Biological Production of 3-Hydroxypropionic Acid: An Update on the Current Status




Abstract
:
1. Introduction
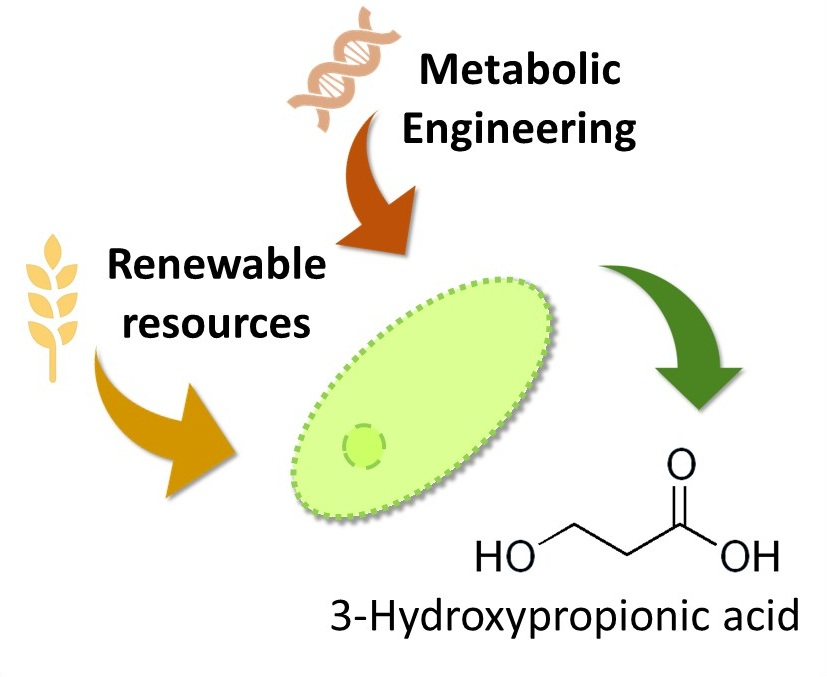
2. Production of 3HP from Glycerol
2.1. Production Using Genes That Are Part of the Dha Operon
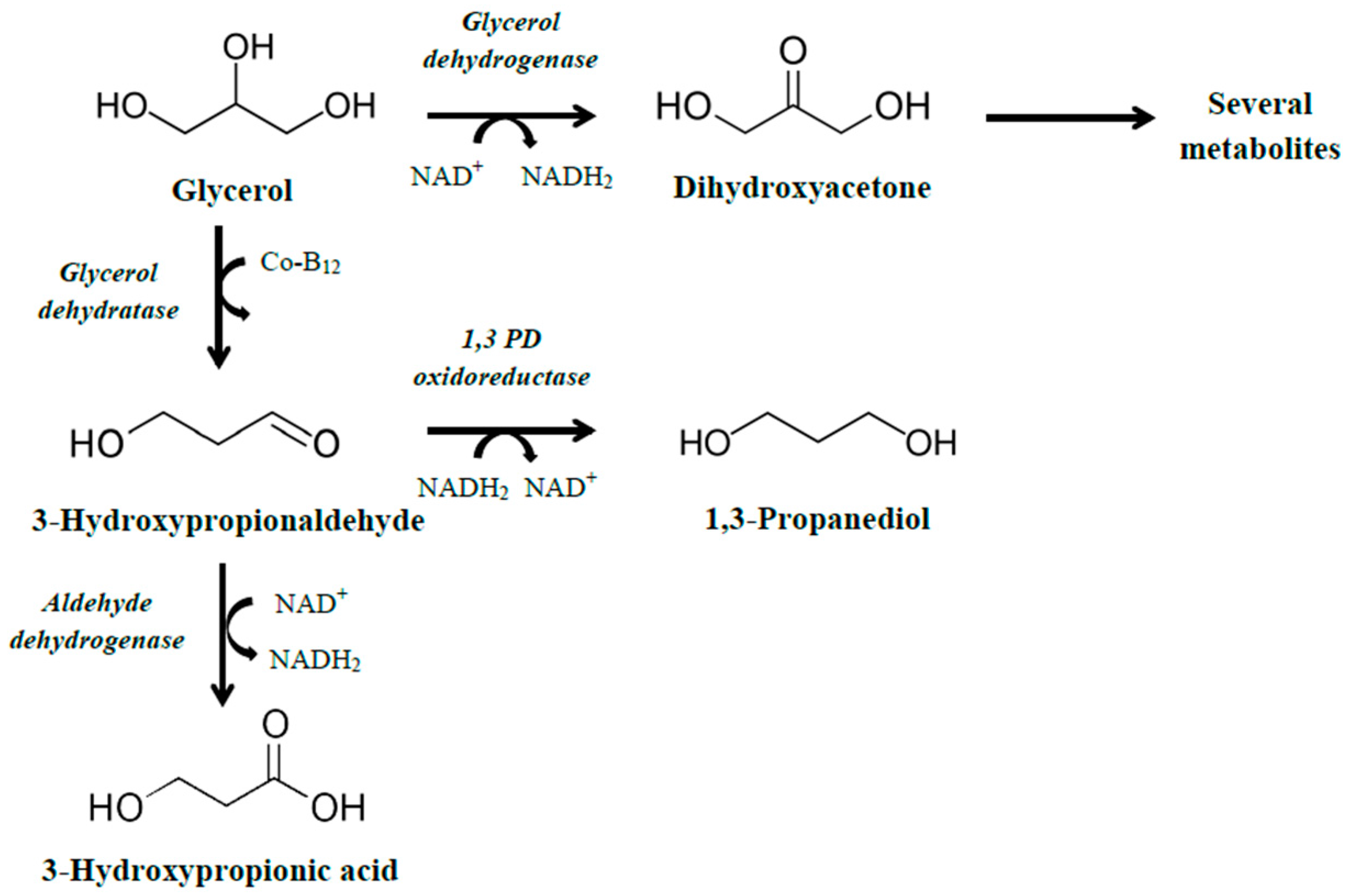
2.2. Production Using Genes That Are Part of the Pdu Operon
2.3. Engineering of K. pneumoniae Cells for the Conversion of Glycerol to 3HP
2.4. Engineering of E. coli Cells for the Conversion of Glycerol to 3HP
