Antifungal Screening of Bioprotective Isolates against Botrytis cinerea, Fusarium pallidoroseum and Fusarium moniliforme

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Pathogens
2.2. Bioprotective Isolates
2.3. Antifungal Screening
2.4. Biosurfactant Screening
2.5. Data Analysis
3. Results and Discussion
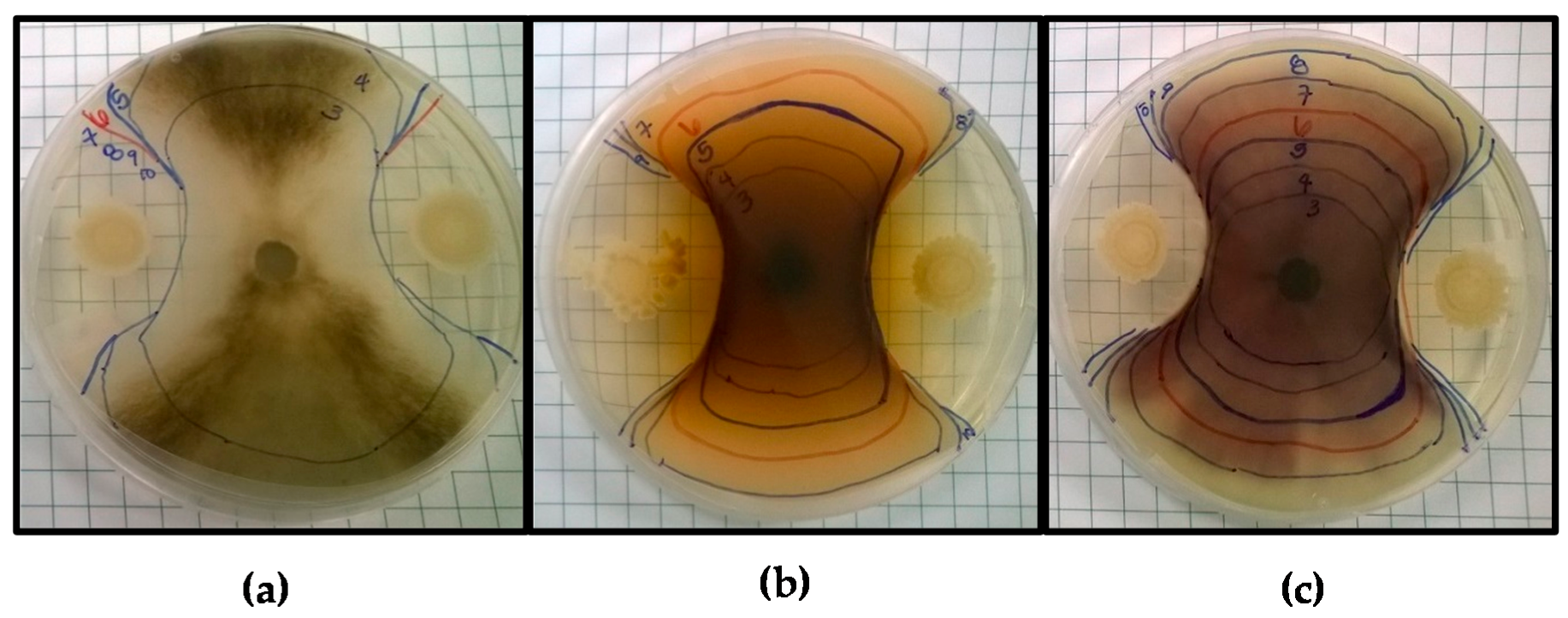
3.1. Antifungal Activity
3.1.1. Bacillus Species
3.1.2. Lactic Acid Bacteria
3.1.3. Serratia Plymuthica
3.1.4. Isolates with No Antifungal Activity
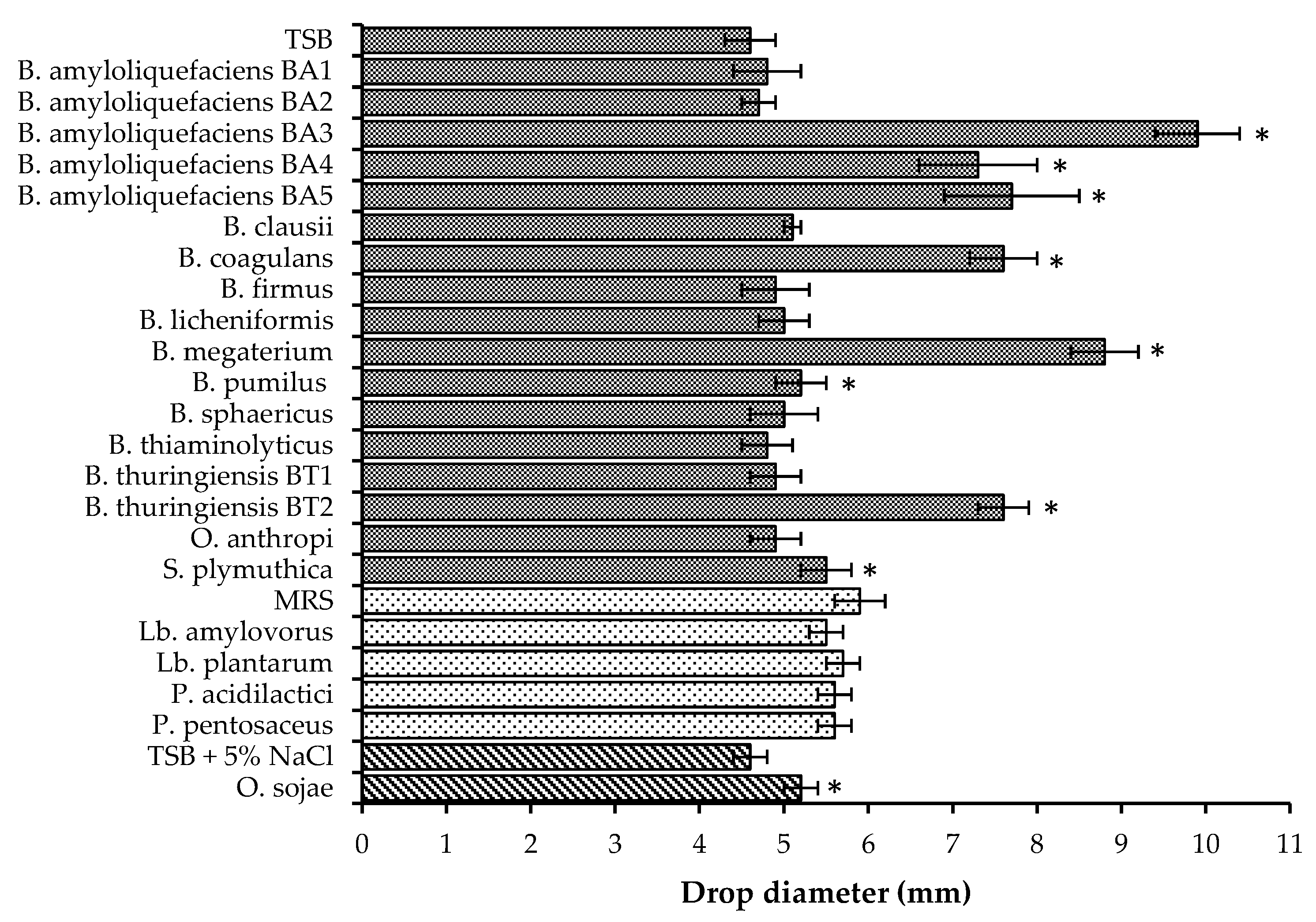
3.2. Biosurfactant Activity
3.2.1. Bacillus Species, Oceanobacillus sojae, and Ochrobactrum anthropi
3.2.2. Lactic Acid Bacteria Isolates
3.2.3. Serratia Plymuthica
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A.L. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Elmer, P.A.G.; Reglinski, T. Biosuppression of Botrytis cinerea in grapes. Plant Pathol. 2006, 55, 155–177. [Google Scholar] [CrossRef]
- Swartzberg, D.; Kirshner, B.; Rav-David, D.; Elad, Y.; Granot, D. Botrytis cinerea induces senescence and is inhibited by autoregulated expression of the IPT gene. Eur. J. Plant Pathol. 2007, 120, 289–297. [Google Scholar] [CrossRef]
- UC IPM: UC Management Guidelines for Fusarium Wilt on Strawberry. Available online: http://www.ipm.ucdavis.edu/PMG/r734101411.html (accessed on 21 June 2014).
