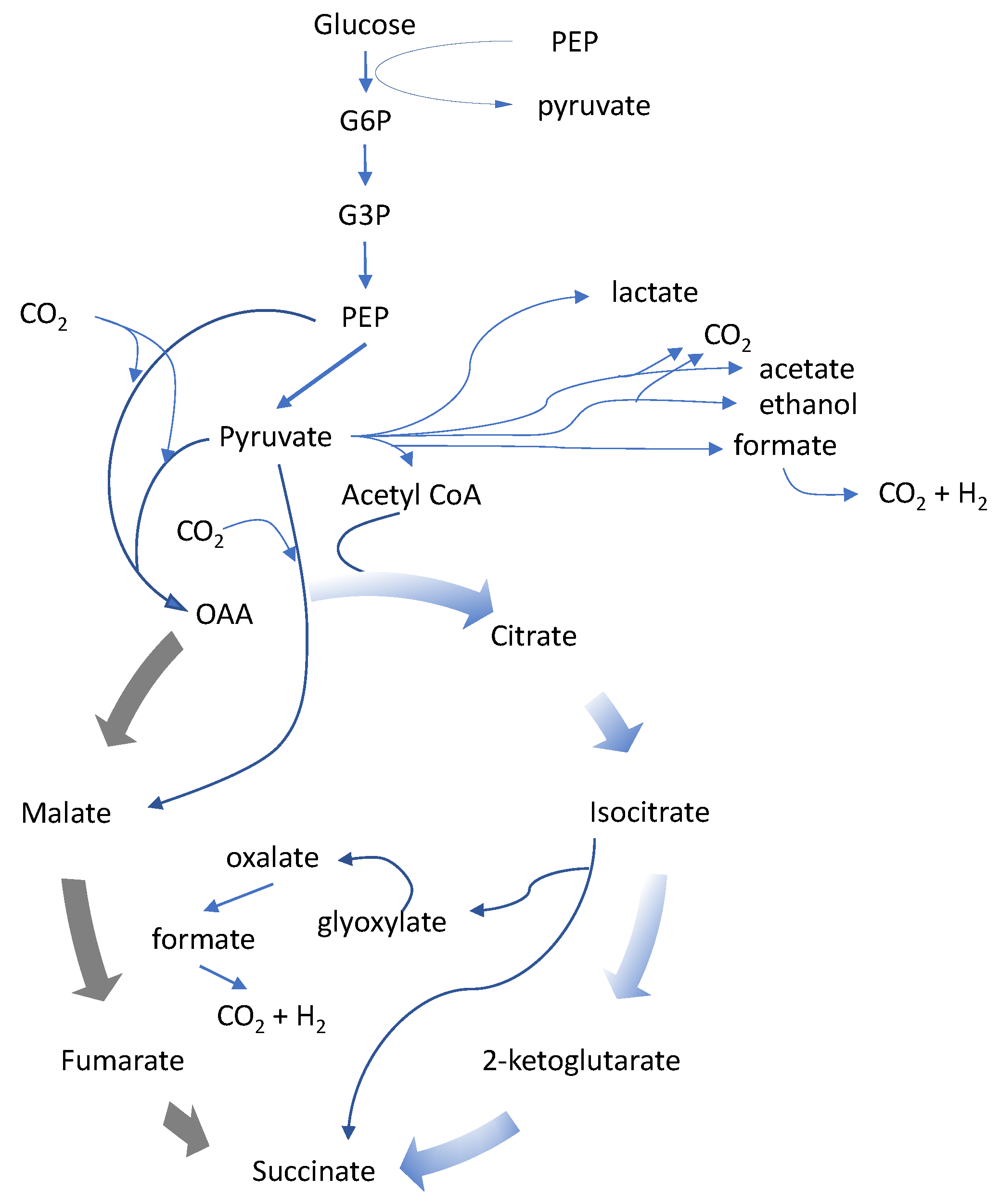{kind=link}
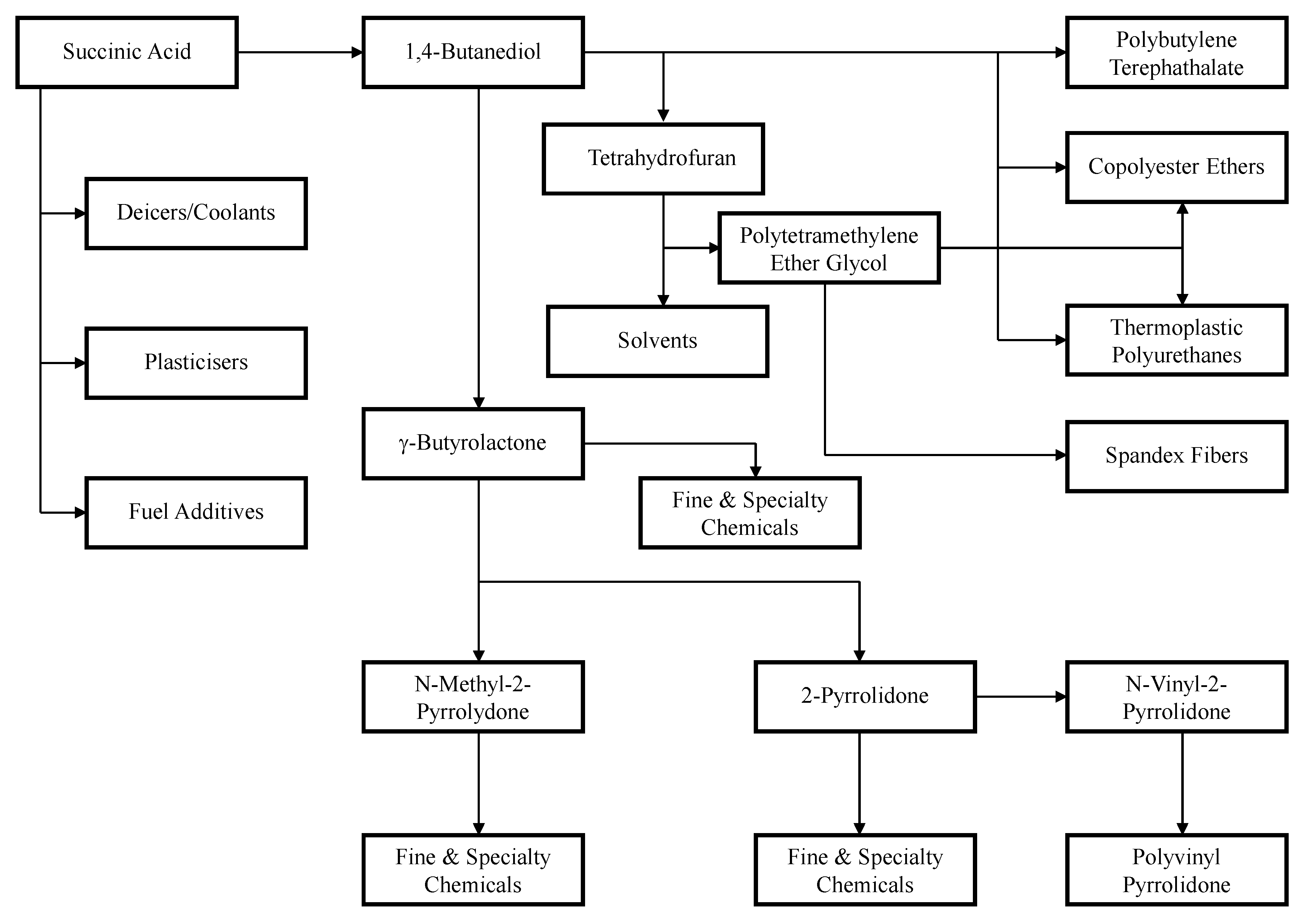{kind=link}
| Product Applications | Market Volume (1000 MT) | Market Share (%) |
|---|---|---|
| BDO 1 | 316 | 52.7 |
| PBS, PBST 2 | 82 | 13.7 |
| Polyester polyols | 51 | 8.5 |
| Plasticizers | 37 | 6.2 |
| Food | 26 | 4.3 |
| Pharmaceutical | 21 | 3.5 |
| Alkyd resins | 21 | 3.5 |
| Resins, coatings | 12 | 2.0 |
| Cosmetics | 12 | 2.0 |
| Solvents & lubricants | 6 | 1.0 |
| De-icer solutions | 3 | 0.5 |
| Others | 13 | 2.2 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nghiem, N.P.; Kleff, S.; Schwegmann, S. Succinic Acid: Technology Development and Commercialization. Fermentation 2017, 3, 26. https://doi.org/10.3390/fermentation3020026
Nghiem NP, Kleff S, Schwegmann S. Succinic Acid: Technology Development and Commercialization. Fermentation. 2017; 3(2):26. https://doi.org/10.3390/fermentation3020026
Chicago/Turabian StyleNghiem, Nhuan P., Susanne Kleff, and Stefan Schwegmann. 2017. "Succinic Acid: Technology Development and Commercialization" Fermentation 3, no. 2: 26. https://doi.org/10.3390/fermentation3020026
APA StyleNghiem, N. P., Kleff, S., & Schwegmann, S. (2017). Succinic Acid: Technology Development and Commercialization. Fermentation, 3(2), 26. https://doi.org/10.3390/fermentation3020026