2.5. Other Microorganisms
3. Production of 3HP from Sugars
3.1. Pathways for the Conversion of Sugars (Mainly Glucose) to 3HP
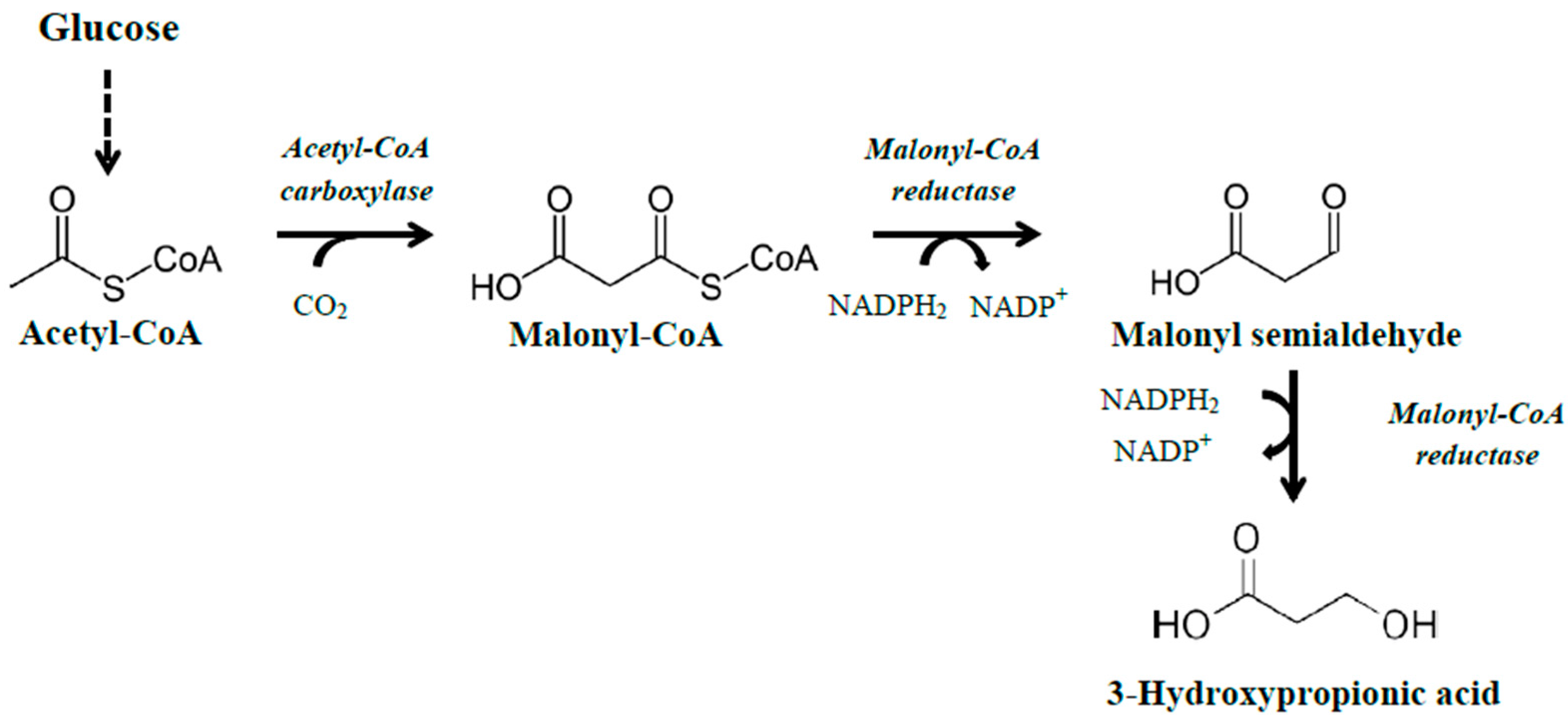
3.1.1. The Malonyl-CoA Pathway
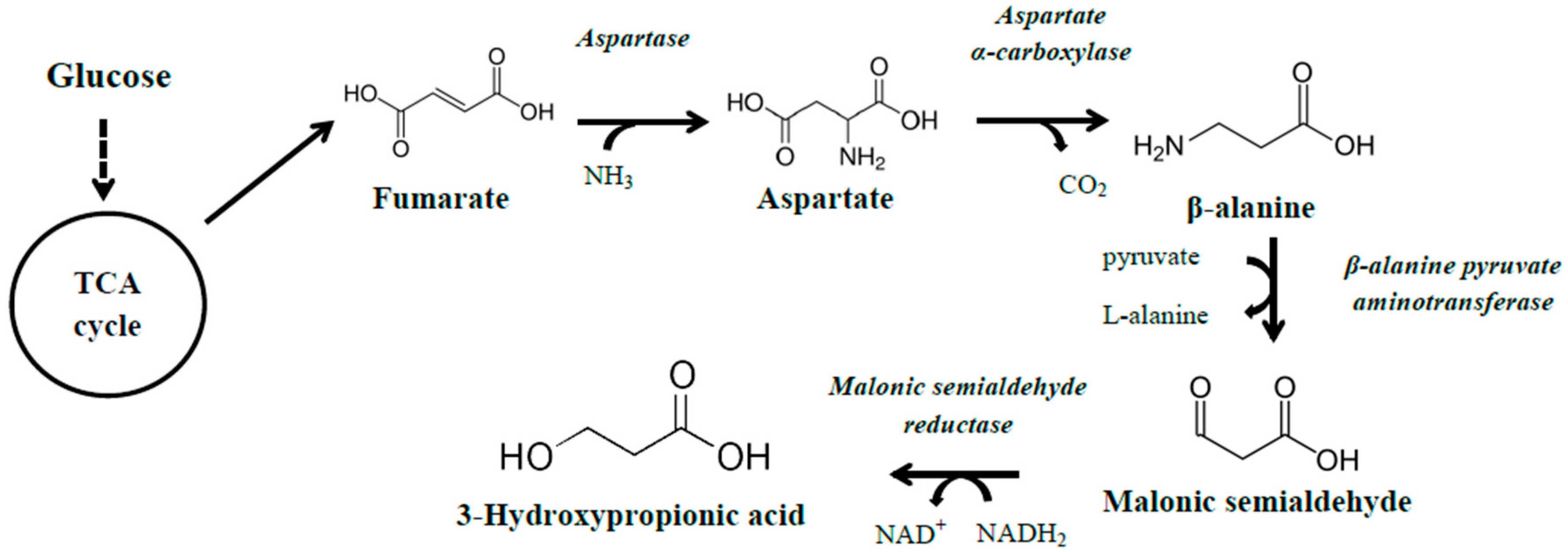
3.1.2. The β-Alanine Pathway
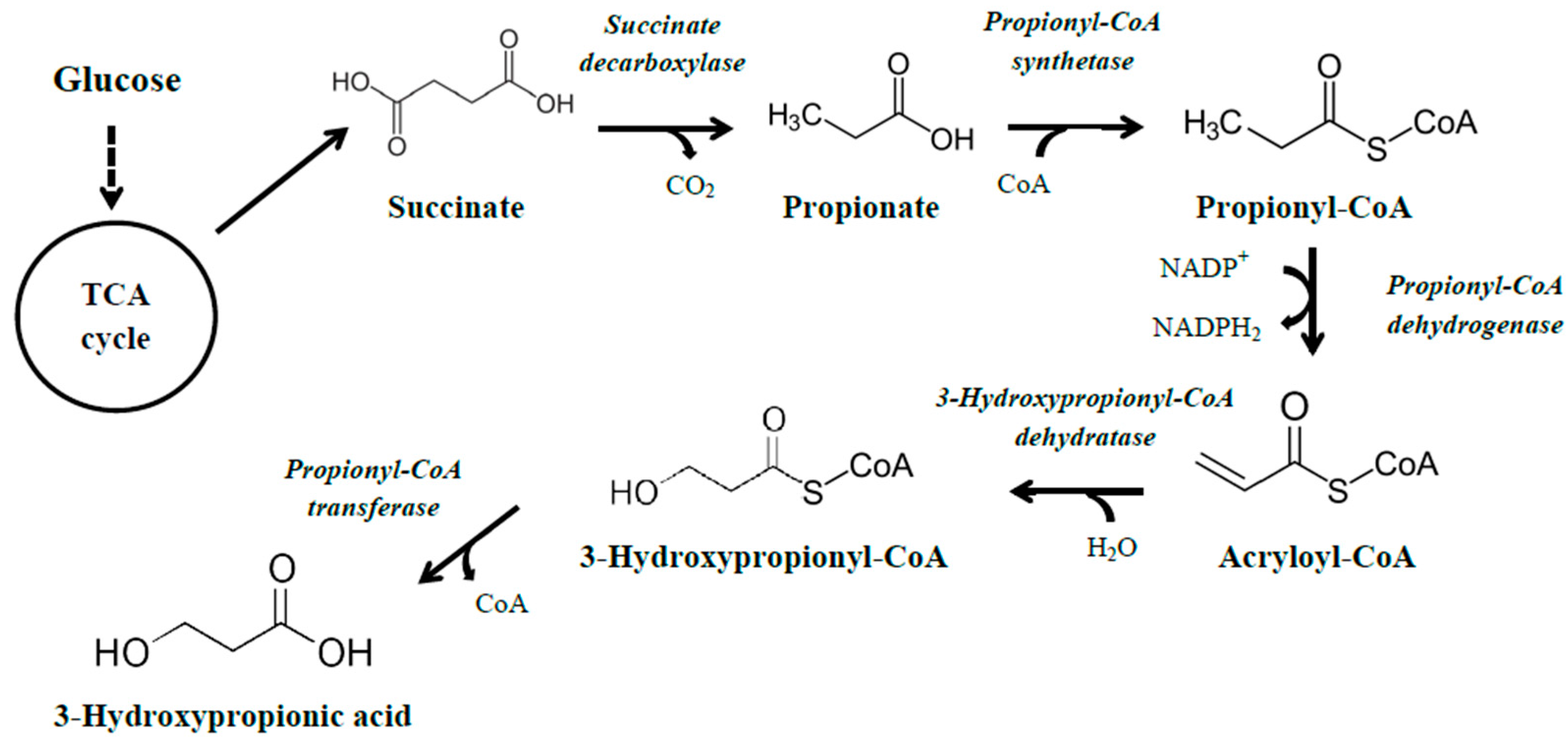
3.1.3. The Propionyl-CoA Pathway
3.1.4. The Glycerate Pathway
3.1.5. The Lactate Pathway
3.1.6. Use of a Glycerol Intermediate
3.2. Use of E. coli as a Host Microorganism
3.3. Use of S. cerevisiae as a Host Microorganism
3.4. Use of Other Host Microorganisms
4. Production of 3HP from Other Sources
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Werpy, T.; Petersen, G.; Aden, A.; Bozell, J.; Holladay, J.; White, J.; Manheim, A.; Eliot, D.; Lasure, L.; Jones, S. Top Value Added Chemicals from Biomass. Volume 1-Results of Screening for Potential Candidates from Sugars and Synthesis Gas; Department of Energy: Washington, DC, USA, 2004.