- Extension FactSheet: Fusarium and Verticillium Wilts of Tomato, Potato, Pepper, and Eggplant. Available online: https://www.oardc.ohio-state.edu/sallymiller/extension/factsheets/fusariumtomato.pdf (accessed on 21 June 2014).
- Díaz Arias, M.M.; Leandro, L.F.; Munkvold, G.P. Aggressiveness of Fusarium species and impact of root infection on growth and yield of soybeans. Phytopathology 2013, 103, 822–832. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, J.P.F.; Placinta, C.M.; MacDonald, A.M.C. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Technol. 1999, 80, 183–205. [Google Scholar] [CrossRef]
- Peraica, M.; Radic, B.; Lucic, A.; Pavlovic, M. Toxic effects of mycotoxins in humans. Bull. World Health Organ. 1999, 77, 754–766. [Google Scholar] [PubMed]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Romero, D.; Vicente, A.; Rakotoaly, R.H.; Dufour, S.E.; Veening, J.W.; Arrebola, E.; Cazorla, F.M.; Kuipers, O.P.; Paquot, M.; Pérez-García, A. The iturin and fengycin families of lipopeptides are key factors in antagonism of Bacillus subtilis toward Podosphaera fusca. Mol Plant-Microbe Interact. 2007, 20, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Quan, C.; Wang, Y.; Wang, J.; Fan, S. Bacillus amyloliquefaciens Q-426 as a potential biocontrol agent against Fusarium oxysporum f. sp. spinaciae. J. Basic Microbiol. 2014, 54, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Arrebola, E.; Jacobs, R.; Korsten, L. Iturin A is the principal inhibitor in the biocontrol activity of Bacillus amyloliquefaciens PPCB004 against postharvest fungal pathogens. J. Appl. Microbiol. 2010, 108, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Vanittanakom, N.; Leoffler, W. Fengycin—A novel antifungal lipopeptide antibiotic produced by Bacillus subtilis F-29-3. J. Antibiot. 1986, 39, 888–901. [Google Scholar] [CrossRef] [PubMed]
- Moyne, A.; Shelby, R.; Cleveland, T.E.; Tuzun, S. Bacillomycin D: An iturin with antifungal activity against Aspergillus flavus. J. Appl. Microbiol. 2001, 90, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Mahata, D.; Paul, D.; Korpole, S.; Franco, O.L.; Mandal, S.M. Purification, biochemical characterization and self-assembled structure of a fengycin-like antifungal peptide from Bacillus thuringiensis strain SM1. Front. Microbiol. 2013, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.I.; Bai, H.; Bai, D.; Chae, H.; Chung, S.; Kim, Y.; Park, R.; Chi, Y.T. Purification and characterization of a lipopeptide produced by Bacillus thuringiensis CMB26. J. Appl. Microbiol. 2004, 97, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Ryan, L.A.M.; Zannini, E.; Dal Bello, F.; Pawlowska, A.; Koehler, P.; Arendt, E.K. Lactobacillus amylovorus DSM 19280 as a novel food-grade antifungal agent for bakery products. Int. J. Food Microbiol. 2011, 146, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Strom, K.; Sjogren, J.; Broberg, A.; Schnurer, J. Lactobacillus plantarum MiLAB 393 produces the antifungal cyclic dipeptides cyclo (L-Phe-L-Pro) and cyclo(L-Phe-trans-4-OH-L-Pro) and 3-phenyllactic acid. Appl. Environ. Microbiol. 2002, 68, 4322–4327. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, J.; Strom, K.; Roos, S.; Sjogren, J.; Schnurer, J. Broad and complex antifungal activity among environmental isolates of lactic acid bacteria. FEMS Microbiol. Lett. 2003, 219, 129–135. [Google Scholar] [CrossRef]