- Rathnasingh, C.; Raj, S.M.; Jo, J.E.; Park, S. Development and evaluation of efficient recombinant Escherichia coli strains for the production of 3-hydroxypropionic acid from glycerol. Biotechnol. Bioeng. 2009, 104, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Matsakas, L.; Topakas, E.; Christakopoulos, P. New trends in microbial production of 3-hydroxypropionic acid. Curr. Biochem. Eng. 2015, 1, 141–154. [Google Scholar] [CrossRef]
- Jung, W.S.; Kang, J.H.; Chu, H.S.; Choi, I.S.; Cho, K.M. Elevated production of 3-hydroxypropionic acid by metabolic engineering of the glycerol metabolism in Escherichia coli. Metab. Eng. 2014, 23, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Matsakas, L.; Giannakou, M.; Vörös, D. Effect of synthetic and natural media on lipid production from Fusarium oxysporum. Electron. J. Biotechnol. 2017, 30, 95–102. [Google Scholar] [CrossRef]
- Yang, F.; Hanna, M.A.; Sun, R. Value-added uses for crude glycerol—A byproduct of biodiesel production. Biotechnol. Biofuels 2012, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Okoye, P.U.; Hameed, B.H. Review on recent progress in catalytic carboxylation and acetylation of glycerol as a byproduct of biodiesel production. Renew. Sustain. Energy Rev. 2016, 53, 558–574. [Google Scholar] [CrossRef]
- He, Q.; McNutt, J.; Yang, J. Utilization of the residual glycerol from biodiesel production for renewable energy generation. Renew. Sustain. Energy Rev. 2017, 71, 63–76. [Google Scholar] [CrossRef]
- Rodrigues, A.; Bordado, J.C.; Dos Santos, R.G. Upgrading the glycerol from biodiesel production as a source of energy carriers and chemicals—A technological review for three chemical pathways. Energies 2017, 10, 1817. [Google Scholar] [CrossRef]
- Garlapati, V.K.; Shankar, U.; Budhiraja, A. Bioconversion technologies of crude glycerol to value added industrial products. Biotechnol. Rep. 2016, 9, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Ashok, S.; Raj, S.M.; Rathnasingh, C.; Park, S. Development of recombinant Klebsiella pneumoniae ΔdhaT strain for the co-production of 3-hydroxypropionic acid and 1,3-propanediol from glycerol. Appl. Microbiol. Biotechnol. 2011, 90, 1253–1265. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.; Seol, E.; Sundara Sekar, B.; Kwon, S.; Lee, J.; Park, S. Metabolic engineering of Klebsiella pneumoniae J2B for co-production of 3-hydroxypropionic acid and 1,3-propanediol from glycerol: Reduction of acetate and other by-products. Bioresour. Technol. 2017, 244, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xiu, Z.; Wang, J.; Zhang, D.; Xu, P. Stoichiometric analysis and experimental investigation of glycerol bioconversion to 1,3-propanediol by Klebsiella pneumoniae under microaerobic conditions. Enzyme Microb. Technol. 2003, 33, 386–394. [Google Scholar] [CrossRef]
- Zaushitsyna, O.; Dishisha, T.; Hatti-Kaul, R.; Mattiasson, B. Crosslinked, cryostructured Lactobacillus reuteri monoliths for production of 3-hydroxypropionaldehyde, 3-hydroxypropionic acid and 1,3-propanediol from glycerol. J. Biotechnol. 2017, 241, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.; Ashok, S.; Ainala, S.K.; Sankaranarayanan, M.; Chun, A.Y.; Jung, G.Y.; Park, S. Coenzyme B12 can be produced by engineered Escherichia coli under both anaerobic and aerobic conditions. Biotechnol. J. 2014, 9, 1526–1535. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.G.; Roth, J.R. Evolution of coenzyme B12 synthesis among enteric bacteria: Evidence for loss and reacquisition of a multigene complex. Genetics 1996, 142, 11–24. [Google Scholar] [PubMed]
- Toraya, T. Radical catalysis in coenzyme B12-dependent isomerization (eliminating) reactions. Chem. Rev. 2003, 103, 2095–2127. [Google Scholar] [CrossRef] [PubMed]
- Daniel, R.; Bobik, T.A.; Gottschalk, G. Biochemistry of coenzyme B12-dependent glycerol and diol dehydratases and organization of the encoding genes. FEMS Microbiol. Rev. 1998, 22, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.L.; Xu, X.L.; Li, C.; Ma, B. Bin Cloning, expression and reactivating characterization of glycerol dehydratase reactivation factor from Klebsiella pneumoniae XJPD-Li. World J. Microbiol. Biotechnol. 2009, 25, 1947–1953. [Google Scholar] [CrossRef]
- Ashok, S.; Sankaranarayanan, M.; Ko, Y.; Jae, K.E.; Ainala, S.K.; Kumar, V.; Park, S. Production of 3-hydroxypropionic acid from glycerol by recombinant Klebsiella pneumoniae ΔdhaTΔyqhD which can produce vitamin B12 naturally. Biotechnol. Bioeng. 2013, 110, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.H.; Seo, J.W.; Baek, J.O.; Oh, B.R.; Heo, S.Y.; Hong, W.K.; Kim, D.H.; Kim, C.H. Identification and characterization of the propanediol utilization protein PduP of Lactobacillus reuteri for 3-hydroxypropionic acid production from glycerol. Appl. Microbiol. Biotechnol. 2011, 89, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Bobik, T.A.; Havemann, G.D.; Busch, R.J.; Williams, D.S.; Aldrich, H.C. The propanediol utilization (pdu) operon of Salmonella enterica serovar Typhimurium LT2 includes genes necessary for formation of polyhedral organelles involved in coenzyme B12-dependent 1,2-propanediol degradation. J. Bacteriol. 1999, 181, 5967–5975. [Google Scholar] [PubMed]