- Arguelles-Arias, A.; Ongena, M.; Halimi, B.; Lara, Y.; Brans, A.; Joris, B.; Fickers, P. Bacillus amyloliquefaciens GA1 as a source of potent antibiotics and other secondary metabolites for biocontrol of plant pathogens. Microbiol. Cell Fact. 2009, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Romano, A.; Vitullo, D.; Senatore, M.; Lima, G.; Lanzotti, V. Antifungal cyclic lipopeptides from Bacillus amyloliquefaciens strain BO5A. J. Nat. Prod. 2013, 76, 2019–2025. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Liu, X.; Li, C.; Tian, W.; Shen, Q.; Shen, B. Isolation of Bacillus amyloliquefaciens S20 and its application in control of eggplant bacterial wilt. J. Environ. Manag. 2014, 137, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Huszcza, E.; Burczyk, B. Surfactin isoforms from Bacillus coagulans. Z. Naturforsch. C 2006, 61, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Troyano Pueyo, M.; Bloch, C.; Carmona-Ribeiro, A.M.; di Mascio, P. Lipopeptides produced by a soil Bacillus megaterium strain. Microbiol. Ecol. 2009, 57, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Kalinovskaya, N.I.; Kuznetsova, T.A.; Ivanova, E.P.; Romanenko, L.A.; Voinov, V.G.; Huth, F.; Laatsch, H. Characterization of surfactin-like cyclic depsipeptides synthesized by Bacillus pumilus from Ascidian Halocynthia aurantium. Mar. Biotechnol. 2002, 4, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Makovitzki, A.; Shai, Y. pH-dependent antifungal lipopeptides and their plausible mode of action. Biochemistry 2005, 44, 9775–9784. [Google Scholar] [CrossRef] [PubMed]
- Bressan, W.; Figueiredo, J.E.F. Chitinolytic Bacillus spp. isolates antagonistic to Fusarium moniliforme in maize. J. Plant Pathol. 2010, 92, 343–347. [Google Scholar]
- Prabhukarthikeyan, R.; Saravanakumar, D.; Raguchander, T. Combination of endophytic Bacillus and Beauveria for the management of Fusarium wilt and fruit borer in tomato. Pest Manag. Sci. 2014, 70, 1742–1750. [Google Scholar] [CrossRef] [PubMed]
- Ilhan, K.; Karabulut, O.A. Efficacy and population monitoring of bacterial antagonists for gray mold (Botrytis cinerea Pers. ex. Fr.) infecting strawberries. Biocontrol 2013, 58, 457–470. [Google Scholar] [CrossRef]
- Donmez, M.F.; Esitken, A.; Yildiz, H.; Ercisli, S. Biocontrol of Botrytis cinerea on strawberry fruit by plant growth promoting bacteria. J. Anim. Plant Sci. 2011, 21, 758–763. [Google Scholar]
- Mari, M.; Guizzardi, M.; Brunelli, M.; Folchi, A. Postharvest biological control of grey mould (Botrytis cinerea Pers.: Fr.) on fresh-market tomatoes with Bacillus amyloliquefaciens. Crop Prot. 1996, 15, 699–705. [Google Scholar] [CrossRef]
- Mari, M.; Guizzardi, M.; Pratella, G.C. Biological control of Gray Mold in pears by antagonistic bacteria. Biol. Control. 1996, 7, 30–37. [Google Scholar] [CrossRef]
- Sathe, S.J.; Nawani, N.N.; Dhakephalkar, P.K.; Kapadnis, B.P. Antifungal lactic acid bacteria with potential to prolong shelf-life of fresh vegetables. J. Appl. Microbiol. 2007, 103, 2622–2628. [Google Scholar] [CrossRef] [PubMed]
- Crowley, S.; Mahony, J.; van Sinderen, D. Broad-spectrum antifungal-producing lactic acid bacteria and their application in fruit models. Folia Microbiol. Praha 2013, 58, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Serenade Optimum (130205). Available online: https://greenbook-assets.s3.amazonaws.com/L1302.05A.pdf (accessed on 19 June 2014).
- Bayer CropScience Safety Data Sheet, Sonata. Available online: http://ag.wilburellis.com/Products/Product%20Documents/Actives/SDS/SONATA-SDS%20US-Default.pdf (accessed on 19 June 2014).
- DoubleNickel LC Biofungicide. Available online: http://www.certisusa.com/pdf-labels/DoubleNickel_LC_label.pdf (accessed on 19 June 2014).
- Turgrul, T.; Cansunar, E. Detecting surfactant-producing microorganisms by the drop-collapse test. World J. Microbiol. Biotechnol. 2005, 21, 851–853. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Zheng, L.; Yang, X.; Liu, H.; Guo, J. Isolation and characterization of an antifungal protein from Bacillus licheniformis HS10. Biochem. Biophys. Res. Commun. 2014, 454, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, E.Z. Chitinase production by Bacillus thuringiensis and Bacillus licheniformis: Their potential in antifungal biocontrol. J. Microbiol. 2012, 50, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Tendulkar, S.R.; Saikumari, Y.K.; Patel, V.; Raghotama, S.; Munshi, T.K.; Balaram, P.; Chattoo, B.B. Isolation, purification and characterization of an antifungal molecule produced by Bacillus licheniformis BC98, and its effect on phytopathogen Magnaporthe grisea. J. Appl. Microbiol. 2007, 103, 2331–2339. [Google Scholar] [CrossRef] [PubMed]
- Saidi, N.; Kouki, S.; Hiri, F.M.; Hajlaoui, M.R.; Mahrouk, M.; Ouzari, H.; Jedidi, N.; Hassen, A. Characterization and selection of Bacillus sp. strains, effective biocontrol agents against Fusarium oxysporum f. sp. radicis-lycopersici, the causal agent of Fusarium crown and root rot in tomato. Ann. Microbiol. 2009, 59, 191–198. [Google Scholar] [CrossRef]
- Agarry, O.O.; Akinyosoye, F.A.; Adetuyi, F.C. Antagonistic properties of microogranisms associated with cassava (Manihot esculenta Crantz) products. Afr. J. Biotechnol. 2005, 4, 627–632. [Google Scholar] [CrossRef]
- Chowdappa, P.; Mohan Kumar, S.P.; Jyothi Lakshmi, M.; Upreti, K.K. Growth stimulation and induction of systemic resistance in tomato against early and late blight by Bacillus subtilis OTPB1 or Trichoderma harzianum OTPB3. Biol. Control. 2013, 65, 109–117. [Google Scholar] [CrossRef]
- Alvindia, D.G.; Natsuaki, K.T. Biocontrol activities of Bacillus amyloliquefaciens DGA14 isolated from banana fruit surface against banana crown rot-causing pathogens. Crop Prot. 2009, 28, 236–242. [Google Scholar] [CrossRef]
- Cortés-Zavaleta, O.; López-Malo, A.; Hernández-Mendoza, A.; García, H.S. Antifungal activity of lactobacilli and its relationship with 3-phenyllactic acid production. Int. J. Food Microbiol. 2014, 173, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Voulgari, K.; Hatzikamari, M.; Delepoglou, A.; Georgakopoulos, P.; Litopoulou-Tzanetaki, E.; Tzanetakis, N. Antifungal activity of non-starter lactic acid bacteria isolates from dairy products. Food Control. 2010, 21, 136–142. [Google Scholar] [CrossRef]
- Gerez, C. L.; Torres, M.J.; Font de Valdez, G.; Rollán, G. Control of spoilage fungi by lactic acid bacteria. Biol. Control. 2013, 64, 231–237. [Google Scholar] [CrossRef]
- Suproniene, S.; Semaskiene, R.; Juodeikiene, G.; Mankeviciene, A.; Cizeikiene, D.; Vidmantiene, D. Seed treatment with lactic acid bacteria against seed-borne pathogens of spring wheat. Biocontrol. Sci. Technol. 2014, 25, 144–154. [Google Scholar] [CrossRef]
- Hamed, H.A.; Moustafa, Y.A.; Abdel-Aziz, S.M. In vivo efficacy of Lactic Acid Bacteria in biological control against Fusarium oxysporum for protection of tomato plant. Life Sci. J. 2011, 8, 462–468. [Google Scholar] [CrossRef]
- Trias, R.; Bañeras, L.; Montesinos, E.; Badosa, E. Lactic acid bacteria from fresh fruit and vegetables as biocontrol agents of phytopathogenic bacteria and fungi. Int. Microbiol. 2008, 11, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Kamensky, M.; Ovadis, M.; Chet, I.; Chernin, L. Soil-borne strain IC14 of Serratia plymuthica with multiple mechanisms of antifungal activity provides biocontrol of Botrytis cinerea and Sclerotinia sclerotiorum diseases. Soil Biol. Biochem. 2003, 35, 323–331. [Google Scholar] [CrossRef]