- Dishisha, T.; Pereyra, L.P.; Pyo, S.H.; Britton, R.A.; Hatti-Kaul, R. Flux analysis of the Lactobacillus reuteri propanediol-utilization pathway for production of 3-hydroxypropionaldehyde, 3-hydroxypropionic acid and 1,3-propanediol from glycerol. Microb. Cell Fact. 2014, 13. [Google Scholar] [CrossRef] [PubMed]
- Johnsont, E.A.; Lin, E.C.C. Klebsiella pneumoniae 1,3-Propanediol:NAD+ Oxidoreductase. J. Bacteriol. 1987, 169, 2050–2054. [Google Scholar] [CrossRef]
- Wang, K.; Tian, P. Engineering plasmid-free Klebsiella pneumoniae for production of 3-hydroxypropionic acid. Curr. Microbiol. 2017, 74, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [PubMed]
- Su, M.; Li, Y.; Ge, X.; Tian, P. 3-Hydroxypropionaldehyde-specific aldehyde dehydrogenase from Bacillus subtilis catalyzes 3-hydroxypropionic acid production in Klebsiella pneumoniae. Biotechnol. Lett. 2015, 37, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, Z.; Shimizu, K.; Ye, Q. Co-production of 3-hydroxypropionic acid and 1,3-propanediol by Klebseilla pneumoniae expressing aldH under microaerobic conditions. Bioresour. Technol. 2013, 128, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, X.; Ge, X.; Tian, P. High production of 3-hydroxypropionic acid in Klebsiella pneumoniae by systematic optimization of glycerol metabolism. Sci. Rep. 2016, 6, 26932. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Lin, J.; Wang, H.; Xie, J.; Wei, D. Development of a two-step process for production of 3-hydroxypropionic acid from glycerol using Klebsiella pneumoniae and Gluconobacter oxydans. Bioprocess Biosyst. Eng. 2015, 38, 2487–2495. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.; Cheng, X.L.; Qin, H.B.; Liu, J.S.; Zheng, Y.G. Investigation of the key factors on 3-hydroxypropionic acid production with different recombinant strains. 3 Biotech 2017, 7, 314. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Ashok, S.; Park, S. Recent advances in biological production of 3-hydroxypropionic acid. Biotechnol. Adv. 2013, 31, 945–961. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.G.; Noh, M.H.; Jeong, J.H.; Park, S.; Jung, G.Y. Optimum rebalancing of the 3-hydroxypropionic acid production pathway from glycerol in Escherichia coli. ACS Synth. Biol. 2016, 5, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.S.; Kim, Y.S.; Lee, C.M.; Lee, J.H.; Jung, W.S.; Ahn, J.H.; Song, S.H.; Choi, I.S.; Cho, K.M. Metabolic engineering of 3-hydroxypropionic acid biosynthesis in Escherichia coli. Biotechnol. Bioeng. 2015, 112, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, M.; Somasundar, A.; Seol, E.; Chauhan, A.S.; Kwon, S.; Jung, G.Y.; Park, S. Production of 3-hydroxypropionic acid by balancing the pathway enzymes using synthetic cassette architecture. J. Biotechnol. 2017, 259, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.; Xiong, T.; Qin, H.B.; Wu, H.; Liu, Z.Q.; Zheng, Y.G. 3-Hydroxypropionic acid production by recombinant Escherichia coli ZJU-3HP01 using glycerol–glucose dual-substrate fermentative strategy. Biotechnol. Appl. Biochem. 2017, 64, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, M.; Ashok, S.; Park, S. Production of 3-hydroxypropionic acid from glycerol by acid tolerant Escherichia coli. J. Ind. Microbiol. Biotechnol. 2014, 41, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Tsuruno, K.; Honjo, H.; Hanai, T. Enhancement of 3-hydroxypropionic acid production from glycerol by using a metabolic toggle switch. Microb. Cell Fact. 2015, 14, 155. [Google Scholar] [CrossRef] [PubMed]
- Honjo, H.; Tsuruno, K.; Tatsuke, T.; Sato, M.; Hanai, T. Dual synthetic pathway for 3-hydroxypropionic acid production in engineered Escherichia coli. J. Biosci. Bioeng. 2015, 120, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Amin, H.M.; Hashem, A.M.; Ashour, M.S.; Hatti-Kaul, R. 1,2 Propanediol utilization by Lactobacillus reuteri DSM 20016, role in bioconversion of glycerol to 1,3 propanediol, 3-hydroxypropionaldehyde and 3-hydroxypropionic acid. J. Genet. Eng. Biotechnol. 2013, 11, 53–59. [Google Scholar] [CrossRef]
- Dishisha, T.; Pyo, S.H.; Hatti-Kaul, R. Bio-based 3-hydroxypropionic- and acrylic acid production from biodiesel glycerol via integrated microbial and chemical catalysis. Microb. Cell Fact. 2015, 14, 200. [Google Scholar] [CrossRef] [PubMed]