- Levenfors, J.J.; Hedman, R.; Thaning, C.; Gerhardson, B.; Welch, C.J. Broad-spectrum antifungal metabolites produced by the soil bacterium Serratia plymuthica A 153. Soil Biol. Biochem. 2004, 36, 677–685. [Google Scholar] [CrossRef]
- Gould, M.; Nelson, L.M.; Waterer, D.; Hynes, R.K. Biocontrol of Fusarium sambucinum, dry rot of potato, by Serratia plymuthica 506. Biocontrol. Sci. Technol. 2008, 18, 1005–1016. [Google Scholar] [CrossRef]
- Mandal, V.; Sen, S.K.; Mandal, N.C. Optimized culture conditions for bacteriocin production by Pediococcus acidilactici LAB 5 and its characterization. Indian J. Biochem. Biophys. 2008, 45, 106–110. [Google Scholar] [PubMed]
- Shih, I.L.; Kuo, C.Y.; Hsieh, F.C.; Kao, S.S.; Hsieh, C. Use of surface response methodology to optimize culture conditions for iturin A production by Bacillus subtilis in solid-state fermentation. J. Chin. Inst. Chem. Eng. 2008, 39, 635–643. [Google Scholar] [CrossRef]
- Hathout, Y.; Ho, Y.; Ryzhov, V.; Demirev, P.; Fenselau, C. Kurstakins: A new class of lipopeptides isolated from Bacillus thuringiensis. J. Nat. Prod. 2000, 63, 1492–1496. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Li, B.; Zhang, N.; Waseem, R.; Shen, Q.; Huang, Q. Production of bacillomycin- and macrolactin-type antibiotics by Bacillus amyloliquefaciens NJN-6 for suppressing soilborne plant pathogens. J. Agric. Food Chem. 2012, 60, 2976–2981. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.; Chen, X.H.; Vater, J.; Franke, P.; Nicholson, G.; Borriss, R.; Süssmuth, R.D. Macrolactin is the polyketide biosynthesis product of the pks2 cluster of Bacillus amyloliquefaciens FZB42. J. Nat. Prod. 2007, 70, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Lin, B.; Zhang, R. A novel antifungal protein of Bacillus subtilis B25. Springerplus 2013, 2, 543. [Google Scholar] [CrossRef] [PubMed]
- Chaiharn, M.; Chunhaleuchanon, S.; Lumyong, S. Screening siderophore producing bacteria as potential biological control agent for fungal rice pathogens in Thailand. World J. Microbiol. Biotechnol. 2009, 25, 1919–1928. [Google Scholar] [CrossRef]
- Aktuganov, G.E.; Melent’ev, A.I.; Kuz’mina, L.Y.; Galimzyanova, N.F.; Shirokov, A.V. The chitinolytic activity of Bacillus cohn bacteria antagonistic to phytopathogenic fungi. Microbiology 2003, 72, 313–317. [Google Scholar] [CrossRef]
- Gudiña, E.J.; Teixeira, J.A.; Rodrigues, L.R. Biosurfactant-producing lactobacilli: Screening, production profiles, and effect of medium composition. Appl. Environ. Soil Sci. 2011, 2011, 1–9. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Antimicrobial and anti-adhesive properties of biosurfactant produced by lactobacilli isolates, biofilm formation and aggregation ability. J. Gen. Appl. Microbiol. 2013, 59, 425–436. [Google Scholar] [CrossRef]
- Velraeds, M.M.; Van Der Mei, H.C.; Reid, G.; Busscher, H.J. Inhibition of initial adhesion of uropathogenic Enterococcus faecalis by biosurfactants from Lactobacillus isolates. Appl. Environ. Microbiol. 1996, 62, 1958–1963. [Google Scholar] [PubMed]

 , MRS
, MRS  , and TSB + 5% NaCl
, and TSB + 5% NaCl  . Asterisks indicate drop diameters significantly larger than that of the media control (α = 0.05).
, MRS , and TSB + 5% NaCl . Asterisks indicate drop diameters significantly larger than that of the media control (α = 0.05).
. Asterisks indicate drop diameters significantly larger than that of the media control (α = 0.05).
, MRS , and TSB + 5% NaCl . Asterisks indicate drop diameters significantly larger than that of the media control (α = 0.05).