- Kalantari, A.; Chen, T.; Ji, B.; Stancik, I.A.; Ravikumar, V.; Franjevic, D.; Saulou-Bérion, C.; Goelzer, A.; Mijakovic, I. Conversion of glycerol to 3-hydroxypropanoic acid by genetically engineered Bacillus subtilis. Front. Microbiol. 2017, 8, 638. [Google Scholar] [CrossRef] [PubMed]
- Sjöblom, M.; Matsakas, L.; Krige, A.; Rova, U.; Christakopoulos, P. Direct electricity generation from sweet sorghum stalks and anaerobic sludge. Ind. Crops Prod. 2017, 108, 505–511. [Google Scholar] [CrossRef]
- Matsakas, L.; Nitsos, C.; Vörös, D.; Rova, U.; Christakopoulos, P. High-titer methane from organosolv-pretreated spruce and birch. Energies 2017, 10, 263. [Google Scholar] [CrossRef]
- Nitsos, C.; Matsakas, L.; Triantafyllidis, K.; Rova, U.; Christakopoulos, P. Investigation of different pretreatment methods of Mediterranean-type ecosystem agricultural residues: Characterisation of pretreatment products, high-solids enzymatic hydrolysis and bioethanol production. Biofuels 2017, 1–14. [Google Scholar] [CrossRef]
- Matsakas, L.; Rova, U.; Christakopoulos, P. Strategies for enhanced biogas generation through anaerobic digestion of forest material—An overview. BioResources 2016, 11, 5482–5499. [Google Scholar] [CrossRef]
- Choi, S.; Song, C.W.; Shin, J.H.; Lee, S.Y. Biorefineries for the production of top building block chemicals and their derivatives. Metab. Eng. 2015, 28, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Borodina, I.; Kildegaard, K.R.; Jensen, N.B.; Blicher, T.H.; Maury, J.; Sherstyk, S.; Schneider, K.; Lamosa, P.; Herrgård, M.J.; Rosenstand, I.; et al. Establishing a synthetic pathway for high-level production of 3-hydroxypropionic acid in Saccharomyces cerevisiae via β-alanine. Metab. Eng. 2015, 27, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.W.; Kim, J.W.; Cho, I.J.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of 3-hydroxypropionic acid and malonic acid through β-alanine route. ACS Synth. Biol. 2016, 5, 1256–1263. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, J.; Ogura, M.; Kanema, H.; Kawaharada, H.; Watanabe, K. Production of β-hydroxypropionic acid from propionic acid by a Candida rugosa mutant unable to assimilate propionic acid: Studies on β-hydroxycarboxylic acids (IV). J. Ferment. Technol. 1982, 60, 591–594. [Google Scholar]
- Luo, H.; Zhou, D.; Liu, X.; Nie, Z.; Quiroga-Sánchez, D.L.; Chang, Y. Production of 3-hydroxypropionic acid via the propionyl-CoA pathway using recombinant Escherichia coli strains. PLoS ONE 2016, 11, e0156286. [Google Scholar] [CrossRef] [PubMed]
- Burgard, A.P.; Dien van, S.J. Methods and Organisms for Growth-Coupled Production of 3-Hydroxypropionic Acid. U.S. Patent 2008/0199926 A1, 22 January 2008. [Google Scholar]
- Chen, Z.; Huang, J.; Wu, Y.; Wu, W.; Zhang, Y.; Liu, D. Metabolic engineering of Corynebacterium glutamicum for the production of 3-hydroxypropionic acid from glucose and xylose. Metab. Eng. 2017, 39, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Jiang, J.; Wu, H.; Li, Z.; Ye, Q. Enhanced production of 3-hydroxypropionic acid from glucose via malonyl-CoA pathway by engineered Escherichia coli. Bioresour. Technol. 2016, 200, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ding, Y.; Zhang, R.; Liu, H.; Xian, M.; Zhao, G. Functional balance between enzymes in malonyl-CoA pathway for 3-hydroxypropionate biosynthesis. Metab. Eng. 2016, 34, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Song, C.W.; Lee, J.; Ko, Y.S.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of 3-aminopropionic acid. Metab. Eng. 2015, 30, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Jung, I.Y.; Lee, J.W.; Min, W.K.; Park, Y.C.; Seo, J.H. Simultaneous conversion of glucose and xylose to 3-hydroxypropionic acid in engineered Escherichia coli by modulation of sugar transport and glycerol synthesis. Bioresour. Technol. 2015, 198, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Suyama, A.; Higuchi, Y.; Urushihara, M.; Maeda, Y.; Takegawa, K. Production of 3-hydroxypropionic acid via the malonyl-CoA pathway using recombinant fission yeast strains. J. Biosci. Bioeng. 2017, 124, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Kildegaard, K.R.; Jensen, N.B.; Schneider, K.; Czarnotta, E.; Özdemir, E.; Klein, T.; Maury, J.; Ebert, B.E.; Christensen, H.B.; Chen, Y.; et al. Engineering and systems-level analysis of Saccharomyces cerevisiae for production of 3-hydroxypropionic acid via malonyl-CoA reductase-dependent pathway. Microb. Cell Fact. 2016, 15, 53. [Google Scholar] [CrossRef] [PubMed]