{kind=link}
{kind=link}
| Isolate | Growth Medium | Incubation Temperature | Screening Medium |
|---|---|---|---|
| Bioprotective isolate | |||
| Bacillus amyloliquefaciens BA1 | TSB | 35 °C | PDA |
| Bacillus amyloliquefaciens BA2 | TSB | 35 °C | PDA |
| Bacillus amyloliquefaciens BA3 | TSB | 35 °C | PDA |
| Bacillus amyloliquefaciens BA4 | TSB | 35 °C | PDA |
| Bacillus amyloliquefaciens BA5 | TSB | 35 °C | PDA |
| Bacillus clausii | TSB | 30 °C | PDA |
| Bacillus coagulans | TSB | 35 °C | PDA |
| Bacillus firmus | TSB | 30 °C | TSA |
| Bacillus licheniformis | TSB | 35 °C | PDA |
| Bacillus megaterium | TSB | 35 °C | PDA |
| Bacillus pumilus | TSB | 35 °C | PDA |
| Bacillus sphaericus | TSB | 30 °C | PDA |
| Bacillus thiaminolyticus | TSB | 30 °C | TSA |
| Bacillus thuringiensis BT1 | TSB | 35 °C | PDA |
| Bacillus thuringiensis BT2 | TSB | 35 °C | PDA |
| Lactobacillus amylovorus | MRS | 35 °C | MRS |
| Lactobacillus plantarum | MRS | 35 °C | MRS |
| Oceanobacillus sojae | TSB w/5% NaCl (w/w) | 35 °C | TSA |
| Ochrobactrum anthropi | TSB | 35 °C | PDA |
| Pediococcus acidilactici | MRS | 35 °C | MRS |
| Pediococcus pentosaceus | MRS | 35 °C | MRS |
| Serratia plymuthica | TSB | 30 °C | PDA |
| Fungal phytopathogen | |||
| Botrytis cinerea | PDA | 24 °C | N/A 1 |
| Fusarium pallidoroseum | PDA | 24 °C | N/A 1 |
| Fusarium moniliforme | PDA | 24 °C | N/A 1 |
| Bioprotective Isolate | Inhibition | ||
|---|---|---|---|
| Botrytis cinerea | Fusarium pallidoroseum | Fusarium moniliforme | |
| B. amyloliquefaciens BA1 | 52% | 62% | 53% |
| B. amyloliquefaciens BA2 | 56% | 63% | 60% |
| B. amyloliquefaciens BA3 | 58% | 67% | 61% |
| B. amyloliquefaciens BA4 | 60% | 67% | 60% |
| B. amyloliquefaciens BA5 | 61% | 69% | 61% |
| B. clausii | 30% | N/I 1 | N/I 1 |
| B. coagulans | 62% | 66% | 59% |
| B. firmus | 49% | N/I 1 | N/I 1 |
| B. licheniformis | N/I 1 | N/I1 | N/I 1 |
| B. megaterium | 62% | 68% | 61% |
| B. pumilus | N/I 1 | N/I 1 | N/I 1 |
| B. sphaericus | N/I 1 | N/I 1 | N/I 1 |
| B. thiaminolyticus | 50% | N/I 1 | N/I 1 |
| B. thuringiensis BT1 | N/I 1 | N/I 1 | N/I 1 |
| B. thuringiensis BT2 | 56% | 68% | 58% |
| L. amylovorus | N/I 1 | N/I 1 | N/I 1 |
| L. plantarum | 36% | N/I 1 | N/I 1 |
| O. sojae | N/I 1 | N/I 1 | N/I 1 |
| O. anthropi | N/I 1 | N/I 1 | N/I 1 |
| P. acidilactici | 56% | N/I 1 | N/I 1 |
| P. pentosaceus | 46% | N/I 1 | N/I 1 |
| S. plymuthica | 51% | 60% | 40% |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Senna, A.; Lathrop, A. Antifungal Screening of Bioprotective Isolates against Botrytis cinerea, Fusarium pallidoroseum and Fusarium moniliforme. Fermentation 2017, 3, 53. https://doi.org/10.3390/fermentation3040053
De Senna A, Lathrop A. Antifungal Screening of Bioprotective Isolates against Botrytis cinerea, Fusarium pallidoroseum and Fusarium moniliforme. Fermentation. 2017; 3(4):53. https://doi.org/10.3390/fermentation3040053
Chicago/Turabian StyleDe Senna, Antoinette, and Amanda Lathrop. 2017. "Antifungal Screening of Bioprotective Isolates against Botrytis cinerea, Fusarium pallidoroseum and Fusarium moniliforme" Fermentation 3, no. 4: 53. https://doi.org/10.3390/fermentation3040053
APA StyleDe Senna, A., & Lathrop, A. (2017). Antifungal Screening of Bioprotective Isolates against Botrytis cinerea, Fusarium pallidoroseum and Fusarium moniliforme. Fermentation, 3(4), 53. https://doi.org/10.3390/fermentation3040053