- Kildegaard, K.R.; Wang, Z.; Chen, Y.; Nielsen, J.; Borodina, I. Production of 3-hydroxypropionic acid from glucose and xylose by metabolically engineered Saccharomyces cerevisiae. Metab. Eng. Commun. 2015, 2, 132–136. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Xiang, A.; Ying, A. Purification of 3-hydroxypropionitrile by wiped molecular distillation. Sci. China Ser. B 2004, 47, 521. [Google Scholar] [CrossRef]
- Yu, S.; Yao, P.; Li, J.; Ren, J.; Yuan, J.; Feng, J.; Wang, M.; Wu, Q.; Zhu, D. Enzymatic synthesis of 3-hydroxypropionic acid at high productivity by using free or immobilized cells of recombinant Escherichia coli. J. Mol. Catal. B Enzym. 2016, 129, 37–42. [Google Scholar] [CrossRef]
- Zhang, Q.; Gong, J.S.; Dong, T.T.; Liu, T.T.; Li, H.; Dou, W.F.; Lu, Z.M.; Shi, J.S.; Xu, Z.H. Nitrile-hydrolyzing enzyme from Meyerozyma guilliermondii and its potential in biosynthesis of 3-hydroxypropionic acid. Bioprocess Biosyst. Eng. 2017, 40, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, T.; Gao, X.; Shi, M.; Wu, L.; Chen, L.; Zhang, W. Biosynthesis of platform chemical 3-hydroxypropionic acid (3-HP) directly from CO2 in cyanobacterium Synechocystis sp. PCC 6803. Metab. Eng. 2016, 34, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Lan, E.I.; Chuang, D.S.; Shen, C.R.; Lee, A.M.; Ro, S.Y.; Liao, J.C. Metabolic engineering of cyanobacteria for photosynthetic 3-hydroxypropionic acid production from CO2 using Synechococcus elongatus PCC 7942. Metab. Eng. 2015, 31, 163–170. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes Transferred | Culture Conditions | Concentration (g/L) | Productivity (g/L·h) | Ref. |
|---|---|---|---|---|
| dhaS gene from B. subtilis | Fed-batch bioreactor 5 L, 3 L, pH 7, 37 °C, 1.5 vvm, 400 rpm | 18.0 | 0.77 | [27] |
| aldH gene from E. coli | Fed-batch bioreactor, 5 L, 2.8 L, pH 6.8–7.0, 37 °C, microaeration 1.5 vvm, 400 rpm | 48.9 | 1.75 | [28] |
| Overexpression of kgsadh, dhaB and gdrAB, deletion of ldhA, frdA, and adhE | Fed-batch bioreactor 1.5 L, 1 L, pH 7.5, 37 °C, 1 vvm, 400 rpm | 43.0 | 0.90 | [12] |
| PuuC overexpression, deletion of ldh1, ldh2, and pta | Fed-batch bioreactor 5 L, 3 L, pH 7, 37 °C, 1.5 vvm, 400 rpm | 83.8 | 1.16 | [29] |
| aldH gene from E. coli | Flasks 250 mL, 100 mL, 37 °C, microaeration, 150 rpm | 0.9 | 0.04 | [25] |
| - | Co-cultivation with Gluconobacter oxydans in fed-batch bioreactor, 7 L, 4.0 L, 1st step: pH 7, 37 °C, 0.2 vvm, 150 rpm 2nd step: pH 5.5, 28 °C, 0.5 vvm, 600 rpm | 60.5 | 1.12 | [30] |
| Genes Transferred | Culture Conditions | Concentration (g/L) | Productivity (g/L·h) | Ref. |
|---|---|---|---|---|
| dhaB and gdrAB from K. pneumoniae and kgsadh from A. brasilense | Flasks 250 mL, 100 mL, 37 °C, 200 rpm | 5.1 | n.a. | [31] |
| dhaB and gdrAB from K. pneumoniae, aldH from E. coli, overexpression of glpF, deletion of ackA-pta, yqhD, and glpR | Fed-batch bioreactor 5 L, 2 L, pH 7, 35 °C, 1 vvm, 500 rpm | 42.1 | 1.32 | [4] |
| dhaB and gdrAB from K. pneumoniae, gabD4 from C. necator with side-directed mutagenesis, deletion of yghD and ackA-pta | Fed-batch bioreactor 5 L, 2 L, pH 7, 35 °C, 1 vvm, 500 rpm | 71.9 | 1.8 | [34] |
| UTR-engineered dhaB, gdrAB from K. pneumoniae and kgsadh from A. brasilense, deletion of yghD and ackA-pta | Fed-batch bioreactor 5 L with the addition of glucose, 2 L, pH 7, 37 °C, 1 vvm, 500 rpm | 40.5 | 1.35 | [33] |
| dhaB and gdrAB from K. pneumoniae and kgsadh from A. brasilense | Fed-batch bioreactor 1.5 L with the addition of glucose, pH 7, 37 °C, 1 vvm, 650 rpm | 56.4 | 1.18 | [35] |
| dhaB and gdrAB from K. pneumoniae and kgsadh from A. brasilense | Fed-batch bioreactor 5 L with the addition of glucose, 3 L, pH 6.5, 37 °C, 450 rpm | 17.2 | n.a. | [36] |
| dhaB and gdrAB from K. pneumoniae and kgsadh from A. brasilense | Fed-batch bioreactor 1.5 L, 1 L, pH 7, 37 °C, 0.5 vvm, 650 rpm | 41.5 | 0.86 | [37] |
| dhaB and gdrAB from K. pneumoniae, AraE from A. brasilense, conditional repression of gapA, deletion of yqhD | Flasks, 300 mL, 37 °C, 150 rpm | 6.06 | 0.13 | [38] |
| dhaB and gdrAB from K. pneumoniae, AraE from A. brasilense, pdu from K. pneumoniae | Flasks 250 mL, 60 mL, pH 6–7, 100–250 rpm | 5.05 | 0.105 | [39] |
| Genes Transferred | Microorganism | Culture Conditions | Concentration (g/L) | Productivity (g/L·h) | Ref. |
|---|---|---|---|---|---|
| Mutation in the CRE upstream of the pdu operon | L. reuteri | Fed-batch bioreactor 3 L with resting cells, 1 L, pH 7, 37 °C, anaerobic, 500 rpm | 10.6 | 1.08 | [23] |
| Mutation in the CRE upstream of the pdu operon | L. reuteri | Immobilized cells with fed-batch feeding, 12 mL, 37 °C, anaerobic, 100 rpm | 3.3 | 0.09 | [14] |
| - | L. reuteri and G. oxydans | Two-step cultivation: (1) anaerobic fed-batch bioreactor 3 L with L. reuteri resting cells, 1 L, pH 5.5, 37 °C (2) aerobic batch system 3 L with G. oxydans resting cells, 1 L, pH 5.5, 28 °C, 0.33 vvm, 800 rpm | 23.6 | n.a. | [41] |
| dhaB, gdrAB, and puuC from K. pneumoniae, deletion of glpK | B. subtilis | Flasks with the addition of glucose, 37 °C, 200 rpm | 10 | n.a. | [42] |
| Genes Transferred | Culture Conditions | Concentration (g/L) | Productivity (g/L·h) | Ref. |
|---|---|---|---|---|
| acc from C. glutamicum and mcr from C. aurantiacus | Fed-batch bioreactor 5 L, 2.5 L, pH 7, 37 °C, 1 vvm | 10.1 | 0.28 | [54] |
| AccADBC, mcr with enhanced activity of the mcr-c fragment and tuning of the expression levels of mcr-n fragment | Fed-batch bioreactor 5 L, 2 L, pH 7, 37 °C | 40.6 | 0.56 | [55] |
| pa0132 from P. aeruginosa, ydfG from E. coli, upregulation of sdhC, overexpression of ppc | Fed-batch bioreactor 6.6 L, 2 L, pH 7, 37 °C, 1 vvm, 200–1000 rpm | 31.1 | 0.63 | [49] |
| dhaB and dhaR from Lactobacillus brevis, aldhH from Pseudomonas aeruginosa, gpd1 and gpp2 from S. cerevisiae, deletion of ptsG, glpK and yqhD, overexpression of xylR operon | Fed-batch bioreactor 2.5 L with co-conversion of glucose and xylose, 1 L, 37 °C > 25 °C, 1 vvm, 600–1300 rpm | 29.7 | 0.54 | [57] |
| Genes Transferred | Culture Conditions | Concentration (g/L) | Productivity (g/L·h) | Ref. |
|---|---|---|---|---|
| Multiple copies of mcr from C. aurantiacus and mutant acc1, acsL641P from S. enterica, overexpression of pdc1 and ald6, gapdh from C. acetobutylicum | Fed-batch bioreactor 1 L, 0.5 L, pH 5, 30 °C, 2 vvm, 800 rpm | 9.8 | 0.1 | [59] |
| yhxA from B. cereus, ydfG from E. coli, panD from T. castaneum, overexpression of aat2, pyc1, and pyc2 | Fed-batch bioreactor 1 L, 0.5 L, pH 5, 30 °C, 2 vvm, 800 rpm | 13.7 | 0.17 | [48] |
| pyc1 and pyc2, bapat from B. cereus, panD from T. castaneum, ydfG from E. coli | Fed-batch bioreactor 2.7 L on xylose, 1 L, pH 5, 30 °C, 1 vvm, 600–1200 rpm | 7.4 | 0.06 | [60] |
| Genes Transferred | Microorganism | Culture Conditions | Concentration (g/L) | Productivity (g/L·h) | Ref. |
|---|---|---|---|---|---|
| mcr from C. aurantiacus and overexpression of cut6p | S. pombe | Flasks 100 mL with high-density cultures, 10 mL, 30 °C, 250 rpm | 7.6 | 0.25 | [58] |
| pdu from K. pneumoniae, gpd1 and gpp2 from S. cerevisiae, gabD from C. necator, xylAB from E. coli, araE from C. glutamicum, deletion of ldhA, pta-ackA, poxB, and glpK, and replacement of ptsH with iolT1 and glk | C. glutamicum | Fed-batch bioreactor 5 L, 2 L, pH 7.2, 30 °C, 1 vvm | 62.6 | 0.87 | [53] |
| pdu from K. pneumoniae, gpd1 and gpp2 from S. cerevisiae, gabD from C. necator, xylAB from E. coli, araE from C. glutamicum, deletion of ldhA, pta-ackA, poxB, and glpK, and replacement of ptsH with iolT1 and glk | C. glutamicum | Fed-batch bioreactor 5 L with glucose and xylose, 2 L, pH 7.2, 30 °C, 1 vvm | 54.8 | n.a. | [53] |
| Genes Transferred | Microorganism | Culture Conditions | Concentration (g/L) | Productivity (g/L·h) | Ref. |
|---|---|---|---|---|---|
| pacd from C. rugora, pct from M. elsdenii and hpcd from C. aurantiacus, and deletion of ygfH and prpC | E. coli | Flasks 100 mL, 20 mL, pH 7, 30 °C, 200 rpm. Propionic acid as carbon source | 2.2 | n.a. | [51] |
| - | M. guilliermondii | Flasks, 20 mL, pH 7.2, 30 °C, 120 rpm. Hydrolysis of 3-hydroxypropionitrile with the addition of glucose | 19.5 | 0.20 | [63] |
| Nitrilase from environmental samples | E. coli | Batch reaction with immobilized cells with 3-hydroxypropionitrile, 30 °C | 184.7 | 36.9 | [62] |
| mcr from C. aurantiacus, overexpression of pntAB and accBCAD-birA, and deletion of phaB and pta | Synechocystis sp. | Flasks 100 mL, 20 mL, pH 7.5, 30 °C, 150 rpm. From CO2 | 0.84 | 0.006 | [64] |
| mcr from S. tokodaii and msr from M. sedula | S. elongates | Flasks, 50 mL, 30 °C. From CO2 | 0.67 | 0.002 | [65] |
| Ppc and aspC from E. coli, SkPYD4 from S. kluyveri, and msr from M. sedula | S. elongates | Flasks, 50 mL, 30 °C. From CO2 | 0.19 | 0.0005 | [65] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsakas, L.; Hrůzová, K.; Rova, U.; Christakopoulos, P. Biological Production of 3-Hydroxypropionic Acid: An Update on the Current Status. Fermentation 2018, 4, 13. https://doi.org/10.3390/fermentation4010013
Matsakas L, Hrůzová K, Rova U, Christakopoulos P. Biological Production of 3-Hydroxypropionic Acid: An Update on the Current Status. Fermentation. 2018; 4(1):13. https://doi.org/10.3390/fermentation4010013
Chicago/Turabian StyleMatsakas, Leonidas, Kateřina Hrůzová, Ulrika Rova, and Paul Christakopoulos. 2018. "Biological Production of 3-Hydroxypropionic Acid: An Update on the Current Status" Fermentation 4, no. 1: 13. https://doi.org/10.3390/fermentation4010013
APA StyleMatsakas, L., Hrůzová, K., Rova, U., & Christakopoulos, P. (2018). Biological Production of 3-Hydroxypropionic Acid: An Update on the Current Status. Fermentation, 4(1), 13. https://doi.org/10.3390/fermentation4010013